


R1. Introduction
We are grateful to the commentators for raising a wide variety of issues. Because these generally fall into a number of distinct categories, we organized our response around them, as follows:
Section R2: Domains of cognition
Section R3: Tasks for test batteries
Section R4: Domain-specificity versus domain-generality
Section R5: What is g/G? (More) on the necessity of validation
Section R6: g/G and brain size
Section R7: g and biological fitness
Section R8: Cultural intelligence
Whenever commentators provided input in more than one of these categories, we discuss them in more than one of the sections. Some of the commentaries show that we did not always present our points with sufficient clarity, so we also take the opportunity to make these clarifications as well as to highlight what we did not claim.
The complicated nature of the issues is illustrated by the fact that different commentators made confident claims that are mutually incompatible. For instance, Ramus and Arden & Zietsch argue that the evolution of g is no puzzle at all, and that it is most likely present in all organisms with a nervous system, Jacobs & Gärdenfors similarly argue that it is hardly surprising or controversial that some individuals consistently perform well, and Pepperberg recites the impressive examples from bird cognition that to her necessarily imply the presence of general intelligence in at least some birds. On the other hand, other commentators question whether there is evidence for g or G in any nonhuman species at all (Huber) or feel the importance of g is overestimated (Amici, Call, & Aureli [Amici et al.]).
The goal of our target article was to make progress on understanding animal intelligence without getting bogged down in terminological debates on what exactly g represents. We suggested complementing the psychometric approaches, which are a necessary first step to establish the possibility of general intelligence, with a variety of validation measures and more demanding tests that look for domain-generality of cognitive processes. This is even more important because animal studies are unlikely to ever reach the sophistication in terms of tests and the sample sizes needed to attain the practical level of utility achieved by human intelligence testing (pace Arden & Zietsch). We will focus, therefore, especially on these forward-looking points.
R2. Domains of cognition
One of the unresolved issues in nonhuman psychometrics is what an ideal test battery should look like. The criteria are obvious: (1) A test battery should be composed of tasks from a broad range of domains rather than tasks that re-sample performance in the same domain, and (2) the tasks should reliably assess the cognitive abilities they are supposed to assess, both within and across species.
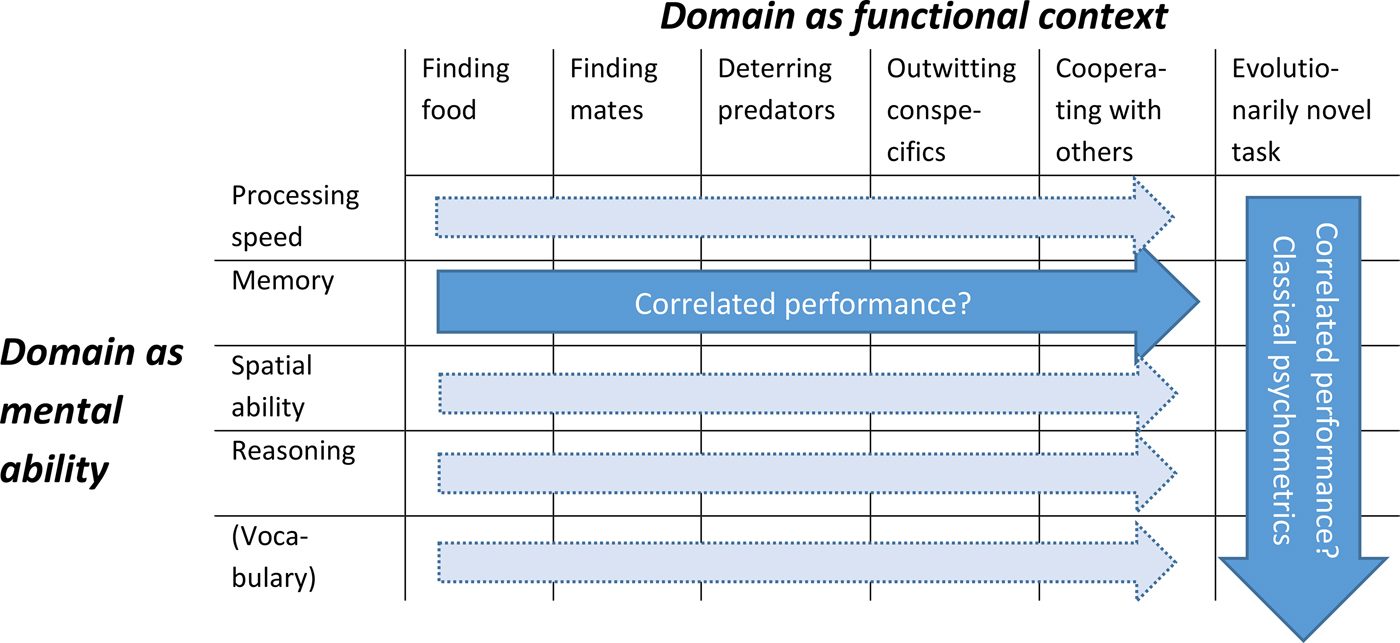
To fulfill criterion (1), it is crucial to know what a domain is. In the classical psychometric tradition, a domain of mental ability refers to a statistically derived group factor on which a set of tasks show strong loadings. For instance, Deary et al. (Reference Deary, Penke and Johnson2010) referred for humans to the group factors processing speed, memory, spatial ability, reasoning, and vocabulary. Thus, the straightforward approach to identify domains in nonhuman animals is to likewise use large batteries of tasks and identify the factor structure statistically. This bottom-up, data-driven approach in animals is ambitious because it requires very large sample sizes. However, this does not mean that, in the meantime, we are unable to correctly identify g in animals. Empirical tests with human subjects have revealed that as long as there is sufficient variety in the tasks of test batteries, g factors derived from different test batteries are almost perfectly correlated (Johnson et al. Reference Johnson, te Nijenhuis and Bouchard2008).
Evolutionary approaches often consider domains to be functional contexts (see also Wang). These can be very specific such as finding food, finding mates, deterring predators, outwitting conspecifics, or cooperating with others (Figure R1), or rather broad, such as in the classic trichotomy in physical, spatial, and social cognition, as followed by Tomasello & Call (Reference Tomasello and Call1997). Obviously, the individual may fail to show a particular ability that it is shown to possess in one context, when tested in a functional context different from the one for which it evolved. Thus, it remains challenging to identify domains.
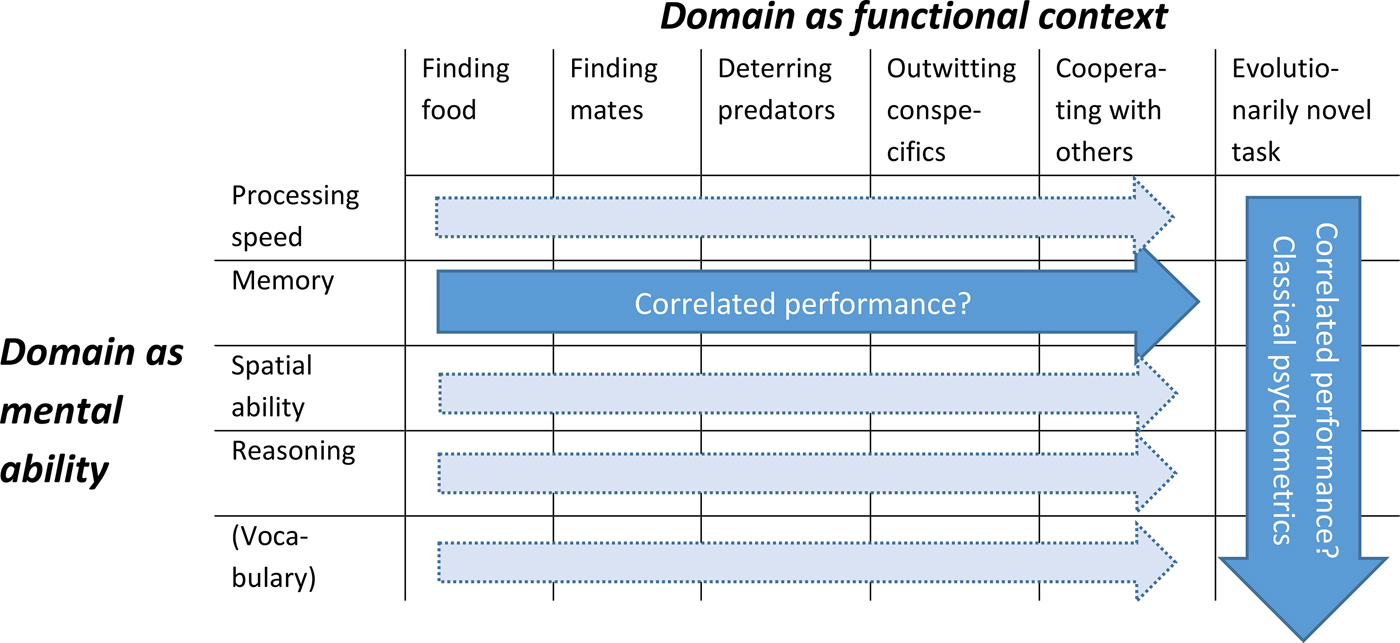
Figure R1. Domains can refer to statistically derived group factors such as processing speed, memory, spatial ability, reasoning, or vocabulary in humans (Deary et al., Reference Deary, Penke and Johnson2010) or to evolutionarily functional contexts. Classical psychometric studies (vertical arrow), in humans and nonhuman animals, typically refer to domain as mental ability. From an evolutionary perspective, however, it is equally informative to ask to what extent a given cognitive ability is correlated across functional contexts (horizontal arrows). Because performance across functional contexts is likely to vary, evolutionarily novel tasks are most suitable for classical psychometric studies that aim at testing individuals across mental ability domains.
Importantly, it is not useful to decide a priori that one notion of domain would be more correct than the other one. Rather, acknowledging that a domain can refer to both a mental ability (as in the psychometric tradition) and a functional context (as in evolutionary approaches; see also the proposal by Shuker, Barrett, Dickins, Scott-Phillips, & Barton [Shuker et al.] or Hauser's examples of abilities that “cut across domains”) allows us to ask the questions that are at the core for understanding the evolution of general intelligence: Under what circumstances can a species that evolves a cognitive ability for a specific context generalize this ability to other contexts as well (cf. Stevens et al. Reference Stevens, Kennedy, Morales and Burks2016)? Can this be true for some abilities, but not others? And are these the same species that also show correlated performance across mental abilities and thus show psychometric g?
Thus, rather than committing to one specific notion of a domain, it is better to identify ways to combine these perspectives to ultimately better understand the evolution of intelligence. Furthermore, this combined perspective will also clarify many of the issues surrounding domain-specificity and domain-generality, and modularity (sect. R4).
Due to the sample size problems in identifying domains using the psychometric approach, some have defined domains a priori (e.g., Herrmann et al. Reference Herrmann, Call, Hernández-Lloreda, Hare and Tomasello2007). However, as we point out in the target article, empirical data do not necessarily confirm that tasks supposed to tap into one such domain actually also cluster around it (see sect. 2.4.1 in target article). Although we agree in principle with the proposal of Amici et al. that it is important to take multifactorial approaches into account (see also Lee & Chabris), we think more empirical work is needed to identify which tasks indeed measure particular cognitive operations (such as inference) or mental capacities (such as working memory). In fact, most tasks probably tap into more than one operation and/or capacity, and it therefore is highly unlikely that one specific test measures one specific cognitive ability, as also pointed out by Huber and Ramus. Even for humans there is often a lack of agreement on how to quantify specific cognitive constructs such as, for instance, working memory (Oberauer et al. Reference Oberauer, Schulze, Wilhelm and Süss2005).
Finally, Figure R1 also helps us to answer how ecologically valid a test should be. An often-made claim is that in order to fully appreciate a given species' cognitive potential, one should look only at problems of high ecological relevance to this species (e.g., Sternberg). However, when we look only at ecologically relevant problems we will never be able to disentangle whether we are dealing with a primary module or with the result of true and flexible problem-solving ability. As indicated in Figure R1, there are at least two ways to demonstrate this kind of flexibility. A psychometric approach would compare different abilities in evolutionarily novel tasks, precisely in order to avoid tapping into predominantly hard-wired solutions to recurring adaptive problems. This approach is particularly powerful in combination with validation studies (sect. R5). The other one is to compare the same ability across functional domains (e.g., asking whether the excellent memory abilities in the ecologically relevant context of caching in food-caching birds also generalize to other functional contexts). Although this second possibility has received surprisingly little attention (but see Stevens et al. Reference Stevens, Kennedy, Morales and Burks2016), it presents an excellent opportunity to externally validate a psychometrically derived g factor (see also Locurto, sect. 2.5 in target article, and sect. R4).
R3. Tasks for test batteries
Many commentators have proposed specific tasks, and we think these are excellent proposals that will help construct broad and diverse test batteries. Van Horik & Lea stress the necessity to also include what they call knowing tasks, which can assess rule-based learning or generalization. Such knowing tasks, as for instance reversal learning, also have an additional advantage, linked to criterion 2 of a good test battery: that the tasks should reliably assess the cognitive abilities they are supposed to assess, both within and across species (sect. R2). The advantage is that knowing tasks are less vulnerable to producing variation in performance due to differences in sensory-motor specializations between species (see Shuker et al., and sect. 2.4.2 in the target article for a detailed discussion of Macphail's critique of species comparisons). We are confident that many of these issues can be resolved, particularly in closely related species, but also agree that it remains problematic for comparisons of very diverse species, such as cephalopods, with completely different sensory-motor traits and even body plans (Holekamp & Miikkulainen).
Commentators Jacobs & Gärdenfors highlight causal cognition tasks; Huber, reasoning by exclusion; and Buskell & Halina, tasks suggestive of de-coupled representations. Tasks in which birds excel may also be useful additions to existing test batteries. As stressed by Pepperberg, corvids and parrots are particularly prime candidates for exhibiting high levels of general intelligence, and we look forward to future empirical studies that explicitly address this question with psychometric test batteries combined with predictive validation studies (Locurto; see also sect. R5). Pennycook & Rand add the possibility to include studies of cognitive control and metacognition.
Commentators Pennycook & Rand also argue that variation in performance between individuals may not only reflect cognitive capacity per se, but also the willingness to engage in effortful cognitive processing when a simpler, more routine solution seems available. They thus refer to the concept of need for cognition, which is an individual predisposition rather than a cognitive ability, even though in humans it is correlated with general intelligence (Hill et al. Reference Hill, Foster, Sofko, Elliott and Shelton2016b). This link is entirely consistent with our view that cognitive skills emanating from general cognitive capacity (i.e., the downward pathway in Figure 3 of the target article) are ontogenetically constructed. Therefore, individuals with higher levels of need for cognition will more systematically expose themselves to situations that require effortful processing and are, therefore, more likely to establish a larger and more powerful set of cognitive skills. To what extent substantial variation in willingness to engage in effortful processing is also available in nonhuman animals remains to be established, but an increasing body of research suggests that this may account for the systematic differences in cognitive performance between captive and wild primates such as orangutans (van Schaik et al. Reference van Schaik, Burkart, Damerius, Forss, Koops, van Noordwijk and Schuppli2016).
R4. Domain-specificity versus domain-generality
Several commentators (Arden & Zietsch; Hauser; Jacobs & Gärdenfors; Martins & Di Paolo) think we advocate a strict dichotomy between a mind made up of modules and one that has general intelligence. This is surprising, because in the target article, we devote an entire section (1.2.2 including Table 1) to exactly why such a dichotomy is not helpful for the debate, and in Figure 3 we present what we believe is the most promising working model given the current state of evidence on g/G in human and nonhuman animals. This working model suggests that the actual skill set of an individual can be construed as a mix of primary or secondary modules. Whereas both can be sensitive to experience (as detailed in Table 2 in the target article and unlike the claim by Lewis, Al-Shawaf, & Anderson [Lewis et al.]), this sensitivity differs: Primary modules are experience-expectant; secondary modules, experience-dependent.
The extent of domain-specificity and domain-generality obviously critically depends on the notion of domain one adopts (see sect. R2). This distinction between domain as mental ability or as functional context, as detailed in Figure R1, is also reflected in the many different notions of modularity that have been the focus in the modularity debate ever since Fodor. Proposals range from defining modules as adaptive behavioral functions localized in delimited neural areas to seeing them as being nothing else than components and processes into which a phenotype can be decomposed (Barrett Reference Barrett, Zeigler-Hill, Welling and Schackelford2015). When referring to domain specificity, we focus on functional specialization in the biological, ultimate sense (as used, for instance, in the commentary by Amici et al.), rather than referring to any other specific notion of domain-specificity (Lewis et al.). This allows us to ask under which conditions a cognitive adaptation to a specific problem, – for instance, the impressive memory of food-caching birds – can also be used in a different context. From comparative studies, we know that cognitive abilities in nonhuman animals sometimes do generalize to other domains, sometimes do so partially, and sometimes do not at all (Cauchoix & Chaine Reference Cauchoix and Chaine2016; Stevens et al. Reference Stevens, Kennedy, Morales and Burks2016). When cognitive abilities do generalize, domain-general cognition emerges, and the fundamental question we are interested in is under what conditions this can happen.
If it is not straightforward to define domain-specificity or modularity, this is even truer for domain-generality in cognition or general intelligence. Not unexpectedly, several authors have commented on this. Sternberg suggests that “an appropriate way to look at the intelligence of any organism is to look at how well it adapts to the range of environments it confronts” (see also Bar-Hen-Schweiger, Schweiger, & Henik [Bar-Hen-Schweiger et al.]). This is problematic because intelligence is only one way among many to adapt to the environment. Under this conception, the thickening of the fur in autumn clearly is an adaptation to the environment but most would agree it has nothing to do with intelligence. For the same reason, it is problematic to refer to domain-generality as phenotypic plasticity (Buskell & Halina) because it is merely one aspect of that. Equally unhelpful is the proposal by Sternberg to look at skills that are relevant to an animal's everyday adaptation. If we go back to our food-caching birds and assess only their memory capacities in the context of food caching, we are simply not able to exclude the possibility that we are dealing with a highly domain-specific capacity.
It is this latter issue that makes psychometric g studies so appealing to comparative psychologists. As highlighted in the target article (sect. 2.4 and sect. 2.5) and by several commentators, this approach comes with a whole suite of issues, addressed here in sections R5, R6, and R7. However, at least in humans, g is well established, known to reflect general intelligence as broadly defined in the target article (following Gottfredson Reference Gottfredson1997 and Nisbett et al. Reference Nisbett, Aronson, Blair, Dickens, Flynn, Halpern and Turkheimer2012), has a variety of genetic correlates, including those linked to neurobiological features, and is a good predictor for various measures of life outcome and thus shows high levels of predictive validity. In nonhumans, more validation is critically required, as also emphasized by Locurto.
R5. What is g/G? (More) on the necessity of validation
A common concern not only to many commentators, but also to ourselves (see sect. 2.5 in the target article), is that once one has statistically established a reliable g factor in a given species, based on adequate tasks from a broad range of domains, it does not necessarily follow that this corresponds to general intelligence as broadly defined and predictive for a variety of life outcome measures, as is the case in humans. Furthermore, even for humans, despite the predictive validity of g, it does not necessarily follow that the predominant hierarchical CHC model (see para. 1.1.1 in the target article) is the only compelling interpretation. Some suggest that a statistical g simply results from variation in non-cognitive factors, such as developmental stability, and thus reflects general health (Lewis et al.), or genetic load or heterozygosity (Arslan, von Borell, Ostner, & Penke [Arslan et al.]). Several commentators discuss plausible alternative models to the hierarchical CHC model. Thus, the process overlap theory of Kovacs & Conway (see also Kovacs & Conway [Reference Kovacs and Conway2016] and the responses in the same issue of Psychological Inquiry) argue that performance in a specific task is determined by several cognitive abilities, and different tasks resample the same abilities to different extents. The mutualism model of van der Maas et al. (Reference van der Maas, Dolan, Grasman, Wicherts, Huizenga and Raijmakers2006) (see Ramus; Shuker et al.) argues that g arises because of mutual interactions during cognitive development. These alternatives thus see g as a composite of independent but overlapping or interacting processes, rather than as a single top-down ability (which Bar-Hen-Schweiger et al. see as the ability to engage in mental manipulation). Hence, they regard g more as a developmental consequence or emergent property, rather than as an underlying latent variable, and are fully compatible with the constructive nature of domain-general cognition.
Nonetheless, these varying interpretations still argue for the presence of some domain-general ability, even if it is entirely emergent, and remain compatible with the various neurobiological and genetic correlates of g. For instance, if g arises as a result of process overlap because cognitive tests from different domains tap the same domain-general executive processes such as working memory, this would exactly correspond to domain-generality as identified by the horizontal arrow in Figure R1.
The exact nature of g continues to be a major puzzle (Deary et al. Reference Deary, Cox and Ritchie2016). But we would argue that this puzzle doesn't have to be fully solved for studies of domain-general processes to be useful, provided g in nonhuman animals survives future tests of external validation, as we discuss next. Some argue that emergent properties or statistical constructs cannot be meaningful individual-level variables subject to natural selection (Arden & Zietsch; Ramus). We do not think this is correct, because even if g is an emergent property of several cognitive and even non-cognitive processes, natural selection should evaluate the degree to which these processes are compatible or even how they synergize to produce organized and appropriate behavior. Indeed, each of the models for g discussed previously is compatible with natural selection having honed g. Hence, considering g a measurable property of an organism is not reification.
These ambiguities in the interpretation of g necessarily spill over into interpretations of animal studies, especially in light of the weaknesses of intraspecific studies, as highlighted by Arden & Zietsch, Lee & Chabris, and Lewis et al. Nonetheless, as we have argued in section 2.4 of the target article, the fact that all purely interspecific studies produce clear evidence for G is incompatible with the notion of g as an artifact of testing the same basic ability or of inadequate test batteries. One can, of course, criticize the use of the rate of naturally occurring behaviors as applied in some G studies (e.g., Huber; Shuker et al. See also Reader et al. [Reference Reader, Hager and Laland2011] for a discussion of the advantages and disadvantages of this procedure). However, if this method would only produce noise, one would not find strong correlations with brain size or aspects of executive functions, or that G is the principal locus of selection in the macroevolution of primate intelligence (Fernandes et al. Reference Fernandes, Woodley and te Nijenhuis2014). Moreover, the anomaly of the lack of success of the mixed intraspecific and interspecific studies to generate a common main factor has been solved by Woodley of Menie, Fernandes, te Nijenhuis, Aguirre, & Figueredo. They suggested that variables with floor or ceiling effects may obscure differences in general intelligence across species because they cannot load on g. Their analysis supports this idea because species differences are especially striking for tests that load highly on g. Overall, then, the increasing plausibility of the idea that g and G can be equated automatically supports the argument that animals have something that closely resembles human g, and may even be homologous to it. This position would also be consistent with the increasing knowledge of the nature of cortical development and especially plastic responses to external inputs during brain development (Anderson & Finlay Reference Anderson and Finlay2014). It also should serve to refute the alternative interpretations of G as totally unrelated to g, offered by Arden & Zietsch and Lewis et al.
In order to resolve current debates, we believe empirical work is needed that confirms the presence of domain-general processes. First, if g were all about health, myelinization, or developmental stability – that is, non-cognitive features – we would expect a correlation between all cognitive abilities, regardless of whether they derive from primary or secondary modules. But if we were dealing with general intelligence, the pairwise correlations between abilities deriving from primary modules should be weaker (largely because they should all be at ceiling or floor values).
Second, more specific tests can be done to provide external validation for the interpretation of domain-general intelligence, along the lines outlined in the target article (sect. 2.5, Table 7) and as also stressed by Locurto. We can derive additional ones from the commentators' comments. Arslan et al. propose to use genetically uniform strains and mutation accumulation lines to help clarify the role of genetic load, and Lee & Chabris propose to use genome-wide association studies to test if distinct cognitive abilities are genetically correlated and thus potentially the target of natural selection. More specifically, Pennycook & Randsuggest to focus on meta-cognitive abilities; van Horik & Lea stress the usefulness of reversal learning paradigms; and Jacobs & Gärdenfors, causal reasoning. Pepperberg summarizes additional fruitful paradigms in which several bird species excel. Bar-Hen-Schweiger et al. propose to also include object manipulation. This is intriguing because recent results show that object manipulation complexity is indeed correlated with brain size (Heldstab et al. Reference Heldstab, Kosonen, Koski, Burkart, van Schaik and Isler2016), but to what extent mental manipulation can be seen as a direct extension of object manipulation requires additional research, in particular in nonhuman animals. For interspecific studies, species differences in basal ganglia–based circuitry may provide further insight into the evolution of G, as highlighted by Nordli & Todd.
In sum, we acknowledge and agree that the mere presence of a first PCA factor (g) in intraspecific psychometric studies is not sufficient evidence for general intelligence, but that such an interpretation critically requires evidence for predictive or criterion-related validity as stressed by Locurto and outlined in section 2.5 in the target article. However, we also argue that if a g factor becomes manifest in psychometric studies, and if this g is not an artifact and shows predictive and criterion-related validity, how exactly the positive intercorrelations arise no longer matters for the claim that g was the target of selection. In other words, it may well be that g is an emergent property of the central neural system, and we in fact think it is unlikely that it can be reduced to any specific psychological or biological trait or construct, and thus agree on this with Ramus or Arden & Zietsch. But this does not imply that g as potentially emergent property cannot be the target of selection. This view is supported by the majority of results from interspecific G studies.
R6. g/G and brain size
Brain size is reliably associated with G in interspecific primate studies, and less strongly with g in intraspecific studies. Matzel & Sauce argue that brain size cannot be important in intelligence. First, they argue that Neanderthals had larger brains than humans, but were “cognitively inferior” to modern humans. However, cultural intelligence can explain this difference because modern humans had much larger groups (Mellars & French Reference Mellars and French2011), and social inputs play a crucial role in affecting the size of a population's skill repertoires (see also Henrich Reference Henrich2016). Thus, differences in brain size (provided they also extended to differences in relative brain size) can be compensated for by differences in the quality and quantity of inputs during development. This observation also deals with their second objection, namely that brain size predicts only a modest proportion of variation in human g.
The second argument by Matzel & Sauce as to why brain size should not matter for intelligence was that intelligence can look quite similar in lineages with very different absolute or relative brain sizes, particularly in birds versus mammals. This can partly be explained by major between-lineage differences in neuron densities, between cetaceans and other mammals, and between mammals and birds (Olkowicz et al. Reference Olkowicz, Kocourek, Lučan, Porteš, Fitch, Herculano-Houzel and Němec2016). Accordingly, comparative studies on relative brain size within birds are consistent with the presence of general intelligence in this lineage (Lefebvre et al. Reference Lefebvre, Reader and Sol2004). Nonetheless, it would be useful to examine the degree to which more extensive tests of general intelligence in birds (e.g., Isden et al. Reference Isden, Panayi, Dingle and Madden2013; Shaw et al. Reference Shaw, Boogert, Clayton and Burns2015) show the same structure of cognition as in humans and presumably other mammals. If multiple independent origins can indeed be demonstrated, as suggested by Matzel & Sauce, this would support the contention that beyond a certain level of complexity, domain-general processes evolve to take over the control of procedures that serve a useful function in many different domains (Wang).
R7. g and biological fitness
If general intelligence is an adaptation, then we expect some link to fitness. Thus, one intuitive way of assessing predictive validity of g is to ask if individuals scoring higher on g have better survival and higher reproductive success. This basic idea is compelling, but both empirical and conceptual caveats need to be carefully considered.
In humans, reproductive success is often no longer maximized in modern societies due to the demographic transition (Coale Reference Coale, Eatwell, Milgate and Newman1989). Survival, however, continues to be a high individual priority and, therefore, is a better measure for this link in humans. There is in fact extensive evidence that g scores predict survival or longevity (Deary Reference Deary2008).
If general intelligence is an adaptation in nonhuman species, too, then the estimates of this ability should also correlate with fitness measures. This correlation has in fact been proposed as a test of the predictive validity of the concept (Locurto). Tests in the wild presuppose that we can actually estimate general intelligence in wild animals. This may be feasible in birds (Isden et al. Reference Isden, Panayi, Dingle and Madden2013; Shaw et al. Reference Shaw, Boogert, Clayton and Burns2015), but will be extremely difficult in many other species that show strong neophobia in the wild (Forss et al. Reference Forss, Schuppli, Haiden, Zweifel and van Schaik2015). Moreover, as stressed by van Horik & Lea, it is more difficult to control for confounds or selective participation, or to retest the same individuals (see also Rowe & Healy Reference Rowe and Healy2014).
Yet, we must also be careful for conceptual reasons. General intelligence is developmentally constructed, and investment in increased intelligence may trade off against other vital activities. It is, therefore, quite conceivable that estimates of general intelligence need not show a positive relationship with fitness but, rather, an inverted U-shaped relationship. Thus, if investment in general intelligence and the brain requires resources that also positively affect other vital processes such as growth and reproduction, this can have a negative impact on an individual's fitness. The positive relationship in modern humans may thus be something of an anomaly, perhaps linked to the absence of such tradeoffs (although it could well be that it existed for reproduction). Indeed, the emerging work examining the link between cognitive abilities and fitness (Morand-Ferron & Quinn Reference Morand-Ferron and Quinn2015) produces mixed results. One reason for this may be that a direct positive relationship with fitness is expected for primary modules, but not for secondary modules. This topic is worth exploring in greater depth if practical obstacles can be overcome (see also Morand-Ferron et al. Reference Morand-Ferron, Cole and Quinn2015; Rowe & Healy Reference Rowe and Healy2014).
R8. Cultural intelligence
The presence of high general intelligence in animals poses something of a conundrum. Individual problem solving based purely on general cognitive ability will only rarely produce useful behavioral innovations, but will often also carry risks of injury or poisoning. Moreover, rarely invented innovations are not transmitted to the next generation; only the ability to make such inventions is. The cultural intelligence hypothesis, therefore, suggests that general intelligence is most likely to evolve where the process of problem solving is socially canalized – that is, in species that strongly rely on social learning, because social canalization increases the odds of actually finding a solution. It stresses that social information can efficiently guide exploration during development. It is, therefore, as Whiten correctly points out, an enabling hypothesis, which concerns the acquisition process of cognitive skills (and the conditions selecting for them), but is silent on the actual nature of the cognitive benefits. The latter could often be cognitive flexibility serving as a cognitive buffer, allowing animals to survive in seasonal habitats, as suggested by Holekamp & Miikkulainen (cf. van Woerden et al. Reference van Woerden, Willems, van Schaik and Isler2012). However, this functional benefit should have been favored in many species, and the cultural intelligence hypothesis argues that selection is more likely to produce this adaptive benefit for behavioral flexibility where costs remain low due to many opportunities to acquire social information.
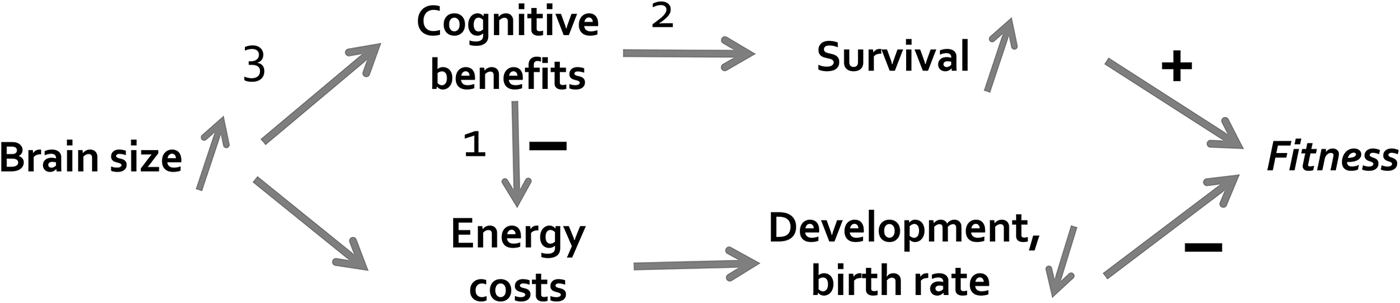
Cognition requires brains, and brains are energetically expensive. Thus, the significant increase in energy need accompanying increased brain size must be met, directly or indirectly. As shown in Figure R2, natural selection would, therefore, favor those mechanisms that either provide a direct energy boost (for instance, ecological effects of cognition leading to reduced fluctuations in food intake: arrow 1) or allow for reduced allocation to growth and reproduction due to improved survival (arrow 2). The far greater efficiency provided by skill acquisition through social learning rather than inefficient and risky innovation (arrow 3) is another selective benefit highlighted by the cultural intelligence hypothesis. This argument clearly implies, although we did not point this out explicitly, that the cultural intelligence hypothesis is, like almost all evolutionary hypotheses, a ceteris paribus hypothesis: Holding all other conditions constant, the availability of social information should make it easier to evolve the supporting infrastructure for increased behavioral flexibility in a particular lineage.
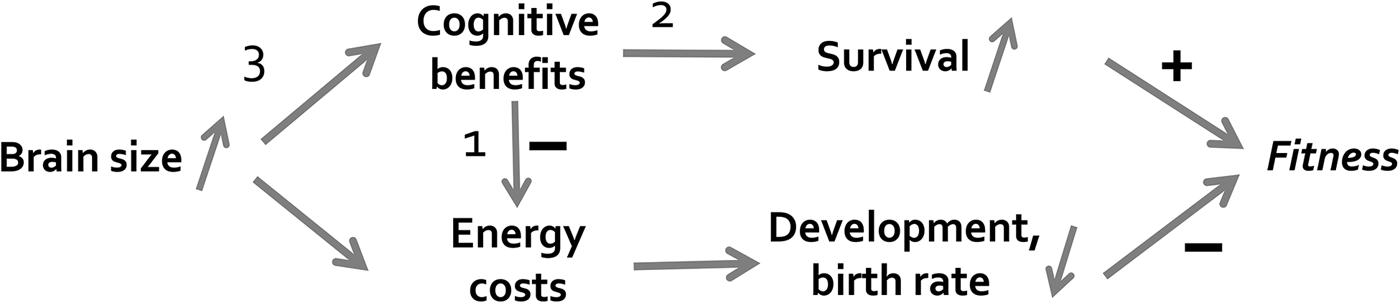
Figure R2. Processes that could be affected by selection for increased brain size: (1) an immediate effect of improved cognition on reduction in energy costs; (2) an unusually large fitness benefit from possessing cognitive skills; or (3) an unusually efficient translation of brain tissue into cognitive skills, as argued by the cultural intelligence hypothesis. The argument may hold especially for domain-general processes, because the latter appear to require exceptional amounts of brain tissue.
There may also be other conditions favoring the evolution of intelligence. Cephalopods, especially octopus, have no contact between generations, and mustelids disperse at a very young age and then remain solitary, yet show signs of strong cognitive performance (Holekamp & Miikkulainen; cf. Holekamp et al. Reference Holekamp, Dantzer, Stricker, Yoshida and Benson-Amram2015). In the expensive brain framework, this would suggest unusual payoffs from exploration and innovation (arrow 2 in Figure R2), or unusually low risks of exploration – which are testable predictions. These seemingly intelligent solitary species, therefore, would appear to be exceptions to the cultural intelligence hypothesis that should be acknowledged, although the impressive imitation ability of solitary octopus (Fiorito & Scotto Reference Fiorito and Scotto1992) is puzzling. The possibility of alternative evolutionary pathways to intelligence makes it even more important to explicitly deploy extensive test batteries to examine if these species truly show evidence of general intelligence, or whether we find specialized (albeit quite impressive) domain-specific cognitive adaptations.
R9. Conclusions
Our goal is to understand the evolutionary origin of general intelligence. The case is still open, even though the body of evidence suggests we also find it in other species and that the cultural intelligence approach can help us understand why it arose in some species but not in others. But identifying the evolutionary origin of g or G in nonhumans is not the full answer to how the human mind evolved. Humans are more than g (Amici et al.; Gardner; Pennycook & Rand) and other components are important too, first and foremost language, but also components summarized as multiple intelligences by Gardner. In principle, the same approach as taken here can be applied to each of these components, by asking what the interspecific distribution of this trait is, and what factors may best explain this distribution. For general intelligence, the most likely factor is brain size, whereas for prosocial attitude, allomaternal care seems to explain interspecific distribution (Burkart et al. Reference Burkart, Allon, Amici, Fichtel, Finkenwirth, Heschl, Huber, Isler, Kosonen, Martins, Meulman, Richiger, Rueth, Spillmann, Wiesendanger and van Schaik2014).
Cultural intelligence effects are particularly pronounced in humans. Among others, this is evident in the Flynn effect, and the strong influence of environment on general intelligence, particularly early in ontogeny when individuals still cannot seek out an environment that matches their abilities (Flynn Reference Flynn2016). Human cultural intelligence is supported by additional adaptations such as language and pedagogy, which involves strong theory-of-mind abilities and intentional teaching as pointed out by Estes & Bartsch (see also Burkart et al. Reference Burkart, Hrdy and van Schaik2009). For humans in particular, one would expect, therefore, that socio-cognitive abilities predict general intelligence, and Fernández-Berrocal, Cabello, & Gutiérrez-Cobo present evidence for humans supporting this prediction (for interspecific tests of this link, Arslan et al. suggest joint phylogenetic tree analyses of sociality and variance explained by g, but this will not work because the amount of variance explained is not a good measure for how much “g” a given species “has,” as detailed in the target article (sect. 2.4.1)).

Target article
The evolution of general intelligence
Related commentaries (28)
G and g: Two markers of a general cognitive ability, or none?
G but not g: In search of the evolutionary continuity of intelligence
A pointer's hypothesis of general intelligence evolved from domain-specific demands
An all-positive correlation matrix is not evidence of domain-general intelligence
Coexistence of general intelligence and specialized modules
Contemporary evolutionary psychology and the evolution of intelligence
Disentangling learning from knowing: Does associative learning ability underlie performances on cognitive test batteries?
Domains of generality
Evolution, brain size, and variations in intelligence
General intelligence does not help us understand cognitive evolution
General intelligence is a source of individual differences between species: Solving an anomaly
General intelligence is an emerging property, not an evolutionary puzzle
Genomic data can illuminate the architecture and evolution of cognitive abilities
Habit formation generates secondary modules that emulate the efficiency of evolved behavior
Hierarchy, multidomain modules, and the evolution of intelligence
It's time to move beyond the “Great Chain of Being”
Negative results are needed to show the specific value of a cultural explanation for g
Of mice and men, nature and nurture, and a few red herrings
Taking a multiple intelligences (MI) perspective
The evolution of analytic thought?
The evolution of fluid intelligence meets formative g
The evolution of general intelligence in all animals and machines
The false dichotomy of domain-specific versus domain-general cognition
Theory of mind: A foundational component of human general intelligence
Understanding the relationship between general intelligence and socio-cognitive abilities in humans
When does cultural transmission favour or instead substitute for general intelligence?
Where is the evidence for general intelligence in nonhuman animals?
“Birdbrains” should not be ignored in studying the evolution of g
Author response
Future directions for studying the evolution of general intelligence