



1. Introduction
Across diverse, contemporary human societies, numerous sex differences appear in physiology, behavior, and experiences that often vary in magnitude depending on the culture (Archer, Reference Archer2019; Berenbaum & Beltz, Reference Berenbaum and Beltz2021; Geary, Reference Geary2021; Hyde, Reference Hyde2005; Wizeman & Pardue, Reference Wizeman and Pardue2001; Wood & Eagly, Reference Wood and Eagly2002; Zell, Krizan, & Teeter, Reference Zell, Krizan and Teeter2015). An evolutionary explanation for these differences relies on sexual selection theory.
Sexual selection occurs when one sex, usually the female, invests more in offspring, and the other sex, typically the male, invests more in mating competition (Darwin, Reference Darwin1871; Janicke, Häderer, Lajeunesse, & Anthes, Reference Janicke, Häderer, Lajeunesse and Anthes2016; Trivers, Reference Trivers and Campbell1972). When applied to humans, sexual selection theory has produced many explanations and testable predictions regarding the reproductive benefits of males' competitive strategies, including direct competition, physical aggression, risk-taking, showing off, impulsivity, sensation-seeking, and resource accumulation (Archer, Reference Archer2009, Reference Archer2019; Walter et al., Reference Walter, Conroy-Beam, Buss, Asao, Sorokowska, Sorokowski and Alm2020). Because such behaviors can improve males' ability to succeed in mating competition, they constitute part of an optimal strategy for fitness maximization even when they reduce survival. Mating competition thus accounts for many “male traits” (those that are expressed more in males than females).
In contrast to male traits, evolutionary theorists have tended to explain female traits in a more piecemeal manner. At least four theories related to sexual selection commonly have been applied to understanding female traits: mating competition, social selection, parental investment, and (in humans) “staying alive” theory (SAT).
The mating competition component of sexual selection theory explains female traits as adaptations for either defeating other females in conflicts over mates or being chosen as mates by males (Janicke et al., Reference Janicke, Häderer, Lajeunesse and Anthes2016). In humans, a high-quality mate can enhance a woman's fitness through providing resources, protection, support, and/or good genes (Campbell, Reference Campbell2004). Human females' competitive strategies to attract high-quality males rarely involve direct competition, but instead rely more on self-promotion (Blake, Bastian, Denson, Grosjean, & Brooks, Reference Blake, Bastian, Denson, Grosjean and Brooks2018; Puts, Reference Puts2010) and indirect tactics including denigration of competitors' reputations and social exclusion (Archer, Reference Archer2004; Campbell, Reference Campbell2004; Hess, Helfrecht, Hagen, Sell, & Hewlett, Reference Hess, Helfrecht, Hagen, Sell and Hewlett2010; Reynolds, Reference Reynolds2021).
Social selection theory applied to females emphasizes contests against other females for resources, territory, or allies (Clutton-Brock, Reference Clutton-Brock2007; Stockley & Bro-Jørgensen, Reference Stockley and Bro-Jørgensen2010). Again, direct contests are relatively uncommon in humans (Campbell, Reference Campbell2004). Instead, human females frequently compete for resources and allies utilizing indirect tactics as in mating competition (Björkqvist, Reference Björkqvist1994; Burbank, Reference Burbank1987; Campbell, Reference Campbell2004; Jankowiak, Sudakov, & Wilreker, Reference Jankowiak, Sudakov and Wilreker2005; Rucas, Reference Rucas and Fischer2017).
Parental investment theory focuses on how adults, typically females, provide optimal levels of care for their young (Trivers, Reference Trivers and Campbell1972). The theory examines adaptations that maximize production and survival of offspring independent of mating relationships, including optimizing interbirth intervals, manipulating sex ratios, and aborting embryos or infanticide (Hrdy, Reference Hrdy1981; Trivers, Reference Trivers1974; Trivers & Willard, Reference Trivers and Willard1973). Parental investment theory has been applied fruitfully to humans to explain diverse patterns of mothering (Hrdy, Reference Hrdy1999).
Building on parental investment theory, Campbell (Reference Campbell1999) proposed SAT. She posited that survival is more fitness-enhancing for females than males, and therefore that human females more than males evolved to avoid physical aggression and invest in alternative traits. Campbell (Reference Campbell1999) emphasized “that lower rates of aggression by women reflect not just the absence of male risk-taking but are part of a positive female adaptation driven by the critical importance of the mother's survival for her own reproductive success” (p. 204). Campbell focused on fear as the critical mechanism that protects females from injurious forms of aggression, and found that females indeed tend to be more fearful than males.
Here we examine whether Campbell's theory has more general applicability by considering the hypothesis that compared to males, human females respond with greater protectiveness to other threats beyond aggression. We define threat as anything potentially damaging to an individual's survival.
Campbell's (Reference Campbell1999) assumption that survival is more fitness-enhancing for females than males predicts that females tend to outlive males. In support, despite large cultural variations, in virtually every country women outlive men as shown in Figure 1 for 2020 (Roser, Ortiz-Ospina, & Ritchie, Reference Roser, Ortiz-Ospina and Ritchie2013). A similar tendency for females to live longer than males is found in mammals (Lemaître et al., Reference Lemaître, Ronget, Tidière, Allainé, Berger, Cohas and Liker2020).
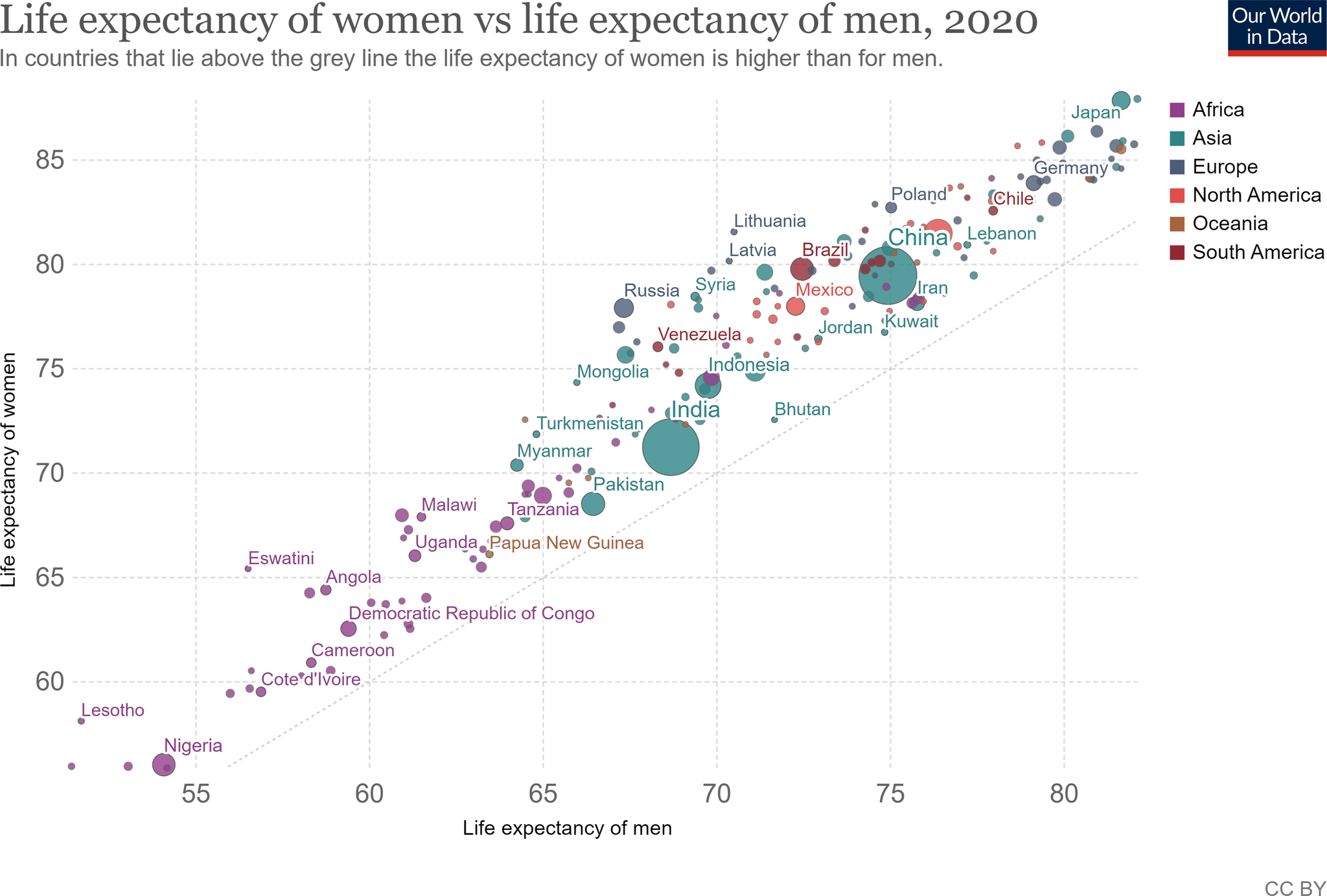
Figure 1. United Nations data for life expectancy by sex for 2020.
Source: UN Population Division (2019 Revision).
Note: Shown is the period life expectancy at birth measured in years.
SAT also predicts that the sex difference in longevity has evolved at least partly because of the benefits that offspring receive from female care, assuming that females contribute more than males to offspring survival. Studies of contemporary societies from hunter-gatherers to Western, Educated, Industrialized, Rich, and Democratic (WEIRD) nations (Henrich, Reference Henrich2020) confirm females' greater investment in and contribution to offspring survival (Atrash, Reference Atrash2011; Hawkes, O'Connell, & Blurton Jones, Reference Hawkes, O'Connell and Blurton Jones2018; Sear & Mace, Reference Sear and Mace2008). According to Wood and Eagly's (Reference Wood and Eagly2002) analysis of 186 geographically and culturally diverse non-industrial societies from the Human Relations Area Files, “all of the cultures in our review revealed an alliance between men and women in a division of labor, which appeared to be organized primarily to enable mothers to bear children and nurse and care for infants” (p. 718). Mothers were the principal caregivers of infants in 92% of these societies (and simultaneously of young children in 26–70% of these societies), whereas fathers and other men never were. Further, across six hunter-gatherer or horticulturalist societies, excluding nighttime co-sleeping, mothers took care of infants between 46 and 53% of the day and fathers between 0 and 6%, with older siblings, grandmothers, and others comprising the additional caregivers (Kramer, Reference Kramer2005).
Besides mothers, female kin, especially grandmothers and older daughters, most often helped mothers with childcare (Wood & Eagly, Reference Wood and Eagly2002). Grandmothers have been found to be particularly important for increasing grandchildren's survival, more so than grandfathers (Hawkes et al., Reference Hawkes, O'Connell and Blurton Jones2018; Lahdenpera, Lummaa, Helle, Tremblay, & Russell, Reference Lahdenpera, Lummaa, Helle, Tremblay and Russell2004; Sear & Mace, Reference Sear and Mace2008). A high adaptive value of grand-mothering is suggested by eight mammalian species in which females contribute to caring for their grand-offspring, because females' lifespans in those species averaged 43% longer than males, compared to only 8% longer in related species without grand-mothering (Péron et al., Reference Péron, Bonenfant, Lemaitre, Ronget, Tidiere and Gaillard2019).
Older daughters also help mothers (Bove, Valeggia, & Ellison, Reference Bove, Valeggia and Ellison2002; Kramer, Reference Kramer2005; Whiting et al., Reference Whiting, Edwards, Ember, Erchak, Harkness, Munroe and Seymour1988). Across 13 traditional societies, girls were more likely than boys to assist in direct care for younger siblings (Whiting et al., Reference Whiting, Edwards, Ember, Erchak, Harkness, Munroe and Seymour1988). In six hunter-gatherer or horticulturalist societies, girls cared for younger siblings between 13 and 33% of the day, with boys childminding 1–14% of the day (Kramer, Reference Kramer2005). In WEIRD societies where children rarely care for younger siblings, girls more than boys cared for dolls (d = 4.12) (Davis & Hines, Reference Davis and Hines2020). In non-human primates, preadult females also attend to and hold infants more than males do (Benenson, Reference Benenson, Welling and Shackelford2019).
These data support the idea that the greater longevity of females than males is an evolutionary response to women's larger role in childcare, such that at all ages selection tends to favor female survival. Thus, females and males confront different life history tradeoffs, with the sex that cares for offspring investing more in survival (Hamilton, Reference Hamilton1966; Kirkwood & Rose, Reference Kirkwood and Rose1991; Trivers, Reference Trivers and Campbell1972; Williams, Reference Williams1966). Preliminary evidence in mammals also links higher female longevity with greater female, relative to male, parental care (Allman, Rosin, Kumar, & Hasenstaub, Reference Allman, Rosin, Kumar and Hasenstaub1998).
To evaluate the applicability of Campbell's (Reference Campbell1999) theory to threats beyond aggression, we searched for articles that analyzed sex differences in response to major threats to survival: internal and external physical dangers, social conflicts, personality styles, emotional reactions to threat, and threat-related clinical conditions. We surveyed highly cited reviews, meta-analyses, and articles that examined human sex differences with large samples from diverse countries, along with authoritative references such as the World Health Organization (WHO) and the Diagnostic and Statistical Manual of Mental Disorders 5th edition (DSM-V). We report sample sizes, effect sizes based on Hyde's (Reference Hyde2005) cutoffs, and developmental effects when these were available. Because many fewer elderly men than women are alive thereby introducing confounds into sex-based comparisons, we focused on young and middle-aged adults. Further, because female investment in childcare increases sharply after puberty, we predicted increased magnitudes of sex differences following puberty and searched for evidence comparing children with adolescents. We included only high-quality datasets for each kind of threat and searched for inconsistencies with other published findings. Following Campbell (Reference Campbell1999), we included only self-protective reactions to threat, rather than reactions that increase the potential for harm, such as impulsivity, sensation-seeking, risk-taking, direct aggression, or homicide which are well-known to be less common in females than males (Archer, Reference Archer2019). Additionally, we reviewed Hyde's (Reference Hyde2005), Zell et al.'s (Reference Zell, Krizan and Teeter2015), and Archer's (Reference Archer2019) meta-analytic summaries of sex differences to identify any additional protective reactions to threats or contrary findings. All sex differences we report were statistically significant unless otherwise stated.
We recognize that human sex is a multidimensional construct influenced by the continuing interplay of biological and environmental components and that the studies we found are limited by including only binary comparisons (Berenbaum & Beltz, Reference Berenbaum and Beltz2021; Eliot, Reference Eliot2009; Fausto-Sterling, Reference Fausto-Sterling2019; Hyde, Bigler, Joel, Tate, & van Anders, Reference Hyde, Bigler, Joel, Tate and van Anders2019). We also acknowledge that many sex differences in specific traits are small, so that sex stereotypes based on single traits mischaracterize the large overlap between the sexes (Hyde, Reference Hyde2005; Zell et al., Reference Zell, Krizan and Teeter2015). Our goal is to suggest that many female-associated traits, which are often pathologized in comparison to male-related traits, can more appropriately be considered, like most evolved features, to enhance females' survival and reproductive success.
2. Protective responses to physical threat
According to our extension of SAT, females should protect themselves more than males in response to all physical threats. We therefore tested the hypothesis that females exhibit stronger defenses than males against pathogens, bodily damage, and environmental dangers.
Recent cross-cultural evidence shows that females were less likely than males to die from disease at every age. The major causes of sex differences in mortality worldwide are shown in Figure 2 with disease the most common contributor (WHO, 2018).
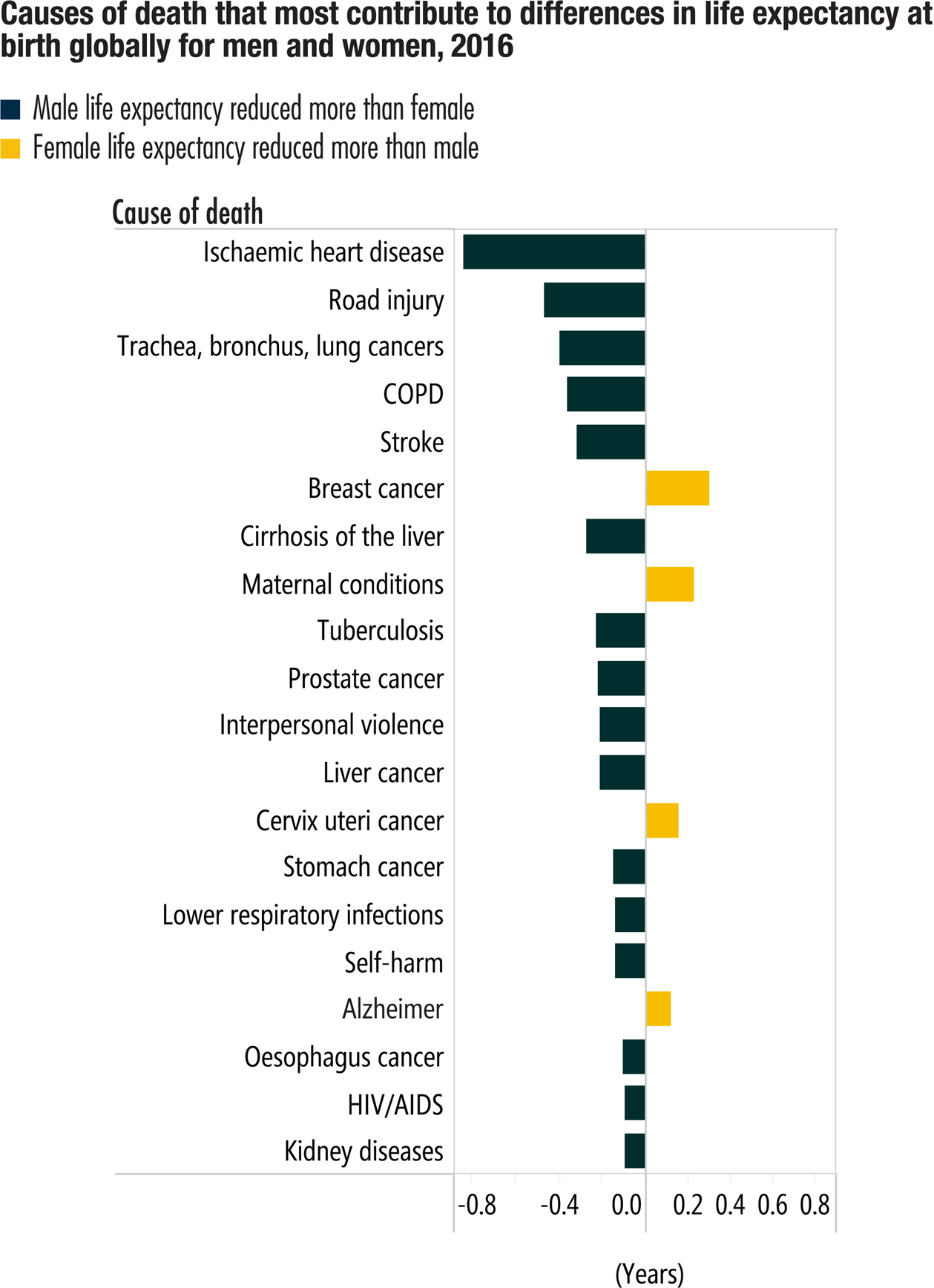
Figure 2. Sex differences in diseases that are major causes of death worldwide from the World Health Organization.
Source: WHO (2018) (2) and see (4) for composition of life expectancy. © Reprinted with permission from the World Health Organization.
Excluding reproductive-related conditions, the fatal illnesses that contributed most to reducing life expectancy afflicted females less frequently or later in life than males (Austad, Reference Austad2006; Verbrugge, Reference Verbrugge1985). Thus, females were less likely than males to develop many diseases, including malaria, leptospirosis, schistosomiasis, brucellosis, rabies, leishmaniasis, pulmonary tuberculosis, hepatitis A, meningococcal and pneumococcal infections (Giefing-Kröll, Berger, Lepperdinger, & Grubeck-Loebenstein, Reference Giefing-Kröll, Berger, Lepperdinger and Grubeck-Loebenstein2015; Mihailova & Krams, Reference Mihailova and Krams2018).
When females developed fatal diseases, they died later than males with small to moderate effect sizes. As examples, adjusting for age, worldwide in 2021 the odds of dying from lung (OR = 0.46), colorectal (OR = 0.68), and stomach (OR = 0.45) cancer were lower for women than men (WHO, 2021). In the United States in 2007, the age-adjusted odds of dying from cardiovascular disease (CVD) were lower for females than males (OR = 0.70) (Mosca, Barrett-Connor, & Kass Wenger, Reference Mosca, Barrett-Connor and Kass Wenger2011). In the United States in 2010, females' age-adjusted death rate was lower than males' for 12 of the most frequent 15 diseases. Two were equal, while only Alzheimer's disease killed more women than men (Austad & Bartke, Reference Austad and Bartke2016). Mortality from the coronavirus disease-2019 (COVID-19) pandemic epitomizes the overall pattern: In 37/38 countries fewer women than men died (Scully, Haverfield, Ursin, Tannenbaum, & Klein, Reference Scully, Haverfield, Ursin, Tannenbaum and Klein2020) with an effect size (OR = 0.63) (Williamson et al., Reference Williamson, Walker, Bhaskaran, Bacon, Bates, Morton and Inglesby2020). Sex differences in fatality rates from COVID-19 appeared even in mice (Scully et al., Reference Scully, Haverfield, Ursin, Tannenbaum and Klein2020). Overall, therefore, females are better protected than males from death by disease.
2.1. Immune defenses
Since immune activation is necessary for survival, SAT predicts that females should exhibit more effective immune responses than males. This prediction is upheld in vertebrates generally.
Across most vertebrate species, females are better able than males to defend themselves against a majority of environmental and vector-borne threats, including fungi, viruses, bacteria, parasites, and allergens, as well as internal threats such as tissue damage and tumors (Giefing-Kröll et al., Reference Giefing-Kröll, Berger, Lepperdinger and Grubeck-Loebenstein2015; Klein, Reference Klein2000; Klein & Flanagan, Reference Klein and Flanagan2016; Oertelt-Prigione, Reference Oertelt-Prigione2012; Ortona, Pierdominici, & Rider, Reference Ortona, Pierdominici and Rider2019). Females' immune systems also respond better than males' to trauma, sepsis, and shock (Angele, Frantz, & Chaudry, Reference Angele, Frantz and Chaudry2006; Angele, Pratschke, Hubbard, & Chaudry, Reference Angele, Pratschke, Hubbard and Chaudry2014). In response to injury, estrogen administration has been shown to promote healing by upregulating the immune system and reducing morbidity and mortality (Angele et al., Reference Angele, Pratschke, Hubbard and Chaudry2014; Bösch, Angele, & Chaudry, Reference Bösch, Angele and Chaudry2018). The American National Health and Nutrition Examination Survey (n = 38,000) from 1988 to 2006 of demographically representative adults showed that women exhibited a stronger immune response than men, as indicated by a higher inflammatory rate, with a large effect size (OR = 4.17) (Yang & Kozloski, Reference Yang and Kozloski2011).
The immune system functions in two waves (Roved, Westerdahl, & Hasselquist, Reference Roved, Westerdahl and Hasselquist2017). First, the general, continually active, costly, rapid, innate part of the immune system responds to a new infection or inflammation. Second, cytokines released by the innate immune system ramp up the adaptive immune system through increased cell-mediated T and humoral-situated B cells which are capable of remembering specific antigens, thereby creating longer-term protection. The type of adaptive immune response varies with T-helper (Th) cells, with one type (Th1, Th2, or Th17) increasing at the expense of another. Th1 immune cells typically produce pro-inflammatory responses that clear infections, whereas Th2 cells produce anti-inflammatory immune responses.
Part of the reason for female mammals' stronger immune response is that the X chromosome encodes more immune-related genes than the Y chromosome, and females have two X chromosomes while males have only one (Klein & Flanagan, Reference Klein and Flanagan2016). Further, in females, one X chromosome typically is randomly inactivated in each cell meaning that females are mosaics permitting increased transcription by both X chromosomes to contribute to immunity. Additionally, some X chromosomes escape inactivation also providing extra immunity (Takahashi & Iwasaki, Reference Takahashi and Iwasaki2021). In mice, experimentally adding an X chromosome to an XY individual to create an XXY individual increases longevity (Davis, Lobach, & Dubal, Reference Davis, Lobach and Dubal2019).
Another reason for females' stronger immune defenses is their higher estrogen and lower androgen levels. Estrogens intricately regulate immunity by both stimulating and suppressing differing types of immune cells, while androgens generally reduce immune functioning (Foo, Nakagawa, Rhodes, & Simmons, Reference Foo, Nakagawa, Rhodes and Simmons2017; Klein & Flanagan, Reference Klein and Flanagan2016; Roved et al., Reference Roved, Westerdahl and Hasselquist2017). Differing concentrations of estrogens can regulate the balance between T-helper cells-2 (Th2) and Th1 and Th17 cells. In contrast, androgens reduce Th2 and Th17 immune processes.
Estrogen levels are not solely responsible for enhanced immunity however, because even pre-pubertal girls activated stronger innate responses than boys, though some sex differences did not appear until puberty (Klein & Flanagan, Reference Klein and Flanagan2016). After puberty, more components of both the innate and adaptive immune systems were upregulated in women than in men.
Some of the clearest evidence for females' stronger inflammatory response comes from reactions to vaccinations. In response to most vaccinations including influenza, pneumonia, hepatitis A and B, tetanus, diphtheria, measles, meningitis, rabies, yellow fever, and smallpox, especially after puberty, human females generally produced more antibodies, and experienced more side effects including fever, soreness, and redness, than males (Flanagan, Fink, Plebanski, & Klein, Reference Flanagan, Fink, Plebanski and Klein2017; Klein, Jedlicka, & Pekosz, Reference Klein, Jedlicka and Pekosz2010). Immune responses were more suppressed in older individuals, but the suppression developed more slowly in women than men. Immune responses to vaccines were so much stronger in women than men that for a number of vaccines, half the dose of a vaccination in women caused the same level of antibodies as a full dose in men (Giefing-Kröll et al., Reference Giefing-Kröll, Berger, Lepperdinger and Grubeck-Loebenstein2015). While ceiling effects in vaccine efficacy obscure sex differences, when differences appeared, vaccines were more effective in females (Flanagan et al., Reference Flanagan, Fink, Plebanski and Klein2017). However, when vaccine doses were too strong, females were more likely to die from the vaccine (Flanagan et al., Reference Flanagan, Klein, Skakkebaek, Marriott, Marchant, Selin and Benn2011), as happened in some African infant inoculation campaigns (Aaby et al., Reference Aaby, Benn, Flanagan, Klein, Kollmann, Lynn and Shann2020). Further, a higher concentration of circulating estrogens was positively related to stronger responses to vaccines in humans and mice (Aaby et al., Reference Aaby, Benn, Flanagan, Klein, Kollmann, Lynn and Shann2020). Table 1 summarizes some of the human findings (Flanagan et al., Reference Flanagan, Fink, Plebanski and Klein2017).
Table 1 Sex differences in immune responses and adverse reactions to vaccinations over the lifespan.
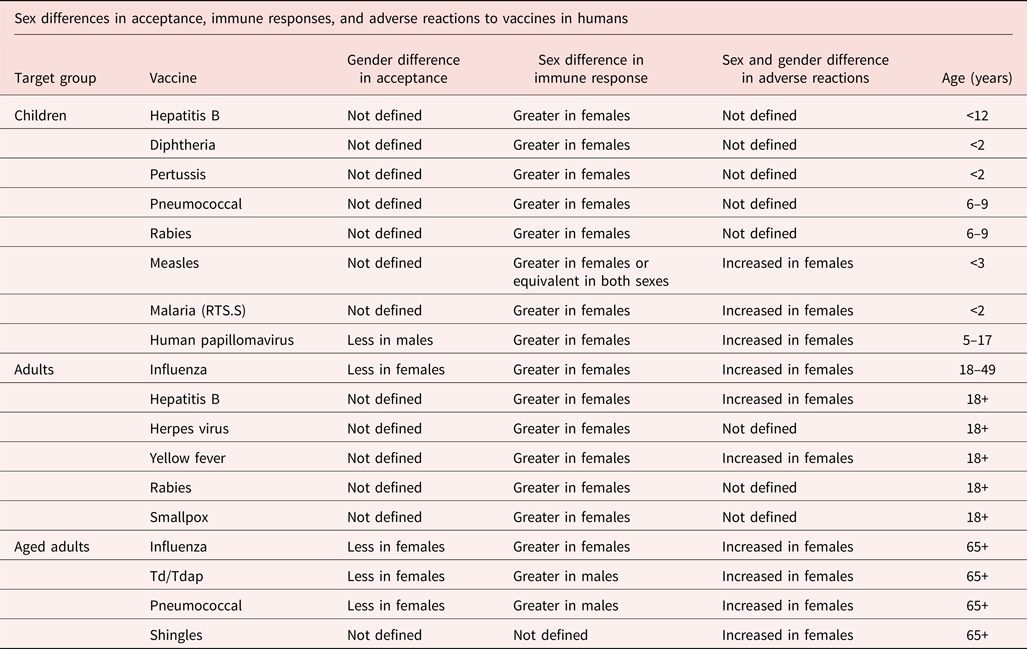
© Used with permission of Annual Reviews, Inc. from Sex and Gender Differences in the Outcomes of Vaccination over the Life Course, Katie L. Flanagan et al., 33, 2017, permission conveyed through Copyright Clearance Center, Inc.
Results with mice demonstrated causality. In mice inoculated against H1N1, females produced more robust IgG and IgA antibodies and B cells and higher quality antibodies than males. When these mice were infected with H1N1, females became less sick, had a lower viral load in their lungs, and recovered more rapidly than males (Fink, Engle, Ursin, Tang, & Klein, Reference Fink, Engle, Ursin, Tang and Klein2018). Observations of people infected with COVID-19 similarly showed that women with higher antibody levels tended to be less sick (Takahashi et al., Reference Takahashi, Ellingson, Wong, Israelow, Lucas, Klein and Tokuyama2020).
Pregnancy illustrates the strength of the immune system in non-pregnant women. During pregnancy, myriad changes occur in the innate and adaptive immune systems that affect maternal and fetal responses to pathogens (Abu-Raya, Michalski, Sadarangani, & Lavoie, Reference Abu-Raya, Michalski, Sadarangani and Lavoie2020). A pregnant woman's immune system must find an optimal way to balance ensuring survival of the genetically foreign fetus with protecting herself and the fetus from pathogens. One way this occurs is by downregulating Th1 cells producing pro-inflammatory cytokines. This switches the preponderance to Th2 cells producing anti-inflammatory cytokines. Sex hormones, including estrogens and progesterone, regulate the balance between Th1 and Th2 lymphocytes so that across pregnancy proinflammatory responses diminish and anti-inflammatory responses increase (Robinson & Klein, Reference Robinson and Klein2012).
The result is that pregnant women are more susceptible than non-pregnant women to, and/or suffer more severe, including fatal, reactions to infections from pathogens, including malaria, tuberculosis, influenza, hepatitis E, invasive pneumonia, herpes simplex virus, Ebola, and measles. In contrast, the severity of inflammatory diseases such as multiple sclerosis and arthritis is reduced (Abu-Raya et al., Reference Abu-Raya, Michalski, Sadarangani and Lavoie2020). Increased maternal morbidity and mortality during pregnancy also occurred with recent severe acute respiratory viruses (H1N1, SARS, and MERS) (Schwartz, Reference Schwartz2020), including COVID-19 (Zambrano et al., Reference Zambrano, Ellington, Strid, Galang, Oduyebo, Tong and Meaney-Delman2020).
Stronger activation of non-pregnant females' immune system however has a downside labeled “the health-survival paradox.” Despite their greater longevity, women felt sicker and were diagnosed with more illnesses than men across North America and Europe, and in Egypt, China, Indonesia, Mexico, and the Tsimane of Bolivia (Alberts et al., Reference Alberts, Archie, Gesquiere, Altmann, Vaupel, Christensen, Weinstein and Lane2014; Austad & Bartke, Reference Austad and Bartke2016; Christensen, Doblhammer, Rau, & Vaupel, Reference Christensen, Doblhammer, Rau and Vaupel2009; Khadr & Yount, Reference Khadr and Yount2012; Macintyre, Ford, & Hunt, Reference Macintyre, Ford and Hunt1999; Mirowsky & Ross, Reference Mirowsky and Ross1995; Roved et al., Reference Roved, Westerdahl and Hasselquist2017; Wheaton & Crimmins, Reference Wheaton and Crimmins2016).
Older women from culturally diverse societies, including the Tsimane of Bolivia, Taiwan, Korea, Mexico, China, Indonesia, reported being sicker and weaker and were evaluated objectively by medical personnel and researchers to be in poorer health than men.
“For instance, women in high-income countries are more likely than men to report difficulties in walking, climbing stairs, dressing, and other common activities. In low-income countries, women report greater difficulties than men in a wide range of common activities such as bending over, pumping water, or walking a specified distance. In addition, women make more doctor visits, spend more days hospitalized, and take more medications than do men. Even in Russia, which has one of the largest sex differences in life expectancy in the world, with a male disadvantage of more than 10 years, males report better health and physical functioning at ages of 55 years and higher.” (Austad & Batke, Reference Austad and Bartke2016, p. 42).
In twentieth-century American nationwide statistics, at all ages women experienced fewer fatal conditions than men, but suffered from more frequent daily symptoms, and more acute (except for injuries) and chronic and disabling conditions as diagnosed by doctors and in hospitals. The sex difference was largest during young adulthood, even after all reproduction-related conditions were excluded. Women reported more daily symptoms, more days with symptoms, more days in bed, more disability days, more health problems across more bodily systems, more prescription drug use, more alternative care visits, and more frequent discussions about their health problems with family and friends (Verbrugge, Reference Verbrugge1976, Reference Verbrugge1985, Reference Verbrugge1986, Reference Verbrugge1989).
The sex difference in the prevalence of immune activation increased at puberty but appeared by early childhood. In a large study of Norwegian adolescents (n = 10,000, ages 17–19 years), 47% of girls versus 21% of boys (OR = 3.34) reported at least one weekly health complaint (Myrtveit Sæther, Sivertsen, Haugland, Bøe, & Hysing, Reference Myrtveit Sæther, Sivertsen, Haugland, Bøe and Hysing2018). Two weekly health complaints were reported by 30% of girls and 10% of boys (OR = 3.86). Employing the Health Behaviour in School-Aged Children (HBSC) scale, a WHO study in 32 countries in Europe and North America plus Israel (n = 1,500 each nation at ages 11, 13, and 15 years, n > 150,000) found that girls reported poorer general health than boys at ages 11 (OR = 1.36), 13 (OR = 1.68), and 15 years (OR = 1.97) (Cavallo et al., Reference Cavallo, Zambon, Borraccino, Raven-Sieberer, Torsheim and Lemma2006). Likewise in the Avon Longitudinal Study of Parents and Children (ALSPAC) (n > 13,900), pregnant women in 1991–1992 in Southwestern England were recruited and later asked to report symptoms experienced by their 4–13-year-old children (Sweeting, Whitley, Teyhan, & Hunt, Reference Sweeting, Whitley, Teyhan and Hunt2017). From 4 years onwards, more girls than boys experienced colds, cold sores, tonsillitis, urinary infections, earaches, headaches, stomachaches, worm infections, head lice or scabies, constipation, and eczema. Beginning at age 7, more girls than boys experienced rashes, and by 10 years, more girls than boys developed fevers, ear and eye infections, and were struggling with poorer overall health. In contrast, in childhood, boys were more likely than girls to suffer chest infections, breathlessness, wheezing, asthma, hay fever, pain in legs and joints, and diarrhea. After puberty, most of boys' excess health problems disappeared with girls then experiencing these problems equally or more frequently.
In summary, evidence supported the hypothesis that with the exception of pregnancy, human females mounted stronger and longer-lasting immune responses than males.
2.2. Preventing injury through the sensation of pain
Pain signals potential danger or actual damage typically following inflammation or trauma (Eisenberger & Lieberman, Reference Eisenberger and Lieberman2004). It therefore constitutes a critical self-protective warning mechanism and ongoing monitoring system that impels corrective action to protect an organism (Grahek, Reference Grahek2001). A heightened sensation of pain should reduce the prevalence of injury and repair damage from an already sustained injury by behaviorally removing or reducing pain-inducing stimuli, thereby enhancing survival. We therefore searched the literature to test the hypothesis that females experience greater pain than males.
Fillingim, King, Ribeiro-Dasilva, Rahim-Williams, and Riley (Reference Fillingim, King, Ribeiro-Dasilva, Rahim-Williams and Riley2009) reviewed clinical and experimental pain studies across diverse countries including rural Brazil, India, Nigeria, Turkey, as well as in European and North American nations. They found that in their daily lives, more women than men reported higher pain prevalence at all sites in the body in the prior week or day, including oral, abdominal, and head pain. Women experienced pain more than men from a variety of sources and across multiple bodily sites, including from cancer, neuropathic, musculoskeletal, and back pain. Likewise in a detailed study of 11,000 patients with 47 diagnoses in an American hospital, patient-reported or practitioner-rated pain scores were higher for women than men for the same diagnosis as pictured in Figure 3 (Ruau, Liu, Clark, Angst, & Butte, Reference Ruau, Liu, Clark, Angst and Butte2012).

Figure 3. Average pain scores by sex for 47 conditions from one American hospital. © Reprinted from the Journal of Pain, 13/3, Ruau et al., Sex Differences in Reported Pain Across 11,000 Patients Captured in Electronic Medical Records, Pages 228–234, 2012, with permission from Elsevier.
The same sex difference appeared also for chronic, widespread pain. Mansfield, Sim, Jordan, and Jordan (Reference Mansfield, Sim, Jordan and Jordan2016) reviewed studies of chronic widespread pain in diverse countries including North and South America, the Middle East, Europe, Oceania, and Asia. Chronic widespread pain was reported more frequently by women than men on average by a 2:1 margin with large variation across ages and cultures and female:male ratios ranging from 1.06 to 4.80.
Even during simple intramuscular injections, women experienced more pain than men (Fillingim et al., Reference Fillingim, King, Ribeiro-Dasilva, Rahim-Williams and Riley2009). While this may be partially due to women's lower muscle density and tissue mass, even 5-year-old girls reported more pain than boys to venipuncture (Chambers, Giesbrecht, Craig, Bennett, & Huntsman, Reference Chambers, Giesbrecht, Craig, Bennett and Huntsman1999).
Sex differences in pain emerge in childhood. The WHO HBSC study in 32 countries in Europe, North America, and Israel (n > 150,000) found that more girls than boys experienced headaches at age 11, 13, and 15 years respectively (ORs = 1.44, 1.88, 2.70), stomachaches (ORs = 1.64, 2.00, 2.31), and backaches (ORs = 1.16, 1.24, 1.28) (Cavallo et al., Reference Cavallo, Zambon, Borraccino, Raven-Sieberer, Torsheim and Lemma2006). Likewise, the English ALSPAC study found that beginning with the youngest children in their study (age 4 years), girls reported more headaches, stomachaches, and earaches than boys did (Sweeting et al., Reference Sweeting, Whitley, Teyhan and Hunt2017). Chronic pain also was reported in childhood by females more than males (Fillingim et al., Reference Fillingim, King, Ribeiro-Dasilva, Rahim-Williams and Riley2009).
Similar sex differences in responses to pain were found in mice and rats (Mogil, Reference Mogil2020). Rodent studies of the midbrain periaqueductal gray and the spinal cord showed that pain and analgesics are modulated differently in females than males (Shansky & Murphy, Reference Shansky and Murphy2021).
In experimental studies that induce pain using identical stimuli, women consistently responded to bodily injury faster and endured it less long than men did across multiple bodily sites and modalities of inflicting pain (Fillingim et al., Reference Fillingim, King, Ribeiro-Dasilva, Rahim-Williams and Riley2009). In a meta-analysis of pain threshold (n > 1,600) and pain tolerance (n > 41,500) for pressure, heat, electrical shock, and ischemic induced pain, women endured pain at a lower stimulus intensity (d = 0.51) and for a shorter time period (d = 1.17) than men did, yielding moderate to large effect sizes (Riley, Robinson, Wise, Myers, & Fillingim, Reference Riley, Robinson, Wise, Myers and Fillingim1998). The sex difference in pain diminished in old age but never disappeared (Girotti et al., Reference Girotti, Trevisan, Fratta, Toffanello, Inelmen, Manzato and Sergi2019).
In experimental tests with children, a meta-analysis showed few sex differences (Boerner, Birnie, Caes, Schinkel, & Chambers, Reference Boerner, Birnie, Caes, Schinkel and Chambers2014). Overall, no sex differences were found in responses to pain intensity, and only 31% of studies found that girls exhibited lower pain thresholds than boys. Some specific types of pain, such as pain due to heat however, produced the same sex differences as in adolescents and adults.
In sum, adult females routinely experience greater pain than males. This suggests that a stronger experience of pain constitutes a female adaptation that functions to enhance survival through reducing injury. In support, the Global Burden of Disease findings showed that disability-adjusted lost years (DALYs) from all types of injuries were typically lower for females than males for ages 15–49 years, as displayed in Figure 4 (Haagsma et al., Reference Haagsma, Graetz, Bolliger, Naghavi, Higashi, Mullany and Alsharif2016).

Figure 4. Disability-adjusted life year (DALY) by injury, sex, and region for ages 15–49 years from the Global Burden of Disease project. DALY rates per 100000. CC BY-NC 4.0
The sex difference was smaller but in the same direction for children 0–14 years. From 1990 to 2017, the same sex difference for adults in DALYs appeared globally for child injuries from all causes except heat (James et al., Reference James, Castle, Dingels, Fox, Hamilton, Liu and LeGrand2020). Likewise, in the WHO HBSC study with 15-year-olds in 36 European and North American countries, girls reported fewer injuries overall than boys in the prior year and fewer physical fights (de Looze, Elgar, Currie, Kolip, & Stevens, Reference de Looze, Elgar, Currie, Kolip and Stevens2019).
2.3. Countering nighttime threats through frequent awakening
Nighttime is a dangerous period because threats are not easily detected. Increased nighttime vigilance improves detection of physical and social threats, including intruders, predators, and fire, that could elude identification during sleep. Accordingly, we tested the prediction from SAT that females are more attuned than males to nighttime threats.
We found that worldwide women reported being awake at night more often than men did. The American Society for Women's Health Research concluded that women's sleep differed objectively and subjectively from men's (Mallampalli & Carter, Reference Mallampalli and Carter2014): women experienced more slow wave sleep and less non-REM sleep than men. Women also experienced longer sleep latency and reported feeling sleepier than men. Further, across humans, mice, and rats, sex hormones affected females' more than males' sleep (Hajali, Andersen, Negah, & Sheibani, Reference Hajali, Andersen, Negah and Sheibani2019; Mallampalli & Carter, Reference Mallampalli and Carter2014).
A meta-analysis (n > 1.2 million, ages 15–103 years) from diverse countries of sex differences in sleep disturbances illustrated the findings: In 26 of 29 studies, at all ages women reported more frequent nighttime awakening than men (risk ratio = 1.41) (Zhang & Wing, Reference Zhang and Wing2006). The Chinese Henan Rural Cohort Study (n > 27,000, ages 18–79 years) employing the Pittsburgh Sleep Quality Index found that females experienced lower sleep quality than males with effect sizes ranging from negligible to small for subjective sleep quality (d = −0.20), sleep latency (d = 0.21), sleep duration (d = −0.03), sleep efficiency (d = −0.13), sleep disturbance (d = 0.15), use of sleep medication (d = 0.08), daytime dysfunction (d = 0.08), and generally (d = 0.22) (Wang et al., Reference Wang, Li, Liu, Liu, Mao, Tu and Jiang2019).
Globally, following puberty women had a 40% higher risk than men of developing insomnia during their lifetimes as depicted in Figure 5 (Mong & Cusmano, Reference Mong and Cusmano2016).

Figure 5. Prevalence of insomnia by sex and age. © Used with permission of The Royal Society from Sex differences in sleep: Impact of biological sex and sex steroids, Jessica A. Mong and Danielle M. Cusmano, 371, 2016, permission conveyed through Copyright Clearance Center, Inc.
Furthermore, a meta-analysis (n > 180,000) showed that more females than males experienced nightmares with negligible to small effect sizes in adolescents (Hedge's g = 0.22), young adults (g = 0.26), and in middle age (g = 0.15). The sex difference was not significant for children (g = −0.03), or >60 years (g = 0.10) (Schredl & Reinhard, Reference Schredl and Reinhard2011).
Although sex differences in nighttime awakenings were negligible to small for children, they increased with age and pubertal status. In the 32 country WHO HBSC survey, girls reported increasing sleep difficulties relative to boys at 11, 13, and 15 years of age (ORs = 1.13, 1.36, 1.56, respectively) (Cavallo et al., Reference Cavallo, Zambon, Borraccino, Raven-Sieberer, Torsheim and Lemma2006). A study in Hong Kong (n > 7,500 ages 6–17 years) linked pubertal development to specific sleep disturbances (Zhang et al., Reference Zhang, Chan, Lam, Li, Liu, Chan and Li2016). Children and adolescents, and their caregivers, reported that girls developed more overall insomnia, difficulty initiating sleep, and difficulty maintaining sleep than boys as they entered pubertal stage 4 on the Tanner scale, although they already had experienced more difficulties with early morning awakening as displayed in Figure 6.

Figure 6. Rates of insomnia in Hong Kong by sex and Tanner stage for (A) overall insomnia symptoms, (B) difficulty initiating sleep, (C) difficulty maintaining sleep, and (D) early morning awakening. © Jihui Zhang et al., Emergence of Sex Differences in Insomnia Symptoms in Adolescents: A Large-Scale School-Based Study, Sleep, 2016, 39, 8, 1563–1570, by permission of Oxford University Press.
Overall, evidence supported the hypothesis that females experienced more nighttime awakenings than males. This is consistent with the hypothesis that nighttime awakenings constitute a female adaptation that enhanced self-protection during an interval of heightened vulnerability.
We also noted that women were less likely than men to suffer from severe, potentially fatal sleep disorders including narcolepsy, obstructive sleep apnea, and hypoventilation (Hajali et al., Reference Hajali, Andersen, Negah and Sheibani2019; Lévy et al., Reference Lévy, Kohler, McNicholas, Barbé, McEvoy, Somers and Pépin2015). Furthermore, laboratory studies of sleep indicated that length, depth, and efficiency of sleep, including enhanced slow wave sleep and slow wave activity, were higher in women than men (Hajali et al., Reference Hajali, Andersen, Negah and Sheibani2019; Mong & Cusmano, Reference Mong and Cusmano2016; Suh, Cho, & Zhang, Reference Suh, Cho and Zhang2018). While these observations are limited to Western research, they suggest that females might compensate for their more frequent awakenings by maintaining a superior quality of sleep.
2.4. Miscellaneous physical threats
Innumerable external physical stimuli threaten survival. We tested the hypothesis, derived from SAT, that females would be more effective than males at avoiding or reducing such threats. We were able to locate evidence for three types of threat: hypothetical physical threats, long-term environmental degradation, and major disease outbreaks.
In the standardized International Affective Picture System (IAPS), participants evaluated the valence of photographs of non-human animals, scenes, and inanimate objects. In the United States, Germany, Switzerland, and China, girls and women reported stronger aversion than their male counterparts to all represented threats, including accidents, illness, pollution, and dangerous animals (Bradley, Codispoti, Sabatinelli, & Lang, Reference Bradley, Codispoti, Sabatinelli and Lang2001; Lang & Bradley, Reference Lang, Bradley, Coan and Allen2007) with moderate to large effect sizes in adults (d = 0.36) (Gomez, von Gunten, & Danuser, Reference Gomez, von Gunten and Danuser2013), (d = 0.55) (Gong, Wong, & Wang, Reference Gong, Wong and Wang2018) (d = 0.43 and 0.80) (McManis, Bradley, Berg, Cuthbert, & Lang, Reference McManis, Bradley, Berg, Cuthbert and Lang2001), and in children (d = 0.64 and 0.67) (McManis et al., Reference McManis, Bradley, Berg, Cuthbert and Lang2001).
Similar sex differences have been found in response to environmental dangers. In the International Social Survey Programme (ISSP) administered in 1993 and 2000 in 26 countries from Asia, Eastern and Western Europe, North and South America, and Oceania (n > 23,000), women expressed more concern about the state of the environment than men did (Franzen & Meyer, Reference Franzen and Meyer2010). Between 2009 and 2011, an expanded ISSP in 32 nations in Africa, Asia, Europe, North and South America, and Oceania (n > 45,000) showed that women exhibited more positive views than men toward protecting the environment, greater awareness of environmental problems, and more willingness to pay to correct environmental problems (Chan, Pong, & Tam, Reference Chan, Pong and Tam2019).
With regard to major outbreaks of disease, a review with Asian, European, and North American participants (n > 25,000, 12 countries) showed that during disease outbreaks women reported being more likely than men to adopt avoidant and protective measures, including handwashing, wearing a mask, and quarantining (Bish & Michie, Reference Bish and Michie2010). Across eight OECD countries (n > 20,000), women were more likely than men to have perceived COVID-19 as a serious health risk and to have complied with restraining public health rules (Galasso et al., Reference Galasso, Pons, Profeta, Becher, Brouard and Foucault2020).
Nonetheless, women can be more reluctant than men to receive vaccines, most likely because of the more adverse side effects of vaccines in women than men (Flanagan et al., Reference Flanagan, Fink, Plebanski and Klein2017). A review (37 studies, 15 diverse countries) of reactions to the 2009 strain of H1N1 influenza showed that both women in the general population and female health care professionals indicated they would be less likely than their male counterparts to be vaccinated (Bish, Yardley, Nicoll, & Michie, Reference Bish, Yardley, Nicoll and Michie2011). An explanation proposed by Bish et al. (Reference Bish, Yardley, Nicoll and Michie2011) was that women tended to believe that the immediate negative effects of the vaccine outweighed its health benefits, consistent with greater side effects in women. A similar concern about negative effects might explain a meta-analysis (n = 88 studies) with African, Asian, Australian, European, North and South American samples, which found that women were 50% more likely than men to have adopted non-pharmacological prevention and avoidant measures in reaction to respiratory pandemics (OR = 1.49), whereas men were slightly more likely to take pharmaceutical protections (OR = 0.89) (Moran & Del Valle, Reference Moran and Del Valle2016).
Overall, for the environmental threats we examined, women reacted more protectively than men with small to large effect sizes, consistent with a female adaptation for increasing longevity.
3. Protective responses to social threat
Social threat includes at least two distinct types: conflicts and isolation. Social conflicts can be physically dangerous, potentially leading to fights, reputation denigration, ostracism, expulsion, and even capital punishment (Boehm, Reference Boehm1999; Wrangham, Reference Wrangham2019). Social isolation predicts psychological and physical morbidity and mortality in young, middle-aged, and elderly adults in a dose-dependent manner (Hawkley & Cacioppo, Reference Hawkley and Cacioppo2010). A meta-analysis of longitudinal studies (n > 3.4 million) showed that objective measures of both social isolation and feeling lonely predicted increased mortality (OR = 1.26–1.83) (Holt-Lunstad, Smith, Baker, Harris, & Stephenson, Reference Holt-Lunstad, Smith, Baker, Harris and Stephenson2015). The longer the loneliness, the greater the risk. Even experimentally inducing feelings of loneliness increased stress and feelings of danger (Holt-Lunstad et al., Reference Holt-Lunstad, Smith, Baker, Harris and Stephenson2015).
Conversely, it is well known that social ties increase survival. A meta-analysis of longitudinal findings from Asia, Australia, Europe, and North America in community and patient samples (n > 300,000) showed that across sex, age, initial health status, and cause of death, social integration increased survival by 50%, and by 91% using multiple measures of integration (Holt-Lunstad, Smith, & Layton, Reference Holt-Lunstad, Smith and Layton2010). Similarly, in a large study in Asia, Europe, and North America (n > 7.5 million), not being married increased all-cause mortality in both sexes, more for males (risk ratio = 1.46) than females (risk ratio = 1.22) (Wang et al., Reference Wang, Jiao, Nie, O'Neil, Huang, Zhang and Yu2020).
SAT would predict that females tend to avoid conflicts and promote social ties more than males do. We found relevant evidence for four strategies that reduce negativity in interactions and facilitate positive connections, namely smiling, politeness, emotion identification, and avoidance of confrontations.
3.1. Smiling
Evidence indicates that women smile more than men with small to moderate effect sizes. A meta-analysis of 162 studies (n > 100,000, 13 countries, 6 continents) reported greater smiling by women from adolescence through old age in every country (d = 0.41) (LaFrance, Hecht, & Paluck, Reference LaFrance, Hecht and Paluck2003). Sex differences were greater in same-sex pairs (d = 0.48) than mixed-sex pairs (d = 0.35). Sex differences also were larger in contexts with greater social tension (d = 0.47) than little tension (d = 0.20), and when participants were unfamiliar (d = 0.45) rather than familiar (d = 0.24). This suggested that an important function of smiling was to reduce threats from other individuals. Prior meta-analyses also concluded that women's smiles more than men's function to reduce social tension (Hall & Halberstadt, Reference Hall, Halberstadt, Hyde and Inn1986).
Computerized coding likewise showed that women smiled more than men. Analyses of photographs of Asian, Black, and White New York City residents posted to Instagram and Twitter (n > 15,000) demonstrated that the more females in a picture, the more smiling occurred as coded by automated software (Singh, Atrey, & Hegde, Reference Singh, Atrey and Hegde2017). Software similarly found that in Argentina, Brazil, China, Colombia, France, Germany, India, Japan, Mexico, Peru, Russia, the United Kingdom, and the United States (n > 740,000), when watching television advertisements women smiled more than men (McDuff, Girard, & El Kaliouby, Reference McDuff, Girard and El Kaliouby2017). Sex differences in smiling however were not found in children (Else-Quest, Hyde, Goldsmith, & Van Hulle, Reference Else-Quest, Hyde, Goldsmith and Van Hulle2006).
3.2. Politeness
SAT would predict that females would be more likely than males to speak politely since this should reduce interpersonal conflicts. We tested the hypothesis that females more than males employed polite language.
Evidence indicated that females exhibited greater politeness than males with small to moderate effect sizes. In a meta-analysis in primarily WEIRD countries (n > 3,500, 29 studies), women used more tentative language (hedges, expressions of uncertainty, intensifiers, and tag questions) than men (d = 0.23), which the authors interpreted as seeking the listener's consent (Leaper & Robnett, Reference Leaper and Robnett2011). Ethnographic reports from Africa, Europe, North and South America, and Oceania also reported that girls and women used more tentative speech than boys and men by employing more qualifiers, euphemisms, and apologies, and speaking with less assertiveness, more agreement, higher numbers of softeners and compliments, and greater supportiveness and modesty (Brown & Levinson, Reference Brown and Levinson1987; Coates, Reference Coates2015; Guadagno & Cialdini, Reference Guadagno and Cialdini2007; Haas, Reference Haas1979; Holmes, Reference Holmes1989; Lakoff, Reference Lakoff1975; Locke, Reference Locke2011).
The same type of sex difference in the use of language has regularly been found in early childhood. In a meta-analysis of speech forms in WEIRD cultures (46 studies, n > 2,600), girls made more general affiliative comments (d = 0.26) than boys which was even stronger in unstructured contexts (d = 0.58) (Leaper & Smith, Reference Leaper and Smith2004). Ethnographic reports in 13 non-WEIRD societies likewise indicated that in middle childhood girls use more prosocial and less egoistic dominance in their speech (Whiting et al., Reference Whiting, Edwards, Ember, Erchak, Harkness, Munroe and Seymour1988).
3.3. Identification of others' emotions
Accurately identifying another's emotions facilitates interactions by enhancing understanding of the actor's emotional state. SAT would predict that females would be more likely than males to accurately identify another's emotions.
Studies utilizing primarily pictures or short videos of non-verbal facial, bodily, and vocal behavior consistently demonstrated that from early in life females detected social signals more accurately than males did with small to moderate effect sizes. A meta-analysis (75 studies, 1923–1978) with WEIRD and non-WEIRD societies showed that girls and women were better able than boys and men to accurately detect others' non-verbally expressed emotions (d = 0.40) (Hall, Reference Hall1978). A follow-up review demonstrated that across cultures, ages, and target sex, girls and women were better able than boys and men to accurately detect the emotional valence of non-verbal facial and bodily behaviors, as well as verbal intonation and prosody (d = 0.41–0.54) (Hall, Carter, & Horgan, Reference Hall, Carter, Horgan and Fischer2000). A more recent meta-analysis showed that in 67% of 32 studies, women were more accurate than men in identifying the valence of emotions, whereas no sex differences occurred in the remainder (Forni-Santos & Osório, Reference Forni-Santos and Osório2015). Similarly, an online study (n > 7,300, ages 18–75+ years) employing the Penn Emotion Recognition Test with happy, sad, angry, and fearful faces showed that women more accurately identified emotions across the lifespan (d = 0.41) (Sasson et al., Reference Sasson, Pinkham, Richard, Hughett, Gur and Gur2010).
Importantly, women were particularly proficient at detecting negative emotions, though the specific negative emotions were not always consistent across studies and the effect sizes were small. In a large meta-analysis (n > 79,000), which encompassed several of the prior meta-analyses with children through adults in nations from Africa, East and Southeast Asia, Europe, North and South America, and including New Guinea, Taiwan, and Tibet, an overall female advantage was found in accurate recognition of emotional reactions to threat (d = 0.24) and of positive emotions (d = 0.19) (Thompson & Voyer, Reference Thompson and Voyer2014). For specific emotions, the effect sizes were highest for anger (d = 0.25), sadness (d = 0.24), and fear (d = 0.22), then slightly lower for happiness (d = 0.18), disgust (d = 0.17), and surprise (d = 0.15), indicating that females were more accurate than males at all ages in decoding emotional signals of threat from non-verbal expressions.
In an Emotion Recognition Test (n > 42,000, ages 11–78 years) conducted in seven South American nations, plus Mexico, Austria, Germany, Spain, Switzerland, and the United States, females were more accurate than males at identifying emotions (d = 0.22) (Merten, Reference Merten2005). For specific emotions however, the sex difference was significant only for anger, fear, and sadness (not for disgust, joy, or surprise). In three online studies (n > 1,900) in which subjects rated static facial and dynamic bodily emotions in less than one second, women recognized disgust and sadness significantly more accurately than men did, but not anger, fear, or happiness (Connolly, Lefevre, Young, & Lewis, Reference Connolly, Lefevre, Young and Lewis2019).
McClure (Reference McClure2000)'s meta-analysis of 58 studies of facial emotion processing in infancy through adolescence likewise demonstrated that girls were more accurate than boys at identifying emotions. The effect size was smaller in older children and adolescents (d = 0.18) than in infants (d = 0.70) however, which may be attributable to the earlier maturity of infant females than males (McClure, Reference McClure2000).
Thus overall we found that females are reported to accurately identify others' emotions more than males.
3.4. Avoidance of confrontation
SAT would predict that females are more likely than males to avoid confrontations and facilitate positive interactions. We found two forms of confrontation with systematic evidence: direct competition and face-to-face aggression.
Evidence indicates that direct competition and aggression are less common in females than males with moderate to large effect sizes. Direct competition has been systematically examined both in economic games and sports. A standardized economic game was created in which individuals chose to either compete against others or work alone with equal expected payoffs (Niederle & Vesterlund, Reference Niederle and Vesterlund2007). In WEIRD and non-WEIRD societies, across tasks, group size, and sex of opponents, two reviews concluded that girls and women were less likely than boys and men to compete against others (Klege, Visser, Barron, & Clarke, Reference Klege, Visser, Barron and Clarke2021; Sutter, Zoller, & Glatzle-Rutzler, Reference Sutter, Zoller and Glatzle-Rutzler2019). Based on Klege et al. (Reference Klege, Visser, Barron and Clarke2021), we calculated effect sizes for 33 studies with adults (d = 0.43) and 12 studies with children (d = 0.55).
Similar evidence comes from sports. Throughout known history, cross-culturally women have participated less than men in competitive sports (Craig, Reference Craig2002; Leibs, Reference Leibs2004). In a geographically and culturally diverse sample of 50 societies, of 248 sports documented, males participated in 95% of the sports and females in 20%, whereas no sex differences existed in participation in amusements (Deaner & Smith, Reference Deaner and Smith2013). In the detailed American Time Use Survey (2003–2010) of activities in the past 24 hours (n > 112,000), 0.64% of females and 2.69% of males (d = −0.92) participated in team sports, and 1.29% of females and 4.45% of males participated in 1:1 sports (d = −0.95), whereas no sex differences occurred in exercise (Deaner et al., Reference Deaner, Geary, Puts, Ham, Kruger, Fles and Grandis2012). Whether in the context of sports or simply informal interactions, ethnographic evidence from Africa, Canada, Europeans, Native Americans, and in Oceania also described the less competitive and conflictual relationships of girls and women compared with boys and men (Locke, Reference Locke2011).
Similar to findings on sex differences in direct competition, females were found to engage in less verbal (d = −0.30) and physical (d = −0.59) confrontation than males (Archer, Reference Archer2019), with moderate effect sizes. Although most studies came from Asia, Europe, and North America, ethnographic reports provided the same conclusion. Despite large cultural variation, females engaged in less frequent and severe direct aggression than males did (Fry, Reference Fry1998; Locke, Reference Locke2011). The sex difference in direct aggression occurred across diverse cultures by age 2 years and continued through adolescence (Maccoby & Jacklin, Reference Maccoby and Jacklin1974). In non-human primates as well, females typically engaged in less intense physical aggression than males (Sabbi et al., Reference Sabbi, Thompson, Machanda, Otali, Wrangham and Muller2021; Smuts, Reference Smuts, Smuts, Cheney, Seyfarth, Wrangham and Struhsaker1987).
Thus evidence from smiling, politeness, emotion identification, and avoidance of direct confrontation indicated that females invested more than males in reducing social conflicts and protecting social bonds. This result is consistent with the finding from 70 countries with student and adult samples (n > 75,000) that women evaluated the “preservation and enhancement of the welfare of people with whom one is in frequent personal contact” as more important than men did (d = 0.19–0.36) (Schwartz & Rubel, Reference Schwartz and Rubel2005).
4. Personality style as a response to threat
Neuroticism consists of a wide range of negative behaviors, moods, emotions, and thoughts. One component of neuroticism, focused on worry and feelings of vulnerability, has been associated with reduced morbidity and mortality (Weiss & Deary, Reference Weiss and Deary2020). Because worry and feeling vulnerable are consistent with promoting self-protective behaviors toward threats, we interpreted SAT as predicting that females should report higher neuroticism than males.
The conclusion from four large cross-cultural studies is that beginning in adolescence, women exhibit higher neuroticism than men. First, in a study of university students (n > 17,500), neuroticism was defined as worrying, tension, nervousness, depression/blue, moodiness, uncalm in tense situations, easily upset, and not relaxed/handling stress poorly. In 49 of 55 countries, women reported more neuroticism than men did, whereas in no country did men report higher neuroticism than women (d = 0.40). Figure 7 displays effect sizes for sex differences in personality styles from world regions with neuroticism showing small to moderate sex differences (Schmitt, Realo, Voracek, & Allik, Reference Schmitt, Realo, Voracek and Allik2008).

Figure 7. Magnitudes of sex differences in neuroticism by world region based on the International Sexuality Description Project. © Reprinted with permission of the American Psychological Association.
Second, in an online study with individuals primarily from Australia, Canada, Ireland, New Zealand, and the United States (n > 1.2 million, ages 10–65 years, 70% Caucasian, 8% Asian, 6% African, 6% Hispanic), neuroticism was defined by worrying, not remaining calm in tense situations, being depressed/blue, and being moody (Soto, John, Gosling, & Potter, Reference Soto, John, Gosling and Potter2011). Overall neuroticism, and sub-clinical levels of anxiety and depression, differed by sex with females' rates increasing in early adolescence and remaining high during their peak reproductive years as displayed in Figure 8.
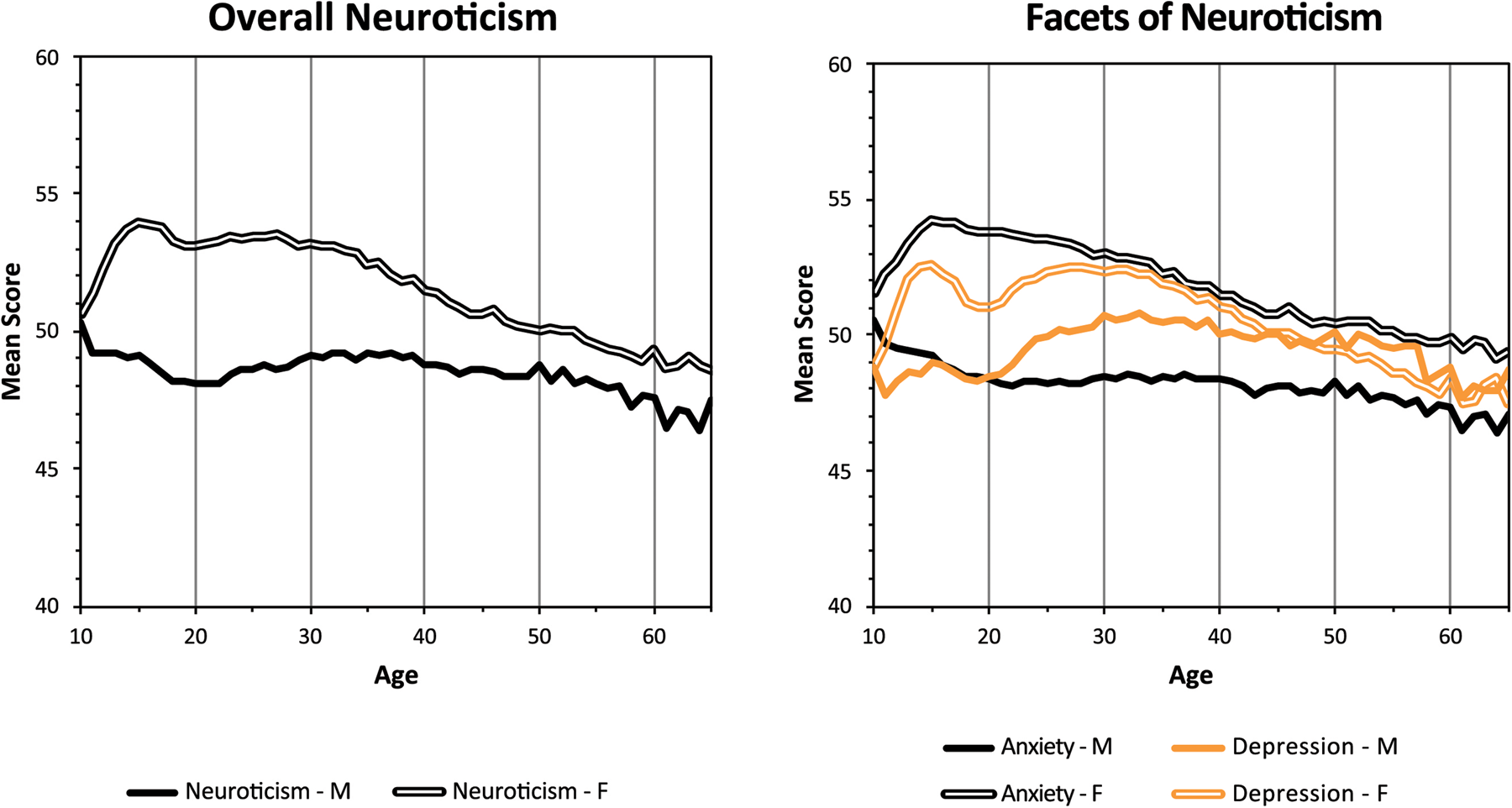
Figure 8. Sex differences in neuroticism by age in a large English-speaking sample. © Reprinted with permission of the American Psychological Association.
Third, a study of US adults (n > 320,000) defined neuroticism as being anxious, angry, depressed, self-conscious, immoderate, and vulnerable. As before, women, particularly during their reproductive years, described themselves as more neurotic than men (d = 0.40) with the anxiety (d = 0.56) and vulnerability (d = 0.54) scores yielding the greatest sex differences (Kajonius & Johnson, Reference Kajonius and Johnson2018).
Finally, in a classic study of personality in 24 highly diverse countries with university students and adults, women rated themselves as more neurotic than men: more vulnerable (d = 0.28–0.44), anxious (d = 0.32–0.43), self-conscious (d = 0.22–0.30), depressed (d = 0.17–0.24), impulsive (d = 0.11–0.23), and angry/hostile (d = 0.09–0.19), although sex differences in angry/hostile were not always significant (Costa, Terracciano, & McCrae, Reference Costa, Terracciano and McCrae2001).
In childhood, as depicted above, sex differences in neuroticism began after age 10 years. Facets of neuroticism, particularly anxiety, however, demonstrated sex differences even earlier (Soto et al., Reference Soto, John, Gosling and Potter2011).
In summary, beginning in adolescence females consistently report higher neuroticism than males, with small to moderate effect sizes. The greater neuroticism of post-pubertal females conforms to the prediction from SAT, suggesting that neuroticism functions to protect females more effectively than males from a diversity of threats.
5. Emotions: immediate reactions to threat
Emotional reactions to threat constitute critical signals that promote staying alive (Darwin, Reference Darwin1872; Ekman & Oster, Reference Ekman and Oster1979; Lazarus, Reference Lazarus1991). Six basic emotions have been identified because they appear across diverse cultures, occur in other primates, and emerge in childhood (Ekman & Cordaro, Reference Ekman and Cordaro2011). Four of these (fear, disgust, sadness, and anger) are considered responses to threat, whereas two (joy and surprise) are not. Each basic emotion arising in response to a threat is believed to produce a unique aversive signal that promotes a defensive action aimed at removing or avoiding the threat.
Assuming that emotional responses to threat promote survival, SAT would predict that females should experience these emotions more than males. We therefore tested the hypothesis that females experience more fear, disgust, sadness, and anger than males.
5.1. Fear
Fear enhances survival by motivating an individual to withdraw from potentially life-threatening danger (Ekman & Cordaro, Reference Ekman and Cordaro2011; Lazarus, Reference Lazarus1991). Campbell (Reference Campbell1999) predicated SAT on female's greater fear of physical aggression, while our extension of SAT predicts that females would exhibit greater fear than males to any threat.
Cross-cultural studies confirm that women tend to display greater fear than men. Archer's (Reference Archer2019) review reported greater fearfulness for adult females than males with small to large effect sizes (d = 0.07–1.16). Additionally, when university students (n = 5,000) from 11 countries were asked to rate the fearfulness of 52 stimuli organized into social, agoraphobic, bodily, and animal categories, women in every nation reported greater fearfulness to every category (d = 0.02–0.90) (Arrindell et al., Reference Arrindell, Eisemann, Oei, Caballo, Sanavio, Sica and Iwawaki2004). Further, in 1975–1980 when university students from 28 diverse countries were asked whether either sex was better described by various adjectives, women were characterized as more fearful than men in every country (Williams & Best, Reference Williams and Best1990).
In the International College Student Data (ICSD) survey (n > 6,000, 39 countries), students reported how frequently and intensely they experienced different emotions. Women described more frequent and intense fear than men (d = 0.31) (Lucas & Gohm, Reference Lucas and Gohm2000). In two studies, one with an Australian sample of students, kin, and friends (n > 2,100), and a second with an international sample of students from 41 countries (n > 6,500), women reported more frequent fear in the past month in Australia (d = 0.14) and in the international sample (d = 0.17) and more intense feelings of fear in the international sample (d = 0.26) (Brebner, Reference Brebner2003).
A small effect has also been found in children. In a cross-cultural meta-analysis (n > 4,500, 3 months–13 years), girls displayed greater fear than boys (d = 0.12) (Else-Quest et al., Reference Else-Quest, Hyde, Goldsmith and Van Hulle2006).
5.2. Disgust
Like fear, disgust is believed to enhance survival by producing withdrawal from potentially life-threatening danger (Ekman & Cordaro, Reference Ekman and Cordaro2011; Lazarus, Reference Lazarus1991). Classically, disgust arises in response to illness-causing objects, such as feces, vomit, mucous, signs of severe illness, rotting flesh, and other potentially contaminating stimuli. Additional, less well-known forms of disgust include sexual, animal, and moral disgust. Experiencing greater disgust or revulsion is directly linked with higher germ avoidance (Hartmann & Siegrist, Reference Hartmann and Siegrist2018), immediately increased immune activation (Schaller, Miller, Gervais, Yager, & Chen, Reference Schaller, Miller, Gervais, Yager and Chen2010; Stevenson et al., Reference Stevenson, Hodgson, Oaten, Moussavi, Langberg, Case and Barouei2012), and disease avoidance (Curtis, Aunger, & Rabie, Reference Curtis, Aunger and Rabie2004; Oaten, Stevenson, & Case, Reference Oaten, Stevenson and Case2009). Since disgust acts as a mechanism for self-protection, SAT would predict that females would experience greater disgust than males.
We found that across varied disgust-producing stimuli, females experienced more disgust than males, with small to large effect sizes. In an online cross-cultural study (n > 390,000, 67 countries), women were more distressed than men about lack of physical/spiritual purity and contamination (Atari, Lai, & Dehghani, Reference Atari, Lai and Dehghani2020). Likewise, on the BBC science website (n > 39,000), children and adults evaluated which of two paired images evoked more disgust. Across ages and continents, females were more likely than males to rate the image designed to appear as if it could transmit disease as more disgusting than its disease-free twin (Curtis et al., Reference Curtis, Aunger and Rabie2004). In another online study (n > 6,100, ages 20–69 years, 10 nations, 5 continents), researchers examined disgust ratings for animal flesh, poor hygiene, human contamination, mold, decaying fruits, fish, decaying vegetables, and living contaminants (Egolf, Hartmann, & Siegrist, Reference Egolf, Hartmann and Siegrist2019). Across countries women's disgust ratings were greater than men's (d = 0.12–0.37). Similarly, participants from Canada, England, and the United States (n > 2,500, mean age 28 years) evaluated degree of disgust toward five targets. Women provided higher ratings than men for animal disgust (d = 0.82), sex disgust (d = 0.70), hygiene disgust (d = 0.48), food disgust (d = 0.44), and lesion disgust (d = 0.31) (Curtis & de Barra, Reference Curtis and de Barra2018). Sexual stimuli produced some of the largest sex differences. Women experienced greater disgust than men to sexual stimuli (d = 0.60–1.54) (Al-Shawaf, Lewis, & Buss, Reference Al-Shawaf, Lewis and Buss2018).
Sex differences in disgust appeared in childhood. As described in the large paired comparison study which began at age 7 years, even the youngest girls exhibited greater disgust than boys (Curtis et al., Reference Curtis, Aunger and Rabie2004).
5.3. Sadness
Sadness is believed to result from the loss of someone or something important to one's life (Ekman & Cordaro, Reference Ekman and Cordaro2011; Lazarus, Reference Lazarus1991). Attachment theory posits that sadness or grief represents separation distress from a support figure (parent, spouse, ally) or from someone with close genetic ties (child, twin) (Archer, Reference Archer, Stroebe, Hansson and Stroebe2008; Bowlby, Reference Bowlby1980). Loss of a relationship partner can threaten the bereaved's survival (Holt-Lunstad et al., Reference Holt-Lunstad, Smith, Baker, Harris and Stephenson2015) or reproductive success (Archer, Reference Archer, Stroebe, Hansson and Stroebe2008) and often leads to withdrawal. Unlike with fear and disgust however, with sadness the loss often has already occurred, and withdrawal is gradual rather than sudden. Withdrawal is believed to protect the bereft individual who likely has lost a current or future ally, and to help elicit support from others (Lomas, Reference Lomas2018). On the assumption that sadness functions to reduce the impact of a threatening factor (the loss of a relationship partner), SAT would predict that females would experience greater sadness than males.
Based on self-reports, females were found to exhibit more sadness than males, with small effect sizes. In the IAPS, standardized photographs depicting illness generated more sadness in women than men (Bradley et al., Reference Bradley, Codispoti, Sabatinelli and Lang2001). In the 39 nation ICSD survey, women reported more frequent and intense sadness than men did (d = 0.26) (Lucas & Gohm, Reference Lucas and Gohm2000). Likewise, women reported more frequent sadness in Australia (d = 0.17) and the 41 country international sample (d = 0.16), and more intense feelings of sadness in Australia (d = 0.19) and the international sample (d = 0.28) (Brebner, Reference Brebner2003).
Higher self-reports of sadness could in theory be accounted for by women's greater willingness to report emotion. To examine this possibility, in 1990 researchers interviewed a demographically representative sample of Americans (n > 2,000, ages 18–90 years) (Mirowsky & Ross, Reference Mirowsky and Ross1995). When asked the number of days in the prior week they felt sad, women reported more days of sadness than men. Then, participants were asked how much they hide their emotions. At every level of emotional reserve, women reported experiencing greater sadness than men, strongly suggesting that the sex difference in reported sadness was not due to a reporting bias.
Limited evidence suggests girls are sadder by adolescence. In a demographically representative American sample (n > 21,000, ages 14–18 years), girls felt sadder than boys (Moeller, Brackett, Ivcevic, & White, Reference Moeller, Brackett, Ivcevic and White2020). Likewise, when Swedish adolescents (n > 1,000, ages 16–18 years) in 49 classrooms were asked how frequently they experienced sadness, 35% of the girls versus 8% of the boys reported feeling sad either “fairly often” or “very often” (OR = 6.19) (Wiklund, Malmgren-Olsson, Öhman, Bergström, & Fjellman-Wiklund, Reference Wiklund, Malmgren-Olsson, Öhman, Bergström and Fjellman-Wiklund2012). In childhood, sex differences in sadness may not exist or are negligible. In a meta-analysis (n > 2,300), in 11 of 16 studies in early and middle childhood exhibited girls greater sadness than boys, but the sex difference was not significant and negligible in size (d = 0.10) (Else-Quest et al., Reference Else-Quest, Hyde, Goldsmith and Van Hulle2006).
Evidence also indicated that females cry (weep) more than males, with moderate to large effect sizes. Crying is often associated with sadness, although it can also serve as a release from emotion and as a means of soliciting support (Rottenberg, Bylsma, & Vingerhoets, Reference Rottenberg, Bylsma and Vingerhoets2008; Vingerhoets & Scheirs, Reference Vingerhoets and Scheirs2000).
In the International Study of Adult Crying (ISAC) (n > 5,500) conducted in 37 nations across six continents, university students were asked how easily and how frequently they had cried in the past month (Van Hemert, van de Vijver, & Vingerhoets, Reference Van Hemert, van de Vijver and Vingerhoets2011). In every country, women reported crying more easily (d = 1.11) and more recently (d = 0.94) than men. Another meta-analysis (15 studies) found that women reported crying more frequently, intensely, and for longer intervals than men across cultures, including Israel, Japan, and the United States (Vingerhoets & Scheirs, Reference Vingerhoets and Scheirs2000). Similarly, across Australia, Croatia, the Netherlands, Thailand, and the United Kingdom (n > 800, ages 18–40 years), women reported crying more frequently (d = 0.74) and intensely (d = 0.45) than men (Sharman et al., Reference Sharman, Dingle, Baker, Fischer, Gracanin, Kardum and Vingerhoets2019). Ethnographic reports from diverse cultures similarly report that the loss of a relationship partner causes sadness and weeping, and more so in women than men (Palgi & Abramovitch, Reference Palgi and Abramovitch1984). Sex differences in crying are not reported before adolescence (Vingerhoets & Scheirs, Reference Vingerhoets and Scheirs2000).
5.4. Anger
Anger occurs after a personal goal has been obstructed, often by a specific perpetrator who purposefully wishes to harm a victim and/or her/his associates (Ekman & Cordaro, Reference Ekman and Cordaro2011; Lazarus, Reference Lazarus1991). Unlike fear, disgust, and sadness however, anger activates approach, not withdrawal. A further contrast is that although anger-motivated responses can reduce a threat through avoidance, they can also increase the risk of being injured through approaching a perpetrator (Sell, Tooby, & Cosmides, Reference Sell, Tooby and Cosmides2009). This means that whether anger functions as self-protection will vary with the context. Anger should be more self-protective if it enhances avoidance of a perpetrator, but less self-protective if it motivates close engagement or cycles of retaliation. Experiencing anger without approaching the perpetrator however should highlight the danger of future interactions with specific perpetrators, thereby enhancing survival.
SAT therefore would predict that females would feel angrier than males toward perpetrators, though only when the approach was suppressed. Even though the prototype of an angry person is a man (Kring, Reference Kring and Fischer2000) most likely because larger individuals face lower likelihood and costs of retaliation (Sell et al., Reference Sell, Tooby and Cosmides2009), we tested the hypothesis that females feel angrier than males.
Evidence suggested at most a negligible effect size indicating females experienced more anger than males (Archer, Reference Archer2004; Kring, Reference Kring and Fischer2000). However, no studies distinguished simple anger from anger tied to aggression. A meta-analysis of 11 countries found a non-significant effect for females to feel angrier than males (d = 0.04) (Archer, Reference Archer2004). In the ICSD study, women reported more frequent anger than men (d = 0.08) (Lucas & Gohm, Reference Lucas and Gohm2000). Likewise, in Brebner's (Reference Brebner2003) Australian and international samples, women reported experiencing anger more frequently than men did both in Australia (d = 0.06) and the international sample (d = 0.05). Women also reported greater intensity of anger in the international sample (d = 0.14). In Mirowsky and Ross (Reference Mirowsky and Ross1995) American study, anger was defined as “feeling annoyed with things or people,” “feeling angry,” and “yelling at someone.” Women reported feeling angrier than men, and “yelling at someone” more than men did. Sex differences in experiencing anger have not been found in children (Potegal & Archer, Reference Potegal and Archer2004).
In conclusion, evidence supported the hypothesis that females experienced more fear, disgust, and sadness than males did. Results did not support an overall sex difference in anger but anger could not be separated from approaching a perpetrator. When a sex difference did arise however, women experienced more anger than men.
6. Clinical conditions: extreme self-protective reactions to threat
Some clinical conditions are believed to be non-adaptive exaggerations of normal adaptive reactions to stress (Del Giudice, Reference Del Giudice2018; Nesse, Reference Nesse2005). If self-protectiveness is an adaptive female strategy, then clinical conditions constituting extreme self-protectiveness to threats should be more prevalent in females than males. We therefore tested whether extreme forms of physical, social, emotional, and generalized reactions to threat occur more frequently in women than men.
6.1. Autoimmune diseases
As we reviewed above, non-pregnant women tend to exhibit stronger immune responses than men including enhanced Th2 reactions. Extrapolating from this result, we reasoned that more females than males would develop extreme immunoenhancing conditions. We therefore searched the literature to test the hypothesis that more females than males develop extreme immune reactions.
The prediction was supported by evidence that globally, women are more likely than men to develop systemic autoimmune diseases (AD) which are linked to proinflammatory responses with moderate effect sizes (Ji, Sundquist, & Sundquist, Reference Ji, Sundquist and Sundquist2016; Moroni, Bianchi, & Lleo, Reference Moroni, Bianchi and Lleo2012; Purnamawati et al., Reference Purnamawati, Ong, Deshpande, Tan, Masurkar, Low and Drum2018) and diminish in severity during pregnancy (Abu-Raya et al., Reference Abu-Raya, Michalski, Sadarangani and Lavoie2020). Of 81 ADs identified worldwide, 47 afflict females more than males, 22 are more common in males, and 9 exhibit no sex difference (Hayter & Cook, Reference Hayter and Cook2012). Despite large cultural differences, overall 6.4% of females and 2.7% of males develop an AD (OR = 2.46) with onset most common between 20 and 29 years. The more prevalent an AD, the higher the proportion of females who develop it, with women comprising 78% of those with ADs (Hayter & Cook, Reference Hayter and Cook2012). Whereas many female-preponderant ADs are believed due to enhanced Th2 responses, in male-preponderant ADs, Th1 reactions often are stronger (Fairweather, Frisancho-Kiss, & Rose, Reference Fairweather, Frisancho-Kiss and Rose2008). Precipitating factors for all ADs include infections or other types of diseases or trauma (Nielsen, Kragstrup, Deleuran, & Benros, Reference Nielsen, Kragstrup, Deleuran and Benros2016). Results therefore are consistent with a female adaptation that promotes immunoenhancing reactions to physical threat.
6.2. Pain disorders
Evidence reviewed above demonstrated that females experience more ordinary pain than males. According to the idea that clinical conditions are non-adaptive extensions of well-adapted responses, extreme pain reactions should be found more often in females than in males.
We found two examples for which cross-cultural evidence is available, migraine headaches and fibromyalgia (FM), both of which were experienced by more females than males. The Global Burden of Diseases report finds that despite large national variations, worldwide 18.9% of women and 9.8% of men experienced migraine headaches (OR = 2.15) with women ages 15–49 years most affected (Stovner et al., Reference Stovner, Nichols, Steiner, Abd-Allah, Abdelalim, Al-Raddadi and Doan2018). The most common precipitating factor was external stress, including odors, foods, weather, pain to the neck or head, or emotional trauma (Peroutka, Reference Peroutka2014; Woldeamanuel & Cowan, Reference Woldeamanuel and Cowan2017).
FM consists of chronic, severe, widespread pain in at least 11 of 18 points in the joints, muscles, and spine. Worldwide, 4.2% of females and 1.4% of males (OR = 3.09) developed FM (Queiroz, Reference Queiroz2013). Onset of FM typically followed a stressor, such as an accident, surgery, infection, or emotional trauma, and was accompanied by sleep disturbance, fatigue, memory, and concentration problems, along with depression and anxiety (Galvez-Sánchez, Duschek, & del Paso, Reference Galvez-Sánchez, Duschek and del Paso2019; Häuser, Thieme, & Turk, Reference Häuser, Thieme and Turk2010). Prevalence ratios of migraine and FM showing moderate effect sizes were consistent with the hypothesis that a female adaptation exists for heightened experience of pain.
6.3. Sleep disorders
Prior evidence showed that females experienced more nighttime awakenings than males, which is plausibly an adaptive response. More extreme, non-adaptive nighttime awakenings were therefore also expected to occur more often in females than in males. We therefore tested whether more females than males developed conditions with severe nighttime awakening.
In line with this prediction, we found that more women than men experienced severe sleep disturbances with small to moderate effect sizes. Although no sharp distinction separates nighttime awakenings from a severe sleep disorder (Suh et al., Reference Suh, Cho and Zhang2018), analyses of major DSM-V clinical conditions showed a female:male prevalence ratio of 1.4 for insomnia disorder and 1.5–2 for restless leg syndrome, which disrupts sleep, as well as a higher female prevalence for nightmare disorder (Hartung & Lefler, Reference Hartung and Lefler2019). A meta-analysis (n > 18,500) further confirmed a female preponderance for restless leg syndrome (OR = 1.63–2.22) (Ohayon & Roth, Reference Ohayon and Roth2002). Additionally, a detailed study in Hong Kong (n > 8,500) found women were more likely than men to experience nightmares ⩾3 times/week (OR = 2.10) (Li, Zhang, Li, & Wing, Reference Li, Zhang, Li and Wing2010).
Furthermore, major sleep disruptions including day-night reversals were associated with clinical conditions with a female preponderance (Lashley, Reference Lashley2003). These included responses to infections, many autoimmune conditions (systemic lupus erythematosus, multiple sclerosis, rheumatoid arthritis), pain-related disorders (migraine, FM), and as described below, anxiety disorders, major depression, post-traumatic stress disorder, and chronic fatigue syndrome. Prevalence ratios of severe sleep disruption were consistent with the thesis that because a female adaptation exists for being especially attuned to nighttime threats, females were also vulnerable to having extreme responses that are likely maladaptive.
6.4. Separation disorder
We have presented evidence that females were more likely than males to avoid social confrontations and to become more distressed about relationship partners' well-being. A more extreme version of this response would consist of becoming extremely distressed about the loss of relationship partners. We therefore tested whether more females than males develop conditions characterized by excessive concern for the maintenance of relationships. We found one condition with cross-cultural evidence that clearly fit this criterion: separation anxiety disorder. Separation anxiety disorder includes severe distress at being away from an attachment figure along with persistent worry about negative events befalling attachment figures.
Examining 18 DSM-IV mental disorders (n > 72,000, across Africa, the Americas, Asia, Europe, the Middle East, and the Pacific), the WHO World Mental Health Survey (WMHS) study found that lifetime prevalence of separation anxiety disorder was more common in females than males with a small effect size (OR = 1.6), whereas the reverse was true for antisocial disorders (Seedat et al., Reference Seedat, Scott, Angermeyer, Berglund, Bromet, Brugha and Jin2009). Similarly, in analyses of the DSM-V categories, separation anxiety disorder was more common in females than males, while males suffered more from antisocial disorders (Hartung & Lefler, Reference Hartung and Lefler2019; Holthausen & Habel, Reference Holthausen and Habel2018; Shear, Jin, Ruscio, Walters, & Kessler, Reference Shear, Jin, Ruscio, Walters and Kessler2006). A demographically representative survey in the United States (n > 14,000) likewise showed that by the end of early childhood and throughout life females were more likely than males to have experienced separation anxiety disorder (OR = 2.2) (Shear et al., Reference Shear, Jin, Ruscio, Walters and Kessler2006). Reported rates of separation disorder are thus consistent with a heightened female adaptation for maintaining relationships.
6.5. Anxiety disorders
Evidence reviewed above demonstrated that females of all ages were more likely than males to experience fear in response to threats. More persistent and intense fear reactions would accordingly be expected to develop in females than males. We therefore tested whether more females than males developed conditions characterized by severe fear.
Anxiety disorders are specific or generalized worries and fears about potential threats often accompanied by unpleasant physical sensations, including increased heart rate and shaking. The worldwide WHO WMHS study showed a higher prevalence for females than males of anxiety disorders with small effect sizes, including panic disorder (OR: 1.9), generalized anxiety disorder (OR: 1.7), agoraphobia (OR: 2.0), social phobia (OR: 1.3), specific phobia (OR: 2.0), separation anxiety disorder (OR: 1.6) as mentioned, and any anxiety disorder (OR: 1.7) (Seedat et al., Reference Seedat, Scott, Angermeyer, Berglund, Bromet, Brugha and Jin2009). Other estimates of the female:male ratio of global prevalence of anxiety disorders were 1.9 (Remes, Brayne, Van Der Linde, & Lafortune, Reference Remes, Brayne, Van Der Linde and Lafortune2016) with the DSM-V analysis finding a ratio of 2:1 (Hartung & Lefler, Reference Hartung and Lefler2019). Thus, anxiety disorders conform to the hypothesis that they represent exaggerated versions of responses that would be adaptive if produced at an appropriate level.
6.6. Major depression and suicide attempts
Earlier we showed evidence that females were more likely than males to experience sadness. Extrapolating from this produces the prediction that more females than males should develop an extreme form of sadness: depression. We tested this hypothesis.
Major depression consists of an array of uncomfortable emotional and physical symptoms including sadness, feelings of worthlessness, reductions in activity and energy levels, changes in appetite and sleep, and difficulty thinking. In 80% of cases, depression is precipitated by stress (LeMoult, Reference LeMoult2020). A meta-analysis of sex differences in major depression across six continents (n > 1.7 million, ages 12–70+ years) showed a higher female prevalence with a moderate effect size (OR = 1.95) (Salk, Hyde, & Abramson, Reference Salk, Hyde and Abramson2017). The DSM-V analysis reported depression to be 1.5–3 times more common in women than men (Hartung & Lefler, Reference Hartung and Lefler2019).
Major depression likely plays a role in suicide attempts. Suicide attempts however may be not only an exaggeration of depression, but also an adaptive request for help across diverse societies (Syme, Garfield, & Hagen, Reference Syme, Garfield and Hagen2016). Although cross-culturally women were less likely than men to commit suicide (Alothman & Fogarty, Reference Alothman and Fogarty2020; WHO, 2014b), women were more likely than men to attempt suicide, beginning in early adolescence (Freeman et al., Reference Freeman, Mergl, Kohls, Székely, Gusmao, Arensman and Rummel-Kluge2017; Lenz et al., Reference Lenz, Röther, Bouna-Pyrrou, Mühle, Tektas and Kornhuber2019; WHO, 2014b). Maximal rates of non-fatal self-injuries for females occurred between 10 and 24 years (OR = 1.72). Females were still more likely than males to attempt suicide between 25 and 44 years (OR = 1.29), but the sex difference became negligible at older ages (Nock et al., Reference Nock, Borges, Bromet, Cha, Kessler and Lee2008). A meta-analysis of suicide attempts by 12–26-year-olds further found that in 23 of 24 studies girls were more likely than boys to attempt suicide (OR = 1.96) (Miranda-Mendizabal et al., Reference Miranda-Mendizabal, Castellví, Parés-Badell, Alayo, Almenara, Alonso and Gili2019). Effect sizes range from small to moderate. The higher rates of depression and lower success of female suicide attempts can be interpreted as females being more invested in self-protection, in line with SAT.
6.7. Psychiatric disorders incorporating disgust
We found previously that females were more likely than males to experience disgust in response to potentially contaminating stimuli. We therefore tested the idea that more extreme forms of disgust would also be more common in females than males.
We found that disgust is integral to three psychiatric illnesses: animal phobias, contamination-related obsessive–compulsive disorders with continual washing (OCD), and blood-injection-injury (BII) phobia which can include fainting. All three of these illnesses were more common in women than men (Arrindell, Mulkens, Kok, & Vollenbroek, Reference Arrindell, Mulkens, Kok and Vollenbroek1999; Davey, Reference Davey2011; Mathis et al., Reference Mathis, Alvarenga, Funaro, Torresan, Moraes, Torres and Hounie2011; Olatunji, Cisler, McKay, & Phillips, Reference Olatunji, Cisler, McKay and Phillips2010; Wani & Ara, Reference Wani and Ara2014). Disgust has also been associated with other serious psychiatric problems that are more prevalent in women than men, including eating disorders, agoraphobia, and female sexual dysfunction (Hartung & Lefler, Reference Hartung and Lefler2019). In all cases, specific threats were the focus. In contrast, most hypersexual-related psychiatric illnesses were more frequent in men than women. Thus, evidence supported the idea that extreme forms of disgust were experienced by more females than males.
6.8. Threat-induced conditions
Given evidence that females reacted more self-protectively than males to threats, it would be expected that conditions having well-established associations with threat should develop in more females than males. We therefore searched for conditions in which identifiable threats constituted clear precipitating factors, to test whether more females than males developed these conditions. We found two conditions known to be caused by stress: post-traumatic stress disorder (PTSD) and chronic fatigue syndrome also known as myalgic encephalomyelitis (CFS/ME).
Both PTSD and CFS/ME demonstrate a female preponderance. PTSD follows trauma and consists of intense, long-lasting emotional and physical symptoms, typically flashbacks, hypervigilance, and nightmares, as well as major depression, anxiety, and sleep disorders. In a WHO WMHS study in 15 diverse societies (n > 72,900), females experienced PTSD more than males (OR = 2.6) (Seedat et al., Reference Seedat, Scott, Angermeyer, Berglund, Bromet, Brugha and Jin2009).
Sex differences in experience of PTSD however must control for the nature of the precipitating stimulus. A detailed meta-analysis (290 studies) of types of trauma producing PTSD in primarily WEIRD societies showed that for every type of trauma except childhood sexual abuse, more females than males developed PTSD (OR = 1.98) (Tolin & Foa, Reference Tolin and Foa2006). Further detail came from a Nordic study (n > 5,200) in which approximately twice as many women (25.6%) as men (13.2%) were categorized as having PTSD (Ditlevsen & Elklit, Reference Ditlevsen and Elklit2012). Using a continuous measure of PTSD, females experienced stronger PTSD than males with effect sizes varying from small to large based on type of trauma: disasters and accidents (d = 0.84), loss (d = 0.47), non-malignant disease (d = 0.47), chronic disease (d = 0.39), violence (d = 0.27), and overall (d = 0.60).
While the female preponderance of PTSD could be due to more females than males experiencing severe trauma, current evidence disputed this. In the meta-analysis in WEIRD countries, fewer females than males confronted severe traumas (OR = 0.77) (Tolin & Foa, Reference Tolin and Foa2006). Likewise, globally, females were less likely than males to die from fatal injuries (WHO, 2014a) or become disabled due to injuries (Haagsma et al., Reference Haagsma, Graetz, Bolliger, Naghavi, Higashi, Mullany and Alsharif2016).
CFS/ME is a debilitating, often life-long condition. Along with extreme exhaustion and weakness, the most common symptoms include immune activation (flu-like symptoms), intense pain (often migraines), severe sleep disturbance, depression or anxiety, concentration problems, and adverse reactions to chemicals (Chu, Valencia, Garvert, & Montoya, Reference Chu, Valencia, Garvert and Montoya2019; Friedman, Reference Friedman2019; Natelson, Reference Natelson2019). Based on a review of 13 Asian, Australian, North and South American studies, despite varying national prevalence rates, after puberty women were 3–4 times more likely than men to develop CFS/ME (Son, Reference Son2012) with a large effect size in an American study (OR = 4.51) (Reyes et al., Reference Reyes, Nisenbaum, Hoaglin, Unger, Emmons, Randall and Gantz2003). Onset typically occurred between ages 20 and 45 years following a specific threat: an infective illness (64% of cases), a major aversive life event (emotional stressor in the family or at work in 39% of cases), and a reaction to environmental toxins such as vaccines (20% of cases), with multiple precipitating stressors for some individuals. CFS/ME is not believed to be a disease in the typical sense, but rather a systemic reaction to stress (Cortes Rivera, Mastronardi, Silva-Aldana, Arcos-Burgos, & Lidbury, Reference Cortes Rivera, Mastronardi, Silva-Aldana, Arcos-Burgos and Lidbury2019). Prevalence rates of both PTSD and CFS/ME thus supported the prediction that more females than males exhibited extreme self-protective reactions to threats.
In conclusion, eight types of threat-related clinical condition, although maladaptive, may reasonably be interpreted as extreme self-protective reactions, and were more prevalent in females than males.
7. Discussion
7.1. Sex differences in self-protective responses
Our goal was to evaluate whether Campbell's (Reference Campbell1999) SAT applied not merely to physical aggression but more generally to all threats. By searching the literature for cross-cultural evidence with large samples, we tested the hypothesis that females demonstrate more self-protective reactions than males to major threats. We found that females exhibited stronger self-protective reactions than males to important biological and social threats; a personality style more geared to threats; stronger emotional responses to threat; and more threat-related clinical conditions suggestive of heightened self-protectiveness. That females expressed more effective mechanisms for self-protection is consistent with females' lower mortality and greater investment in childcare compared with males. Table 2 summarizes our major findings. As our hypothesis would predict, the magnitudes of sex differences in self-protective reactions were largest for the most potent threats to life – including heightened antibody production, reduced endurance of pain, and greater avoidance of direct competition, with fear, disgust, and crying sometimes also producing large effect sizes.
Table 2 Threats, self-protective responses, extreme protective reactions, and fatal conditions with estimates of adult female:male odds ratios (OR)*.
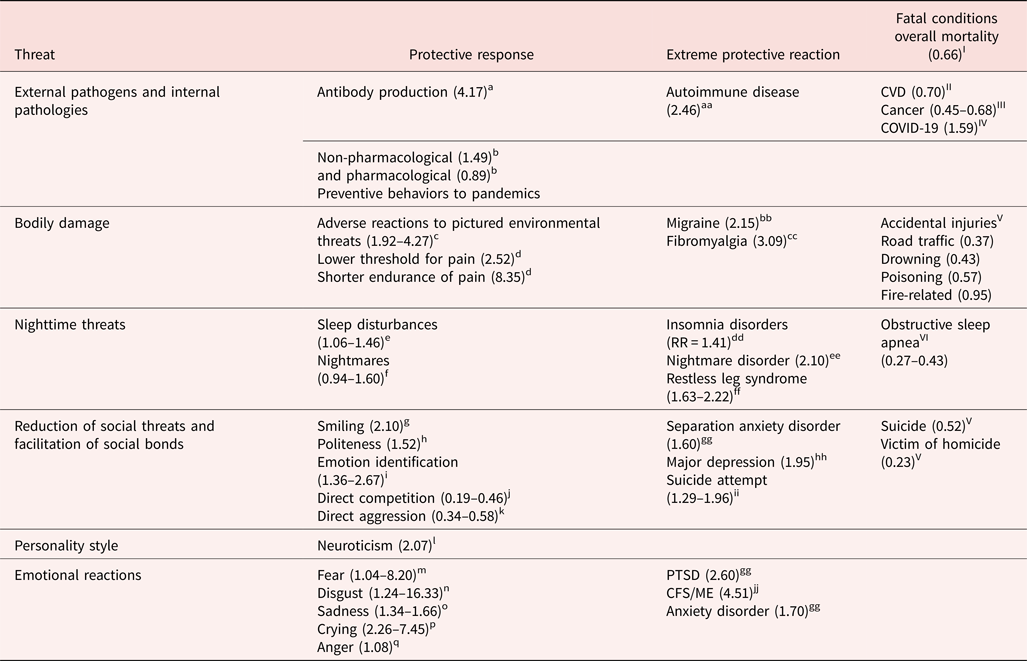
* Odds ratios >1 indicate higher prevalence in females than males and <1 indicate higher prevalence in males than females.
aYang and Kozloski (Reference Yang and Kozloski2011) antibodies; bMoran and Del Valle (Reference Moran and Del Valle2016) pandemic prevention; cBradley et al. (Reference Bradley, Codispoti, Sabatinelli and Lang2001), Gong et al. (Reference Gong, Wong and Wang2018), Gomez et al. (Reference Gomez, von Gunten and Danuser2013), Lang and Bradley (Reference Lang, Bradley, Coan and Allen2007), McManis et al. (Reference McManis, Bradley, Berg, Cuthbert and Lang2001) adverse reactions to photographs of environmental threat; dRiley et al. (Reference Riley, Robinson, Wise, Myers and Fillingim1998) pain; eWang et al. (Reference Wang, Li, Liu, Liu, Mao, Tu and Jiang2019) sleep disturbances; fSchredl and Reinhard (Reference Schredl and Reinhard2011) Hedge's g nightmares; gLaFrance et al. (Reference LaFrance, Hecht and Paluck2003) smiling; hLeaper and Robnett (Reference Leaper and Robnett2011) politeness; iConnolly et al. (Reference Connolly, Lefevre, Young and Lewis2019), Hall (Reference Hall1978), Hall et al. (Reference Hall, Carter, Horgan and Fischer2000), Sasson et al. (Reference Sasson, Pinkham, Richard, Hughett, Gur and Gur2010), Thompson and Voyer (Reference Thompson and Voyer2014) emotion identification; jKlege et al. (Reference Klege, Visser, Barron and Clarke2021), Deaner and Smith (Reference Deaner and Smith2013) direct competition; kArcher (Reference Archer2019) direct aggression; lKajonius and Johnson (Reference Kajonius and Johnson2018), Schmitt et al. (Reference Schmitt, Realo, Voracek and Allik2008) neuroticism; mArcher (Reference Archer2019), Arrindell et al. (Reference Arrindell, Eisemann, Oei, Caballo, Sanavio, Sica and Iwawaki2004), Brebner (Reference Brebner200) fear; nAl-Shawaf et al. (Reference Al-Shawaf, Lewis and Buss2018), Atari et al. (Reference Atari, Lai and Dehghani2020), Curtis and de Barra (Reference Curtis and de Barra2018), Egolf et al. (Reference Egolf, Hartmann and Siegrist2019) disgust; oBrebner (Reference Brebner2003), Lucas and Gohm (Reference Lucas and Gohm2000) sadness; pSharman et al. (Reference Sharman, Dingle, Baker, Fischer, Gracanin, Kardum and Vingerhoets2019), Van Hemert et al. (Reference Van Hemert, van de Vijver and Vingerhoets2011) crying; qArcher (Reference Archer2004) anger.
aaHayter and Cook (Reference Hayter and Cook2012) autoimmune disorder; bbStovner et al. (Reference Stovner, Nichols, Steiner, Abd-Allah, Abdelalim, Al-Raddadi and Doan2018) migraine; ccQueiroz (Reference Queiroz2013) fibromyalgia ddZhang and Wing (Reference Zhang and Wing2006) risk ratio insomnia disorder; eeLi et al. (Reference Li, Zhang, Li and Wing2010) Hong Kong ⩾3 nightmares/week; ffOhayon and Roth (Reference Ohayon and Roth2002) restless leg syndrome; ggSeedat et al. (Reference Seedat, Scott, Angermeyer, Berglund, Bromet, Brugha and Jin2009) separation anxiety disorder, PTSD, anxiety disorder; hhSalk et al. (Reference Salk, Hyde and Abramson2017) major depression; iiNock et al. (Reference Nock, Borges, Bromet, Cha, Kessler and Lee2008) non-fatal self-injury; Miranda-Mendizabal et al. (Reference Miranda-Mendizabal, Castellví, Parés-Badell, Alayo, Almenara, Alonso and Gili2019); jjReyes et al. (Reference Reyes, Nisenbaum, Hoaglin, Unger, Emmons, Randall and Gantz2003) CFS in the USA.
IGBD (2018) mortality; IIMosca et al. (Reference Mosca, Barrett-Connor and Kass Wenger2011) cardiovascular disease; IIIWHO (2021) cancer mortality; IVWilliamson et al. (Reference Williamson, Walker, Bhaskaran, Bacon, Bates, Morton and Inglesby2020); VWHO (2014a) for deaths from injuries, suicides, and homicides (we calculated ORs based on graph). We assume being a victim of homicide can be avoided to some extent through self-protective reactions. VILévy et al. (Reference Lévy, Kohler, McNicholas, Barbé, McEvoy, Somers and Pépin2015) obstructive sleep apnea.
Our test of Campbell's extended SAT is limited by our inability to examine every conceivable kind of threat, and by the incomplete availability of cross-cultural data. Nevertheless, we found evidence from many large samples and diverse cultures. Strikingly, despite intense searching, we failed to find large studies or meta-analyses that showed sex differences in the opposite direction to those expected from SAT. We conclude that the tendency for females to show more self-protective physiology, social interactions, personality styles, emotional reactions, and threat-related clinical conditions than males provides a strong first test for the extension of Campbell's SAT.
Many additional potential examples of greater female self-protectiveness occur in important areas that we excluded due to limited cross-cultural evidence. As examples, females more than males exhibit a lower threshold for detecting many sensory stimuli (Velle, Reference Velle1987); remain closer to home (Ecuyer-Dab & Robert, Reference Ecuyer-Dab and Robert2004); overestimate the speed of incoming stimuli, discuss threats and vulnerabilities more frequently, find punishment more aversive, demonstrate higher effortful control, and experience deeper empathy (Archer, Reference Archer2019); express greater concern over friend's (Hall, Reference Hall2011) and romantic partner's (Carpenter, Reference Carpenter2012) loyalty; and seek more frequent help (Möller-Leimkühler, Reference Möller-Leimkühler2002; Tamres, Janicki, & Helgeson, Reference Tamres, Janicki and Helgeson2002; Whiting & Whiting, Reference Whiting and Whiting1975).
Developmental evidence generally supported the prediction that females would exhibit greater self-protective reactions than males throughout life, but that the magnitude of the sex difference would increase following puberty when females can reproduce. Sex differences in immune functioning, pain, and nighttime awakenings appeared in childhood, but increased post-pubertally. Sex differences in politeness, emotional identification, and avoidance of confrontations emerged by early childhood, but in smiling only after puberty. Sex differences in neuroticism arose only after puberty, but girls displayed some components of neuroticism more than boys pre-pubertally. Girls experienced greater fear and disgust than boys pre-pubertally, but not sadness, for which the sex difference only clearly arose post-pubertally.
We did not systematically examine sex differences in children's clinical conditions due to lack of cross-cultural evidence. However, sex differences in some conditions, such as anxiety (Rescorla et al., Reference Rescorla, Achenbach, Ivanova, Dumenci, Almqvist, Bilenberg and Döpfner2007) and depression (Salk et al., Reference Salk, Hyde and Abramson2017), emerge only after puberty, whereas in others, such as immune functioning, shifts occur from childhood to puberty (Klein & Flanagan, Reference Klein and Flanagan2016).
7.2. Complexities in interpreting sex differences
That numerous sex differences conform to SAT is consistent with the thesis that self-protective reactions constitute a series of evolved female adaptations. If sex differences are to be fully understood however, additional factors must be included.
First, sex/gender is a multidimensional construct resulting from the continuing interplay between biological and environmental factors (Berenbaum & Beltz, Reference Berenbaum and Beltz2021; Eliot, Reference Eliot2009; Fausto-Sterling, Reference Fausto-Sterling2019; Hyde et al., Reference Hyde, Bigler, Joel, Tate and van Anders2019). Further, some of its important components and related constructs, including epigenetic configurations, hormonal balances, reproductive capacity, gender identity, gender roles, and sexuality, often fluctuate over the lifespan. Thus, menarche and menopause typically produce diverse changes for human females. Males too undergo transitions. Testosterone diminishes in bachelors who marry and even further after they father children (Gettler, McDade, Feranil, & Kuzawa, Reference Gettler, McDade, Feranil and Kuzawa2011; Gray, Kahlenberg, Barrett, Lipson, & Ellison, Reference Gray, Kahlenberg, Barrett, Lipson and Ellison2002). Because the studies we found do not define sex/gender and treat it as binary, we cannot specify which characteristics of sex/gender relate to self-protection. More precise structural, epigenetic, cellular, hormonal, cultural, social, emotional, cognitive, and behavioral indices will greatly improve understanding of the relation between self-protection and sex/gender.
Second, the magnitude of sex differences in self-protectiveness depends on local threats and safeguards which may differentially affect females and males. For example, in matrilineal societies where females receive greater protection from kin (Smuts, Reference Smuts1992), girls and women appear to be as directly competitive as their male counterparts in economic games (Klege et al., Reference Klege, Visser, Barron and Clarke2021). Likewise, when women's rights are protected by laws, the normally higher rates of men physically battering their female partners (WHO, 2014a) can disappear (Archer, Reference Archer2006). In impoverished societies that provide infants with both nutritional supplementation and breastmilk, females' immune systems are strengthened more than males' (Khulan et al., Reference Khulan, Cooper, Skinner, Bauer, Owens, Prentice and Affara2012; Osrin et al., Reference Osrin, Vaidya, Shrestha, Baniya, Manandhar, Adhikari and Anthony2005). Prevalence of physical illness itself varies depending on gender identity, role, and status (Mauvais-Jarvis et al., Reference Mauvais-Jarvis, Merz, Barnes, Brinton, Carrero, DeMeo and Klein2020) and societal kinship structure (Reynolds et al., Reference Reynolds, Wander, Sum, Su, Thompson, Hooper and Blumenfield2020). Even sex differences in mortality are influenced by societal factors, including patients', physicians', and researchers' belief systems (Mauvais-Jarvis et al., Reference Mauvais-Jarvis, Merz, Barnes, Brinton, Carrero, DeMeo and Klein2020); rates of female infanticide, sexual abuse of and violence against women (Solotaroff & Pande, Reference Solotaroff and Pande2014; WHO, 2014a); and men's willingness to seek help (Verbrugge, Reference Verbrugge1989).
More individual factors also likely regulate the magnitude of sex differences in self-protective reactions. For example, grandmothers typically invest more in daughters' than sons' young children, thereby tending to maximize their own fitness (Sear & Mace, Reference Sear and Mace2008). The extent of a particular grandmother's investment, however, should regulate her daughter's self-protective reactions. Similarly, although worldwide mothers care for infants (Wood & Eagly, Reference Wood and Eagly2002), a particular family's norms regarding the exclusivity of maternal responsibility for childcare likely influences maternal self-protective reactions. As another example, females may have lower thresholds than males to even perceive physical symptoms, social conflicts, or other threats, as extrapolation from research on pain would suggest (Riley et al., Reference Riley, Robinson, Wise, Myers and Fillingim1998). Nevertheless, the degree to which members of an individual's social network reward or punish females' versus males' expressions of vulnerability should regulate sex differences in even recognizing threats (Jansz, Reference Jansz and Fischer2000).
Third, more research is necessary to understand the societal impact of gender equality, measured by women's participation and power in public life, on self-protectiveness. Greater gender equality is associated with even fewer women than men committing suicide (Alothman & Fogarty, Reference Alothman and Fogarty2020); and increased proportions of women than men expressing concerns about environmental degradation (Chan et al., Reference Chan, Pong and Tam2019), reporting neuroticism (Costa et al., Reference Costa, Terracciano and McCrae2001; Schmitt et al., Reference Schmitt, Realo, Voracek and Allik2008), and experiencing depression (Salk et al., Reference Salk, Hyde and Abramson2017). These somewhat paradoxical findings require further inquiry. Gender equality however is intertwined with other ecological and cultural variables (Kaiser, Reference Kaiser2019) and may not adequately capture the realities of life of people in non-WEIRD societies (Markus, Reference Markus2021).
Fourth, a large overlap exists between the sexes in many self-protective reactions as is found in many studies of sex differences (Eliot, Reference Eliot2009; Hyde et al., Reference Hyde, Bigler, Joel, Tate and van Anders2019; Zell et al., Reference Zell, Krizan and Teeter2015). Consequently, a particular self-protective response cannot be predicted simply from knowing an individual's binary sex. Our evidence merely demonstrates that there is a population-wide tendency in the direction predicted by Campbell's theory.
Distinct self-protective responses, however, frequently co-occur. Myriad studies report links among somatic, social, neurotic, emotional, and clinical responses (Okur Güney, Sattel, Witthöft, & Henningsen, Reference Okur Güney, Sattel, Witthöft and Henningsen2019; Yunus, Reference Yunus2007). This suggests that aggregating self-protective reactions could more accurately describe the effect of greater self-protectiveness on females' than males' lives than simply comparing the sexes on only one self-protective response at a time. Thus, multivariate statistical techniques could provide a more qualitative distinction between the sexes (Del Giudice, Reference Del Giudice, Vander Laan and Wong2022).
Fifth, many female-prevalent illnesses and clinical conditions could impede women's ability to care for their children, thereby reducing their fitness. An adaptive approach however suggests that the benefits accrued from women's greater longevity outweigh the costs of chronic illness (Del Giudice, Reference Del Giudice2018; Nesse, Reference Nesse2005).
Overall, considering the diversity of our evidence, the consistency of our findings is striking. Accordingly, we regard sex differences in self-protective responses as important to investigate not only for theoretical reasons but also for practical value related to medical and psychological health (Clayton, Reference Clayton2016; Shansky & Murphy, Reference Shansky and Murphy2021).
An analysis of rates of COVID-19 illustrates how sex differences can vary widely in magnitude without undermining their significance. Both absolute frequencies and sex differences vary markedly by ethnicity, age, geographical location, socioeconomic status, and baseline health status even within the same country, as depicted in Figure 9 for England (Economist, Reference Economist2020).

Figure 9. Sex differences in frequency of COVID-19 infections across varied demographic factors in England. © The Economist Group Limited, London (November 21, 2020).
Despite this variation, women are almost always less likely than men to die within each demographic group. Thus, sex constitutes an important biological variable that can enhance causal understanding of a phenomenon which varies on many factors (Clayton, Reference Clayton2018).
Finally, further research is necessary to understand whether some threats elicit stronger self-protective reactions in men than women. Examples include male concerns over status and warfare. Current evidence however indicates no sex differences in the desire to attain status (Anderson, Hildreth, & Howland, Reference Anderson, Hildreth and Howland2015), even though the sexes may choose different strategies to achieve it (Benenson & Abadzi, Reference Benenson and Abadzi2020). Whether men are more concerned than women about imminent military attacks has not been investigated.
7.3. Complementary explanations
Our findings provide support for self-protective responses as a female adaptation. Alternative, but potentially compatible, explanations however could also apply.
First, males' higher levels of androgens, larger body size, more negative social interactions, greater risk-taking, and other sexually selected characteristics related to mate competition are expected to partially explain why males exhibit lower self-protectiveness than females (Dunsworth, Reference Dunsworth2020; Klein, Reference Klein2000; Kruger & Nesse, Reference Kruger and Nesse2006; Zuk, Reference Zuk2009). Individual and population variation among males in their optimal strategies and biological constraints will therefore contribute to explaining their lower self-protectiveness. Nevertheless, it is not simply degree of mate competition that produces sex differences in self-protectiveness. Rather, females invest in costly self-protective mechanisms, including immune functioning and conflict reduction measures, that elevate the probability of survival. Independent selective pressures on both male and female traits influence the nature and degree of sex differences in self-protectiveness.
Unquestionably, women's smaller size and muscle mass enhances their self-protectiveness with men. Nonetheless, sex differences in size and strength alone cannot explain all self-protective reactions. For example, in children where sex differences in size and muscularity are minimal, girls often exhibit more self-protective reactions than boys, including greater pain to vaccines (Chambers et al., Reference Chambers, Giesbrecht, Craig, Bennett and Huntsman1999) or politer language (Leaper & Smith, Reference Leaper and Smith2004; Whiting et al., Reference Whiting, Edwards, Ember, Erchak, Harkness, Munroe and Seymour1988).
Second, a “trauma hypothesis” might suggest that traumas specific to women increase their self-protectiveness. Worldwide, one in three women suffers domestic or sexual violence; 20% of girls are sexually abused; and reproductive-related morbidity and mortality are more frequent where women have fewer rights (WHO, 2014a). However, universally more men than women are exposed to lethal threats (WHO, 2014a). If trauma alone were responsible for greater self-protectiveness, then men should exhibit stronger self-protective responses than women in some areas, such as in response to violent social interactions or accidents which affect men more (WHO, 2014a), but they do not (Ditlevsen & Elklit, Reference Ditlevsen and Elklit2012). Many of the sex differences we reviewed therefore are not explicable by the trauma hypothesis.
Third, women's greater self-protectiveness could result from the high costs of pregnancy and lactation which could render women less physically, socially, emotionally, and clinically functional than men. This view is commonly held by medical and psychological professionals who perceive “self-protective” reactions as maladaptive or compensatory (Chesler, Reference Chesler1972; Cleghorn, Reference Cleghorn2021). If reproduction itself were the only critical factor however, sex differences in self-protective reactions would be highest during pregnancy and lactation, but this is not the case. Some sex differences in self-protectiveness appear by infancy (e.g., immune functioning, fear); others arise at puberty (e.g., smiling, sadness); some are maximal throughout the reproductive years (e.g., neuroticism, migraine); and others increase with age (e.g., insomnia).
Fourth, worldwide men tend to hold higher status than women, particularly in the public sphere (Fiske, Dupree, Nicolas, & Swencionis, Reference Fiske, Dupree, Nicolas and Swencionis2016; Rosaldo & Lamphere, Reference Rosaldo and Lamphere1974). This difference almost certainly heightens females' self-protective reactions compared to when females hold power such as within the confines of families or in matrilineal societies (Smuts, Reference Smuts1992). Status differences do not readily explain other socially self-protective reactions, however, such as females' greater fearfulness (Else-Quest et al., Reference Else-Quest, Hyde, Goldsmith and Van Hulle2006) or more accurate emotional identification (McClure, Reference McClure2000) from infancy onwards.
In sum, the evolutionary explanation offered by SAT complements other accounts of women's greater responses to threats by integrating divergent domains. Further, it views females' strategies as evolved adaptive functions, rather than being constrained features that are sub-optimal compared to those of males. It also helps resolve many apparent paradoxes in which women are less likely than men to die from pathogens, injuries, social conflicts, and suicide, yet more likely to experience physical symptoms, pain, sleep disturbances, avoidance of social conflicts, generalized worry, fear, disgust, and sadness, and make suicide attempts. SAT posits that stronger self-protective reactions to threat enhance survival, rendering females less vulnerable than males. Furthermore, SAT would predict that individuals who assume primary responsibility for children's well-being will increase their self-protective reactions so as to enhance their own survival.
7.4. Conclusion
Girls' and women's heightened responsiveness to physical and social threats, neuroticism, emotional reactions to threats, and related clinical conditions are often pathologized, that is, attributed to hysteria, mental health disorders, or other abnormalities, because men are the reference point (Chesler, Reference Chesler1972; Cleghorn, Reference Cleghorn2021). Our extension of Campbell's SAT suggests, by contrast, that females' “sensitivity” should be construed not as a constrained weakness or compromise enforced by reproductive biology but as a strategic approach to counteracting threats. Male traits, equivalently, that are often seen as the norm, are suboptimal for enhancing survival, but serve to maximize men's fitness given the constraints that they confront (Seager, Reference Seager, Barry, Kingerlee, Seager and Sullivan2019). Thus, independent consideration of each sex, along separate dimensions, is necessary for understanding the ways in which each sex's traits are optimized. Had the evidence existed, we would have compared the reproductive success of females who varied in their degree of reactions to threats, and not included males at all.
A richer understanding of the adaptive nature of female self-protection ultimately will depend on discovering its underlying mechanisms and how they may have co-evolved. Promising mechanisms undergirding self-protectiveness include sex chromosomes (Schurz et al., Reference Schurz, Salie, Tromp, Hoal, Kinnear and Möller2019), sex hormones (Klein & Flanagan, Reference Klein and Flanagan2016), centralized reactivity (Yunus, Reference Yunus2007), support of attachment figures (Archer, Reference Archer, Stroebe, Hansson and Stroebe2008; Bowlby, Reference Bowlby1980) and the larger community (Smuts, Reference Smuts1992), and degree of responsibility for offspring survival (Allman et al., Reference Allman, Rosin, Kumar and Hasenstaub1998).
In conclusion, in response to a wide diversity of threats, human females exhibit greater self-protective responses than males. This finding suggests an opportunity for researchers and clinicians to better understand the adaptive nature of diverse female traits, both in humans and other species.
Acknowledgements
The authors sincerely thank Dr. Barbara Finlay and the six reviewers for their excellent recommendations which greatly improved the manuscript.
Funding statement
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Conflict of interest
None.












Target article
Self-protection as an adaptive female strategy
Related commentaries (22)
An expanded “staying alive” theory (SAT) underplays complexity in Homo sapiens
Beyond individual sex differences: “Staying alive theory” as an adaptive complex
Biological sex, by-products, and other continuous variables
Female advantage in threat avoidance manifests in threat reaction but not threat detection
Harm or protection? Two-sided consequences of females' susceptible responses to multiple threats
Only as a last resort: Sociocultural differences between women and men explain women's heightened reaction to threat, not evolutionary principles
Pathological complexity and the evolution of sex differences
Psychological and behavioral implications of self-protection and self-enhancement
Sex differences are insufficient evidence of ecological adaptations in human females
Sex differences in longevity are relative, not independent
Sex-dependent selection, ageing, and implications for “staying alive”
Societies also prioritize female survival
Somatic maintenance/reproduction tradeoffs and human evolution
Staying alive enhances both women's and men's fitness
Staying alive includes adaptations for catalyzing cooperation
The pregnancy compensation hypothesis, not the staying alive theory, accounts for disparate autoimmune functioning of women around the world
The “staying alive” theory reinforces stereotypes and shows women's lower quality of life
Toward a more domain-specific conceptualization of female traits: A commentary on Benenson et al. (2022)
Women amid the COVID-19 pandemic: Self-protection through the behavioral immune system
Women need to stay alive and protect reproductive choice
Women take risks to help others to stay alive
“Staying alive” in the context of intimate partner abuse
Author response
Females undergo selection too