



Introduction
Since 2008, Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) has spread across much of North America, South America, and Europe (Grassi et al. Reference Grassi, Giongo and Palmieri2011; Hauser Reference Hauser2011; Walsh et al. Reference Walsh, Bolda, Goodhue, Dreves, Lee and Bruck2011; Calabria et al. Reference Calabria, Máca, Bächli, Serra and Pascual2012; Cini et al. Reference Cini, Anfora, Escudero-Colomar, Grassi, Santosuosso, Seljak and Papini2014; Deprá et al. Reference Deprá, Poppe, Schmitz, De Toni and Valente2014; Asplen et al. Reference Asplen, Anfora, Biondi, Choi, Chu and Daane2015; Funes et al. Reference Funes, Kirschbaum, Escobar and Heredia2018). Unlike most Drosophilidae, D. suzukii is a serious pest of soft fruits and berries. Female flies use a serrated ovipositor to lay eggs in ripening rather than overripe fruits, causing the fruit to spoil and become unmarketable. Male flies are more readily recognised by their characteristic wing colourisation that gives the species their common name, spotted-wing drosophila. Costs associated with crop losses and increased pesticide use worldwide exceed one billion USD annually (Bolda et al. Reference Bolda, Goodhue and Zalom2010; Lee et al. Reference Lee, Bruck, Curry, Edwards, Haviland, Van Steenwyk and Yorgey2011; Cini et al. Reference Cini, Anfora, Escudero-Colomar, Grassi, Santosuosso, Seljak and Papini2014; Wiman et al. Reference Wiman, Dalton, Anfora, Biondi, Chiu and Daane2016). To date, studies have demonstrated that fruits of 151 plant species from 31 families are at least somewhat susceptible to infestation (Lee et al. Reference Lee, Dreves, Cave, Kawai, Isaacs and Miller2015; Kenis et al. Reference Kenis, Tonina, Eschen, van der Sluis, Sancassani and Mori2016; Lee and Sial Reference Lee and Sial2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017; Elsensohn and Loeb Reference Elsensohn and Loeb2018).
The full potential for continued range expansion by D. suzukii in North America and abroad has yet to be determined. Dynamic acclimation to cold conditions may allow D. suzukii to successfully overwinter in colder climates and further expand its geographic range (Stockton et al. Reference Stockton, Wallingford and Loeb2018). Overwintering populations of D. suzukii have been found in numerous locations in Canada and northern Europe where winter temperatures drop well below −17 °C and laboratory studies confirm physiological adaptations suitable to cold hardiness (Hamby et al. Reference Hamby, Bellamy, Chiu, Lee, Walton and Wiman2016; Rossi Stacconi et al. Reference Rossi Stacconi, Kaur, Mazzoni, Ometto, Grassi and Gottardello2016; Thistlewood et al. Reference Thistlewood, Gill, Beers, Shearer, Walsh and Rozema2018). Fruits and berries grown commercially or that are endemic to boreal regions at the presumed limit of their current range may be at risk as D. suzukii continues to extend its range northward.
Host choice by D. suzukii is generally thought to be a factor of changes in fruit characters associated with ripening, particularly firmness or skin resistance (penetration force), sweetness (Brix), and acidity (pH) (Lee et al. Reference Lee, Bruck, Curry, Edwards, Haviland, Van Steenwyk and Yorgey2011, Reference Lee, Dreves, Cave, Kawai, Isaacs and Miller2015; Burrack et al. Reference Burrack, Fernandez, Spivey and Kraus2013). Fruits susceptible to D. suzukii tend to be limited to soft-skinned fruits and berries, with some exceptions, particularly when fruits are damaged or overripe (Lee et al. Reference Lee, Dreves, Cave, Kawai, Isaacs and Miller2015; Kenis et al. Reference Kenis, Tonina, Eschen, van der Sluis, Sancassani and Mori2016; Lee and Sial Reference Lee and Sial2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017; Elsensohn and Loeb Reference Elsensohn and Loeb2018). Other fruit characters thought to be important for host selection and suitability to D. suzukii, including size, shape, structure, volatile odour profile, and colour of fruits (Poyet et al. Reference Poyet, Le Roux, Gibert, Meirland, Prévost, Eslin and Chabrerie2015). Fruit firmness is also a limiting factor in oviposition success by D. suzukii (Entling et al. Reference Entling, Anslinger, Jarausch, Michl and Hoffmann2018).
Most previous studies of fruit preference by D. suzukii that investigated fruit colour did not quantify colour parameters (Lee et al. Reference Lee, Bruck, Curry, Edwards, Haviland, Van Steenwyk and Yorgey2011, Reference Lee, Dalton, Swoboda-Bhattarai, Bruck, Burrack and Strik2016; Karageorgi et al. Reference Karageorgi, Bräcker, Lebreton, Minervino, Cavey and Siju2017; Jaffe et al. Reference Jaffe, Avanesyan, Bal, Feng, Grant and Grieshop2018). Other studies that investigated colour preference in D. suzukii used measures of perceived colour based on human vision (Kirkpatrick et al. Reference Kirkpatrick, McGhee, Hermann, Gut and Miller2016, Reference Kirkpatrick, Gut and Miller2018). Parameters such as hue, saturation, chroma, and brightness are comparative measures of colour appearance based on human perception (Fairchild Reference Fairchild2005). Colour appearance models compare similarities or differences among perceived colours and assume identical environmental conditions (Fairchild Reference Fairchild2005; Kelber and Osorio Reference Kelber and Osorio2010; Lunau Reference Lunau2014; Cuthill et al. Reference Cuthill, Allen, Arbuckle, Caspers, Chaplin and Hauber2017). The colour of an object is a property of both the object being perceived and the perception of the animal that perceives it (Glover and Whitney Reference Glover and Whitney2010). Drosophila Fallén and human visual sensitivity differ; therefore, we chose a percentage reflectance across a range of wavelengths as an objective measure of fruit colour, independent of the observer (Paulk et al. Reference Paulk, Millard and van Swinderen2013; Little et al. Reference Little, Chapman and Hillier2018).
Drosophila suzukii is highly polyphagous, infesting fruits of widely diverse characters and colours (Lee et al. Reference Lee, Dreves, Cave, Kawai, Isaacs and Miller2015; Kenis et al. Reference Kenis, Tonina, Eschen, van der Sluis, Sancassani and Mori2016; Lee and Sial Reference Lee and Sial2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017; Elsensohn and Loeb Reference Elsensohn and Loeb2018). We hypothesise that fruit reflectance may play an important role in host selection among diverse fruit species available in an area. Host choice among blueberries (Vaccinium corymbosum Linnaeus (Ericaceae)) by D. suzukii is partially dependent upon visual contrast cues or conspicuousness against the background of foliage (Little et al. Reference Little, Chapman and Hillier2018). Similar behaviour in female Delia radicum Linnaeus (Diptera: Anthomyiidae) flies has been described for host choice based on “appropriate-inappropriate landings,” with appropriate landings defined as being on host plants and inappropriate landings as being on nonhost plants (Finch and Collier Reference Finch and Collier2000).
We identified 11 fruit-producing plant species of interest that were found across Nova Scotia and Newfoundland and Labrador, Canada. Pin cherries, Prunus pensylvanica Linnaeus (Rosaceae), and bittersweet nightshade, Solanum dulcamara Linnaeus (Solanaceae), are common in hedgerows and wooded areas throughout much of Canada. Lowbush or wild blueberry, Vaccinium angustifolium Aiton (Ericaceae), is found in wooded areas and hedgerows and is grown commercially throughout much of Canada. Haskap, Lonicera caerulea Linnaeus (Caprifoliaceae), also known as blue honeysuckle and honeyberry, is a relatively new commercial and ornamental berry plant in North America but widely grown in northern Asia (Celli et al. Reference Celli, Ghanem and Brooks2014). Amelanchier alnifolia Nuttall (Rosaceae), commonly known as Saskatoon berry or service berry, is native to boreal regions of North America but is now grown in northern regions of Europe (Bakowska-Barczak and Kolodziejczyk Reference Bakowska-Barczak and Kolodziejczyk2008; Lavola et al. Reference Lavola, Karjalainen and Julkunen-Tiitto2012). Crowberry, Empetrum nigrum Linnaeus (Ericaceae), is an ecologically and culturally important species in the northern heathlands of North America, Europe, and Asia (Koskela et al. Reference Koskela, Anttonen, Soininen, Saviranta, Auriola, Julkunen-Tiitto and Karjalainen2010; Buizer et al. Reference Buizer, Weijers, van Bodegom, Alsos, Eidesen and van Breda2012). Cloudberry, Rubus chamaemorus Linnaeus (Rosaceae), also known as bakeapple in Newfoundland and Labrador, is a perennial herbaceous plant found in boreal regions of North America, Europe, and Asia (Thiem Reference Thiem2003). Fruit colour changes from red to orange-yellow as it ripens. Strawberries, Fragaria × ananassa Duchesne (Rosaceae); highbush blueberries, Vaccinium corymbosum Linnaeus (Ericaceae); raspberries, Rubus idaeus Linnaeus (Rosaceae); and cherries, Prunus avium Linnaeus (Rosaceae), are commercially grown in boreal regions of Canada.
Here we tested whether fruit colour is a determinant of host choice in the absence of background foliage. We also examined whether aspects of fruit colour can be used during host search as a reliable proxy for other fruit characters. We quantified potential preference of fruits to D. suzukii based on host selection in choice assays and host suitability based on larval success in no-choice assays. Host preference and suitability measures were then compared to fruit characters including firmness, sweetness, acidity, and colour. We also assessed the susceptibility of a variety of fruits that are of commercial or cultural importance in boreal regions across Canada.
Key message
Fruits growing in boreal regions are suitable hosts for D. suzukii and are at risk of infestation as its invasive range expands.
How D. suzukii find and discriminate among diverse potential host fruits is poorly understood.
Characters such as sweetness, acidity, fruit firmness, and colour can be signals for fruit ripeness and host suitability.
This study gives first evidence that D. suzukii can use fruit colour reflectance as cues for host suitability across multiple fruit species.
Selection for fruit varieties with greater reflectance across the visible spectrum may reduce attractiveness to D. suzukii.
Materials and methods
Fruit and insect sources
Ripe fruits from 11 plant species were collected and tested between 2014 and 2017. Pin cherry, bittersweet nightshade, and wild blueberry were picked from wild plants in the Annapolis Valley, Nova Scotia, Canada. Haskap and Saskatoon berry were picked at u-pick farms in Nova Scotia. Crowberry was picked by a colleague in Iqaluit, Nunavut, Canada. Fresh cloudberry was not available due to adverse weather conditions in 2015 and 2016 throughout much of Newfoundland and Nova Scotia. We obtained frozen cloudberry that had been picked in Newfoundland in 2014. Host selection assays and oviposition and larval development assays were also conducted using bittersweet nightshade (Solanum dulcamara) collected from wild plants in Nova Scotia after hearing anecdotal accounts of D. suzukii associated with these plants. Strawberry (Fragaria × ananassa varieties “Wendy,” “Honeyeye,” and “Cabot”) and cherry (P. avium variety “Stella”) were obtained from local growers at farm markets in the Annapolis Valley, Nova Scotia. Highbush blueberry (V. corymbosum varieties “Jersey” and “Coville”) and raspberries (R. idaeus variety “Caroline”) obtained from commercial growers in Nova Scotia were also used to determine relative preference in host selection assays.
Drosophila suzukii used in the experiments were obtained from colonies maintained since 2013 at Acadia University from stocks originally reared by the Kentville Research and Development Centre (Agriculture and Agri-Food Canada, Kentville, Nova Scotia) and replenished with wild flies reared from local fruit in 2016. Colonies were housed in 250-mL Drosophila flasks (Genesee Scientific, San Diego, California, United States of America) containing 50-mL Formula 4–24 Instant Drosophila Medium (Merlan Scientific, Mississauga, Ontario, Canada) mixed with 50-mL dH2O. Approximately two hours prior to the start of each assay, reproductively mature females aged up to two weeks were removed from colonies created one month earlier and held in 30-mL vials without access to food or water. Vouchers are held on deposit at the Acadia University Wildlife Museum, Wolfville, Nova Scotia.
Fruit characters
Fruits were assessed for colour, firmness (gfmm2), sweetness or sugar content (brix), and acidity (pH) to elucidate characters that could affect susceptibility and suitability to D. suzukii (Arnó et al. Reference Arnó, Solà, Riudavets and Gabarra2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017, Reference Little, Chapman and Hillier2018). Ten intact ripe fruits were selected randomly from each plant species. Mean values of each character for each fruit species were used for comparison and analysis. Due to limited quantities of available undamaged nightshade fruits, fruit characters were not measured for that species.
First, fruit colour was quantified. Reflectance spectra for each fruit were measured with an Alta II reflectance spectrometer (Vernier Software & Technology, Beaverton, Oregon, United States of America) to quantify colour. All reflectance values were measured in a dark room with the spectrometer providing the sole light source. Percentage reflectance measures were obtained for seven visible colour wavelengths (470–700 nm).
Fruits were then tested for firmness using a Wagner Fruit Penetrometer FT02 gram force gauge (Wagner Instruments, Greenwich, Connecticut, United States of America), sweetness was measured with a Westover Model RHR – 32ATC handheld brix refractometer with automatic temperature compensation (Cole-Parmer, Montréal, Québec, Canada), and acidity was measured using a Fisher Scientific Accumet Basic AB15 pH metre (Fisher Scientific, Toronto, Ontario, Canada) (Lee et al. Reference Lee, Bruck, Curry, Edwards, Haviland, Van Steenwyk and Yorgey2011; Little et al. Reference Little, Chapman, Moreau and Hillier2017, Reference Little, Chapman and Hillier2018).
Fruit preference
We used two-choice trap assays to assess the relative preference of D. suzukii among fruit species (Abba et al. Reference Abba, Olusakin, Dare, Mohammed, Ajayi and Okpanachi2012; Brodie et al. Reference Brodie, Smith, Lawrence and Gries2015). Two-choice assays were conducted within a sealed 600-mL airtight arena (18 × 12 × 6.5 cm) (Little et al. Reference Little, Chapman, Moreau and Hillier2017, Reference Little, Chapman and Hillier2018). Each arena contained two clear plastic traps, each consisting of a 30-mL portion cup with a truncate 200-μL pipette tip inserted through the lid (Little et al. Reference Little, Chapman, Moreau and Hillier2017, Reference Little, Chapman and Hillier2018). Approximately 10 mL of fruit was placed in each trap (Little et al. Reference Little, Chapman, Moreau and Hillier2017, Reference Little, Chapman and Hillier2018). Fifteen mated female D. suzukii were placed in each arena (Little et al. Reference Little, Chapman, Moreau and Hillier2017, Reference Little, Chapman and Hillier2018). Host choices were checked after 24, 48, and 72 hours. Each fruit species was tested against each other and against a blank (control) trap. Three to five replicates were completed for each trial. Due to limited fruit availability and differences in fruit phenology, we were unable to conduct trials with all possible combinations of fruit. Response index (RI) scores ranging from −1 to +1 were calculated as a measure of host preference based on Dweck et al. (Reference Dweck, Ebrahim, Kromann, Bown, Hillbur and Sachse2013), where
 $${\rm{RI = \;}}\left( {{\rm{A - B}}} \right){\rm{/T}}$$
$${\rm{RI = \;}}\left( {{\rm{A - B}}} \right){\rm{/T}}$$A and B are the number of flies in each trap
T is the total number of flies per trial
Fruit suitability
Approximately 10 mL of each fruit and two sexually mature, mated female D. suzukii were placed in a 50-mL Drosophila vial (Genesee Scientific) capped with cellulose acetate plugs. Flies were removed after 72 hours, and fruits were visually inspected under a dissecting microscope at 120–250 times magnification for eggs and larvae. Fruits were visually inspected weekly for six weeks thereafter for emerging flies. Nondestructive examination methods were employed to reduce risk of damage to developing larvae. Adult flies were removed for counting and sexing. We assessed 10 replicates for each fruit, with each vial representing one replicate.
Statistical analysis
We accounted for effect of treatment (fruit type), time, and trial on retention indices using repeated measures Friedman χ 2 analyses. We used Kruskal–Wallis χ 2 tests and Tukey and Kramer (Nemenyi) post hoc tests (P < 0.05) to assess differences in host selection among fruit types. Fractional analysis and multiple-factor analyses of variance were used to assess relationships between host selection and interactions among fruit characters. All other data were analysed with one-way analysis of variance and Tukey post hoc tests performed using RStudio (Version 1.1.419, https://rstudio.com/products/rstudio/download; using R version 3.4.3 (www.r-project.org)).
Results
Fruit characters
Ripe fruits of all species tested had higher mean percentage reflectance values at longer wavelengths than at shorter wavelengths (Fig. 1A). Mean percentage reflectance values of crowberry and wild blueberry were relatively consistent across wavelengths tested, with no strong peak discernable for either fruit species (Fig. 1A). Pin cherry and Saskatoon berry were the sweetest, although differences in sweetness among pin cherry, Saskatoon berry, and cherry were not significant (Fig. 1B). Saskatoon berry and wild blueberry were the least acidic (Fig. 1C). Cloudberry was most difficult to puncture, whereas haskap was very easily punctured (Fig. 1D).
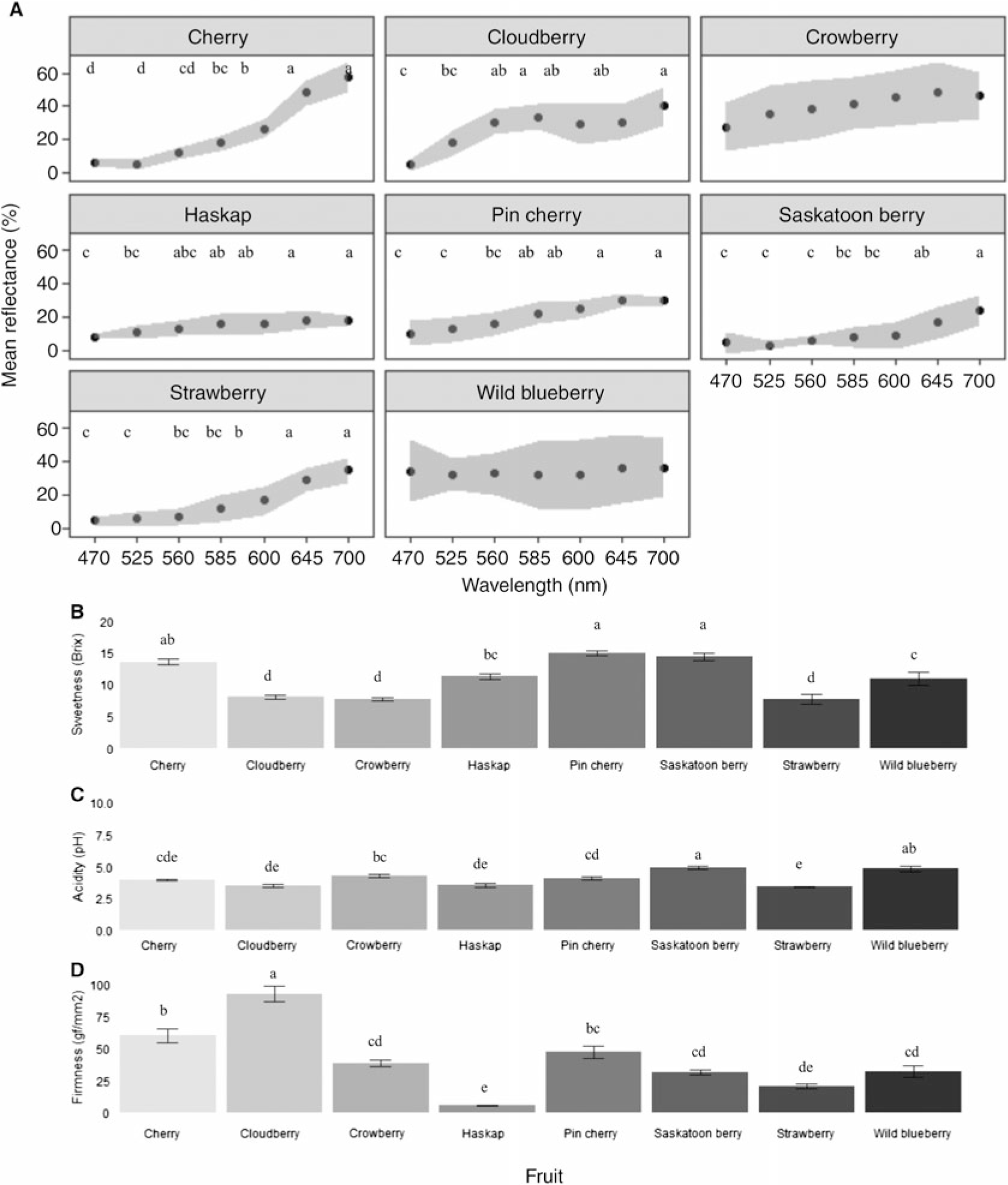
Fig. 1. Comparison of fruit colour and characters. A, Spectra of mean percentage reflectance (± standard error) at each wavelength. Differences were observed in reflectance among wavelengths for each fruit and among fruits at each wavelength (see Table 3) (two-way analysis of variance, fruit: F 7,544 = 65.39, P < 0.0001, wavelength: F 1,544 = 306.41, P < 0.0001, interaction: F 7,544 = 16.81, P < 0.0001). Differences were also observed among fruits sweetness, acidity, and firmness as follows. B, Brix levels (sweetness) (one-way analysis of variance F 7,72 = 30.31, P < 0.0001); C, acidity (pH) (one-way analysis of variance F 7,72 = 19.09, P < 0.0001); D, firmness (resistance to puncture) (one-way analysis of variance F 7,72 = 46.82, P < 0.0001). Whiskers denote minimum and maximum response index values. Different letter values above the figure denote significant differences among fruits (Tukey and Kramer (Nemenyi) post hoc, P < 0.05).
Among ripe fruits, percentage reflectance was strongly correlated across short-wavelength colours (blue–orange (470–600 nm)) and across long-wavelength colours (red–deep red (645–700 nm)) (Supplementary Table 1). Fruits with low reflectance values for blue (470 nm), therefore, also had proportionally low reflectance values for cyan (525 nm) and green (560 nm). However, reflectance was not correlated between short- and long-wavelength colours (Supplementary Table 1). Therefore, reflectance values at short wavelengths such as blue (470 nm) are not predictive of reflectance values at longer wavelengths such as red (645 nm).
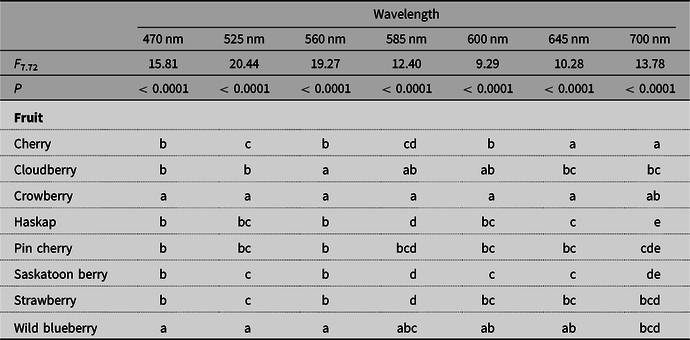
Fruit colour across most of the measured spectra was interrelated with other fruit attributes, irrespective of the fruit species (Table 2). Patterns of relative proportions of reflectance measures during ripening are fruit species-specific or variety-specific and can be a reliable proxy for fruit ripeness and quality (Li et al. Reference Li, Lee and Wang2014; Hu et al. Reference Hu, Zhai, Zhao and Wang2018). Among the fruits tested, fruit sugar content or sweetness (brix) was positively correlated with cyan (525 nm) to orange (600 nm), which is well within the visual range of Drosophila species (Hernández de Salomon and Spatz Reference Hernández de Salomon and Spatz1983). In a separate study, we found that the visual range of D. suzukii is consistent with that of D. melanogaster (Little et al. Reference Little, Rizzato, Charbonneau, Chapman and Hillier2019). Fruit firmness (gf/mm2) is positively correlated with green (560 nm) to deep red (700 nm) and thus is also detectable within the visual range of Drosophila. Fruit acidity is positively correlated with blue (470 nm). Further testing would be required to confirm if correlations also occur in the ultraviolet range.
Table 1. Mean response index scores (± standard error) for two-choice trials of each potential fruit combination. Positive response index values represent net attraction (shown in bold) towards the fruit listed in the first column versus the fruit listed in the top row of the table.

Table 2. Correlations among reflectance measures at each wavelength versus other fruit attributes, response index values during two-choice trials (see Fig. 2A, Table 1), and adult D. suzukii emergence during fruit suitability trials. Pearson’s rank correlation; R is on first line and P is on second line. Significant results are shown in bold.

Table 3. Differences were observed in reflectance among fruits at each wavelength (two-way analysis of variance, fruit: F 7,544 = 65.39, P < 0.0001, wavelength: F 1,544 = 306.41, P < 0.0001, interaction: F 7,544 = 16.81, P < 0.0001).
Fruit preference
Mean retention indices for each two-choice fruit trial showed that preference for any given fruit was in part dependent upon the alternative available (Table 1). Mean retention indices for each fruit type from two-choice assays were consistent across time periods measured (Friedman χ 2 = 0, df = 2, P = 1); therefore, host selection results, hereafter, are presented for retention indices at 72 hours. Fewest flies were found in highbush blueberry and pin cherry with mean retention indices not statistically different than empty control traps (Fig. 2A). Numerical counts of D. suzukii were highest in strawberry, Saskatoon berry, raspberry, and bittersweet nightshade; however, differences were significant only between Saskatoon berry and highbush blueberry or pin cherry (Fig. 2A). Response indices were not correlated with fruit firmness overall (Spearman’s rank correlation, Rs = −0.07, P = 0.87), fruit sweetness (Rs = −0.21, P = 0.62), or fruit acidity (Rs = −0.19, P = 0.65). However, in two-choice assays with fruits of differing characteristics, the relatively softer fruit was consistently preferred over firmer fruits (t-test, t = 2.3, df = 208.9, P = 0.03) and relatively more acidic fruits were preferred over those with higher pH (t = −3.4, df = 441.4, P < 0.001). No preference was observed between sweeter and less sweet fruits in two-choice tests (t = −1.1, df = 209.1, P = 0.29). Response indices were negatively correlated with percentage reflectance across most of the visible spectrum (Table 2). Percentage reflectance within short-wavelength (470–560 nm) and long-wavelength (575–700 nm) ranges were highly correlated for fruits tested (Supplementary Table 1).
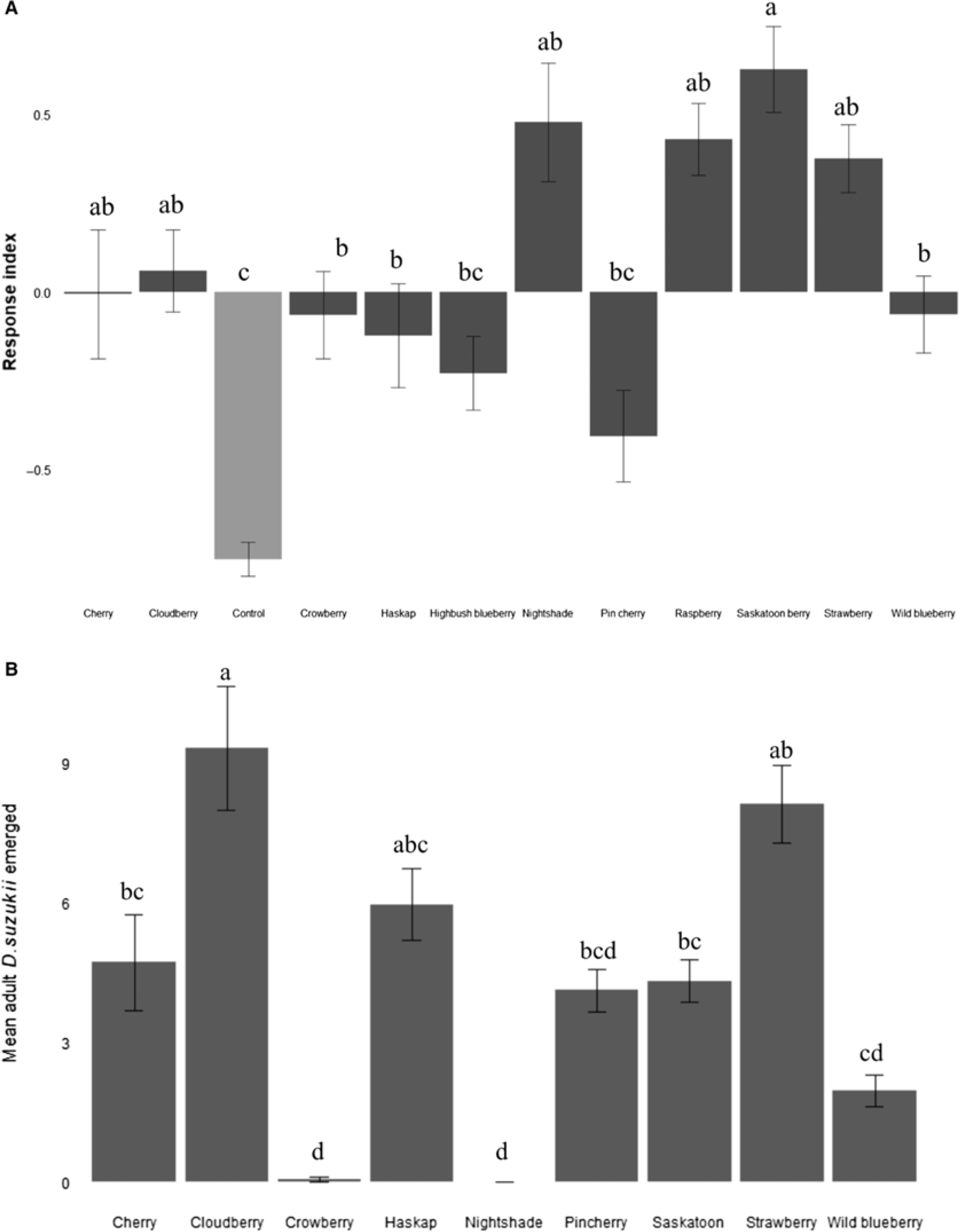
Fig. 2. A, Mean response indices for fruits and controls in two-choice attraction assays with adult female unmated Drosophila suzukii (n = 10 per fruit species). A positive response index score indicates preference for that fruit. Different letter values above the figure denote significant differences among fruits (Kruskal–Wallis χ 2 = 79.29, df = 11, P < 0.0001, Tukey and Kramer (Nemenyi) post hoc, P < 0.05). B, Mean adult Drosophila suzukii emerged from equal amounts of different fruits. Different letter values above the figure denote significant differences among fruits (analysis of variance; F 8,81 = 12.21, P < 0.0001, Tukey and Kramer (Nemenyi) post hoc, P < 0.05).
Fruit suitability
We were not able to accurately count eggs, larvae, and pupae in all fruit species using nondestructive methods, so fruit suitability was assessed based on emergence of adult flies. Adult D. suzukii that emerged from test fruits in no-choice assays were removed for counting and sexing. Counts differed among fruits; however, emergence of female and male D. suzukii were consistent within fruit species (two-way analysis of variance, fruit: F 8,162 = 19.78, P < 0.0001; sex: F 1,162 = 0.28, P = 0.60; fruit:sex: F 8,162 = 0.53, P = 0.84). We observed eggs, larvae, and emerging adults in all fruit species except bittersweet nightshade. Highest numbers of adult D. suzukii emerged from cloudberry, strawberry, and haskap; however, differences among strawberry, haskap, cherry, pin cherry, and Saskatoon berry were not significant (Fig. 2B). Ten mL of cloudberry produced a maximum count of 41 emergent adult flies.
We found no correlation between number of adult flies emerged from fruits and fruit sweetness (Spearman’s rank correlation, R s = −0.05, P = 0.67) or fruit firmness (R s = −0.06, P = 0.60). The number of emerged flies was negatively correlated with pH, with higher numbers of adult D. suzukii emerged from species with more acidic fruit (R s = −0.38, P < 0.001). Numbers of emerged flies were also negative correlated with percentage reflectance across most of the visible spectrum (Table 2).
Discussion
Given the rapid expansion of D. suzukii across Asia, Europe, North America, and South America, significant attention has been focussed on risk to commercial fruit and berry crops. Studies addressing the potential attractiveness or host preference of D. suzukii have suggested fruit characters, including fruit sweetness, acidity, and firmness, as important factors (Lee et al. Reference Lee, Bruck, Curry, Edwards, Haviland, Van Steenwyk and Yorgey2011, Reference Lee, Dalton, Swoboda-Bhattarai, Bruck, Burrack and Strik2016; Burrack et al. Reference Burrack, Fernandez, Spivey and Kraus2013). Volatile profiles of ripening fruit and fruit-associated yeasts have also been identified as potentially important olfactory cues for host-finding by D. suzukii (Cha et al. Reference Cha, Adams, Rogg and Landolt2012, Reference Cha, Adams, Werle, Sampson, Adamczyk, Rogg and Landolt2014; Hamby et al. Reference Hamby, Hernández, Boundy-Mills and Zalom2012; Yu et al. Reference Yu, Zalom and Hamby2013; Abraham et al. Reference Abraham, Zhang, Angeli, Abubeker, Michel, Feng and Rodriguez-Saona2015; Keesey et al. Reference Keesey, Knaden and Hansson2015; Revadi et al. Reference Revadi, Vitagliano, Rossi Stacconi, Ramasamy, Mansourian and Carlin2015; Hamby and Becher Reference Hamby and Becher2016; Hickner et al. Reference Hickner, Rivaldi, Johnson, Siddappaji, Raster and Syed2016; Huang et al. Reference Huang, Gut and Grieshop2017; Karageorgi et al. Reference Karageorgi, Bräcker, Lebreton, Minervino, Cavey and Siju2017; Cloonan et al. Reference Cloonan, Abraham, Angeli, Syed and Rodriguez-Saona2018; Liu et al. Reference Liu, Dong, Zhang, Kenis, Griepink and Zhang2018). Changes in fruit profiles following damage due to injury or prior insect oviposition can alter attraction of D. suzukii (Yu et al. Reference Yu, Zalom and Hamby2013; Lasa et al. Reference Lasa, Tadeo, Dinorín, Lima and Williams2017; Wang et al. Reference Wang, Kaçar and Daane2019). Fruit phenology has also been identified as a risk factor (Sward et al. Reference Sward, Glass and Philips2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017; Elsensohn and Loeb Reference Elsensohn and Loeb2018). However, as D. suzukii continues to expand its geographic range, predicting which fruits are at risk is of increasing importance. We tested fruit susceptibility and preference among fruits at the northern range limit of D. suzukii.
In two-choice assays, numerical counts of D. suzukii were higher in Saskatoon berries, cloudberries, strawberries, raspberries, and bittersweet nightshade than in highbush blueberries and pin cherries. We note that in choice tests among these fruits, only Saskatoon berries had significantly higher mean retention-index values compared to the other fruits. This is consistent with previous findings that D. suzukii oviposits more eggs in raspberry than in many other commercially grown berries (Burrack et al. Reference Burrack, Fernandez, Spivey and Kraus2013). However, in two-choice assays of fruit versus a blank control, D. suzukii were attracted to all fruit species tested, and differences in attraction among fruit species were significant only between Saskatoon berry and highbush blueberry or pin cherry. This is consistent with previous findings that D. suzukii is highly opportunistic and makes use of less preferred hosts when options are limited (Diepenbrock et al. Reference Diepenbrock, Swoboda-Bhattarai and Burrack2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017).
In no-choice trials, eggs were observed in all fruit species except bittersweet nightshade. Lee et al. (Reference Lee, Dreves, Cave, Kawai, Isaacs and Miller2015) previously documented oviposition in this fruit but larvae did not develop. We observed eggs, larvae, and a single emergent adult in crowberry. Under laboratory conditions, crowberry fruit became increasingly desiccated. In a more natural environment, fruits attached to the plant may be more resistant to desiccation and therefore more suitable to larval development. For the purposes of this study, equal volumes of fruits were used for host preference two-choice assays and host suitability no-choice trials. This would not be the case in a field setting, where fruit size, proximity of fruits to each other, relative abundance of fruits, and foliage characteristics may play important roles in host fruit selection and/or suitability for larvae. Commercial berry crops are typically bred for large fruit size, high fruit abundance, and clustered fruit for ease of harvest. These characteristics, which are beneficial for commercial growers, also promote host fruit selection by pest insects.
Host selection by D. suzukii among fruits of a single species has previously been correlated with firmness, acidity, and sweetness in raspberries and blackberries but not grapes (Vitis Linnaeus; Vitaceae) (Burrack et al. Reference Burrack, Fernandez, Spivey and Kraus2013; Pelton et al. Reference Pelton, Gratton and Guédot2017). This study confirms that fruit firmness and acidity are factors in host selection among several host fruit species, but that fruit sweetness is less reliable as a host selection factor.
Fruit firmness or resistance to penetration is a limiting factor in host suitability among undamaged fruits for D. suzukii (Diepenbrock et al. Reference Diepenbrock, Swoboda-Bhattarai and Burrack2016; Lee et al. Reference Lee, Dalton, Swoboda-Bhattarai, Bruck, Burrack and Strik2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017). Oviposition has previously been correlated with fruit firmness, and we found that softer fruits were preferred in two-choice assays, but fruit firmness did not limit larval development (Lee et al. Reference Lee, Dalton, Swoboda-Bhattarai, Bruck, Burrack and Strik2016; Little et al. Reference Little, Chapman, Moreau and Hillier2017). Fruit sweetness was not a factor in either host selection or host suitability among the fruits tested. No more or fewer adult flies emerged relative to fruit sweetness levels. Among the fruits tested, more flies were attracted to and emerged from fruits with lower pH. This is contrary to results in previous studies, suggesting that D. suzukii may rely on a combination of factors for determining host selection and host suitability, even under controlled laboratory conditions (Lee et al. Reference Lee, Dalton, Swoboda-Bhattarai, Bruck, Burrack and Strik2016). Little is yet known of the effects of different larval host fruits on adult D. suzukii fecundity and longevity. However, previous studies have demonstrated that D. suzukii maternal life history, particularly maternal diet, can significantly influence the viability of their offspring regardless of the fruit host of the larvae (Plantamp et al. Reference Plantamp, Estragnat, Fellous, Desouhant and Gibert2017). Further research is needed to determine the role these fruits may play in the growth of local D. suzukii populations.
Within a fruit species or cultivar, fruit colour has been used as a proxy for assessing fruit ripeness (Li et al. Reference Li, Lee and Wang2014; Hu et al. Reference Hu, Zhai, Zhao and Wang2018; Little et al. Reference Little, Chapman and Hillier2018). These relationships are more pronounced within a single fruit species or variety but can also be useful across diverse fruit species. Across the fruit species tested, mid-length-wavelength reflectance values (cyan–orange (525–600 nm)) were characteristic of fruit sweetness, and reflectance across most of the visible spectrum was representative of fruit firmness. Fruit acidity was not correlated with reflectance.
Previous studies that investigated relationships between host choice and fruit colour used broad categories of colour based on what is perceptible to human vision, rather than Drosophila visual sensitivities (Lee et al. Reference Lee, Dreves, Cave, Kawai, Isaacs and Miller2015). This research builds on our previous study of the role of fruit colour and contrast between fruit and foliage in host selection of ripening blueberries (Little et al. Reference Little, Chapman and Hillier2018). We observed a negative correlation between fruit preference and reflectance across most of the visible spectrum. Adult emergence was also negatively correlated with reflectance. Thus, fruits that were more reflective were less preferred by D. suzukii and potentially less suitable hosts. This is the first study to explore relationships between host choice among different fruits and objective, quantified assessments of fruit colour, with recognition that the visual range of Drosophila differs from that of humans (Little et al. Reference Little, Chapman and Hillier2018).
This study is the first confirmation that Saskatoon berry, crowberry, and cloudberry are susceptible to infestation by D. suzukii. These fruits currently occur at the presumed northern range limit for D. suzukii; however, exposure of fruit to this invasive fly will most likely increase with continuing effects of climate change (Hamby et al. Reference Hamby, Bellamy, Chiu, Lee, Walton and Wiman2016; Rossi Stacconi et al. Reference Rossi Stacconi, Kaur, Mazzoni, Ometto, Grassi and Gottardello2016; Langille et al. Reference Langille, Arteca and Newman2017; Stockton et al. Reference Stockton, Wallingford and Loeb2018; Thistlewood et al. Reference Thistlewood, Gill, Beers, Shearer, Walsh and Rozema2018). There may also be sufficient plasticity in D. suzukii cold tolerance that northern populations may slowly become more cold adapted (Jakobs et al. Reference Jakobs, Gariepy and Sinclair2015; Stockton et al. Reference Stockton, Wallingford and Loeb2018).
Drosophila suzukii can assess relative risks and benefits of different fruit species and has demonstrated behavioural plasticity in host selection behaviour (Diepenbrock et al. Reference Diepenbrock, Swoboda-Bhattarai and Burrack2016; Sward et al. Reference Sward, Glass and Philips2016). This lack of host fidelity by D. suzukii may be key to its rapid near global expansion (Diepenbrock et al. Reference Diepenbrock, Swoboda-Bhattarai and Burrack2016). Host selection by D. suzukii is subject to numerous factors, including fruit characteristics, fruit and foliage volatile odours, fruit abundance, availability of other fruit species, competition with other insect species, and risk of predation. Fruit colour, particularly reflectance of short-length to mid-length-wavelength (blue–orange (470–600 nm)) light, provides D. suzukii with a reliable proxy for fruit quality and ripeness, regardless of fruit species, and may contribute to host selection. Growers of fruits at greatest risk from D. suzukii may benefit from selecting fruit varieties that retain high reflectance values, particularly at shorter wavelengths (470–600 nm), as the fruit ripens to reduce the attractiveness of those fruit crops to D. suzukii. Future research is needed to determine how fruit colour and host volatile odours interact in host selection by D. suzukii and to determine the importance of fruit colour relative to other host cues. Commercial and native fruits in boreal and even Nearctic regions are suitable hosts for D. suzukii and, with growing effects of climate change, are at increasing risk of infestation.
Acknowledgements
We are grateful for the assistance of Rebecca Rizzato, Aaron Spares, and Velma Dixon for collecting fruits for testing. We thank Heather Crozier for her assistance measuring fruit characters. We are grateful for the generosity of Nova Agri Group and MacBerry Farms for their continuing support and access to their blueberry fields. We gratefully acknowledge the financial support of Atlantic Canada Opportunities Agency Atlantic Innovation Fund (197853), Canada Foundation for Innovation (22087), Natural Sciences and Engineering Research Council of Canada (356109-2008 RGPIN and PGSD2-475743-2015), and Growing Forward 2 in Newfoundland and Labrador (GF21415-309, GF21516-519, GF21617-773, and GF21718-956).
Author contribution
C.L. and N.H. designed research. P.D. and T.C. advised on suitable fruits for testing and methodology. C.L. conducted research. C.L. analysed data and prepared manuscript. N.H., P.D., and T.C. edited manuscript. All authors read and approved manuscript.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.4039/tce.2020.1.







