

The insects examined are preserved in concretions or tabular bands of very fine-grained micrite, known as Insect Limestone. The unit where these concretions/bands occur is known as the Insect Bed, which lies towards the base of the Bembridge Marls Member (Solent Group: Bouldnor Formation). Most of the specimens at the Natural History Museum in London (NHMUK) belong to the A'Court Smith (purchased 1877, 1883), P. B. Brodie (purchased 1898) and R. W. Hooley (purchased 1924) collections. They are labelled ‘Gurnard Bay' or ‘Gurnet Bay' (which is an old name for Gurnard Bay). However, Smith collected specimens all the way from West Cowes to Newtown River on the NW side of the Isle of Wight (Jarzembowski Reference Jarzembowski1980). Most of the specimens probably came from Thorness Bay (Jarzembowski Reference Jarzembowski1976). Brodie and Hooley acquired parts of Smith's collection, so parts and counterparts of individual insects have turned up in all three collections. The parts and counterparts often have different numbers because they were registered at different times and sometimes deposited in different museums. An additional collection was discovered at the Sedgwick Museum, Cambridge, by A. J. Ross. This collection has also yielded counterparts of specimens at the NHMUK, which indicates that this is another part of the Smith collection. A label with ‘1883' on it suggests that the Sedgwick Museum acquired this collection in 1883, the same year that the NHMUK purchased specimens from Smith.
The age of the Bembridge Marls has been discussed in many publications, and they are currently considered to be latest Eocene (Priabonian) in age (see Ross & Self 2014). Many publications are devoted to the insect assemblages, some of them list several beetle names (Brodie Reference Brodie1878, Reference Brodie1894; Goss Reference Goss1878; Cockerell Reference Cockerell1921; Crowson Reference Crowson1962). For example, P. B. Brodie mentioned the following Coleoptera in 1878: Dytiscus (Dytiscidae), Staphylinus (Staphylinidae), Hydrophilus (Hydrophilidae), Dorcus (Lucanidae; now Pentodon dorcus sp. nov. in Scarabaeidae), Anobium (Anobiidae) and Curculio (Curculionidae), and he added ‘Telephoridae' and ‘Buprestidae' to these names in 1894. Nevertheless, in 1921 Cockerell described only one Carabid species (Pterostichus gurnetensis), and in 1962 Crowson published the description of one Cupedid species (Tetraphalerites oligocenicus), although by that time most specimens had already been deposited in museum collections.
The collections studied (NHMUK; Booth Museum, Brighton; Isle of Wight County Museum; Maidstone Museum and Art Gallery; Sedgwick Museum of Earth Sciences, Cambridge) contain many coleopterous remains from the Bembridge Marls, comprising nearly complete beetles or fragmentary remains. In total, 994 specimens have been identified as beetles. There is no conclusion on the systematic position of 414 of the specimens, about half of which have not been studied because of limited time for investigation. Among the determined remains there are 31 families present, although the whole number of coleopterous families in the Bembridge Marls should be expected to exceed at least 50. The overwhelming majority of remains available for study are isolated elytra, the precise identification of which is difficult because of similarity in elytra belonging to different groups. Some elytra have a rather characteristic structure and can be identified, but because of the huge diversity of the order it is difficult to do this precisely. Most difficulties arise in determining the family attribution of isolated elytra, and often below the family level they cannot be clarified without particular consideration and special study, as the characteristics of extant families in handbooks and manuals, as a rule, do not include a description of the elytra in sufficient detail.
The manuscript of this paper was prepared in 2007. Two of the authors, A. V. Alexeev and V. G. Gratshev, are now deceased. In 2018 the manuscript was updated with necessary, but minimum, corrections and additions, including important publications that have appeared since the submission of this manuscript.
AGK compiled the manuscript. Individual systematic sections were written by the following: Archostemata by AGP and AGK; Dytiscoidea, Caraboidea and Spercheidae by AGP; Hydrophilidae by AGP and CS; Hydraenidae, Pselaphidae, Throscidae, Eucnemidae, Byrrhoidea, Cantharoidea, Cleroidea, Latridiidae and Tenebrionoidea by AGK; Staphylinidae by AYS; Scarabaeoidea by FTK; Scirtoidea by AGK and AGP; Buprestidae by AVA, AGK and AGP; Elateridae by AVA; Nitidulidae, Cryptophagidae and Corylophidae by AGK and ASK; Chrysomeloidea by ASK and AGK; Curculionoidea by VG and AGP.
The complete lists of fossil taxa and references to them are represented in a Web catalogue on the site ‘Beetles (Coleoptera) and coleopterists' (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko2018), which also includes some unpublished information.
DEPOSITORIES:
BMB, Booth Museum of Natural History, Brighton.
CAMSM, Sedgwick Museum of Earth Sciences, University of Cambridge.
IWCMS, Isle of Wight County Museum Service.
NHMUK, Department of Earth Sciences, Natural History Museum, London.
1. Systematic palaeontology
Suborder Archostemata Kolbe, Reference Kolbe1908
Family Cupedidae Laporte, Reference Laporte1836

Figure 1 Beetles of families of Cupedidae sensu lato and Carabidae, Coleoptera incertae sedis. Tetraphalerites oligocenicus Crowson, Reference Crowson1962, holotype, NHMUK I. 8611: (a) dorsal view, part; length=9.9mm; (b) ventral view, counterpart; length=10.9mm; (c) Cupedinae sp.: NHMUK PI II 2717, elytral apex, dorsal view; length=1.8mm. (d) ?Calosoma (Callitropa) sp., NHMUK I. 25573, elytron, dorsal view; length=8.0mm. Neothanes europaeus Ponomarenko, sp. nov.: (e) holotype NHMUK In. 25679, elytron, dorsal view; length = 12.0 mm. (f) Paratype NHMUK In. 25580, elytron, dorsal view; length=12.0mm. (g) Pterostichus gurnetensis Cockerell, Reference Cockerell1921, holotype, NHMUK In. 24325' elytra, dorsal view; length of elytra 1.1, width=4.0mm.
Material. Two specimens (identified by R. A. Crowson, A. G. Kirejtshuk and A. G. Ponomarenko). NHMUK, I. 8611 (Fig. 1a, b), holotype of Tetraphalerites oligocenicus Crowson, Reference Crowson1962 and NHMUK PI II 2717 (Fig. 1c), N end of Thorness Bay, remains of apex of the left elytron apex of cupedine specimen, collected by A. G. Kirejtshuk and A. S. Kurochkin.
This family consists of usually elongate beetles of medium-sized body. This group is well recognisable in both the Recent fauna and fossils due to the characteristic elytral structure maintaining the primary membrane in longitudinal rows of cells and primary veins (with frequent fusion of long anal vein (A1) and cubitus anterior (CuA) at elytral apices), squamose vestiture of dorsum, more or less tuberculose dorsal surface of head and pronotum, procoxal cavities externally broadly open and rather narrow and long legs. The fossils of it can be identified even by the characters of separated elytra thanks to the peculiar ‘venation' and particular tubercles and scales around cells of elytra and other parts of body. There are more than 30 known species in the Recent fauna distributed through many areas, but outside Europe (and Western Palaearctics in general), mostly in South East Asia. Larvae of the Recent species are known to develop in dead wood of angiosperm trees only, and adults occur at the surface of tree trunks and branches and frequently visit flowers. The comparatively great number of Caenozoic fossils of this family (Spahr Reference Spahr1981; Tröster Reference Tröster1993; Kirejtshuk Reference Kirejtshuk2005a; Kirejtshuk et al. Reference Kirejtshuk, Nel and Colomb2010a, Reference Kirejtshuk, Nel and Kirejtshuk2016; etc.) indicates that this group had strongly decreased in abundance, even after the Palaeogene (fossil Cupedinae sensu stricto were recorded from the Miocene and even reached the Pliocene (Ponomarenko Reference Ponomarenko1973; Gersdorf Reference Gersdorf1976; Tröster Reference Tröster1993; Kirejtshuk et al. Reference Kirejtshuk, Nel and Kirejtshuk2016; etc.). In most large Eocene and Oligocene localities of Europe the reticulated beetles are represented mostly by the genus Cupes Fabricius, Reference Fabricius1801 sensu lato. Having appeared in fossil records in the Middle-Upper Triassic, the family Cupedidae is found from all periods of the Meso- and Caenozoic eras, although Triadocupedinae Ponomarenko, Reference Ponomarenko1966 do not reach the Caenozoic (Ponomarenko Reference Ponomarenko1969). A piece of limestone with one tip of a cupedine elytron was found in Thorness Bay as a result of the authors' fieldwork of 2005.
Note. The holotype of Tetraphalerites oligocenicus was re-examined and transferred from the tribe Ommatini to Brochocoleini, previously regarded as an extinct Mesozoic group (only up to Cenomanian) (Ponomarenko Reference Ponomarenko2006).
Suborder Adephaga
Superfamily Dytiscoidea Leach, Reference Leach1815
Family Dytiscidae Leach, Reference Leach1815

Figure 2 Beetles of the family Dytiscidae. (a) Agabus latissimus Ponomarenko, sp. nov.: holotype, NHMUK I. 10246, pterothorax and abdomen, ventral view; length = 6.0mm. Ilybius gratshevi Ponomarenko, sp. nov.: (b) holotype, NHMUK I. 9984, body, ventral view; length=7.7 mm; (c) paratype NHMUK I. 26124, elytron, dorsal view; length = 5.5mm. (d) ?Hygrotus sp.: NHMUK I. 9840, abdomen, ventral view and elytron, dorsal view; length = 2.0mm. Tribe ?Methlini or Vatellini, gen. & sp. indet.: (e) NHMUK I. 10293, body, ventral view (negative print); length = 2.0mm; (f) NHMUK I. 9346, elytra, dorsal view; length = 3.0mm. (g) ?Laccophilus sp.: NHMUK In. 25581, pterothorax and abdomen, ventral view; length = 2.2mm. (h) ?Oreodytes sp. or ?Hydrovatus sp.: NHMUK I. 17437, elytron, dorsal view; length = 2.9mm. (i) Dytiscid larva: NHMUK I. 10008, fragments of body, dorsal view; length of print = 6.0mm.

Figure 3 Suborder Adephaga, family Dytiscidae. (a) Holotype of Agabus latissimus Ponomarenko, sp. nov., NHMUK ‘I. 10246', pterothorax and abdominal base, ventral view. (b) Holotype of Ilybius gratshevi Ponomarenko, sp. nov., NHMUK ‘I. 9984', body, ventral view.
Material. A total of 18 specimens (identified by A. G. Ponomarenko, A. G. Kirejtshuk and A. A. Prokin), including remains of larvae.
Diving beetles, or Dytiscidae, are a rather diverse aquatic group with predaceous larvae and adults. The adults have various body sizes (1.5–45.0mm in length) and are characterised by a streamlined body, usually short head with widely arcuate anterior edge of frons, rather large and contiguous metacoxae meeting elytra, and long posterior legs with dilated and long-haired apical segments. Predaceous diving beetles are widespread in all zoogeographic regions, although the faunas in temperate and tropical climate are rather different. Both larvae and adults of Recent taxa usually live in stagnant fresh and brackish water basins, because they breathe air. Predaceous diving beetles appeared in the Jurassic, but during the Mesozoic and most part of the Palaeogene they were rare and not diverse. In the Oligocene their representation and diversity increased, and in the Miocene they became dominant in many sites with insect fossils. Only two extinct Caenozoic genera have been described from Upper Eocene Florissant (Miodytiscus Wickham, Reference Wickham1911) and Lower Miocene Rott (Palaeogyrinus Schlechtendal, Reference Schlechtendal1894). The diversity of predaceous diving beetles in the Bembridge Marls corresponds to other cases of the Oligocene situation or, probably, are somewhat higher. All remains are on the surface of the laminae. They present 5–7 species, including 1–2 species of medium size (with a length of 5–6mm, looking like Agabus Leach, Reference Leach1817), while the remainder consist of small beetles (with a length of 2.0–2.5mm and similar to Hydroporus Schellenberg, Reference Schellenberg1806 and Hygrotus Stephens, Reference Stephens1828). It is noteworthy that species of the genus Copelatus Erichson, Reference Erichson1832 are absent, although they are quite characteristic of the areas with tropical climate and usual in other sites with fossils (one species of this genus is known from Baltic amber). Besides adults, there are three large segments of a larva, more similar to predaceous diving beetles than to water scavenger beetles (Hydrophilidae). At the same time, no adult with such a body size has been found among the agabine specimens in the examined limestome. The composition of the family suggests moderate climatic circumstances of the past environment of this Palaeogene fauna, which consisted mainly of inhabitants of shallow water basins.
Subfamily Agabinae Thomson, Reference Thomson1867
Genus Agabus Leach, Reference Leach1817
Type species. Agabus paykullii Leach, Reference Leach1817, Recent.
These incomplete coleopterous remains are put into the genus Agabus because of the shape of metaventrite and metacoxae. Over 100 Recent and around ten fossil species have been described. Extant species of the genus Agabus are known from the uppermost Eocene of Florissant, USA, and numerous Neogene localities.
Agabus latissimus Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:7AA79AD3-CC25-48FF-983D-2395BC29B7C2
Derivation of name. The name latissimus means ‘widest'.
Materal. Holotype. NHMUK I. 10246 (Figs 2a, 3a): meso- and metathorax and base of abdomen; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species seems to be closest to Agabus rottensis Nilsson, Reference Nilsson2001 (=latus Statz, Reference Statz1939–1940) of Aquitanian Rott, Germany, differing from it in the wider body, wider wings of metaventrite and parallel-sided metacoxal plates. A somewhat similar beetle was figured, but not described by Schawaller (Reference Schawaller1986).
Description. Holotype I. 10246: length of elytra about 6.0mm, width 2.0mm, entire length of specimen about 9.0mm. Broad oval, lateral edges weakly curved between pronotum and the middle of abdomen. Metaventrite long, along the middle about as long as metacoxae, submetacoxal line nearly rectangular, wings of metaventrite wide, not linguiform. Length of metacoxa smaller than its width, metacoxal plates not pointed behind, their edge subparallel and tips rounded. Abdominal ventrite 2 longer than the first, ventrite 3 somewhat longer than second one. Metaventrite and metacoxae with coarse puncturation.
Probable ecology. Most Recent species of Agabus live in stagnant lakes with rich vegetation.
Genus Ilybius Erichson, Reference Erichson1832
Type species. Dytiscus fenestratus Fabricius, Reference Fabricius1781, Recent.
These incomplete coleopterous remains are regarded as the genus Ilybius because of the size and shape of body, which is widest behind the middle, and the very narrow wings of the metaventrite. Some isolated elytra are considered as the same species because they are the same size and have long longitudinal meshes of elytral reticulation. Some dozens of Recent species of the genus are known. No fossil species has been described before, although the genus was mentioned from Oeningen (Miocene) (Schöberlin Reference Schöberlin1888).
Ilybius gratshevi Ponomarenko, sp. nov.
(Figs 2b, c, 3b)
Zoobank ID. urn:lsid:zoobank.org:act:0774D13A-2A98-4BCC-8560-645E087B116A
Derivation of name. The name of the new species is devoted to the memory of V. G. Gratshev.
Material. Holotype. NHMUK I. 9984 (Figs 2b, 3b): beetle with missing antennae and legs, probably female; Insect Limestone, NW Isle of Wight, Brodie Collection.
Additional specimen. NHMUK In. 26124 (Fig. 2c): right isolated elytron; Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. The metaventrite of this new species is most similar to that of the Recent Ilybius fenestratus Fabricius, Reference Fabricius1781, but differs from the latter in the shape of body and colouration of elytra.
Description. Holotype I. 9984: length of body 7.7mm, length of elytra 5.5mm, width 1.8mm. Body elongate oviform, widest in posterior third, lateral edges regularly curved between head and apex of abdomen. Head subtriangular, at base twice as wide as long. Eyes drop-shaped and large. Prothorax four times as wide as long, its anterior angles weakly projecting. Procoxae small, rounded. Prosternal process sharply pointed and with a longitudinal median keel. Mesocoxae rounded and contigous. Metaventrite short, along the middle twice as long as diameter of mesocoxa, metacoxae twice as long as metaventrite, submetacoxal line nearly rectangular, wings of metaventrite narrow, linguiform. Metacoxa as long as wide, coxal plates not pointed behind, its edges subparallel, apices rounded. Abdominal ventrites 1 and 3 subequal in length, ventrites 4 and 5 shorter, ventrite 6 longest. Epipleura narrow, ending near base of abdomen.
Additional specimen. Length of elytra 5.5mm, width 1.8mm. Elytra elongate, three times as long as wide in the middle. Adsutural striae present. Disc of elytra in anterior third with reticulation of very long, narrow longitudinal meshes. There are two pale elongate stripes on each elytron: one is wider and disposed from shoulder to apex of the elytra, and another is along sutural edge in anterior two-thirds.
Probable ecology. Most Recent species of Ilybius live in stagnant lakes with rich vegetation. The beetle examined has rather narrow wings of the metaventrite and probably did not fly very often.
Subfamily Laccophilinae Gistel, Reference Gistel1856 &
Hydroporinae Aubé, Reference Aubé and Dejean1836
There are seven specimens that possibly belong to these subfamilies based on their body size, shape and colouration.
Material. NHMUK I. 9840 (Fig. 2d) is a small dytiscid with wide oval body and missing head, prothorax and legs; Insect Limestone, NW Isle of Wight, Brodie Collection. No details of meso- and metaventrite are visible. Length of elytra 2.0mm. Possible attribution of it could be Hygrotus Stephens, Reference Stephens1828.
NHMUK I. 10293 (Fig. 2e) is a small dytiscid with widely oviform body and missing head, prothorax and legs, Insect Limestone, NW Isle of Wight, Brodie Collection. No details of meso- and metaventrite are visible. Apex of abdomen is acuminate. Length of elytra 1.8mm. A possible attribution of it could be Methlini Branden, Reference Branden1885 or Vatellini Sharp, Reference Sharp1882. Another small dytiscid with similar shape of body without visible details (I. 9346 – Fig. 2f) possibly belongs to the same species.
NHMUK In. 25581 (1) (Fig. 2g) is a small dytiscid with wide oviform body and with missing head, prothorax and legs; Insect Limestone, NW Isle of Wight, Hooley Collection. Metatergum is well visible, but the details of the underside are invisible. Elytra with wide dark spots arranged in transverse bands. Length of elytra 2.3mm. It is possible that it should belong to the genus Laccophilus Leach, 1915.
Gen. & sp. indet.
(Fig. 2i)
Material. Dytiscid larva NHMUK I. 10008: Insect Limestone, Isle of Wight, Brodie Collection; length of segments about 1.2mm, width about 2.6mm. Total length of larva could be about 10–12mm. It most probably belongs to a species of Agabinae. A somewhat similar dytiscid larva was described from the lowermost Miocene (or Upper Oligocene) of Rott, Germany, by G. Statz (Reference Statz1939–1940) as ‘Dytiscites sp.'
Superfamily Caraboidea Latreille, Reference Latreille1802
Family Carabidae Latreille, Reference Latreille1802
Material. A total of 28 specimens (identified by T. D. A. Cockerell, A. G. Ponomarenko and A. G. Kirejtshuk), including holotype of Pterostichus gurnetensis Cockerell, Reference Cockerell1921.
Carabidae or ground beetles present one of the largest and most diverse families in the Recent fauna. They are characterised by a wide range of body sizes, usually elongate head narrowing to truncate to emarginate (or excised) anterior edge of frons, moderately short and narrowly separated metacoxae not meeting elytra, long posterior legs with narrow and simple tarsomeres. Recent ground beetles are mostly predators and are distributed worldwide. Some Recent groups of the family are mycetophagous or phytophagous and usually these groups show some corresponding structural adaptations. Ground beetles are quite a diverse coleopterous group in the Bembridge Marls as well. According to the examination by S. Kuzmina, the specimen labelled as Pterostichus gurnetensis (NHMUK In. 14325, Hooley Collection; Fig. 1g) indeed belongs to the genus Pterostichus Bonelli, Reference Bonelli1810. Five isolated elytra seem to belong to 2–3 species of the genus Calosoma Weber, Reference Weber1801. The latter is recorded from the Lower Oligocene onwards and represented by 20 fossil species. One elongate elytron with 12 rows of rather large pits and sharp costae between them is rather similar to those in representatives of the subgenus Callitropa Motschulsky, Reference Motschulsky1865 (NHMUK I. 25573; Fig. 1d). Two elytra with 18 rows of punctures differ from most species of the genus in the lack of rows with larger pits (foveae). In addition to the Calosoma species, Neothanes Scudder, Reference Scudder1890 from Middle Eocene Green River (USA) differs from them in the higher number of grooves (sulci) with punctures (numbering up to 25), but without clear foveae. However, in the elytra under consideration, such grooves are not so numerous. Nevertheless, a possibility to regard them as the elytra of Calosoma still exists. Two other elytra have rather bad preservation and details of their structure are not accessible for study. The remaining fragments comprise isolated elytra or incomplete bodies.
Subfamily Carabinae Latreille, Reference Latreille1802
Genus Neothanes Scudder, Reference Scudder1890
Type species. Cychrus testaceus Scudder, Reference Scudder1878, from the Middle Eocene of Green River, USA.
Two isolated elytra are put in to the genus Neothanes because of their shape and size, numerous punctured striae and absence of foveae. Ataktosites palustris Haupt, Reference Haupt1956 seems to be similar, but the latter is much smaller. The isolated elytra are not enough to decide the systematic position of these beetles. They could belong to species of Cychrus Fabricius, Reference Fabricius1794, Calosoma or Carabus Linnaeus, Reference Linnaeus1758. This type of elytral striae multiplication arose in many different families of beetles (see Omophron Latrielle, Reference Latreille1802 in Carabidae and Laccobius Erichson, Reference Erichson1837 in Hydrophilidae).
Neothanes europaeus Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:C2AB4A2F-8A4B-4254-855B-F865ECBF490C
Derivation of name. The name of this new species is formed from “Europa”.
Material. Holotype. NHMUK In. 25679 (Fig. 1e): left isolated elytron; Insect Limestone, NW Isle of Wight, Hooley Collection.
Paratype. NHMUK In. 25580 (Fig. 1f): counterpart of right isolated elytron; Insect Limestone, Isle of Wight, Hooley Collection.
Diagnosis. This new fossil species differs from the type species of the genus in the smaller number of longitudinal rows of punctures (18 instead of 25) and larger punctures in the row.
Description. Holotype In. 25679: length of elytra about 12.0mm, width 4.0mm. Surface of elytra with at least 18 regular longitudinal rows of round punctures. Punctures and distance between them subequal. Six rows located along sutural margin to apical part of outer edge of elytron. Four rows along outer edge reaching the end of the first of sutural groups of rows. Four next rows along the sutural margin terminate at the last rows in the outer group. The last four rows terminate at the first row in the previous group. Epipleural border very short.
Probable ecology. According to the structures characteristic of the genus, the new fossil species could be, like its close relatives, a predator of terrestrial snails.
Suborder Polyphaga
Infraorder Staphyliniformia Lameere, Reference Lameere1900
Superfamily Hydrophiloidea Latreille, Reference Latreille1802

Figure 4 Beetles of families Spercheidae and Hydrophylidae. Spercheus wightensis Ponomarenko, sp. nov.: (a) holotype, NHMUK In. 25625, elytron, dorsal view; length = 5.0mm; (b) paratype, NHMUK In. 25621, elytron, dorsal view; length = 5.1mm; (c) paratype, NHMUK ‘I. 8613', elytron, dorsal view; length = 5.6mm. (d) Spercheus punctatus Ponomarenko, sp. nov.: holotype, NHMUK In. 26104, elytron, dorsal view; length = 3.1mm. Hydrochara woodwardi Ponomarenko, sp. nov.: holotype NHMUK In. 17424: (e) pterothorax and abdomen, ventral view, part; (f) elytra, dorsal view, counterpart; length = 18.5mm. Berosus barclayi Ponomarenko, sp. nov.: (g) holotype, NHMUK In. 25659, elytron, dorsal view; length = 2.9mm; (i) paratype, BMB 024613, body, ventral view; length = 4.6mm; (j) paratype, NHMUK In. 26128, elytron, dorsal view; length = 2.5mm; (k) paratype, NHMUK ‘In. 25608', elytron, dorsal view; length = 3.0mm. (h) Berosus sp.: NHMUK I. 10180, pterothorax and abdomen, ventral view; length = 3.4mm. (l) Larva Berosus barclayi Ponomarenko, sp. nov.:, NHMUK I. 10199, dorsal view; length = 5.8mm.
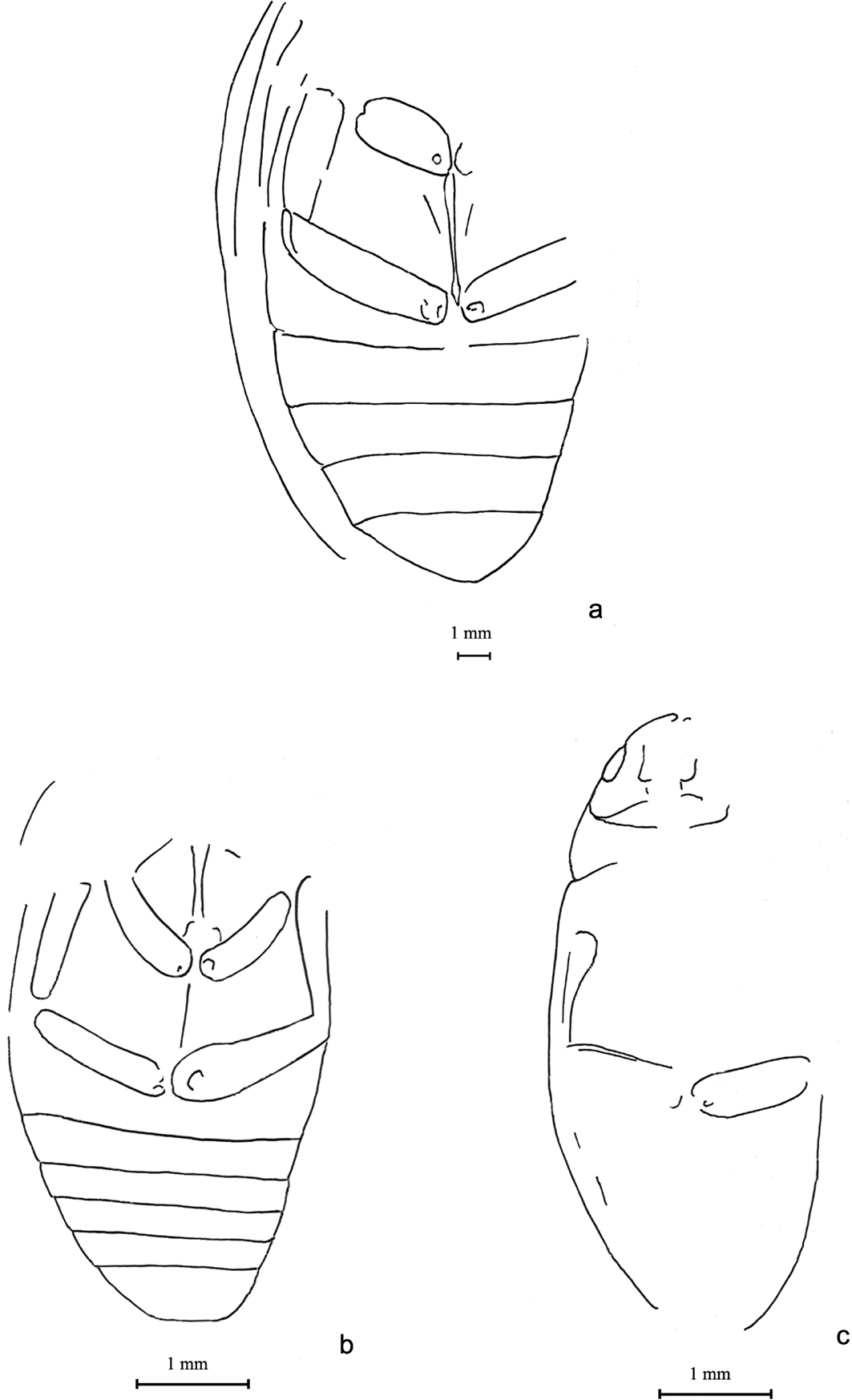
Figure 5 Suborder Polyphaga, family Hydrophilidae. (a) Holotype of Hydrochara woodwardi Ponomarenko, sp. nov., NHMUK I. 17424, pterothorax and abdomen, ventral view. (b) Berosus sp., NHMUK I. 11080, pterothorax and abdomen, ventral view. (c) Paratype of B. barclayi Ponomarenko, sp. nov., BMB 024613, body, ventral view.
Material. A total of 77 specimens (identified by A. G. Ponomarenko and A. G. Kirejtshuk).
Water scavenger beetles or Hydrophiloidea are represented in the Bembridge Marls by two families: Spercheidae and Hydrophilidae. Spercheidae is recorded as fossil for the first time. Hydrophilidae is one of the most diverse families of fossil beetles.
Family Spercheidae Erichson, Reference Erichson1837
Material. Five specimens (identified by A. G. Ponomarenko).
Several isolated elytra are put into the genus Spercheus Kugelann, Reference Kugelann1798 of the monotypic family Spercheidae because of the shape and size of body, as well as the shape of elytra with keel-like longitudinal ridges and large punctures between them. The genus was unknown in fossils before this study. Isolated elytra are not enough for an exact decision on the systematic position of the beetles; they resemble some species from Hydrochus Leach, Reference Leach1817, Epimetopus Lacordaire, Reference Lacordaire1854 and Georissus Latreille, Reference Latreille1809, but elytra of the first are more elongate, those of the second have rows of very large punctures between ridges and those of the third have ridges of another type.
Genus Spercheus Kugelann, Reference Kugelann1798
Type species. Dytiscus emarginatus Schaller, Reference Schaller1783, Recent.
Description of elytra. Elytra convex and wide. Length of elytra about one- and two-thirds as long as wide. Disc of elytra with four ridges and 2–4 rows of punctures between them. Many species have horn-like structures in apical part of ridges. Scutellary striola present. Elytral apices without interlocking tongue. Epipleura complete, not or gradually narrowed. Lateral margin arcuate.
Distribution. More than ten species are known in the Recent fauna, mostly from Africa, several species from South East Asia and Australia, and one species from both Europe and South America. There is usually no more than one species in one area.
Probable ecology. Recent species of Spercheus are phytophagous and live in stagnant, muddy or swampy freshwater bodies, most commonly on roots of water plants. Adults are often found on surface films of water, feeding on algae. Larvae are predaceous. The ecology of the new fossil species was probably similar.
Spercheus wightensis Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:C4F280C1-0ABC-4C3C-BDF2-B821940E9BCC
Derivation of name. This new species is named after the Isle of Wight.
Material. Holotype. NHMUK In. 25625 (Fig. 4a): left isolated elytron; Insect Limestone, NW Isle of Wight, Hooley Collection.
Paratypes. NHMUK I. 8613 (Fig. 4c), Brodie Collection; In. 25621 (Fig. 4b), In. 25634, Hooley Collection: isolated elytra; Insect Limestone, NW Isle of Wight.
Diagnosis. This new fossil species is closest to the African Spercheus crenulatus Fairmaire, Reference Fairmaire1897 owing to the flat platform, instead of horns on ridges, but differs from it in the very small punctures and filiform dark stripes between ridges.
Description. Holotype In. 25625: length of elytra about 4.5–5.1mm, width 2.1–2.3mm. Ratio of elytral length to width about 2.2:1. Disc of elytra with three flattened ridges, with small flattened plates in apical part of elytra. Ridges not joined in apical part of elytra, but the adsutural ridge bending to next one just before end. Fourth ridge at outer part of elytra and one short ridge before sutural striola weak. Short ridges between outer ridges absent. There are four rows of small, round, dark punctures between ridges and one pair of filiform dark stripes between rows of punctures. Epipleura gradually narrowed.
Spercheus punctatus Ponomarenko, sp. nov.
(Fig. 4d)
Zoobank ID. urn:lsid:zoobank.org:act:41CE6562-C52B-4DA7-BBC0-CF45D2AE79F7
Derivation of name. The name of this new species refers to punctures on its elytra.
Material. Holotype NHMUK In. 26104 (Fig. 4d): left isolated elytra; Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. This new fossil species seems to be closest to the African Spercheus senegalensis Laporte, Reference Laporte1832 due to the characteristic flat platform, though it has horns on ridges (a short ridge between two outer ridges); it differs from the latter species in having only four rows of punctures between ridges.
Description. Holotype In. 26104: length of elytra about 3.1mm, width 1.5mm. Ratio of elytral length to width about 2.1:1. Disc of elytra with three flattened ridges, with small flattened plates in apical part of elytra. Middle pair of ridges joined in apical part of elytra and, after that, with the adsutural one just before end. Outer ridge in outside part of elytra not weak. Short ridges present between outer ridges. There are four rows of punctures between ridges. Epipleura gradually narrowed.
Family Hydrophilidae Latreille, Reference Latreille1802
Material. A total of 72 specimens (identified by A. G. Ponomarenko and A. G. Kirejtshuk).
Water scavenger beetles or Hydrophilidae are, at present, the most diverse family of water beetles. Recent species live in different types of water basins and in wet terrestrial localities, mainly in stagnant and slow waters. Their larvae are predaceous, while the adults usually feed on decaying or, more rarely, live water plants. Larvae have gills and prefer to live in fresh water because of osmosis problems in saline water. They have a subcosmopolitan range, many genera have worldwide distribution, but they are more diverse and numerous in warm areas. Water scavenger beetles were described as fossils from the Jurassic (Geinitz Reference Geinitz1894; Bode Reference Bode1953; Ponomarenko Reference Ponomarenko and Rasnitsyn1985a, Reference Ponomarenko1985b, Reference Ponomarenko1987) and beetles of modern appearance started to come in deposits from the Lower Cretaceous (Ponomarenko Reference Ponomarenko, Arnoldi, Zherikhin, Nikritin and Ponomarenko1977, 1986; etc.). More than 20 genera of water scavenger beetles have been recorded from the Caenozoic, including two extinct genera (although the state of the latter is still doubtful). In the Palaeogene and Neogene, there is a difference in composition of predominant groups. In the Bembridge Marls, water scavenger beetles are numerous but with a small diversity (more than ten species). Some of the remains examined not infrequently belong to the same species. Besides the remains of adults, there are seven remains of larvae. All remains are on the surface of the laminae. Among adults, species of Berosus Leach, Reference Leach1817 (B. barclayi sp. nov. and B. sp.) and Hydrochara Berthold, Reference Berthol1827 (H. westwoodi sp. nov.) were established. Two small species, each with a body size of about 2.5mm, were found (each represented by only one specimen: NHMUK In. 25581 (2) and PI II 2842) from the subfamily Hydrophilinae and also ten separate elytra of the Hydrobiites-type (two or three species according to body size). The larvae examined seem to be conspecific with the most common beetle species of Berosus. Among the observed dominants, the genus Berosus is equally characteristic of both the Palaeogene and Neogene; however, the genus Hydrochara has been recorded mainly from the Neogene (Oligocene and Miocene deposits: Aix, Rott, most numerous in Oeningen, and others). In the materials examined, there are no characteristic tropical species. Thus, the Palaeogene fauna under consideration represents dwellers of shallow water basins.
Subfamily Hydrophilinae Latreille, Reference Latreille1802
Genus Hydrochara Berthold, Reference Berthol1827
Type species. Dytiscus caraboides Linnaeus, Reference Linnaeus1758, Recent.
Several fragmentary beetle remains can be put into the extant genus Hydrochara due to their short metaventral keel and correspondent body size. This genus was known as Hydrophilus for a long time and fossil representives were described under this name. Recent species of Hydrochara can be discriminated by body size, but many fossil isolated elytra with intermediate size are known. More than 20 extinct species were described during the 19th Century, but the descriptions are not informative enough for the determination of specimens; some of them can be treated as members of Hydrochara, Hydrophilus or belonging to a new genera. A further revision of the genus is needed.
Distribution. About ten Recent species in all continents, except Australia, are known. The genus is one of the most common Caenozoic fossil beetles, especially for the Miocene. The most ancient Hydrochara from Middle Eocene Green River, USA, remain still undescribed. This genus was mentioned in the ‘Insect Bed' by Woodward (Reference Woodward1879) as Hydrophilus.
Probable ecology. Adults of species of this genus are omnivorous water beetles, and larvae are predaceous.
Hydrochara woodwardi Ponomarenko & Soriano, sp. nov.
(Figs 4e, f, 5a)
Zoobank ID. urn:lsid:zoobank.org:act:EDF6CD65-27BA-4B1C-A55F-FDEB2ADBA0C4
Derivation of name. This new species is named after H. Woodward.
Material. Holotype NHMUK In. 17424 (Figs 4e, f, 5): beetle without head, prothorax and legs; Insect Limestone, NW Isle of Wight, A'Court Smith Collection.
Paratypes. NHMUK I. 8624, I. 9895, In. 25558 and counterpart In 25599, In. 26111, In. 26122: isolated elytra; Insect Limestone, NW Isle of Wight, Brodie and Hooley Collections.
Diagnosis. This new species is one of the largest and narrowest beetles among Recent and extinct members of Hydrochara and differs in the shape of the body, which is wedge-like and narrowed posteriorly. It seems to be close to ‘Hydrophilus' antiquus Oustalet, Reference Oustalet1874 from Aix, France (Oligocene), because of the body shape; however, it is not as large and has weaker elytral striae.
Description. Holotype In. 17424: length of elytra about 15–20mm, width 7–9mm. Ratio of elytral length to width about 2:1. Middle and metacoxae large, oblique transverse. Metepisterna narrow. Metasternal keel reaching the middle of metacoxa. Abdominal ventrite 2 widest, twice as wide as base of hypopygidium. Elytra convex and narrow. Length of elytra about 1.75 as long as wide. Disc of elytra with ten rows of punctures. Scutellary striola long. Elytra confluent at the apex. Elytral apices without interlocking tongue. Epipleura absent or incomplete. Lateral edge weakly convex. All fossil species of the genus have elytral apices untapered and without denticles. Elytra widest before middle. Disc of elytra with ten very weak striae, two nearest to suture more distinct. Epipleura not so wide and gradually narrowed.
Variations. Elytra vary in size and shape. Paratype I. 8624: Brodie Collection: length of print 18.0mm, width 7.0mm. Body outline somewhat wider than the holotype; discal rows of punctures are very weak.
Paratype I. 9895: Brodie Collection; length of elytra 19.0mm, width 6.0mm. Outline narrower than the holotype; discal rows of punctures are more evident.
Paratype In. 25558: Hooley Collection; length of elytra 18.0mm, width 9.0mm. Body outline somewhat wider than the holotype; discal rows of punctures are more evident. Counterpart In. 25599: Hooley Collection; body outline wider than the holotype; discal rows of punctures are more evident.
Paratype In. 26111: Hooley Collection; length of elytra 18.5mm, width 6.0mm. Body outline more narrow than the holotype; discal rows of punctures are evident.
Paratype In. 26122: Hooley Collection; length of elytra 19.0mm, width 8.0mm. Body outline like that of the holotype; discal rows of punctures are more evident.
Additional material. There are several smaller elytra, which may be small specimens of the same species or similar ones, detailed as follows.
NHMUK I. 8610: Insect Limestone, NW Isle of Wight, Brodie Collection; length of print near 15.0mm, width 5.8mm. Body outline somewhat wider, disc of elytron with rows of sparse punctures.
NHMUK In. 26102: Insect Limestone, NW Isle of Wight, Hooley Collection; length of print 10.0mm, width 4.3mm. Elytral outline wider, gradually narrowed in the apical half. Elytral disc with eight evidently punctured striae not joining the apex of elytron. External part of elytra flattened. Epipleura wide, with large, rounded punctures on its medial part.
NHMUK In. 26146: Insect Limestone, Isle of Wight, Hooley Collection; length of print 15.0mm, width 7.0mm. Body outline wider; discal rows of punctures are more clear.
Genus Berosus Leach, Reference Leach1817
Type species. Dytiscus luridus Linnaeus, Reference Linnaeus1760, Recent.
Many isolated elytra, two complete beetles and several larval remains can be put into the extant genus Berosus because of the body structures, puncturation and colouration of elytra, and abdominal gills of larvae. They are regarded as members of the subgenus Berosus sensu stricto because of the lack of denticles at their elytral apices.
Distribution. More than 100 extant species in all continents, except Antarctica. Four Caenozoic species, including the two most ancient species from Green River, USA (Middle Eocene), can scarcely belong to this genus. Other two species are known from Rott, Germany (Aquitanian).
Probable ecology. Adults are omnivorous water beetles, their larvae are predaceous.
Berosus barclayi Ponomarenko & Soriano, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:9221898B-7106-4056-B494-5BE7208ECFCE
Derivation of name. This new species is named after the coleopterist from the NHMUK Maxwell V. L. Barclay.
Material. Holotype NHMUK In. 25659 (Fig. 4g): isolated elytron; Insect Limestone, Isle of Wight, Hooley Collection. The specimen is selected as holotype, because all other specimens with body structures examined do not have well-preserved elytra and the comparison of them with other numerous isolated elytra is not possible.
Paratypes. Adult: NHMUK I. 10180, I. 10323, beetles with missing head, prothorax and legs; In. 17426, In. 25529, In. 25608 (Fig. 4k), In. 25634 (2), In. 26128 (Fig. 4j), isolated elytra. Larvae: I. 8478, I. 8486, I. 8488, I. 9361, I. 10020, I. 10199 (1) (Fig. 4l); Insect Limestone, NW Isle of Wight, Brodie and Hooley Collections; BMB, 024613 (Figs 4i, 5c), beetle without legs, ventral side only; Insect Limestone, NW Isle of Wight.
Diagnosis. This new species seems to be regarded as closest to Berosus morticinus Heyden & Heyden, Reference Heyden and Heyden1866 from Rott, Germany (Aquitanian) because of similarity in the shape of head and prothorax, dark elytral striae and a pair of dark spots at lateral part of elytra, but the new fossil species is much smaller, with the head shorter and punctures on elytral striae are dark.
Description. Adult: length of body about 4.0–4.5mm, length of elytra 3–4mm, width 1.5–2.0mm. Ratio of elytral length to their width about 2:1. Body small, convex, long oviform, narrower anteriorly, widest at base of abdomen. Head wider than long, triangular before eyes. Prothorax short, about twice as wide as long. Mesoventrite with one longitudinal keel. Mesocoxae large, oblique, metacoxae transverse. Metepisterna narrow. Metaventrite about twice longer at sides than in the middle. Abdomen with six ventrites, ventrite 1 with one longitudinal keel, ventrite 2 widest, about twice as wide at base as hypopygidium. Scutellum long and tapered. Elytra convex, confluent at apices. Disc of elytra with ten punctured furrows and with one row of small punctures between them. Scutellary striola long, falling into the previous one. Three lateral striae reaching apices of elytra, two next ones conjoining, others coming to the distal third along sutural edge and parallel. Epipleura absent or very narrow. Lateral edge weak, sutural edge very convex. Apices of elytra without denticles. Six internal elytral striae with dark stripes and their punctures dark as well. Two irregular dark spots along lateral edge and one along sutural edge.
Larvae. All specimens examined are in a bad condition: they seemed to be decayed and dessicated before fossilisation. Length of body 5.8–10mm. They are more slender than Recent Berosus larvae. Structure of head including characteristic epistome is invisible, legs short and first–seventh segments of abdomen bear filamental gills.
Variations. Paratype MHN I.10323: body length (without head) about 4.5mm, width 3.0mm; elytron length 4.3mm. Body elongate oval, widest behind the middle. Mesocoxae contiguous, about 2.5 times as wide as long. Metathorax short, more than three times as wide as long. Metacoxae very oblique. Abdominal ventrite 1 longest, abdomen gradually narrowed behind ventrite 2, hypopygidium about 1.3 times longer than penultimate ventrite.
Paratype BMB O24613: body length (with head) 4.6mm; body width 2.0mm; elytral length 3.0mm. Body elongate oval, widest at base of abdomen. Length of head 1.6mm, shorter than wide. Metaventrite short, more than three times as wide as long. Metacoxae transverse. Abdominal ventrite 1 longest, abdomen gradually narrowed from base. Elytra about 1.5 times as long as wide, epipleura narrow.
Isolated elytra. Holotype NHMUK In. 25659: length 2.9mm, width 1.2mm. There are six main striae with narrow dark stripes containing dark punctures, and punctures in intermediate rows also dark; two unclear infuscations located along lateral edge.
Paratype NMH In. 17426: length 3.0mm, width 1.4mm. Elytra with deep striae in sutural part and narrow dark stripes containing dark and deep punctures. There is one irregular infuscation along sutural edge and another at base along lateral edge.
Paratype NMH In. 25529: length 4.0mm, width 2.0mm. Elytra with deep striae along suture containing small and dark punctures. There is one large irregular infuscation along sutural edge and several small ones on disk.
Paratype NMH In. 25608: length 2.9mm, width 1.2mm. Elytral striae weak and with narrow dark stripes containing dark punctures. There are two irregular infuscations along lateral edge, one at base and another along sutural edge.
Paratype NMH In. 26128 (Fig. 4j): length 3.0mm, width 1.4mm. Elytral striae weak and with narrow dark stripes containing dark and small punctures. Irregular infuscations weak.
Berosus sp.
Material. NHMUK I. 10180 (Figs 4h, 5b): beetle lacking part of head and elytra; Insect Limestone, Isle of Wight, Brodie Collection. Body length 8.2mm, total body width 4.0mm, elytral length 4.2mm. Body elongate oval, widest at the middle of elytra. Prothorax about 2.3 times as wide as long, procoxae small and rounded. Procoxal process nearly a third of procoxal width. Mesocoxae projecting externally, contiguous. Metathorax short, more than three times as wide as long. Metacoxae slightly longer than procoxae. Abdominal ventrite 1 about 1.4 times as long as each of the other ventrites, abdomen gradually narrowed behind ventrite 2. Elytra about 2.8 times as long as wide combined, epipleura narrow. Disk of elytra with dark punctured striae and without denticles at apices. This specimen can be included in the genus Berosus because of the characteristic structures, puncturation and colouration of apical part of elytra. It is regarded as a member of the subgenus Berosus sensu stricto because of the lack of denticles at elytral apices. This specimen cannot be included in the previously described species Berosus barclayi sp. nov., as its body is about twice the size.
Superfamily Staphylinoidea Latreille, Reference Latreille1802
Family Hydraenidae Mulsant, Reference Mulsant1844

Figure 6 Beetles of family Hydraenidae. (a) Ochthebius (subgenus incertus) rossi Kirejtshuk, sp. nov.: holotype, NHMUK In. 43411, body, ventral view; length of print = 1.6mm. (b) Hydraenites gracilimmus Kirejtshuk, gen. & sp. nov.: holotype, NHMUK I. 10189, body, ventral view; length of print = 2.6mm. (c) Metacoxites ventritalis Kirejtshuk, gen. & sp. nov.: holotype, NHMUK In. 26131, body, ventral view; length of print = 1.4mm. (d) Davidraenites gratshevi Kirejtshuk, gen. & sp. nov.: holotype, NHMUK I. 10171, body, ventral view; length of print = 1.6mm. (e) Davidraenites interruptus Kirejtshuk, gen. & sp. nov.: holotype, NHMUK I. 10014, body, ventral view; length of print = 1.4mm. (f) Davidraenites spurcus Kirejtshuk, gen. & sp. nov.: holotype, IWCMS 2018.41.1, body, ventral view; length of print = 1.5mm.

Figure 7 Beetles of families Hydraenidae, ? Hydraenidae, Staphylinidae (Pselaphinae). (a) Hydraenidae, gen. & sp. indet.: IWCMS 2018.41.2, pterothorax and abdomen, ventral view; length of specimen = 1.4mm. (b) Eolimnebius fossilis Kirejtshuk, gen. & sp. nov.: holotype, NHMUK I. 8592, body, ventral view; length of specimen = 1.4mm. (c) Pselaphinae, gen. & sp. indet.: CAMSM X.50140.206, metaventrite and abdomen, ventral view; length of specimen = 1.6mm.

Figure 8 Suborder Polyphaga, family Hydraenidae. (a) Holotype of Ochthebius (subgenus incertus) rossi Kirejtshuk, sp. nov., NHMUK In. 43411, body, ventral view. (b) Holotype of Hydraenites gracilimmus Kirejtshuk, gen. & sp. nov., NHMUK I. 10189, body, ventral view. (c) Holotype of Metacoxites ventritalis Kirejtshuk, gen. & sp. nov., NHMUK In. 26131, body, ventral view. (d) Additional specimen of M. ventritalis Kirejtshuk, gen. & sp. nov., NHMUK In. 26147, body, ventral view.
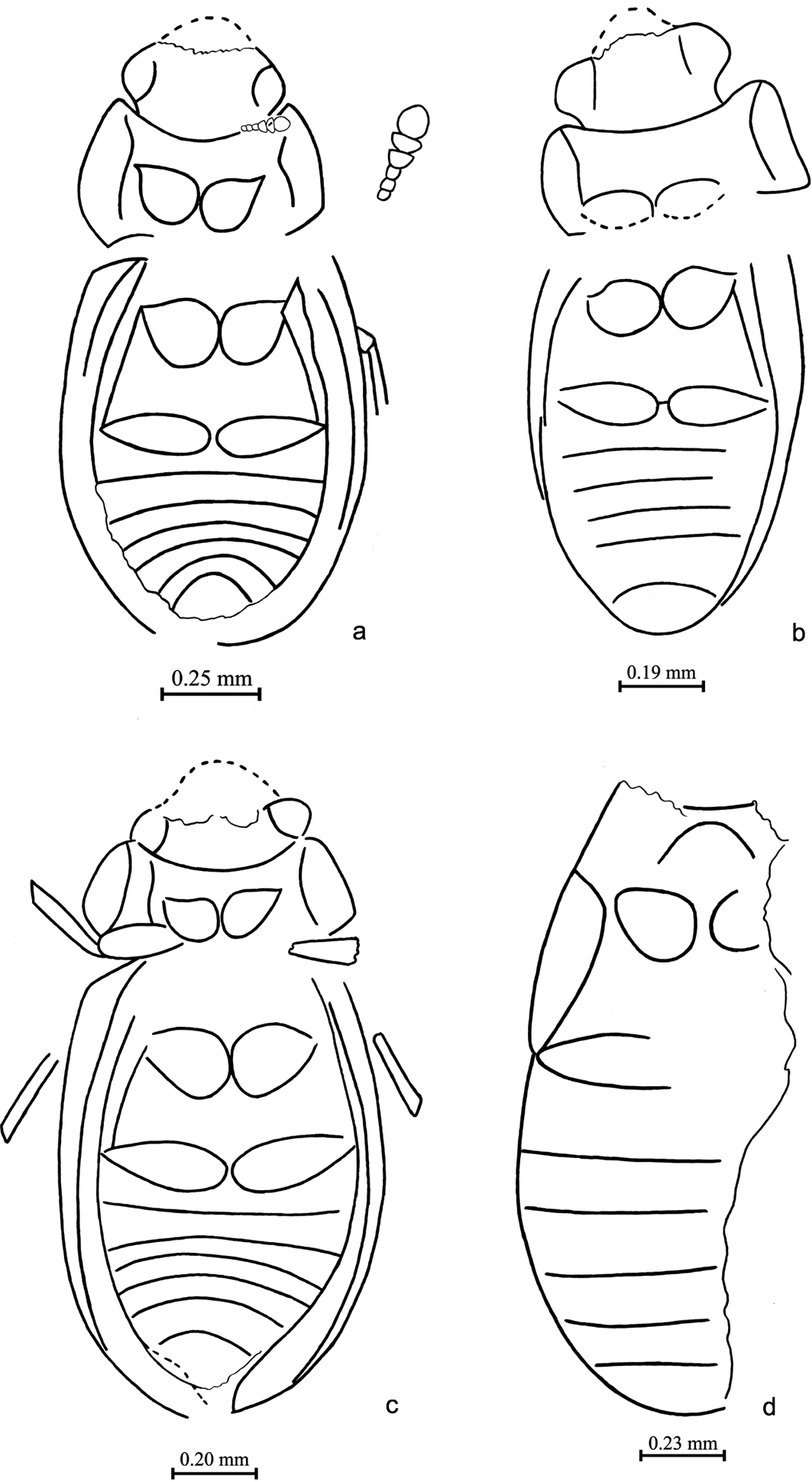
Figure 9 Suborder Polyphaga, family Hydraenidae. (a) Holotype of Davidraenites gratshevi Kirejtshuk, gen. & sp. nov., body, ventral view, NHMUK I. 10171. (b) Holotype of D. interruptus Kirejtshuk, gen. & sp. nov., body, ventral view, NHMUK I. 10014. (c) Holotype of D. spurcus Kirejtshuk, gen. & sp. nov., IWCMS 2018.41.1, body, ventral view. (d) Hydraenidae, gen. & sp. Indet., IWCMS 2018.41.2, body, ventral view.

Figure 10 Suborder Polyphaga, ? Hydraenidae: holotype of Eolimnebius fossilis Kirejtshuk, gen. & sp. nov., NHMUK I. 8592, body, ventral view.
Material. A total of 18 specimens (identified by A. G. Kirejtshuk).
This family represents a not very numerous group of rather small beetles (1.0–3.2mm), usually more or less elongate and slightly or moderately convex dorsally. They have a quite peculiar combination of characters, including eight nine-segmented antennae with a club, very long maxillar palpi, a prothoracic segment with the hyaline cuticula along sides and base, a rather long metaventrite and an abdomen with 6–7 ventrites. Adults of most Recent species have well-developed aquatic adaptations, including hydrofuge pubescence and hydrophilid-like antennae involved in the maintenance of an air bubble. The Recent species are distributed worldwide, except for pre-polar areas. Both larvae and adults occur in aquatic, semiaquatic, riparian and littoral habitats, mostly at shallow depth or at the water edge; both are considered to feed on algae. This family can be recognised in fossils due to the combination of the small body, structure of coxae and 6–7 abdominal ventrites. The earliest records of it with an unclear subfamiliar attribution are from Novospasskoye (Middle–Upper Jurassic), and it was also recorded from the Karatau shales (Upper Jurassic), Manlay (Lower Cretaceous) (Ponomarenko Reference Ponomarenko, Arnoldi, Zherikhin, Nikritin and Ponomarenko1977), Albian-Cenomanian Burmese amber (Jäch & Yamamoto Reference Jäch and Yamamoto2017) and others (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko2018). In the Caenozoic, representatives of Hydraeninae are known from Castle Eden (Pliocene: Lesne Reference Lesne1920), and those of Ochtebiinae Thomson, Reference Thomson1859 are known from Staratschin (Starostin) (Palaeocene: Heer Reference Heer1870) and Rott (Lower Miocene: Heyden & Heyden Reference Heyden and Heyden1866). The fossils described here belong to seven species of the subfamilies Hydraeninae and Ochtebiinae. Unfortunately, a precise generic discrimination is impossible for many Bembridge remains of this family. Important structures such as maxillary palpi are invisible or scarcely visible in all of the specimens examined. Some of these specimens are more reminiscent, in many characters, of the members of Ochthebius Leach, Reference Leach1815 and Limnebius Leach, Reference Leach1815. In other cases, when a generic attribution raises doubt or cannot be correlated to the genera accepted in the Recent fauna, the species examined are regarded in a composition of formal genera or morphogenera rather than genera proposed for the Recent fauna and need further clarification.
Hydraenidae, commonly called minute moss beetles (in contrast to somewhat similar mould beetles (Latridiidae) with six abdominal ventrites), have the head partly more or less retracted inside the prothoracic segment and with long maxillary palpi, larger prothorax, narrowly separated from (sub)contiguous coxae in each pair of legs. Representatives of this group could be confused with small oval species of Staphylinidae with long elytra; however, the pronotum of Hydraenidae in most cases is characteristic enough to distinguish small specimens of both families. Classification and division into subfamilies, tribes and genera is still problematic and requires further study (Perkins Reference Perkins1997). A traditional interpretation of the system is accepted here, despite difficulties in using the diagnostic characters to identify fossil specimens.
Subfamily Ochthebiinae Thomson, Reference Thomson1859
This subfamily is frequently regarded as more archaic than the subfamily Hydraeninae and other groups of the family because of presence in some Recent groups of some plesiotypic features (Perkins Reference Perkins1997) and being less diverse. Discrimination of the subfamilies of fossil remains is quite problematic because of missing or lack of most or all diagnostic characters elaborated by neontologists (palpi of mouthparts, gland concentrations on head or prothorax, prohypomeran antennal pockets, etc.). In most cases subfamily attribution was determined by additional unusual diagnostic characters.
Genus Ochthebius Leach, Reference Leach1815
Type species. Elophorus marinus Paykull, Reference Paykull1798 (by subsequent designation by Orchymont Reference Orchymont1942), Recent.
The specimen In. 43411 (holotype of Ochthebius rossi sp. nov.), like Recent members of the genus, is characterised by the comparatively short maxillary palpi, maximal width of pronotum somewhat before the middle, not very long metaventrite, broadly arcuate elytral sides. It looks somewhat like the Recent species of Hydraenida Germain, Reference Germain1901 from the Patagonian Region, but differs from them in the markedly shorter maxillary palpi (allowing to determine it as a member of Ochtebiinae). The genus Ochthebius is found in all zoogeographical regions, although the majority of species are known from the Northern Hemisphere. It was recorded from the Palaeogene (Ochthebius nitidulus Heer, Reference Heer1870) and Neogene (Ochthebius plutonis Heyden and Heyden, Reference Heyden and Heyden1866), although the specimens used for proposal of these species need to be re-examined to clarify their generic attribution. The Recent members of the genus are characteristic of stagnant water basins, they are algophagous inhabitants of shallow water or litter at the water edge. Unfortunately, the characters available for study do not allow us to determine any reliable attribution of this new species. The specimens In. 26131 and In. 26147 (Metacoxites ventritalis gen. & sp. nov.) are somewhat similar to its type specimen (In. 43411), although metacoxae of In. 26131 and In. 26147, in contrast to the Recent members of the genus Ochthebius, are not transverse, but rather oblique.
Ochthebius (subgenus incertus) rossi Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:1AB77FC2-0A74-455B-9CAF-BFAF295550A4
Derivation of name. The epithet of this new species is devoted to the palaeontologist A. J. Ross, coordinator of the INTAS project, in the framework of which these studies on insects became possible.
Material. Holotype NHMUK In. 43411 (Figs 6a, 8a): most part of the ventral surface from inside (negative print) somewhat rotated, with the well-preserved right side of all sclerites (prothoracic segment and head somewhat detached) and abdomen somewhat turned and with apex extended inside the limestone matrix, with clear right maxillary palp, left profemur, right mesofemur, left intermediate leg and both posterior legs; Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. This new species has an abnormally short prothoracic segment, which makes it possible to distinguish it from the Recent congeners. Besides, it has comparatively wide procoxae studied from inside. It differs from the Metacoxites ventralis gen. & sp. nov. in the longer mesoventrite, shorter metaventrite, transverse metacoxae, shorter abdominal ventrite 1 and shorter ultimate ventrite. The Palaeogene Ochthebius nitidulus (Heer, Reference Heer1870) and Neogene Ochthebius plutonis are described without clear indication of the characters for their proper diagnostics and need to be revised.
Description. Holotype In. 43411: length of specimen 1.6mm, width 0.6mm, length of elytra apparently 1.0mm. Body elongate, apparently slightly convex dorsally and moderately convex ventrally; light brown in colouration. Thoracic ventrites, metacoxae and abdominal ventrites apparently with smooth sculpture and indistinct punctures and without visible ‘frontoclypeal' suture. Head of usual shape (somewhat shorter than wide at eyes), subtriangular, with moderately large oval eyes. Labrum invisible. Maxillary palpi about three-fifths as long as the head is wide, ultimate palpomere somewhat narrower and slightly longer than penultimate one. Pronotum about twice as wide as long, apparently convex; widest somewhat before the middle and gently narrowing both anteriorly and posteriorly; anterior edge moderately excised and posterior one somewhat convex; anterior angles rounded and posterior ones with distinct tops. Elytra about 1.6 times as long as wide combined; slightly narrowed at base, widely rounded along the middle and slightly wider than pronotum, apparently leaving uncovered the apex of abdomen. Procoxae distinctly transverse and extremely narrowly separated (nearly contiguous). Mesoventrite very long and longer than metaventrite. Mesocoxae comparatively large, suboval and slightly transverse, narrowly separated. Metaventrite clearly short (much shorter than usually seen in Hydraenidae). Metepisterna moderately narrow, at anterior edge slightly narrower than tibiae and gradually narrowing posteriorly. Metacoxae transverse and of usual outline, narrowly separated. Abdomen more than twice as long as metaventrite, with seven ventrites. Ventrite 1 about twice as long as ventrite 2, ventrites 2–5 subequal in length, ventrites 6 and 7 with rather arcuate anterior edge and almost as long as ventrite 1, hypopygidium widely rounded at apex. Epipleura much narrower than femora and somewhat wider than tibiae, more than twice as wide as distance between mesocoxae or that between metacoxae, very gradually narrowing posteriorly. Femora comparable in size and shape, about 3.5 times as long as wide and somewhat extended beyond outer edge of epipleura. Meso- and metatibiae rather narrow and comparable in size and shape, subparallel-sided in most length, somewhat shorter and somewhat less than half as narrow as femora. Meso- and metatarsi narrow and slightly longer than half the length of tibiae.
Subfamily Hydraeninae Mulsant, Reference Mulsant1844
This subfamily is the most numerous and diverse in the family and, therefore, appears to be the most successful in the Recent fauna. However, discrimination of the subfamilies in fossils is quite problematic because of missing or lack of most or all diagnostic characters, and in most cases the subfamily attribution was determined due to additional diagnostic characters, not the usual ones used by neontologists.
Genus Hydraenites Kirejtshuk, gen. nov.
Zoobank ID. urn:lsid:zoobank.org:act:75E95C48-454C-4EA7-BCE9-AAB0590EFF95
Type species. Hydraenites gracilimmus sp. nov.
Derivation of name. The name of this new genus is formed from Hydraena. Gender: masculine.
Notes. This new genus is represented by one species.
Diagnosis. The specimen examined has unclear outline of prothoracic and abdominal sclerites. The general configuration of the sclerites accessible for study is very similar to that in the representatives of Hydraena Kugelann, Reference Kugelann1794, although the Recent species of the genus Orchymontia Broun, Reference Broun1919 (Orchymontiinae) from the Australian and Novozealandian regions have somewhat similar sclerites. The fossil examined has a rather long head, which is rather narrowed apically and comparatively long metaventrite. Therefore, the treatment of this fossil as possibly a member of somehow a formal genus with the name produced from Hydraena and, respectively, in the composition of Hydraeninae seems to be reasonable. The new species is characterised by the rather elongate body, very long head and metaventrite. The new species seemed to have a bionomy similar to that in most Recent representatives of the family; it could be an algophagous inhabitant of shallow water or litter at the water edge.
Hydraenites gracilimmus Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:0339AD63-2C64-4309-B6C9-2CE3A3236110
Derivation of name. The epithet of this new species means ‘very graceful', ‘very elegant'.
Material. Holotype NHMUK I. 10189 (Figs 6b, 8b): underside from inside with the abdomen detached and somewhat moved to the right, right maxillary palpus and a part of left maxillary palpus, pro- and metafemora, mesofemora and mesotibiae as well as left epipleuron; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Holotype I. 10189: length of specimen 2.6mm, width apparently 0.8mm, length of elytra apparently 1.6mm. Body rather elongate, apparently moderately convex dorsally and ventrally; very light brown in colouration. Integument of mesoventrite and left metepisternum smoothed and without clear puncturation. Head of usual shape and rather long (somewhat longer than wide at eyes), subtriangular, with distinct ‘frontoclypeal' suture. Labrum invisible. Maxillary palpi apparently about as long as width of head, without clear separation of palpomeres. Prothoracic segment without clear outline. Elytra apparently leaving apex of abdomen uncovered, apparently about twice as long as their combined width; slightly narrowed at base, widely rounded along the middle and apices forming a joint arc. Prosternum apparently not carinate. Procoxae invisible. Mesoventrite moderately long. Mesocoxae comparatively large, suboval and slightly transverse, apparently narrowly separated. Metaventrite moderately long, somewhat longer than mesoventrite. Metepisterna moderately narrow, at anterior edge about as wide as tibiae and gradually narrowing posteriorly. Metacoxae transverse and of usual outline, moderately widely separated. Abdomen somewhat longer than metaventrite, very widely rounded at apex and with outlines of ventrites invisible. Epipleura very narrow, much narrower than tibiae, very gradually narrowing posteriorly. Femora comparable in size and shape, about three times as long as wide and somewhat extended beyond outer edge of epipleura. Mesotibiae rather narrow, somewhat shorter and somewhat less than two fifths as narrow as femora, narrowing at base and at apex.
Genus Metacoxites Kirejtshuk, gen. nov.
Zoobank ID. urn:lsid:zoobank.org:act:9D2EED21-AFD6-46AC-945A-0DE8B8A49042
Type species. Metacoxites ventritalis sp. nov.
Derivation of name. The name of this new genus is formed from ‘metacoxa'. Gender: masculine.
Notes. This new genus is represented by one species.
Diagnosis. The specimens In. 26131 and In. 26147 (1) (Metacoxites ventritalis sp. nov.) have metacoxae rather oblique in contrast to those in members of most generic groups of the family and, therefore, it seems to be reasonable to regard these specimens as a separate genus until there is a detailed generic revision of the family. Some modern species of Tympasnogaster Perkins, Reference Perkins1979 and Prototympasnogaster Perkins, Reference Perkins2018 with similar outline of pronotum and elytra, in contrast to Hydraenites gracilimmus sp. nov., have only slightly oblique metacoxae and also apparently rather wider tibiae. The type species of this new genus looks like some Ochthebius, some Hydraena and different members of several genera, but has very distinct metacoxae. All other characters accessible for study also give enough reason to regard this new species as belonging to the family Hydraenidae.
Metacoxites ventritalis Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:397E4E7B-B532-4112-839F-1D50C1BB1D54
Derivation of name. The epithet of this new species is formed from ‘ventrite' and refers to particular configurations of abdominal ventrites.
Material. Holotype NHMUK In. 26131 (Figs 6c, 8c): most of the ventral surface of thorax and abdomen from inside (negative print) as well as basal parts of epipleura, with the well-preserved right side of many sclerites, except for the place between prosternum and mesoventrite, right side of metaventrite and right side of abdomen; most visible parts with remains of chitin; Insect Limestone, Isle of Wight, Hooley Collection.
Additional specimen. NHMUK In. 26147 (1) (Fig. 8d): most of the ventral surface of head, thorax and base of abdomen from inside as well as basal parts of epipleura, profemora, left protibia and basal protarsomeres, bases of metafemora; prothorax and pterothorax slightly separated; print without trace of chitin and partly unclear outline of sclerites (together with remains of Scirtidae and Curculionoidea); Insect Limestone, Isle of Wight, Hooley Collection.
Description. Holotype In. 26131: length of specimen 1.4mm, width 0.6mm. Body elongate, apparently slightly convex dorsally and moderately convex ventrally; dark brown to blackish in colouration. Thoracic ventrites, metacoxae and abdominal ventrites apparently with coarse sculpture, small and moderately dense punctures. Head invisible. Pronotum about twice as wide as long, apparently convex; widest somewhat before the middle and gently narrowing both anteriorly and posteriorly. Elytra slightly narrowed at base, widely rounded along the middle and slightly wider than pronotum. Procoxae distinctly transverse and apparently extremely narrowly separated (nearly contiguous). Mesoventrite moderately long and as long as metaventrite. Mesocoxae comparatively large, suboval and slightly transverse, narrowly separated. Metaventrite moderately short. Metepisterna moderately narrow and gradually narrowing posteriorly. Metacoxae oblique and narrowly separated. Abdomen almost twice as long as metaventrite, with seven ventrites, abdominal ventrite 1 about as long as ventrite 2, ventrites 3–5 each somewhat longer than previous ones and subequal in length, ventrite 6 very short and ventrite 7 (hypopygidium) longest and widely rounded at apex (a narrow stripe exposed behind it – apparently the apex of pygidium and/or genital capsule). Epipleura narrow and very gradually narrowing posteriorly.
Variation. Additional specimen In. 26147: length 1.6mm, width 0.7mm. Body elongate, apparently slightly convex dorsally and moderately convex ventrally. Head somewhat longer than wide and subarcuate at labral apex (rather pressed in the limestone piece). Pronotum about twice as wide as long, apparently convex; widest somewhat before the middle and gently narrowing to both anteriorly and posteriorly. Elytra slightly narrowed at base, widely rounded along the middle and slightly wider than pronotum. Procoxae distinctly transverse and contiguous. Mesoventrite moderately long and somewhat shorter than metaventrite. Mesocoxae comparatively large, suboval and slightly transverse, narrowly separated. Metaventrite moderately short. Metepisterna moderately narrow and gradually narrowing posteriorly. Metacoxae oblique and narrowly separated. Epipleura narrow and very gradually narrowing posteriorly. Pro- and metafemora moderately wide, somewhat wider than epipleura. Protibia rather narrow. Protarsus very narrow.
Genus Davidraenites Kirejtshuk, gen. nov.
Zoobank ID. urn:lsid:zoobank.org:act:8ADD14ED-7546-4332-A827-C077431F224E
Type species. Davidraenites gratshevi sp. nov.
Derivation of name. The name of this new morphogenus is formed from Davidraena. Gender: masculine.
Note. This genus is represented by only three new species from Isle of Wight described here.
As a reliable generic discrimination according to the characters available in the remains examined would be hypothetical, the author decided to propose a new formal genus for the three species described here. However, the likelihood of a reasonable separate generic (or subgeneric) state for them is high.
Diagnosis and discussion. The structures of the three fossil remains are somewhat similar to the Recent members of the genus Davidraena Jäch, Reference Jäch1994 from India in the rather wide pronotum (markedly wider than elytra) and contiguous procoxae, and, therefore, they can be linked to this generic group. These fossil remains also resemble the Recent species of the Indo-Malayan genus Gonraena Jäch, Reference Jäch1994 (apparently nearest to the last mentioned genus), but differ from it in having the pronotum not wider than elytra and distinctly separated procoxae. Thus, all these fossil species have an intermediate appearance between Davidraena and Gonraena, and now it seems to be impossible to find a reliable character for decision of this taxonomic dilemma. Moreover, Davidraenites gratshevi sp. nov. shows the pronotum, which is similar to that in species of Gonraena rather than that in representatives of Davidraena. This new species also shows anterior pronotal angles somewhat projecting (as those in both Davidraena and Gonraena), while anterior angles of the prothoracic segment of the other two new species are seen only from below (excised anterior edge) and it is impossible to trace the anterior edge of the dorsal surface of this segment. It seems the other two species have the anterior angles of pronotum not projecting anteriorly rather than anterior edge. Taking into consideration that, in contrast to the Recent species of both mentioned groups, all new fossil species are only somewhat similar to them and have much larger eyes and frons gradually narrowing anteriorly, it could be reasonable to propose for these new species a new genus. The antennal club of Davidraenites gratshevi sp. nov. is distinctly three-segmented and almost compact, while Davidraena antennalis Jäch, Reference Jäch1994 has antennae without club, Davidraena boukali Jäch, Reference Jäch1994 with three-segmented loose club, and in both known species of Gonraena with two-segmented compact club. Finally, the lateral lines on prohypomera in the fossil species described here are closer to the prothoracic sides than in the Recent species of comparable genera. These fossils also partly resemble the representatives of the genus Macrogasma J. Sahlberg, Reference Sahlberg1900 from the Mediterranean in the shape of pronotum and head, but differ from them in the shape of head narrowing anteriorly, three-segmented antennal club (not five-segmented), pronotum with more or less arcuate sides, not carinate prosternum and (sub)contiguous procoxae. Some genera of Afrotropical, Capean and Malagasy Hydraeninae have a somewhat similar appearance to the new species described here as Davidraenites gen. nov. Unfortunately, all fossils treated here as this genus have no visible palpomere and, therefore, their subfamily and generic attribution was elaborated after the study of some additional (not principal) diagnostic characters. This particularly concerns the Recent species of the genus Decarthrocerus Orchymont, Reference Orchymont1948, also with a somewhat similar appearance, which, however, differs from the fossil species considered here as Davidraenites gen. nov. in having a subhexogonal pronotum, five-segmented antennal club and (sub)contiguous procoxae. The Recent species of Coelometopon Janssens, Reference Janssens1972 are also similar to the members of Davidraenites gen. nov., but differ from the fossil species in having a wider and more projecting head apex, strongly narrowed pronotal base and uneven elytral sides, somewhat sinuate behind shoulders.
Davidraenites gratshevi Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:D5413336-739A-4B5E-A19B-A6EAB5A4A40A
Derivation of name. The epithet of this new species is devoted to V. G. Gratshev, a brilliant coleopterist and participant of the project on Bembridge Insect Beds, who passed away suddenly and could not complete his valuable contribution to the Isle of Wight coleopterous fauna.
Material. Holotype NHMUK I. 10171 (Figs 6d, 9a): negative print, demonstrating the most part of ventral surface with remains of chitin from inner side, with the contour of epipleura, elytral sides, six left terminal antennomeres, apical part of left mesofemur and left mesotibia; only basal part of head clearly visible and its anterior part turned inside limestone matrix; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from Davidraenites intermixta sp. nov. and Davidraenites spurcus sp. nov. in the somewhat narrower and longer pronotum (pronotum of both about three times as wide as long), and is arcuately narrowing anteriorly and posteriorly; from the former also in the more flattened body and somewhat smaller eyes; and from the latter in the more slender body. The lateral lines on prohypomera in this new species are more or less S-shaped and the anterior angles of the prothoracic segment appear to project anteriorly. The lateral lines on prohypomera in other new species described here are C-shaped rather than S-shaped and the anterior angles of the prothoracic segment are very slightly projecting.
Description. Holotype I. 10171: male; length of specimen 1.6mm, width 0.8mm, length of elytra apparently 1.1mm. Body elongate, apparently slightly to moderately convex ventrally; light brown to reddish in colouration with darker head. Thoracic ventrites, metacoxae and abdominal ventrites apparently with smoothed sculpture and indistinct punctures and without visible ‘frontoclypeal' suture. Head rather wide, with moderately large oval eyes. Labrum and palpi invisible. Antennae with three-segmented not very compact club. Pronotum about twice as wide as long, apparently convex; widest behind the middle and gently narrowing both anteriorly and posteriorly; posterior edge somewhat concave; anterior angles apparently rather projecting anteriorly; posterior angles with distinct tops. Elytra about 1.4 times as long as combined width; slightly narrowed at base, widely rounded along the middle and slightly wider than pronotum, complete. Prothorax with S-shaped lateral lines on prohypomera; anterior edge rather excised and anterior angles narrowly rounded and posterior ones with distinct tops. Procoxae distinctly transverse and extremely narrowly separated (nearly contiguous). Mesoventrite very long and longer than metaventrite. Mesocoxae comparatively large, suboval and slightly transverse and contiguous. Metaventrite about as long as prosternum. Metepisterna moderately narrow, at anterior edge rather narrower than mesotibia and gradually narrowing posteriorly. Metacoxae transverse and of usual outline, rather narrowly separated. Abdomen more than twice as long as metaventrite, with seven ventrites, ventrite 1 about as long as ventrites 2 and 3 combined, ventrites 2–5 subequal in length, ventrites 6 and 7 with rather arcuate anterior edge and almost as long as ventrite 1, hypopygidium very convex at base and widely rounded at apex. Epipleura somewhat wider than metepisterna, very gradually narrowing posteriorly. Mesotibia very narrow and about as wide as ultimate antennomere.
Davidraenites interruptus Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:21F18337-7D1C-4F35-8B0C-09ED137E6A97
Derivation of name. The epithet of this new species means ‘particular', ‘single' and ‘disturbed'.
Material. Holotype NHMUK I. 10014 (Figs 6e, 9b): female; negative print, demonstrating the most part of the underside from inside, without visible appendages; only basal part of head clearly visible and its anterior part turned inside limestone matrix.
Diagnosis. This new species is more similar to Davidraenites spurcus sp. nov., but distinct from it in the more slender body with more convex underside, different shape of the prothoracic sides, C-shaped lateral line of the prohypomera and wider epipleura. See also the diagnosis of Davidraenites gratshevi sp. nov.
Description. Holotype I. 10014: female; length of specimen 1.4mm, width 0.6mm, length of elytra apparently 0.9mm. Body elongate, apparently rather convex ventrally; light brown to reddish in colouration. Thoracic ventrites, metacoxae and abdominal ventrites apparently with smoothed sculpture and indistinct punctures and without visible ‘frontoclypeal' suture. Head rather wide, with very large oval eyes. Labrum, palpi and antennae invisible. Pronotum about three times as wide as long, apparently convex; widest at the middle and gently narrowing both anteriorly and posteriorly; posterior edge apparently subtruncate; posterior angles apparently blunt. Elytra about 1.5 times as long as combined width; slightly narrowed at base, widely rounded along the middle and slightly narrower than pronotum, apparently leaving abdominal apex uncovered. Prothorax with C-shaped lateral lines on prohypomera; anterior edge shallowly emarginate and anterior angles widely rounded. Procoxae distinctly transverse and extremely narrowly separated (nearly contiguous) and comparatively wide. Mesoventrite moderately long and somewhat shorter than metaventrite. Mesocoxae comparatively large, suboval and slightly transverse and (sub)contiguous. Metaventrite somewhat longer than prosternum. Metepisterna rather narrow and very gradually narrowing posteriorly. Metacoxae transverse and of usual outline, rather narrowly separated. Abdomen more than twice as long as metaventrite, with six ventrites, ventrite 1 about 1.5 times as long as each of ventrites 2–4 combined, ventrite 5 longest, ventrite 6 moderately convex at base and widely rounded at apex. Epipleura about twice as wide as metepisterna, very gradually narrowing posteriorly.
Davidraenites spurcus Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:017A7955-5CEB-4D95-B85A-AE6D813C82E6
Derivation of name. The epithet of this new species means ‘dirty', ‘muddy', ‘sordid'.
Material. Holotype IWCMS 2018.41.1 (Figs 6f, 9c): most part of the ventral surface with remains of chitin from inner side, with the contour of epipleura, elytral sides, profemora and right protibia, mesotibiae; only basal part of head visible, and its anterior part turned inside limestone matrix and invisible; Insect Limestone, NW Isle of Wight.
Diagnosis. See the diagnoses of the previous species of the genus described here. In addition, the prosternum of this new species is markedly shorter than in the other species of the genus.
Description. Holotype IWCMS 2018.41.1: male; length of specimen 1.5mm, width 0.7mm, length of elytra apparently 0.8mm. Body elongate, apparently slightly to moderately convex ventrally; light brown to reddish in colouration with somewhat darker head. Thoracic ventrites, metacoxae and abdominal ventrites apparently with smoothed sculpture and indistinct punctures and without visible ‘frontoclypeal' suture. Head rather wide, with very large oval eyes. Labrum, palpi and antennae invisible. Pronotum about three times as wide as long, apparently convex; widest behind the middle, subangular at widest place and narrowing both anteriorly and posteriorly; anterior and posterior angles invisible. Elytra about 1.3 times as long as wide combined; slightly narrowed at base, widely rounded along the middle and slightly wider than pronotum, complete. Prothorax with C-shaped lateral lines on prohypomera; anterior edge gently emarginate and anterior angles widely rounded. Procoxae distinctly transverse and extremely narrowly separated (nearly contiguous). Mesoventrite very long and longer than metaventrite. Mesocoxae comparatively large, suboval and slightly transverse and contiguous. Metaventrite longer than prosternum. Metepisterna moderately narrow, at anterior edge slightly wider than tibiae and gradually narrowing posteriorly. Metacoxae transverse and of usual outline, rather narrowly separated. Abdomen more than twice as long as metaventrite, with seven ventrites, ventrite 1 about as long as ventrites 2 and 3 combined, ventrites 3–5 subequal in length and rather short, ventrites 6 and 7 with rather convex anterior edge and they are almost as long as ventrite 1, hypopygidium very convex at base and widely rounded at apex. Epipleura somewhat wider than metepisterna, very gradually narrowing posteriorly. Pro- and mesotibiae very narrow. Profemora of usual shape and about 2.5 times as wide as tibiae.
Gen. & sp. indet.
The remains, which seem to have an appearance somewhat like a member of Hydraena, cannot be determined further and, therefore, it is regarded here as an uncertain genus. Nevertheless, this species looks rather different from the rest of the species because of its very wide metepisterna and comparatively long abdominal ventrites.
Material. IWCMS 2018.14.2 (on the same block as the holotype of Davidraenites spurcus Kirejtshuk sp. nov.) (Figs 7a, 9d): right part of the ventral surface from inner side, with the contour of meso- and metaventrites, and also abdomen (female). Length 1.4mm. Body elongate and light in colouration. Mesocoxae narrowly separated. Metaventrite slightly longer than mesoventrite. Right metepisternum rather wide. Abdominal ventrites comparatively long.
NHMUK I. 9427: Hydraenidae gen. & sp. indet. or Staphylinidae gen. & sp. indet.; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 8499 (3): part of head, part of pterothorax and abdomen as well as some sclerites of legs (but no characters important for generic diagnostics); Insect Limestone, Isle of Wight, Brodie Collection.
NHMUK In. 25634: mostly part of the underside (negatively) with unclear outlines, although the apical ventrite characteristic of the Hydraenid male is seen (together with two pairs of small elytra of Curculionoidea, one elytron of Hydrophilidae, some ventrites and two small bodies of Coleoptera incertae sedis); Insect Limestone, Isle of Wight, Hooley Collection.
Genus Eolimnebius Kirejtshuk, gen. nov.
Zoobank ID. urn:lsid:zoobank.org:act:77A432BE-6270-44DD-B771-97A5F8654747
Type species. Eolimnebius fossilis sp. nov.
Derivation of name. The name of this new genus is formed from the Greek prefix ‘eos' (dawn) and generic name ‘Limnebius'. Gender: masculine.
Notes. This new genus is represented by one species.
This new genus seems to be more closely related to the Hydraena genera-complex rather than Limnebius Leach, Reference Leach1815 because of similarity in proportions of sclerites of pterothorax and structure of abdomen, although the appearance of the species included is rather reminiscent of the genus Limnebius.
Diagnosis. This new genus is most similar in body shape to Limnebius, but Eolimnebius fossilis sp. nov. differs from many species of Limnebius in the almost complete elytra with narrowly rounded apices. The head of the new species examined is comparatively narrower than in the species of Limnebius and the abdomen of the new species, in contrast to the latter, appears to consist of only six ventrites. It differs from the species of Hydraena in having the pronotum widest before the base and with an even lateral outline along the elytra and pronotum; the type species of this new fossil genus is distinct in having a much shorter body.
Eolimnebius fossilis Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:D8BED4B9-9969-4B3F-8EE3-9BB9CB603DC1
Derivation of name. The epithet of new species means ‘fossil'.
Material. Holotype NHMUK I. 8592 (Figs 7b, 10): negative impression, showing most of the pterothorax and abdomen from inner side; also the contour of epipleura, elytral sides and apices; prothorax and head pressed in the limestone matrix; right profemur, right metafemur and metatibia; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Holotype I. 8592: length of specimen 1.4mm, width 0.8mm, length of elytra apparently 1.0mm. Body oval, apparently slightly convex dorsally and flattened ventrally; light brown in colouration with darkened prothorax and head. Integument without trace of sculpture. Head declined and looking like the usual shape for Limnebius (about half as wide as pronotal base), widely arcuate anterior edge and with moderately large oval eyes. Labrum and palpi invisible. Pronotum more than twice as wide as long, apparently convex; widest at base and gently narrowing anteriorly; anterior edge invisible; posterior angles seemingly with distinct tops. Elytra about 1.3 times as long as combined width; slightly narrowed at base, widely rounded along the middle and slightly wider than pronotum, their narrowly rounded apices extended beyond apex of abdomen. Prosternum and procoxae invisible. Mesoventrite moderately long and less than 1.5 times as long as metaventrite. Mesocoxae comparatively large, suboval and slightly transverse, very narrowly separated to contiguous. Metaventrite moderately short. Metepisterna moderately narrow, at anterior edge slightly wider than tibiae and gradually narrowing posteriorly. Metacoxae transverse and of usual outline, narrowly separated. Abdomen more than twice as long as metaventrite, apparently with six ventrites (although ultimate segment with unclear outline and may consist of two ventrites), ventrite 1 about twice as long as ventrites 2, ventrites 2–5 subequal in length, last apparent segment somewhat longer than ventrite 1, subtriangular and rounded at apex. Epipleura slightly narrower than femora and somewhat wider than tibiae, more than twice as wide as distance between mesocoxae or that between metacoxae, very gradually narrowing posteriorly. Metafemur about as wide as length of metacoxa. Metatibia moderately narrow and almost half as wide as metafemur, slightly widening apically.
Discussion. Taking into consideration the data on the bionomy of the Recent Hydraenidae, their extinct relatives seem to be one of the most indicative components of swamps during the deposition of the Bembridge Marls. If these water basins were quite shallow and overgrown by Typha and other swamp plants, they provided optimal ecological conditions for species of this family. They could inhabit the water basin and as litter along the shore. It is not clear why this group is so rare in other examined localities with lacustrine deposits. If attribution of the Mesozoic members of the family (Ponomarenko Reference Ponomarenko, Arnoldi, Zherikhin, Nikritin and Ponomarenko1977) is correct, this family should be regarded as a rather old group.
Family Staphylinidae Latreille, Reference Latreille1802

Figure 11 Beetles of the family Staphylinidae. (a) Gen. & sp. indet. (Tachyporinae, Mycetoporini): NHMUK I. 9570, body, ventral view; length of specimen = 4.2mm. (b) ?Bledius sp.: NHMUK I. 9612, body without head, lateral view; length of specimen = 5.3mm. (c) ?Bledius sp.: NHMUK In. 26087, thorax and abdomen, ventral view; length of specimen = 5.0mm. (d) specimen of Euaesthetinae gen. & sp. indet.: NHMUK I. 10271, body, dorsal view; length of print = 2.8mm. (e) Gen. & sp. indet. (Paederinae, Pinophilini, Pinophilina): NHMUK In. 17423, pronotum, elytra and part of abdomen, dorsal view; length of specimen = 6.0mm. (f) Gen. & sp. indet. (Paederinae, Paederini, Lathrobiina): IWCMS 2014.133 part, body, dorsal view; length of specimen = 4.8mm. (g) Gen. & sp. indet. (Paederinae, Paederini, Lathrobiina): IWCMS 2014.133 counterpart, body, ventral view; length of specimen = 4.8mm. (h) Gen. & sp. indet. (Paederinae, Paederini, Lathrobiina): NHMUK I. 8580, head and prothorax, ventral view; length of specimen = 1.7mm. (i) Gen. & sp. indet. (Paederinae, Paederini, Stilicopsina): NHMUK I. 10077, body, ventral view (opened and reversed elytra in dorsal view); length of specimen = 1.0mm.

Figure 12 Beetles of the family Staphylinidae. Subfamily incertae sedis. (a) NHMUK I. 10269, body, lateral view; length of specimen = 5.3mm. (b) NHMUK I. 9998, body, dorsal view; length of specimen = 6.3mm. (c) NHMUK I. 8526, body, ventral view; length of specimen = 9.5mm. (d) NHMUK I. 10233, elytra and abdomen, dorsal view; length of specimen = 4.9mm. (e) NHMUK I. 17384 (counterpart to NHMUK I. 10233), meso-, metathorax and abdomen, ventral view; length of specimen = 6.3mm.
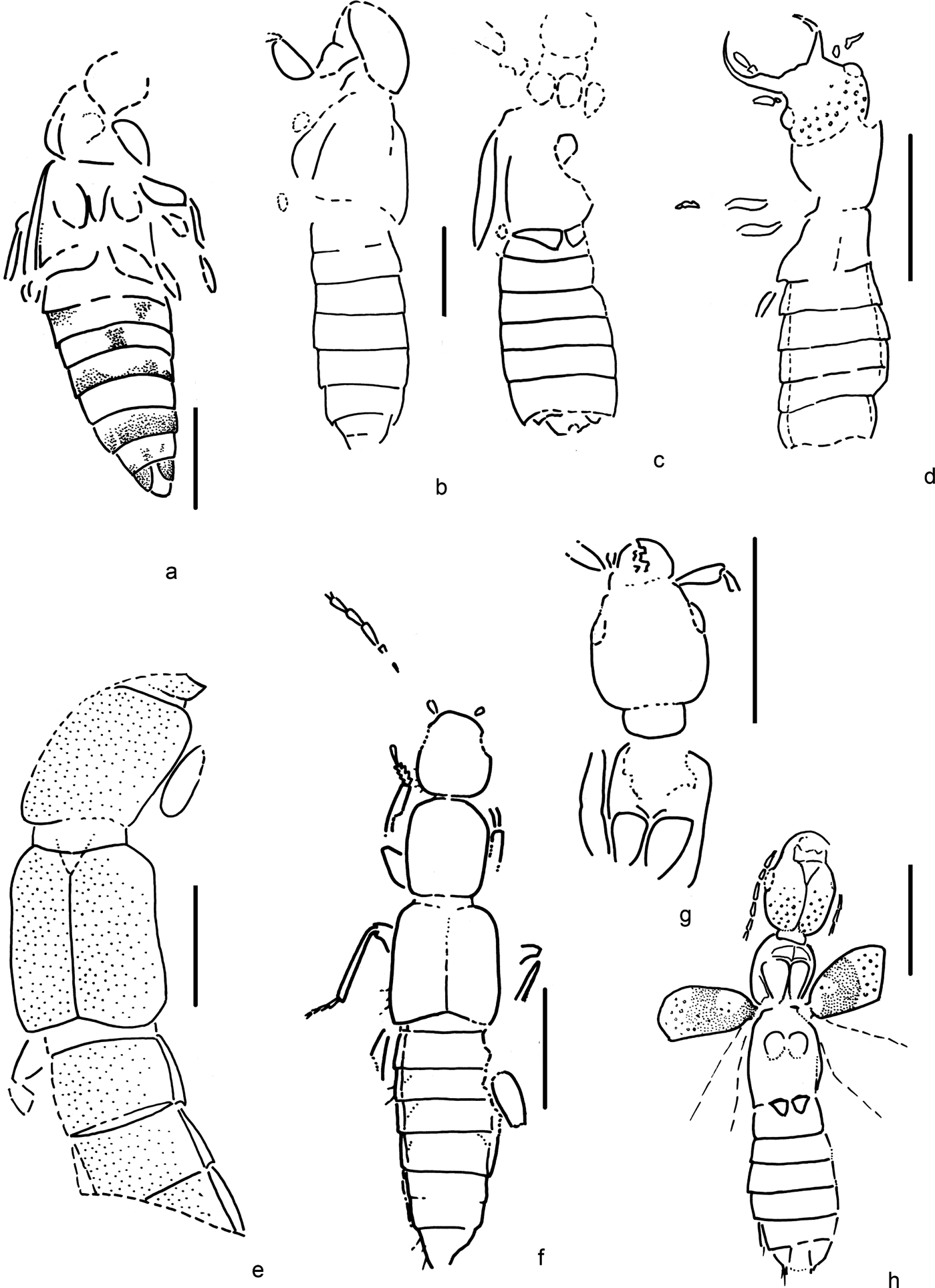
Figure 13 Suborder Polyphaga, family Staphylinidae. (a) Gen. & sp. indet. (Tachyporinae, Mycetoporini), NHMUK I.9570, body, ventral view. (b) ?Bledius sp., NHMUK I. 9612, body without head, lateral view. (c) ?B. sp., NHMUK In. 26087, thorax and abdomen, ventral view. (d) Specimen of Euaesthetinae gen. & sp. indet., NHMUK I. 10271, body, dorsal view. (e) Gen. & sp. indet. (Paederinae, Pitophylini), NHMUK I. 17423, pronotum, elytra and part of abdomen, dorsal view. (f) Gen. & sp. indet. (Paederinae, Paederini), IWCMS 2014.133, body, dorsal view. (g) Gen. & sp. indet. (Paederinae, Paederini), NHMUK I. 8580, head and prothorax, ventral view. (h) Gen. & sp. indet. (Paederinae, Paederini), NHMUK I. 10077, body, ventral view (opened and reversed elytra in dorsal view). Scales in mm.
Material. A total of 83 specimens including one larva with length 5.5mm (NHMUK I. 9119 (1)) (preliminarily identified at the family level by A. G. Kirejtshuk, A. G. Ponomarenko and A. S. Kurochkin); 11 best preserved specimens studied by A. Yu. Solodovnikov.
With nearly 3900 genera and more than 63,000 species worldwide in the Recent fauna, the family Staphylinidae (rove beetles) is one of the largest and most biologically diverse groups of Coleoptera. The majority of Recent Staphylinidae are predators inhabiting various decaying substrates on the soil surface. Most Staphylinidae can be easily recognised as fossils at the family level by short truncate elytra exposing more than a half of the flexible abdomen combined with large, projecting procoxae. However, identification of fossil Staphylinidae to any level below, and especially to genus, is very difficult, because important diagnostic characters are often not visible in fossils. The earliest fossil specimen attributed to Staphylinidae is with unclear sister relationships inside the family known from the Triassic (Chatzimanolis et al. Reference Chatzimanolis, Grimaldi, Engel and Fraser2012b). Beginning from the Middle Jurassic (e.g., Ryvkin Reference Ryvkin and Rasnitsyn1985; Cai et al. Reference Cai, Huang, Thayer and Newton2012, Reference Cai, Yan, Beattie, Wang and Huang2013a, Reference Cai, Yan and Vasilenkob), diverse fossil Staphylinidae mostly assigned to modern subfamilies and presenting sites of different origin were found through the Mesozoic (Chatzimanolis Reference Chatzimanolis, Betz, Irmler and Klimaszewski2018). Overall, groups of rove beetles from the subfamilies Omaliinae, Glypholomatinae, Oxytelinae and Tachyporinae, considered more basal, are predominant among fossils from the Jurassic. During the Cretaceous the more derived subfamilies like Staphylininae, Paederinae, Scydmaeninae and Aleocharinae were becoming represented and abundant. The Caenozoic Staphylinidae are also notable and comparatively diverse, especially from the comparable Upper Eocene outcrops relevant for this study (Florissant fossil beds and Green River formation in the USA, Baltic amber in Europe). Although the Caenozoic fossils are mainly known from outdated literature that needs a substantial revision, often they clearly represent Recent genera that demonstrate occurrence in places not correspondent with the Recent distribution. Like the better known Baltic amber inclusions (Chatzimanolis & Engel Reference Chatzimanolis and Engel2011; Bogri et al. Reference Bogri, Solodovnikov and Żyła2018), Staphylinidae from the Bembridge Marls seem to represent Recent, or at least very close to Recent, genera, many of which are no longer distributed in Europe. Mostly, they present entire beetle bodies with lack of some organs. Therefore, a precise identification of such specimens to species, genus or even tribe or subfamily level is often impossible.
Subfamily Tachyporinae MacLeay, 1825
The earliest member of this subfamily is a species from the Middle Jurassic Jiulongshan Formation in China, originally misidentified as the omaliine-like Protostaphylinus mirus Lin, 1976 (Cai & Huang Reference Cai and Huang2010). A greater number of Tachyporinae were described from the Upper Jurassic (Tikhomirova Reference Tikhomirova and Rohdendorf1968; Cai et al. Reference Cai, Yan, Beattie, Wang and Huang2013a), a few others are known from the Cretaceous (the Mesozoic fossil record on Tachyporinae reviewed in Cai et al. Reference Cai, Huang and Solodovnikov2011). The Caenozoic Tachyporinae have been hitherto represented by nine extinct species from the Oligocene belonging to six genera: members of the extinct genus Paleosepedophilus Paśnik & Kubisz, Reference Paśnik and Kubisz2002, and five Recent genera, Bolitobius Leach, Reference Leach1819 (Schülke Reference Schülke2000), Lordithon Thomson, Reference Thomson1859 (Scudder Reference Scudder1900; Schülke Reference Schülke2000), Tachyporus Gravenhorst, Reference Gravenhorst1802 (Paśnik Reference Paśnik2005), Sepedophilus Gistel, Reference Gistel1856 (Paśnik & Kubisz Reference Paśnik and Kubisz2002) and Mycetoporus Mannerheim, Reference Mannerheim1831 (Scudder Reference Scudder1890). Also, a species of the Recent genus Coproporus Kraatz, 1857 has been recently described from the Miocene Dominican amber (Yamamoto & Takahashi Reference Yamamoto and Takahashi2016).
Tribe Mycetoporini Thomson, Reference Thomson1859
The specimen NHMUK I. 9570 is attributed to Tachyporinae based on its overall sublimuloid shape of the body, retractile head, large prominent procoxae and tapered abdomen. The characters usually used for diagnostics of tribes within this subfamily (Smetana Reference Smetana1983; Newton et al. Reference Newton, Thayer, Ashe, Cchandler, Arnett and Thomas2000) are not available for observation in this fossil. It can be a representative of either the tribe Tachyporini, or Mycetoporini because of its general appearance. Based on the characteristic body size, shorter elytra in relation to abdomen (often longer in Tachyporini) and colouration pattern, displaying pale abdomen with dark spots, it is more likely a member of Mycetoporini.
Gen. & sp. indet.
Material. NHMUK I. 9570 (Figs 11a, 13a): represents well-preserved shape of the body and its ventral surface from inside, with traces of the well-preserved shape of the body, left anterior procoxae and both mesocoxae, and well-preserved abdominal ventrites; head poorly preserved without trace of antennae or mouthparts; outline of prothorax poorly visible, but left anterior leg well preserved, except missing tarsus; entire right mesotibia; epipleural part of right elytron well preserved along its entire length; seven abdominal segments (III–IX) (ventrites I–VII) visible; segment IX (ventrite VII) with lateral tergal sclerites and entire (as a single plate) ventrite suggesting the male sex for the I. 9570; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Length of print 4.2mm, maximum width (at base of abdomen) 1.2mm, length of elytra (from shoulder to apex) 1.2mm; body sublimuloid, abdominal ventrites bicoloured, mostly pale with dark spots; head without distinct neck constriction; prothorax wider than long; procoxae large and prominent, profemora slightly longer than coxa, protibiae slightly shorter than profemora; elytra slightly longer than wide; mesocoxae large, positioned close to each other but not contiguous; abdomen tapered apically, surface of abdominal ventrites apparently smooth, without puncturation; lateral sclerites of abdominal segment IX dark, sternite IX (ventrite VII) pale.
Subfamily Oxytelinae Fleming, Reference Fleming1821
The earliest member of the subfamily is known from the Upper Jurassic (Tikhomirova Reference Tikhomirova and Rohdendorf1968); then a few other oxytelines are known from the Cretaceous (Ryvkin Reference Ryvkin and Rasnitsyn1990; Yue et al. Reference Yue, Zhao and Ren2010; Cai et al. Reference Cai, Yan and Vasilenko2013b). Three Recent genera – Bledius Leach, Reference Leach1819, Platystethus Mannerheim, Reference Mannerheim1831 and Oxytelus Gravenhorst, Reference Gravenhorst1802 – were described from the Oligocene deposits in North America (Scudder Reference Scudder1876, Reference Scudder1878, Reference Scudder1890, Reference Scudder1900a, Reference Scudderb) and Europe (Heer Reference Heer1862; Förster Reference Förster1891). All records in the latter papers need to be revised. The fossil record of Oxytelinae has been recently reviewed in Lü et al. (Reference Lü, Cai and Huang2017).
Genus Bledius Leach, Reference Leach1819
Type species. Bledius armatus (Panzer, Reference Panzer1799), by monotypy, Recent.
The fossil record for this genus consists of seven species described from the Oligocene (Scudder Reference Scudder1878, Reference Scudder1890, Reference Scudder1900a) and Miocene (Heer Reference Heer1862). However, as far as can be judged from their descriptions and illustrations, at least neither species described by Scudder belong to this genus. Bledius morsei Scudder, Reference Scudder1900a and Bledius soli Scudder, Reference Scudder1900a can be clearly identified as species of the genus Stenus (Steninae); Bledius faecorum Scudder, Reference Scudder1900a as some species from the subfamily Aleocharinae; Bledius osborni Scudder, Reference Scudder1900a as an oxyteline from the genus Oxytelus or similar. The identity of Bledius primitarium Scudder, Reference Scudder1900a is unclear, but that species clearly does not have geniculate antennae and its general appearance is very different from that of Bledius. Based on the shape of metacoxae, it might be a member of Staphylininae or Paederinae. Definition of Bledius adamus Scudder, Reference Scudder1878, Bledius glaciatus Scudder, Reference Scudder1890 and Bledius speciosus Heer, Reference Heer1862 also needs to be clarified by revision of the type specimens.
Two specimens NHMUK I. 26087 (Figs 11c, 13c) and NHMUK I. 9612 (Figs 11b, 13b), both incompletely preserved, are attributed to the subfamily Oxytelinae because of the following reasons. The specimen I. 26087 has oxyteline-like metacoxae and an indication of seven visible entire abdominal ventrites, a unique synapomorphy for most oxytelines (Newton Reference Newton1982). Specimen ‘I. 9612' resembles the genus Bledius in habitus, and also has an indication, although weaker, of seven entire visible segments of the abdomen. Finally, both specimens match each other in size and, thus, they are assumed to be conspecific. Some available structural features are not diagnostic for the genus Bledius or even for the tribes of Oxytelinae (Herman Reference Herman1970; Newton Reference Newton1982); however, since they do not contradict the diagnosis of Bledius, these specimens are tentatively identified as one species of this genus.
?Bledius sp.
Material. NHMUK I. 26087: ventral surface from inside, with the cast of abdominal ventrites; well-preserved prints of meso- and metathoraces, poorer preserved impressions of pro- and mesocoxae, and good impressions of metacoxae, especially right one; epipleural part of right elytron is well preserved as impression along the entire length of elytron; Insect Limestone, NW Isle of Wight, Hooley Collection. NHMUK I. 9612; lateral view of the body with the cast of posterior part of prothorax, entire part of meso- and metathoraces (i.e., these parts preserve left side of the body); abdomen represented as lateral print of its right side; no legs are preserved except partial print of profemora, probably left; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Length from base of elytra to apex of abdomen 3.8mm (length of entire body appears to be over 5mm); width of abdomen at sternites VI–VII (ventrites IV–V) 1.3mm; length of elytra 1.2mm; body parallel-sided along most of its length, rather cylindrical, microsculpture and colouration not preserved; pronotum robust, only slightly narrower than meso- and metathoraces; procoxae large, somewhat prominent, contiguous; abdomen cylindrical, with seven visible entire sternites (II–VIII) (ventrites I–VI), although sternite VIII (ventrite VI) poorly visible.
Subfamily Euaesthetinae Thomson, Reference Thomson1859
Libanoeuaesthetus pentatarsus Lefebvre et al., Reference Lefebvre, Vincent, Azar and Nel2005 from the Lower Cretaceous Lebanese amber, Octavius electrospinosus Clarke & Chatzimanolis, Reference Clarke and Chatzimanolis2009 from the boundary of Lower/Upper Cretaceous Burmese amber and Stenaesthetus sp. (Puthz, Reference Puthz2006) from Baltic amber were hitherto the only known fossil representatives of this subfamily.
Specimen I. 10271 is identified as a member of this subfamily owing to the small size and shape of the body, structure of the mandibles, which are slender, falcate and without molar lobes, and also the insertion of antennae in front of eyes and distinctly in from side of head. Because of incomplete preservation, this specimen displays no characters diagnostic for tribe (Scheerpeltz Reference Scheerpeltz, Hanström, Brink and Rudebeck1974) or genus. It cannot be easily matched with any of the Recent genera, partly because the latter can be quite similar to each other. From Libanoeuaesthetus pentatarsus and previously mentioned species of Stenaesthetus, it differs in the larger body size, shape of the head with smaller eyes, pronotum with sides gradually converging posteriorly and shorter elytra without pronounced shoulders. Therefore, the specimen I. 10271 is named here as uncertain genus and species.
Gen. &. sp. indet.
Material. NHMUK I.10271 (Figs 11d, 13d): dorsal surface from inside with well-preserved body sclerites and mandibles, fragments of bases of antennae, right maxillary palpus and femora; head with rough puncturation, surface of thorax and abdomen unclear because of bad preservation; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Body length 2.8mm; head slightly wider than long (length 0.5mm, width 0.7mm), with rather small but prominent eyes, with temples broadly rounded towards neck, which seems to be more than 0.5 times as wide as head; mandibles 0.6 mm long, falcate, without molar lobes or teeth internally; antennae preserved by traces of their basal parts, indicating their insertions anteriorly from eyes and distinctly at sides of head; puncturation of head coarse; pronotum about as wide as long, widest at anterior angles, converging posteriorly, about as wide as head; elytra poorly preserved, but seemingly shorter than pronotum, widening posteriorly; abdomen moderately elongate, narrowing at base, widest at segments V–VI, its apical part poorly preserved; laterosternites distinct.
Subfamily Paederinae Fleming, Reference Fleming1821
There are many ambiguous fossils recorded in literature (e.g., Rayner & Waters Reference Rayner and Waters1991) that could be preliminarily identified as Paederinae; however, all known fossils that are unambiguously attributed to this subfamily belong to the Eocene–Oligocene, and most of them have been attributed to the Recent genus Lathrobium Gravenhorst, Reference Gravenhorst1802. But, as found very recently in Solodovnikov et al. (Reference Solodovnikov, Yue, Tarasov and Ren2013), the genus Mesostaphylinus Zhang, Reference Zhang1988, described as Staphylinidae incertae sedis from the Lower Cretaceous, seems to represent the oldest known and probably extinct lineage of Paederinae. Staphylinidae from the Bembridge Marls include five specimens (three species) that can be unambiguously identified as members of the subfamily Paederinae. Although neither of them has an entire set of tribal-, subtribal- or genus-level diagnostic characters preserved, based on the traits available for observation and the habitus, two species are placed in the tribe Paederini, and one in the tribe Pinophilini. Within Paederini, NHMUK I.17423 and IWCMS 2014.133 can be attributed to a species in the Lathrobium-complex of genera of the subtribe Lathrobiina, while the specimen I. 10077 apparently belongs to some genus from the subtribe Stilicopsina. The bad preservation of the available specimens makes it difficult to reach a more detailed identification.
Tribe Paederini Fleming, Reference Fleming1821
Subtribe Stilicopsina Casey, 1905
Until now this subtribe was not recorded as fossil. The specimen NHMUK I. 10077 is assigned to Stilicopsina based on the shape of the body, fused gular sutures, as well as the structure of the prosternum, which is expanded laterally and posteriorly, and narrowly separated from prohypomera. Thin and elongate antennae, long mandibles, confluent gular sutures, neck about 0.3 times as wide as head, very characteristic rough sculpture of the body surface and colour pattern of elytra, with pale spots at their shoulders and apical portions, all characters together support such placement. Within the Stilicopsina, the Recent members of the widespread Recent genera Dibelonetes Sahlberg, Reference Sahlberg1847 and Stilicopsis Sachse, Reference Sachse1852 could be the most likely match for this species based on the habitus. However, none of the characters diagnostic at the genus level in this complex are preserved in the specimen under consideration.
Gen. & sp. indet.
Material. NHMUK I. 10077 (Figs 11i, 13h): ventral surface from inside of the entire body, except elytra; elytra moved aside and dorsal surface of each elytron preserved as a cast; fragment of right antenna and obscure contour of the mouthparts with closed mandibles, prints of procoxae and metacoxae visible; slight traces of wings without clear venation exposed; puncturation and sculpture of integument unclear, although puncturation on elytra quite distinct; traces of darker areas on pale colouration on elytra visible; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Head oblong (length about 1.0mm, width about 0.7mm), oval because of broadly rounded temples; eyes poorly traced but seem to be relatively large, slightly shorter than temples; neck about third of head width; mandibles seemingly long and sickle-shaped, closed and thus poorly visible; gular sutures confluent; ventral surface of head roughly punctured; antennae only slightly shorter than length of head and pronotum combined, their presumed fourth–sixth antennomeres elongate, each much longer than wide; pronotum about as long and as wide as head, widest before its middle, with broadly rounded anterior and posterior angles; procoxae large, prominent, contiguous; elytra 0.7 mm long, each elytron 0.5mm wide, with rough and moderately dense puncturation, sparser than that on head; elytra bicoloured with pale spot at each shoulder and broad pale area in posterior half; mesocoxae (sub)contiguous; metacoxae narrow, subtriangular, with apex projecting posteriorly; abdomen widening from base, widest at segments VI–VII; lateral tergal sclerites (laterosternites) of abdominal segment IX apically sharply pointed.
Subtribe Lathrobiina Laporte, Reference Laporte1835
The subtribe is known in the fossil record by nine species of the genus Lathrobium Gravenhorst, Reference Gravenhorst1802 (Abdullah & Abdullah Reference Abdullah and Abdullah1968; Zhang Reference Zhang1989; Paśnik & Kubisz Reference Paśnik and Kubisz2002).
Three prints from the Bembridge Marls (two of them could be part and counterpart of the same beetle) are attributed to one species of this subtribe based on their habitus, and (in NHMUK I.8580) visible short robust mandibles with strong internal cuspidate tooth. Although none of the specimens has well-preserved diagnostic characters for the genus-level identification, their membership of the Lathrobium-complex of genera can be certainly suggested. Based on the relative abundance of species identified as Lathrobium in Baltic amber (e.g., Paśnik & Kubisz Reference Paśnik and Kubisz2002), it can be assumed that around the Eocene–Oligocene boundary this relatively thermophilous complex of several morphologically similar genera was at least as numerous in species as it is today.
Gen & sp. indet.
(Figs 11f–h, 13f, g)
Material. Part IWCMS 2014.133 (on the same block as the holotype of the wasp Exeristes gurnetor Khalaim, IWCMS 2012.574) (Figs 11f, 13f): dorsal surface from inside (positive print), with well-preserved body sclerites, fragment of right antenna, almost entire right anterior and middle legs; fragments of left anterior (part of femur and tibia) and middle (part of femora and tibia) legs; fragments of metafemora. Counterpart (Fig. 11g): ventral surface from inside (negative print) with well-preserved body sclerites, fragment of right antenna, almost entire right anterior and middle legs; fragments of left anterior leg (part of femur and tibia) and left middle leg (part of femur and tibia); fragments of metafemora; well-preserved shape of mesocoxae.
NHMUK I. 8580 (Figs 11h, 13g): only head and prothorax, ventral surface from inside (negative print); well-preserved head with mandibles, fragment of right maxillary palpus, and basal segments of antennae; well-preserved procoxae and fragment of right anterior femur; Insect Limestone, NW Isle of Wight, Brodie Collection.
Note. All examined specimens are considered as conspecific based on the match in their size, and shape of head and prothorax.
Description. Length of body (Part IWCMS 2014.133) 4.8mm; head elongate (length 0.7mm, width 0.6mm), with temples slightly longer than eyes and broadly rounded posterior angles; antennae slightly longer than head and pronotum combined; mandibles short, robust, with broad base and long, strong tooth internally; pronotum slightly longer than wide (length 0.8mm, maximum width 0.7mm), widest before its middle, with rounded but distinct angles; elytra as long as combined width (1.0mm), wider than head and pronotum, with rounded but distinct shoulders; legs moderately long; procoxae large, elongate and prominent, contiguous.
Tribe Pinophilini Nordmann, Reference Nordmann1837
Subtribe Pinophilina Nordmann, Reference Nordmann1837
Relatively large body size; shape of the pronotum and elytra; dense, even puncturation of the body surface; visible posterior margin of a large eye demarking very short tempora; relatively long abdominal segments, each of them with basal tergal carina that has a pair of short lateral lines protruding towards apex of segment; these characters make it possible to assign the specimen I. 17423 to the tribe Pinophilini. Within the Pinophilini, it can be placed in the subtribe Pinophilina based on the seemingly well-developed abdominal paratergites. Based on the habitus, this species must be a member of, or be very similar to, Recent genera of Pinophilina such as Gridellius Fagel, 1963, Metapinophilus Gridelli, 1928 or Pinophilus Gravenhorst, Reference Gravenhorst1802. This is the first fossil record of the tribe.
Gen. & sp. indet.
Material. NHMUK I.17423 (Figs 11e, 13e): cast of right latero-dorsal part of pronotum, base of neck, dorsal part of elytra and abdominal tergites II–V (at least part of each) with well-preserved integument, fragment of right femora, right abdominal laterosternites; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Length of body (extrapolated from available size of pronotum, elytra and part of abdomen) about 9.5mm; pronotum slightly longer than wide, with distinct angles and distinct moderate puncturation; elytra as long as wide (1.8mm), parallel-sided, with distinct shoulders and with puncturation as that on head; abdominal tergites with puncturation similar to that on elytra and pronotum, but somewhat more faint.
Subfamily incertae sedis
Gen. & sp. indet.
(Fig. 12)
Material. NHMUK I. 9998 (Fig. 12b): dorsal surface from inside (positive print), with well-preserved body sclerites; length of the body 6.3mm; no appendages, except fragments of profemora and left antenna, are preserved; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 10269 (Fig. 12a): lateral right surface from inside, with well-preserved body sclerites; length of the body 5.3mm; no appendages are preserved, except extremely incomplete antennae, fairly good right procoxa and profemur, fragments of middle legs and presumably right metafemur; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 8526 (Fig. 12c): ventral surface from inside (negative print); head and pronotum seemingly detached from the rest of the body and slightly distorted basally; only head is preserved in the print, no trace of mouthparts or even eyes; parts of procoxae and metacoxae; overall shape of the body visible, length of the body 9.5mm; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 10233 (Fig. 12d), part: dorsal surface from inside (positive print); meso- and metathoraces (visible as elytra) and abdomen; incomplete procoxae; length of this posterior part of the print (meso-, metathoraces and abdomen) 4.9mm; Insect Limestone, Isle of Wight, Brodie Collection.
NHMUK In. 17384: ventral surface from inside; meso-, metathoraces, abdomen and all coxae somewhat visible; Insect Limestone, Isle of Wight, Smith Collection.
There is no doubt that all these specimens belong either to subfamily Paederinae or Staphylininae because of their body shape. Unfortunately, these specimens do not display any characters that would have made it possible to separate these subfamilies (like antennal insertions, hypomera of pronotum, shape of posterior coxae), and they also do not have enough details of general appearance to assign them unambiguously to any genus within either of these subfamilies. The frontal part of head, fragments of base of antennae inserted before eyes and close to each other, and possibly narrow neck (the latter character is not very clearly preserved) in the specimen I. 9998 suggest that it may belong to the tribe Xantholinini from Staphylininae. The shape of the abdominal sternite VIII (ventrite VI) with deep emargination, and apically sharp lateral tergal sclerites of segment IX, suggest that this male specimen, represented by two parts (I. 10233 and In. 17384), belongs to the subfamily Paederinae. The identity of I. 10269 and I. 8526 remains even more uncertain.
Subfamily Pselaphinae Latreille, Reference Latreille1802
(Fig. 7c)
Material. One specimen CAMSM, X.50140.206, Insect Limestone, Isle of Wight (identified by A. G. Kirejtshuk).
This subfamily is moderate in number, subcosmopolitan and particularly well represented in wet tropics. Recent Pselaphinae beetles are rather small in body-size (0.9–3.5mm). Their adults and larvae are associated mostly with mites, living in decaying vegetation, humus and moss, and require damp substrata (some of them are also myrmecophilous). In contrast to most rove beetles, their short elytra and abdomen are wider and heavily sclerotised; the latter is not very flexible and the ommatidia of their compound eyes are rather coarser. The remains examined are represented only by the underside of the pterothorax and the abdomen from inside, but easily diagnosed owing to the oval outline, widely separated meso- and metacoxae and rather long abdominal ventrite 1 in comparison with the remainder. The oldest remains of this groups were recorded from Lower Cretaceous Alava amber (Peris et al. Reference Peris, Chatzimanolis and Declós2014) and boundary of Lower/Upper Cretaceous Burmese amber (Rasnitsyn & Ross Reference Rasnitsyn and Ross2000; Grimaldi et al. Reference Grimaldi, Engel and Nascimbene2002), although the most fossil records have been described from the lowermost Eocene (Kirejtshuk & Nel Reference Kirejtshuk and Nel2013) and Upper Eocene Baltic amber (Spahr Reference Spahr1981; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984; etc.), where all subfamilies have been found. Only a few findings of this group originated from other deposits (Oke Reference Oke1957; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984).
Discussion. Taxonomically and ecologically, this small sample of identified Staphylinidae from the Bembridge Marls seems to be very diverse. The comparatively variegated tachyporins from the tribe Mycetoporini could possibly be associated with forest macrofungi. This assumption is based on the fact that among Recent Mycetoporini, the tribe associated with leaf litter or macrofungi, the fungi-associated species often have contrasting bi- or multicoloured bodies contrary to more unicoloured litter-dwellers. Two species, one Euaesthetinae and one presumed Paederinae (represented by complementing pieces NHMUK I. 10233 and NHMUK In. 17384), both with indistinct humeri and shortened elytra, are obviously flightless forest-leaf-litter dwellers confined to habitats with stable environmental conditions, as can be judged by analogy with many Recent members of these subfamilies with similar habitus and well-known bionomics. Two Paederinae species with obviously well-developed wings, one from the subtribe Stilicopsina and one from the Lathrobium-complex (subtribe Lathrobiina), could be confined to less stable habitats like wet leaf litter or debris at the edges of waterbodies. The examined fossil Bledius species could be a specialised inhabitant of the muddy banks of some brackish ponds or lagoon, where it dug tunnels in soil, similarly to many Recent species of this genus. Finally, by analogy with Recent species, the large-eyed fossil member of Pinophilini could be an arboreal species living in forest foliage. An overall, rather thermophilous aspect of the sampled fossil fauna is noteworthy, which even includes a predominantly tropical group like Pinophinini. Its overall composition suggests subtropical or, at most, warm temperate subtropical conditions in the area of fossilisation. A mixture of possible inhabitants of some wetland shores, and a forest, suggests that the former could be fossilised on site while the latter could have been carried to the lacustrine deposits for a short distance from their habitats by water.
Superfamily Scarabaeoidea Latreille, Reference Latreille1802
Family Scarabaeidae Latreille, Reference Latreille1802
Figure 14 Beetles of the family Scarabaeidae, and an unidentifiable specimen. (a) Holotype of Aphodius vectis Krell, sp. nov., holotype NHMUK In.17436: dorsal view; width of specimen = 6.78mm. (b) Unidentifiable remains of unknown family association, superficially resembling a head of a Dorcus-like stag-beetle: NHMUK, I. 10050. Pentodon dorcus Krell, sp. nov., holotype, NHMUK In.17431: (c) body, dorsal view; (d) counterpart; (e) oblique view. Scales in mm.

Figure 15 Beetles of the family Scarabaeidae. Abdomen specimen A, Scarabaeidae: Dynastinae, NHMUK In. 10370, ventral view: (a) part; (b) counterpart. Abdomen specimen B, Scarabaeidae: Dynastinae, ventral view: (c) counterpart, NHMUK In. 10390; (d) part, NHMUK In. 24652. (e) Abdomen specimen C, Scarabaeidae: Dynastinae, ventral view: NHMUK In. 25642. (f) Abdomen specimen D, possibly Scarabaeidae: Hopliinae, NHMUK I. 9838, ventral view. Scale in mm.

Figure 16 Suborder Polyphaga, family Scarabaeidae: holotype of Aphodius vectis Krell, sp. nov., NHMUK In. 17436, dorsal view; width of specimen = 6.78mm.
Material. Six specimens examined (nine specimens identified by A. G. Ponomarenko and A. G. Kirejtshuk as Scarabaeoidea; three of them considered by Krell to lack sufficient characters to be assigned to Scarabaeoidea).
With about 31,000 described extant species (Jameson & Ratcliffe Reference Jameson, Ratcliffe, Arnett, Thomas, Skelley and Frank2002), the Scarabaeoidea are one of the largest superfamilies in the Coleoptera. They are distributed worldwide and comprise such varied groups as dung beetles, stag beetles and chafers, ranging from just over 1 mm long to 170mm in body length. The larvae (grubs) develop in soil, dung, decaying wood and other organic matter. In the fossil record they are fairly well represented, with about 230 species described from the Upper Jurassic to the Pleistocene (Krell Reference Krell2007). During the Cretaceous this superfamily reached a considerable diversity, with most extant families represented (Krell Reference Krell2000, Reference Krell2006, Reference Krell2007; Nikolajev Reference Nikolajev2007). Scarabaeoidea are a monophyletic group diagnosed by two autapomorphies: antenna with a lamellate club and anterior border of hindwings with sclerotised field proximal to a pinch, as part of a spring folding mechanism (Krell Reference Krell2006). Both characters are rarely preserved in fossils. Most fossil scarab beetles were identified on the basis of other characters typical for Scarabaeoidea, such as enlarged prothorax adapted for digging with rather short and powerful legs, with tibiae toothed along outer edge, rather large and narrowly separated to contiguous pro- and mesocoxae, and transverse and narrowly separated to contiguous metacoxae. Most of those characters are not preserved in the specimens described here, which are classified on the basis of overall similarity to extant forms or similarities of body parts such as head, sternites or elytral sculpture.
Subfamily Aphodiinae Leach, Reference Leach1815
Genus Aphodius Illiger, Reference Illiger1798
Type species. Scarabaeus fimetarius Linnaeus, Reference Linnaeus1758, Recent.
Aphodius vectis Krell, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:CCEB807D-7EC9-439E-AC5E-66C3EDE93769
Derivation of name. Insula Vectis was the Roman name for the Isle of Wight. The species epithet vectis is used as a noun in apposition.
Material. Holotype NHMUK In. 17436 (Figs 14a, 16): a single piece showing the beetle from dorsal with spread elytra and remains of the right ala, with remains of the cuticula mainly on elytra, head and thorax; dorsal part of thorax and abdomen missing with imprint of the ventrum visible; counterpart indicated on the label, but missing; Insect Limestone, Isle of Wight, Smith Collection (NHMUK).
Diagnosis. Aphodius of 4.5mm body length with punctured elytral striae, probably smooth elytral intervals and truncated clypeus. Aphodius vectis is the only described species of Aphodiini from the Eocene (Krell Reference Krell2007). Two Aphodius species from Baltic amber (description in preparation) differ from A. vectis either by keeled elytral intervals or smaller body size.
Description. Holotype In. 17436: body length 4.5mm; object width (body with open elytra) 6.8mm; visible length of left elytra 2.8mm. Clypeus blackish brown, broadly truncate, obtuse-angled. Head almost as broad as pronotum. Pronotum dark brown, without traces of keels or impressions (but surface damaged). Elytra reddish brown, convex, with nine punctured striae, a further distal impression is likely to be the remains of a tenth stria; distance between points of striae one to three times their diameter; no humeral denticle; intervals flat and smooth; with possible traces of sparse large punctures of the size of the stria punctures. Traces of a fully developed right hindwing present. All coxae touching, hind femora slender. Five visible abdominal ventrites without traces of transversal rows of longitudinal impressions.
Remarks. The overall body shape indicates a species of Aphodiinae or Aegialiinae. Belonging to Aegialiinae cannot be excluded, but mandibles are not visible, which are likely to leave some traces in Aegialiinae fossils. Body shape combined with missing transversal rows of longitudinal impressions on the sternites indicate that the specimen belongs to Aphodiini. Ten punctured elytral striae, flat elytral intervals and pronotum without keels are typical characters for the mega-genus Aphodius. We use Aphodius in the traditional sense (Löbl & Smetana Reference Löbl and Smetana2006) and do not follow the upgrading of all subgenera of Aphodius to genera as proposed by Dellacasa et al. (Reference Dellacasa, Bordat and Dellacasa2001). Assignment to a subgenus (or genus sensu Dellacasa) is impossible since the relevant character-bearing body parts (legs, scutellum) are missing or invisible.
Subfamily Dynastinae MacLeay, 1819
Genus Pentodon Hope, Reference Horn1837
Type species. Scarabaeus punctatus Villers, Reference Villers1789, Recent.
Pentodon dorcus Krell, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:BE5EB818-437C-47F7-A56A-253319E19B25
Derivation of name. The species was originally reported as a stag beetle of the genus Dorcus MacLeay, 1819 (Lucanidae). The species epithet dorcus is used as a noun in apposition.
Material. Holotype NHMUK In. 17431 (Fig. 4c–e). Part: dorsal part of beetle with elytra ripped off; head missing, mediobasal part of pronotum damaged; right posterior part of abdomen missing. Counterpart: impression of inside of elytra and pronotum, colour and sculpture visible; elytral margins, left pronotal margin and posterior part of right pronotal margin incomplete; Insect Limestone, Isle of Wight, Smith Collection (NHMUK).
Note. The specimen is labelled as Dorcus In. 17431 (handwritten) and was reported as Dorcus sp. (Lucanidae) by Brodie (1878), Goss (Reference Goss1878), Woodward (Reference Woodward1878, Reference Woodward1879), Houlbert (Reference Houlbert1914), Franciscolo (Reference Franciscolo1997) and Krell (Reference Krell2000, Reference Krell2007), but has never been described.
Diagnosis. Ca. 22 mm-long beetle of the general shape of Pentodon, with three paired, punctured, elytral stripes, but pronotum with right hind angles, which distinguishes the species from other Pentodontini. It has half the body width of the only other dynastine described from the Eocene, Oryctoantiquus borealis Ratcliffe et al., Reference Ratcliffe, Smith and Erwin2005 from the Clarno Formation, Oregon, USA. The fossil congener, Pentodon bellerophon Heyden & Heyden, Reference Heyden and Heyden1866, from Rott (Lower Miocene or Upper Oligocene), is less than half as long as Pentodon dorcus. The other possible fossil congener, Pentodon proserpinae Heer, Reference Heer1862, from Oeningen (Miocene), is represented by a single elytron, which has similar size and sculpture as P. dorcus, but is almost 22 myr younger, hence unlikely to be conspecific.
Description. Holotype In. 17431. Part: specimen length 21.8mm; height (over sediment) 6.2mm; max width 11.3mm; length of pronotum 6.6mm. Counterpart: length of impression 21.9mm; max width of impression 10.8mm; length of left elytra 14.6mm; scutellum width 2.0mm; scutellum length 1.4mm. Pronotum smooth, broader than long, brown with greenish metallic shine (inside); laterally (visible on the right anterior angle), but neither anteriorly not posteriorly marginated; anterior margin with membranous hem; anterior angles slightly protruding from the straight anterior margin, posterior angles almost right-angled; sides slightly convex, more so closely behind the anterior angles. Scutellum triangular, 1.45 times as broad as long, with sharp or narrowly blunt tip. Elytra (as visible from the inside) brown with greenish metallic shine, with two longitudinal double lines of punctures, one in the middle and one in the distal half; traces of a third double line are visible on the proximal third of the right elytra; the double lines of punctures seem to be associated with (now flattened) keels or bulges. Remnants of dense punctures present in the posterior three quarters of the elytra (distance of punctures about their diameter). Traces of fully developed left hindwing present.
Remarks. Overall body shape, form of the pronotum and sculpture and puncturation pattern of the elytra resemble extant Dynastinae of the tribe Pentodontini. With the preserved set of characters, lacking head and legs, a reliable generic assignment cannot be made. The assignment to Pentodon is tentative and was chosen because it is the type genus of Pentodontini. Most characters fit Pentodon, but in extant species of this genus the hind angles of the pronotum are rounded or obtuse, whereas in Pentodon dorcus they are right-angled, which is unusual for Dynastinae. Metallic, structural colours differ between layers of the cuticula (as seen in fossils from the Messel Formation, pers. obs.). The metallic shine seen on the inside of elytra and pronotum need not be the original surface colour.
The specimen was originally identified as a stag beetle of the genus Dorcus (Lucanidae). Dorcus is generally more flattened than the fossil imprint and has a leathery cuticular microsculpture. Also the sutural stripe, if visible at all, is restricted to the anterior third to two-thirds of the elytra (Mizunuma & Nagai Reference Mizunuma and Nagai1994), whereas it reaches almost the tip of the elytra in Pentodon dorcus. In Dorcus, the posterior angles of the pronotum are truncated or even truncated-emarginated, not rectangular. Despite the paucity of key characters, the species is described and named to establish it in the palaeontological record and to correct its misidentification as Dorcus.
Gen. & sp. indet.
Four separate abdomina of two types are known from the Bembridge Marls that can be assigned to Scarabaeidae.
Type 1 (Scarabaeidae: Dynastinae)
Material. Specimen A – NHMUK I. 9854 (Fig. 15a) (part): median length of abdomen 10.5mm; maximum width of abdomen 14.4mm. NHMUK I. 10370 (Fig. 15b) (counterpart): median length of imprint 10.6mm; maximum width of imprint 14.6mm; Insect Limestone, NW Isle of Wight, Brodie Collection.
Specimen B – NHMUK In. 24652 (Fig. 15d) (part): maximum width of abdomen 11.6mm; median length of abdomen 8.7mm; Hooley Collection. NHMUK I.10390 (Fig. 15c) (counterpart): maximum width of imprint 9.6mm; Insect Limestone, NW Isle of Wight, Brodie Collection.
Specimen C – NHMUK In. 25642 (Fig. 15e): maximum length of specimen 15.6mm; no counterpart present; Insect Limestone, NW Isle of Wight, Hooley Collection.
All these abdomina show a large, broad and convex pygidium, with the apex bent to the ventral side so that the cloacal opening sits ventrally and not apically. The pygidia have a pronounced apical margin, broadened in the lateral angles. The border between penultimate and last ventrite is more deeper incised than the borders between the anterior ventrites, and the last ventrite is more convex than the anterior ones. All these characters are typical for Dynastinae and not found in this combination in any other group of Scarabaeoidea. It is possible that one or more of these abdomina are conspecific with Pentodon dorcus.
Type 2 (possibly Scarabaeidae: Melolonthinae: Hopliini)
(Fig. 15f)
Material. Specimen D – NHMUK I.9838 (Fig. 15f): maximum length of specimen 7.1mm; underside of abdomen with five visible ventrites bearing large and dense punctures, ventrite 5 broadly fused to the strongly sclerotised propygidium surrounding the narrow pygidium; pygidium apically broadly rounded, with large and dense punctures; Insect Limestone, Isle of Wight, Brodie Collection.
Within Scarabaeoidea, Glaresidae, Trogidae, most Lucanidae and Passalidae have only five visible ventrites (Scholtz Reference Scholtz1990). None of them resemble specimen D with its last ventrite (hypopygidium) broadly fused with the well-sclerotised propygidium, indicating that the propygidium was not fully covered by the elytra. This does not occur in extant representatives of these families. However, if a propygidium is fused with a ventrite, it is fused to the penultimate visible ventrite. If this is the case here, the last visible ventrite might be very short and invisible due to imperfect preservation. A strongly sclerotised propygidium fused to the penultimate ventrite, relatively narrow pygidium and shortened last ventrite are found in Hopliini (Scarabaeidae: Melolonthinae).
Family indet.
(Fig. 14b)
Material. Another specimen from the Brodie collection (NHMUK In. 10050), tentatively identified as Lucanidae or Scarabaeidae by A. G. Kirejtshuk, is not sufficiently preserved for any reliable taxonomic classification (Fig. 14b). It superficially looks like a head of a Dorcus-like stag beetle, but the sclerite to the right of the left ‘mandible' does not support this interpretation since Dorcus does not have a large labrum. It could be a misplaced sclerite, or the fragments could be remains of another beetle or arthropod.
Discussion. The representatives of Scarabaeoidea identified from the Bembridge Marls of the Isle of Wight belong to different trophic guilds. Most Aphodiinae feed on dung, some on rotten organic matter such as carrion or plant debris. Aphodius vectis is likely to indicate the presence of mammals in the area. Aphodiinae are distributed from subartic to tropical regions, hence not indicating a particular climate. Dynastinae are xylophagous, saproxylophagous or feed on roots or dead, rarely living, plant matter (the pollinophagous Cyclocephalini have a less convex pygidium). Distributed worldwide apart from arctic and subarctic regions, they prefer warmer habitats and show their highest diversity and abundance in the tropics. Four of the six Scarabaeoidea from the Bembridge Marls are Dynastinae, indicating a warm climate. Hopliini feed in flowers and serve as pollinators (Micó & Galante Reference Micó and Galante2002; Mayer et al. Reference Mayer, Soka and Picker2006). They are a predominantly Old World taxon with its centre of diversity in the Afrotropics, representatives in the Palaearctic and Indo-Malayan regions and a few species in America. Hopliini avoid cooler climates with only a few species occurring in the northern Palaearctic. The possible hopliine abdomen from the Bembridge Marls indicates an at least moderately warm climate and the presence of flowering plants. The composition of the Scarabaeoidea assemblage does not contradict McCobb's et al. (Reference McCobb, Duncan, Jarzembowski, Stankievicz, Wills and Briggs1998) interpretation of a sub-tropical/tropical palaeoenvironment.
Infraorder Elateriformia Crowson, Reference Crowson1960
Superfamily Scirtoidea Fleming, Reference Fleming1821
Family Scirtidae Fleming, Reference Fleming1821
(=Sinodryopitidae Hong, Reference Hong2002)

Figure 17 Beetles of family Scirtidae. Species of the genus Scirtes. Scirtes calcariferens Kirejtshuk & Ponomarenko, sp. nov.: (a) holotype, NHMUK I. 10083 (1), body, ventral view; length of print 3.1mm; (b) counterpart to holotype (ventral view), NHMUK 10066 (1) and additional specimen (lateral view), NHMUK I. 10066 (2). Scirtes khnzoryani Kirejtshuk & Ponomarenko, sp. nov.; ventral view: (c) holotype, NHMUK In. 25597; (d) paratype, NHMUK I. 8593.

Figure 18 Beetles of family Scirtidae. Species of the genus Scirtes. (a) Scirtes metepisternalis Kirejtshuk & Ponomarenko, sp. nov.: holotype, NHMUK I. 8504, body, ventral view. (b) Scirtes wightensis Kirejtshuk & Ponomarenko, sp. nov.: holotype, IWCMS 2018.42, body, ventral view; length of specimen = 3.8mm. Contacyphon insularis Kirejtshuk & Ponomarenko, sp. nov.: (c) holotype, NHMUK I. 8507, body, ventral view; (d) paratype, NHMUK I. 10306, pterothorax and abdomen, ventral view. Scale in mm.

Figure 19 Beetles of family Scirtidae. Species of the genus Contacyphon. (a) Contacyphon insularis Kirejtshuk & Ponomarenko, sp. nov.: paratype, NHMUK I. 26088, body without head, ventral view. Contacyphon involutus Kirejtshuk & Ponomarenko, sp. nov., ventral view: (b) holotype, NHMUK I. 9466, body without head; (c) additional specimen, NHMUK I. 9131, pterothorax and abdomen. (d) Contacyphon kozlovi Kirejtshuk & Ponomarenko, sp. nov.: holotype, NHMUK I. 9124, body, ventral surface from inside (negative print), with lateral part of left elytron and complete right elytron. Scale in mm.

Figure 20 Suborder Polyphaga, family Scirtidae. (a) Holotype of Scirtes calcariferens Kirejtshuk & Ponomarenko, sp. nov., NHMUK I. 10083 (1), body, ventral view. (b) Additional specimen of of S. calcariferens Kirejtshuk & Ponomarenko, sp. nov., NHMUK I. 10083 (2), body, lateral view. (c) Holotype of Scirtes khnzoryani Kirejtshuk & Ponomarenko, sp. nov., NHMUK In. 25597, body, ventral view. (d) Holotype of Scirtes metepisternalis Kirejtshuk & Ponomarenko, sp. nov., NHMUK I. 8504, body, ventral view. (e) Holotype of Scirtes wightensis Kirejtshuk & Ponomarenko, sp. nov., IWCMS 2018.42, body, ventral view.

Figure 21 Suborder Polyphaga, family Scirtidae, Contacyphon insularis Kirejtshuk & Ponomarenko, sp. nov. (a) Holotype, NHMUK I. 8507, body, ventral view. (b) Paratype, NHMUK I. 10161, body, ventral view. (c) Paratype, NHMUK I. 10306, body, ventral view. (d) Additional specimen, NHMUK I. 10300, body, ventral view.

Figure 22 Suborder Polyphaga, family Scirtidae. (a) Holotype of Contacyphon involutus Kirejtshuk & Ponomarenko, sp. nov., NHMUK I. 9466, body, ventral view. (b) Additional specimen of the same species, NHMUK I. 9131, body, ventral view. (c) Holotype of Contacyphon kozlovi Kirejtshuk & Ponomarenko, sp. nov., NHMUK I. 9124, body, ventral view. (d) Paratype of the same species, NHMUK I. 10291, body, ventral view (with intermixed edges of ventrites and tergites).
Material. A total of 64 specimens (identified by A. G. Kirejtshuk and A. G. Ponomarenko).
Scirtidae, commonly known as marsh beetles, are a comparatively small family. All species of Scirtidae with known bionomy need water for larval development, because the mouthparts of their larvae can only get food by the filtration of algae and other small particles. Adults usually adhere to vegetation near water basins. Therefore, marsh beetles not infrequently use water-filled tree holes, leaf bases, wet soil, rotten logs and similar sources of water. Larvae of some Recent species develop in groundwater as deep as 10m. Adults of some species can occur in many dry localities, including even arid places in deserts. Scirtidae are one of the predominant groups in the Bembridge Marls. This place is the only limestone site among deposits with many findings of this family, although Scirtidae are quite common in the Upper Eocene Baltic amber (Iablokoff-Khnzorian Reference Iablokoff-Khnzoryan1961; Klausnitzer Reference Klausnitzer1976, Reference Klausnitzer2004, Reference Klausnitzer2011; Kubisz Reference Kubisz2000) and also recorded in lowermost Eocene Oise amber (Kirejtshuk & Nel Reference Kirejtshuk and Nel2008, Reference Kirejtshuk and Nel2013) and Lower Eocene Fushun amber (Hong Reference Hong2002). Most specimens examined are nearly complete and of fairly good preservation, sometimes with sclerites of legs and antennae. Most remains originated from homogenous lithology, but some were recovered on the surface of laminae. Specimens of seven species from two genera have been named, while the rest are regarded as Scirtidae incertae sedis. Among the specimens examined, beetles of the genus Scirtes Illiger, Reference Illiger1807 are the most common (previously this genus was recorded by only one species both in Baltic and Fushan amber). Many clear representatives of this family are also present in Karatau (Upper Jurassic) (undescribed in the collection of the Palaeontological Institute of the Russian Academy of Sciences, Moscow) and some other outcrops of a different age. Members of this family also occur in Burmese amber from the boundary of Lower/Upper Cretaceous (Grimaldi et al. Reference Grimaldi, Engel and Nascimbene2002) and Lower Cretaceous Lebanese amber (Kirejtshuk & Azar Reference Kirejtshuk and Azar2008, Reference Kirejtshuk and Azar2013). Some species of Contacyphon Gozis, Reference Gozis1886 were recovered in the lowermost Eocene amber deposits in the Paris Basin (Kirejtshuk & Nel Reference Kirejtshuk and Nel2008). The genera in the studied materials lived mostly in areas with temperate climate, although species of Scirtes are also known from the northern (tropical) territories of Australia. The comparatively large numbers of recognisable specimens and species of this family seems to give some support for the interpretation of wet conditions, not only in places where the sediments were deposited, but also in the surrounding landscape.
Note. H. F. Wickham (Reference Wickham1914b) described one extinct genus Miocyphon in the family Dascillidae from Florissant shales (Upper Eocene), which can be scarcely compared as a close relative with representatives of both Dascillidae and Scirtidae. In contrast to Scirtidae, according to the original description and drawings, it has the comparatively large body (8mm), prosternal segment not rather short, head not clearly declined and subtruncate elytral apices. Expansifemoralis orientalis Hong, Reference Hong2002 (‘Familia incerta sedis': Hong Reference Hong2002) from Fushun amber could be a member of Scirtidae; however, in contrast to the species under description here, it is characterised according to the original description by the longitudinally arranged elytral sculpture and very short head.
Genus Scirtes Illiger, Reference Illiger1807
Type species. Chrysomela haemispherica Linnaeus, Reference Linnaeus1767 (by subsequent designation by Westwood Reference Westwood and Westwood1838), Recent.
=Eohelodes Hong, Reference Hong2002
Type species. Eohelodes orbiculatus Hong, Reference Hong2002 (by monotypy), Fushun amber (Eocene/Lower Eocene), Guchengdzi Formation.
The most characteristic features of Scirtes are the wide and large metafemora as well as long metatibial spurs. They are also somewhat similar to the Recent members of Prionoscirtes Champion, Reference Champion, Godman and Salvin1897, but their metatibial spurs are not as short as those in the latter genus and their metafemora are wider, and the body is not so regularly oval and convex. Species of Scirtes are also somewhat similar to some Recent species of Ora Clark, Reference Clark1866, differing from the latter in the metathoracic coxae meeting along the full length of the median line and their posterior edge conjointly forming a subquadrate plate, which is not in the same plane as the intercoxal process of the abdomen. Sometimes the length of metatibial spurs is used to discriminate these genera; however, this character is quite variable in both genera. This genus was recorded from Baltic amber without description (Klebs Reference Klebs1910; Larsson Reference Larsson1978; Kubisz Reference Kubisz2000) and also from Fushun amber of comparable age (Hong Reference Hong2002) described as Eohelodites orbiculatus Hong, Reference Hong2002 (the latter seems to be a member of Scirtes or Ora with rather short oval body and strongly thickened metafemur: Kirejtshuk & Nel Reference Kirejtshuk and Nel2013). It seems to be very distinct from the species under consideration by its more widely oval body. Sinodryopites ovalifemoralis Hong, Reference Hong2002 (‘Sinodryopitidae': Hong Reference Hong2002) from Fushun amber has also rather large metafemora and very long metatibial spurs, but this species differs from Scirtes species in the very elongate body with comparatively long prosternum, longitudinal costae on elytra, subpectinate antennomeres 3 and 4 (antennomeres 5–11 could be missing), something like those in species of Amplectopus Sharp, Reference Sharp1886; also, the author mentioned four-segmented tarsi, perhaps because tarsomere 4 is very small and hardly visible through the amber.
The species collected from the Bembridge Marls are characterised by comparable body size, but differ mostly in body outline, proportion of head, length of abdominal ventrites, width of the metafemur and shape of the metepisternum. It is rather difficult to compare them with the species described from the Recent fauna, because neontologists use the characters of the dorsal side of the body, some sclerites of appendages and genitalia, which are not usually accessible in fossil specimens obtained from carbonate rocks. Discrimination of specimens of this genus and those of Contacyphon in fossils is possible mostly when at least one metafemur is present.
The genus Scirtes is one of the most abundant coleopterous groups in the Bembridge Marls. There are more than 20 specimens of these beetles belonging to several species. Recent Scirtes has a very wide distribution throughout all continents except Antarctica. More than 150 species of the genus are known from Europe, Asia, South Africa, Australia, North and South America, and sometimes more than one species of the genus occur in the same locality. All specimens placed here in Scirtes resemble some Recent Holarctic representatives of the genus and also some from the Indo-Malayan Region, although the fossil species from the Isle of Wight are more regularly oval. All the new species differ from Scirtes orbiculatus (Hong Reference Hong2002) in having a wide oval body shape, and from Scirtes circumcises Kirejtshuk & Nel, Reference Kirejtshuk and Nel2013 in that the metepisterna is rather strongly narrowing posteriorly, with shorter metaventrite and metafemora rather wide at base and more narrow distally, and also in the somewhat wider base of its pronotum in relation to the posterior part of body. Scirtes metepisternalis sp. nov. seems to have more oval metafemora that are reminiscent of those in S. circumcises, but the former, in addition to other aforementioned diagnostic characters, has a much more slender body.
Scirtes calcariferens Kirejtshuk & Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:824B7C19-847C-4371-BDB8-796B5309CDEC
Derivation of name. The epithet of this new species is formed from the Latin ‘calcar' (spur) and ‘ferens' (bearing, wearing, carrying).
Material. Holotype NHMUK I. 10083 (1) (Figs 17a, 20a): ventral surface from inside (negative print) represented by all body sclerites as well as by right mesotibia and posterior legs and the lateral side of left elytron (together with two additional specimens of this species, an elytron of Curculionoidea and also with remains of Diptera, Hymenoptera and Typha); Insect Limestone, NW Isle of Wight, Brodie Collection. Counterpart to holotype NHMUK I. 10066 (1) (Fig. 17b): dorsal surface of pronotum, pterothorax, lateral part of left elytron and abdomen from inside with some traces of wing venation and also right mesotibia, right metafemur, metatibia with a spur and metatarsomere 1 (with an additional specimen of this species, and also with remains of Diptera, Hymenoptera and Typha); Insect Limestone, NW Isle of Wight, Brodie Collection.
Paratype. NHMUK I. 9119 (2): Middle Oligocene, Bembridge Limestone, NW Isle of Wight, Brodie Collection; ventral surface of head, thorax and basal ventrites from inside and epipleura (negative print) as well as base of left metafemur (together with lateral view of two specimens Curculionidae, a larva of ?Staphylinidae and plenty of Typha remains).
Paratype. NHMUK In. 17437 (2): ventral surface from inside (negative print) with clear outline of most sclerites, including exposing left metafemur (together with three elytra of different specimens of Coleoptera); Insect Limestone, NW Isle of Wight, Smith Collection.
Paratype. NHMUK I. 9977: ventral surface with obscure outline of most sclerites (together with Dipteran wings); Insect Limestone, NW Isle of Wight, Brodie Collection.
Additional specimens. NHMUK In. 25581 (4): dorsum (pronotum, scutellum and elytra) (together with remains of one specimen of ‘Cercyon', ‘Hemisphaera' or ‘Paracymus', elytron of Curculionoidea, an additional pair of small Hydrophilidae, about 2.4mm in length); Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK I. 10083 (2) (Fig. 20b): specimen printed laterally with some traces of an antenna on very slightly declined head and traces of all legs (including protarsus), but it cannot be regarded here with confidence as an additional specimen probably conspecific with the holotype (see the comments to the latter); Insect Limestone, NW Isle of Wight, Brodie Collection. Counterpart NHMUK I. 10066 (2) (Fig. 17b): specimen printed laterally with some traces of an antenna on very slightly declined head and traces of all legs (including protarsus), but it cannot be regarded here with confidence as an additional specimen probably conspecific with the holotype; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 10083 (3): remains represented by dorsal surface of two elytra of the same specimen, but it also cannot be regarded here with confidence as an additional specimen probably conspecific with the holotype; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. In comparison with both of the following new species described here, this new species has its head comparatively narrower, pronotum longer and elytra shorter; its metafemur is somewhat narrower in general and widest at base; it also differs from Scirtes metepisternalis sp. nov. and Scirtes wightensis sp. nov. in the shorter ventrites as well as from Scirtes wightensis sp. nov. in the narrower metepisterna. Finally, the epipleura of this new species are somewhat wider than in Scirtes khnzoryani sp. nov. and Scirtes metepisternalis sp. nov., but distinctly narrower than in Scirtes wightensis sp. nov.
Description. Holotype I. 10066(1)/I. 10083(1): length of specimen 3.1mm, width 2.5mm. Body elongate oval and arcuately narrowed both anteriorly and posteriorly. Integument of underside very finely microreticulated, and that on elytra with very dense, small and rather distinct punctures and finely alutaceous on interspaces between them. Head transverse and rather short, declined ventrally, about twice narrower than pronotum, with moderately large eyes. Pronotum partly deepened into the limestone matrix, transverse and distinctly much more than twice as wide as long, convex at disc, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides; posterior angles almost distinct. Prosternum looking like an extremely narrow stripe before very (sub)contiguous and rather transversely oblique procoxae. Mesoventrite seemingly subcarinate and slightly longer than prosternum, with distinct and comparatively large mesepisterna and mesepimera; mesocoxae (sub)contiguous and somewhat transversely oval and laterally acuminate. Metaventrite with very wide metepisterna (about twice as long as wide, somewhat wider than mesocoxa long), distinct parametacoxal line (parallel to anterior edge of metacoxae), anterior edge of metacoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly, their inner posterior angles forming an angle between them. Elytra about as long as their combined width, rather widened to the middle and seemingly with apices forming a joint arc (with most length at suture); epipleura rather wide at base (about three-fifths as wide as metafemur) and gradually narrowing apically. Abdominal ventrites 1–5 with comparable length. Legs moderately developed. Mesotibia narrow and subparallel-sided (about a third as wide as metepisternum). Metafemur widest at base and about 2.5 times as long as wide, its posterior edge nearly straight. Metatibia rather narrow and very slightly widened apically, with a spur distinctly shorter than metatarsomere 1 and slightly curved.
Counterpart of holotype I. 10066(1): length of print 2.9mm, width apparently 2.5mm. Integument of pronotum and elytra densely, finely and shallowly punctured, and also smoothly alutaceous on interspaces between punctures. Head with appendages invisible. Pronotum convex at disc, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides; posterior angles almost distinct. Metatibia with a spur distinctly shorter than metatarsomere 1 and distinctly straight.
Variation. Paratypes. I. 9977: length 3.2mm, width 2.8mm; differs from holotype in the widest place of body, at abdominal base, not at metaventrite as in other specimens of this species, although the metafemur and configuration of the other sclerites are very similar to those in holotype. I. 17437: length 3.5mm, width 2.8mm; showing all visible sclerites (including metepisterna) as those in the holotype, but with missing trace of legs. I 9119: width 2.3mm; epipleura somewhat narrower than mesocoxa long. Additional specimens. In. 25581: length of print 3.0mm, width 2.5mm; body outline as that in the holotype; dorsal surface quite smooth; scutellum about 1.5 as wide as long, subtriangular and widely rounded at apex. In. 10083 (2) (lateral view): length 3.1mm, height 0.6mm; body moderately convex dorsally and seemingly very slightly ventrally; head not strongly declined ventrally; antennae consisting of subcylindrical segments (seven of them visible); profemur about a third as wide as metafemur of the holotype; protibia slightly shorter than profemur and about as narrow as meso- and metatibiae of the holotype; protarsus somewhat shorter than protibia with tarsomeres 3 and 4 lobed. Counterpart. I. 10066 (2) (lateral view): length 3.2mm, height 0.6mm. I. 10083 (3): elytra distinctly acuminate at apex and with distinct adsutural lines along the whole length.
Scirtes khnzoryani Kirejtshuk & Ponomarenko, sp. nov.
(Fig. 17c, d, 20c)
Zoobank ID. urn:lsid:zoobank.org:act:C62C3032-5DAE-47CF-AB23-F2F2630B80C8
Derivation of name. The epithet of this new species is devoted to the outstanding coleopterist who first started to describe fossil Scirtidae (Iablokoff-Khnzoryan Reference Iablokoff-Khnzoryan1961).
Material. Holotype NHMUK In. 25597 (Figs 17c, 20c): inner side of almost all ventral sclerites (including epipleura at base), and also with sides of elytral apices, more or less clear outline of both (left and right) meso- and metafemora; Insect Limestone, NW Isle of Wight, Hooley Collection.
Paratypes. NHMUK I. 8593 (Fig. 17d): ventral surface from inside represented by most body sclerites with not quite clear outlines, but with distinct configuration of left posterior legs (with a long spur, but without metatarsus) and also with base of left epipleuron, part of pronotum and right elytron (positive print); Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 10340: ventral surface from inside represented by all body sclerites with clear outlines of coxae and both metafemora; Insect Limestone, Isle of Wight, Hooley Collection.
Diagnosis. This new species has the body more narrowing both anteriorly and posteriorly than in Scirtes calcariferens sp. nov.; other differences of it from the latter are mentioned in the diagnosis of S. calcariferens sp. nov. (see above). From Scirtes metepisternalis sp. nov. and Scirtes wightensis sp. nov. it differs in the longer ventrites and metafemur seemingly less narrowing apically; from S. metepisternalis sp. nov. it differs also in the configuration of anterior edge of its pronotum and narrower metepisterna; from S. wightensis sp. nov. it differs also in the markedly more slender body and narrower epipleura. The print of the holotype also has a trace of the median carina on prosternum. The mesocoxae of this new species are (sub)contiguous like those in S. wightensis sp. nov., but in contrast to those in both S. calcariferens sp. nov. and S. metepisternalis sp. nov.
Description. Holotype In. 25597: length of specimen 3.0mm, width 2.0mm. Body elongate oval and arcuately narrowed both anteriorly and posteriorly. Head transverse and rather short, declined ventrally, less than twice narrower than pronotum, with moderately large eyes. Pronotum transverse and markedly more than twice as wide as long, convex at disc, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides from the base; posterior angles almost distinct. Prosternum carinate in the middle and looking like a very narrow stripe before subcontiguous and rather transversely oblique procoxae. Mesoventrite seemingly subcarinate and slightly longer than prosternum, with distinct and comparatively large mesepisterna and mesepimera; mesocoxae subcontiguous and somewhat transversely oval and laterally acuminate. Metathorax with very wide metepisterna, distinct parametacoxal lines, anterior edge of metacoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly. Elytra at least 1 and 1/15 as long as combined width (probably much longer), rather widened at the middle and seemingly with most length at suture, epipleura rather wide at base (about two-fifths as wide as metafemora) and gradually narrowing apically. Abdominal ventrites 1–4 with comparable length, although hypopygidium somewhat longer than previous ventrites. Legs seemingly moderately developed. Mesofemur moderately narrow (narrower than mesocoxae long); metafemora widest distally from base (but before the middle) and about 2.5 times as long as wide.
Variations (paratypes). Length of body 2.5–3.6mm. Paratype I. 8593: length of specimen 3.1mm, width 2.1mm; configuration of body somewhat more slender than in the holotype, although most sclerites, including head, metepisterna and metafemora with outlines as those in the holotype; metatibia narrow and markedly longer than metafemur, its spur slightly curved and about a quarter as long as metatibia. Paratype I. 10340: length of specimen 2.5mm, width 2.1mm; configuration of body and most sclerites, including metepisterum, as that in the holotype.
Scirtes metepisternalis Kirejtshuk & Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:0089263D-B178-4D26-AD0D-2E2C3784139C
Derivation of name. The epithet of this new species refers to the peculiarly wide metepisterna in this new species.
Material. Holotype NHMUK I. 8504 (1) (Figs 18a, 20d): ventral surface from inside represented by all body sclerites, as well as by base of left epipleuron, left metafemur and side of right elytron (together with elytron of Cantharidae); Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species is very distinct from others described here in the narrow head and seemingly narrower epipleura. However, its metafemur is much wider than in other congeners from the Bembridge Marls and it has comparatively long ventrites, somewhat like those in Scirtes wightensis sp. nov., but, in contrast to the latter Scirtes metepisternalis sp. nov., is markedly more slender. Finally, the metepisterna of this new species appears to have the shortest inner side among the congeners. See also the diagnoses of two previous species.
Description. Holotype I. 8504: length of specimen 3.2mm, width 2.0mm. Body elongate oval and arcuately narrowed both anteriorly and posteriorly. Integument of elytra with very fine, dense and shallow punctures and alutaceous interspaces between them. Head transverse and rather short, declined ventrally, less than twice narrower than pronotum, with moderately large eyes. Pronotum transverse and about twice as wide as long, convex at disc, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides; posterior angles distinct. Prosternum rather obscure in the middle and looking like a very narrow stripe before subcontiguous, and rather transversely oblique procoxae. Mesoventrite with distinct median depression and rather longer than prosternum, with distinct and comparatively large mesepisterna and mesepimera; mesocoxae subcontiguous and somewhat transversely oval and laterally acuminate. Metathorax with rather wide metepisterna, distinct parametacoxal lines, anterior edge of metacoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly. Elytra seemingly nearly one and a quarter as long as combined width, rather widened to the middle and seemingly with most length at suture; epipleura rather wide at base and gradually narrowing apically (about a quarter as wide as metafemur). Abdominal ventrites 1–4 with comparable length, but hypopygidium markedly shorter. Legs moderately developed. Metafemur extremely wide and widest at the middle and almost twice as long as wide.
Scirtes wightensis Kirejtshuk & Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:55F71417-3E5D-438F-925A-1AE9B649B0C4
Derivation of name. The name of this new species is named from the toponym of the island of its origin.
Material. Holotype IWCMS 2018.42 (Figs 18b, 20e): underside represented by all body sclerites with impression of right metafemur; Insect Limestone, NW Isle of Wight.
Additional specimen. NHMUK I. 10199 (1): underside represented by many body sclerites, most of them with obscure outline; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from all fossil species of the genus from the Isle of Wight fauna in the very small head. Also, it is very distinct from others described here in the combination of very wide metafemur, comparatively less wide metepisterna and rather wide epipleura. In contrast to the previous species with wide metafemur and comparatively long ventrites, Scirtes wightensis sp. nov. has a wider body and much narrower metepisterna and wider epipleura, while in contrast to the first two new species described here with comparatively narrow metepisterna and comparatively wide epipleura, S. wightensis sp. nov. is characterised by a rather wide metafemur and longer ventrites.
Description. Holotype IWCMS 2018.42: length of specimen 3.8mm, width 2.2mm. Body oval and arcuately narrowed both anteriorly and posteriorly. Head transverse and rather short, declined ventrally, about three times wider than pronotum, with moderately large eyes. Pronotum transverse and very short, convex at disc, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides; posterior angles seemingly distinct. Prosternum rather obscure in the middle and looking like a very narrow stripe before (sub)contiguous, and rather transversely oblique procoxae. Mesoventrite obscure in the middle and almost as long as prosternum, with distinct and comparatively large mesepisterna and mesepimera; mesocoxae contiguous and somewhat transversely oval and laterally acuminate. Metathorax with moderately wide metepisternum, distinct parametacoxal lines (parallel to anterior edge of metacoxae), anterior edge of procoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly. Elytra seemingly nearly one and sixth as long as combined width, rather widened to the middle and seemingly with most length at suture, epipleura rather wide at base and gradually narrowing apically (about two-fifths as wide as metafemur). Abdominal ventrite 2 and hypopygidium somewhat longer than the other ventrites. Legs moderately developed. Metafemur widest before the middle, almost twice as long as wide.
Variation. Additional specimen I. 10199 has the general outline as that in the holotype, although most sclerites of it are obscure in outline.
Scirtes spp.
Material. NHMUK I. 10088: part of prothorax dorsally, part of posterior wing, abdomen from inside and right elytron dorsally; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK In. 17239: lateral view with large metafemur; Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK In. 25641 (1): body represented by a mixture of negative and positive impressions, showing base of elytra dorsally with trace of adsutural lines, pronotum dorsally, posterior part of metaventrite and abdomen from inside, right posterior leg partly exposed, with large metafemur (but without clear outline) (together with the holotype of Themus polyaki sp. nov. and two other coleopterous remains); Insect Limestone, NW Isle of Wight, Hooley Collection.
Genus Contacyphon Gozis, Reference Gozis1886
Type species. Cantharis variabilis Thunberg, Reference Thunberg1787 (original designation by Gozis Reference Gozis1886), Recent.
This genus is very abundant in both Recent fauna and the fossil records. About 100 Recent species are found throughout different zoogeographical regions, with most of them recorded in the Northern Hemisphere. As in the case with Scirtes species, specimens collected in the Bembridge Marls are difficult to compare with most descriptions, as characters used in them are taken usually from the dorsal side of body, some sclerites of appendages and genitalia, which are not accessible in fossil specimens obtained from carbonate rocks. Nevertheless, a possible comparison was made on the basis of specimens deposited in St Petersburg, Moscow, London and Paris collections. Iablokoff-Khnzoryan (Reference Iablokoff-Khnzoryan1961) described three species of Contacyphon from Baltic amber. All of these fossil species are markedly more slender than the new species from the Bembridge Marls and their metepisterna seem to be narrower in general. Klausnitzer (Reference Klausnitzer2004, Reference Klausnitzer2011) and Heuss (Reference Heuss2008) described five species of Contacyphon from Baltic amber with an exposed apical part of the aedeagus, which have various structures of the lateral lobes of tegmen. Unfortunately, these authors did not mention any character allowing comparison of these members of the genus with other congeners, even with the congeners described by Iablokoff-Khnzoryan (Reference Iablokoff-Khnzoryan1961) from the same fossil resource. Therefore, at this stage the species described here as new cannot be compared with Recent and fossil species of the genus, although most of these species from Baltic amber have a metepisterna less narrowing posteriorly than those in the new congeners from the Bembridge Marls. Kirejtshuk & Nel (Reference Kirejtshuk and Nel2008) described two species as members of Contacyphon from the lowermost French amber of Oise also having their metepisterna less narrowing posteriorly than those in the new species. Hong (Reference Hong2002) published the picture of the species of this genus from Fushun amber without species epithet, which differs from the species described here in having a much more oval body. However, this species from Fushun cannot belong to the genus Contacyphon at all because of its strongly declined metacoxae (not a frequent character among Recent congeners). Finally, species of this genus are very similar to those of Hydrocyphon Redtenbacher, Reference Redtenbacher1858, which could be diagnosed in fossil materials mostly because of the different shape of pronotum. However, the prothorax in most remains examined is within the limestone matrix and, therefore, its shape is often not clearly visible.
Contacyphon insularis Kirejtshuk & Ponomarenko, sp. nov.
(Figs 18c, d, 19a, 21)
Zoobank ID. urn:lsid:zoobank.org:act:99477579-FE64-4320-9D0E-AD9D9CB0D849
Derivation of name. The epithet of this new species refers to the isolation of its type locality from the European continent.
Material. Holotype NHMUK I. 8507 (Figs 18c, 21a): ventral surface from inside represented by all body sclerites, right profemur and left metafemur, and also sides of elytra (with an abdomen of Staphylinidae); Insect Limestone, NW Isle of Wight, Brodie Collection.
Paratypes. NHMUK I. 10306 (Figs 18d, 21c): underside represented by pterothorax, abdomen and epipleura, as well as by left base of elytra and scutellum; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK In. 26088 (Fig. 19a): inner view of underside with outline of right meso- and metafemora (together with Oecophylla sp. and Dipteran wing); Insect Limestone, NW Isle of Wight, Hooley Collection.
Additional specimens. NHMUK I. 10161 (Fig. 21b): ventral surface from inside represented by many sclerites, but partly not clearly outlined (particularly at sides), right mesofemur visible as a deep cavity at outer edge of mesocoxa and a trace of the right metafemur; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 10300 (Fig. 21d): ventral surface from inside represented by many sclerites with obscure outline of pterothoracic sclerites; at sides, dorsal surface of elytra well exposed; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from Contacyphon involutus sp. nov. in the more oval and more robust body shape, distinctly separated pro- and mesocoxae, shorter elytra with more arcuate sides and distinctly acuminate apex, wider metepisterna and distinctly carinate prosternum (in the paratype, prosternal process seemingly narrower and its median carina not expressed). It also differs from Contacyphon kozlovi sp. nov. in the more oval and more robust body shape, larger eyes, distinctly separated procoxae and narrower metepisterna.
Contacyphon insularis sp. nov. is distinct from many species of the genus also in the wider epipleura and rather long ventrite 1. It can be compared with the Holarctic Contacyphon variabilis (Thunberg, Reference Thunberg1787), although C. insularis sp. nov. has the pronotum narrowing much more anteriorly and wider metepisterna. This new species also can be compared with Contacyphon doctus Lea, Reference Lea1910 (type specimen examined) and Contacyphon genalis Sharp, Reference Sharp1878 (type specimen examined), but it differs from the former in the wider and more oval and not so oblique metacoxae; and from the latter in the wider pronotum, wider anterior part of metepisterna and longer abdominal ventrite 1.
Description. Holotype: length of print 2.6mm, width 1.6mm. Body oval and arcuately narrowed both anteriorly and posteriorly; rather light reddish in colouration. Integument with indistinct and very fine puncturation, densely microreticulated on thorax and abdomen, as well as smoothly and densely alutaceous on elytra. Head transverse and very short, declined ventrally, about three-fifths as wide as pronotum, with moderately large eyes. Anterior edge of head and mouthparts invisible. Pronotum transverse and rather deep inside the limestone matrix, seemingly more than four times as wide as long, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides from posterior angles; posterior angles almost distinct. Prosternum comparatively long and medially sharply carinate; procoxae narrowly separated and rather transversely oblique. Mesoventrite obscure in the middle and distinctly shorter than prosternum, with partly distinct mesepisterna and mesepimera; mesocoxae very narrowly separated (about twice as long as wide at its anterior edge), somewhat transversely oval and laterally acuminate. Metaventrite with rather wide metepisterna, slight parametacoxal line, anterior edge of metacoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly. Elytra seemingly nearly 1.5 times as long as their combined width, slightly widened to the middle and seemingly with most length at suture, epipleura rather narrow at base and gradually narrowing apically (about half as wide as mesocoxa long). Abdominal ventrites 1–5 with comparable length. Legs, perhaps, moderately developed. Profemur comparatively wide (about as wide as mesocoxae long); metafemur not wider than profemur and more or less elliptical, more than four times as long as wide.
Variations. Paratype I. 10306: length of print 2.8mm (without prothorax and head), width 2.1mm. The paratype is somewhat larger and appears more robust, with epipleura somewhat narrower, abdominal ventrites 2–4 and hypopygidium slightly longer than in the holotype. Scutellum subtriangular with widely rounded apex. Integument of elytral base with small dense and shallow punctures. Paratype In. 26088: length of print 2.2mm, width 1.3mm. The specimen appears smaller and somewhat more slender, with metepisterna less narrowing posteriorly. Meso- and metafemora are slightly narrower than length of mesocoxae. Additional specimens (I. 10300 and I. 10161): length of print 2.0–2.1mm, width 1.5mm. These specimens are characterised by comparatively bad preservation, although apparently they belong to the genus Contacyphon rather than Scirtes, and they could be conspecific with the type species of Contacyphon insularis sp. nov., although they are certainly smaller and much more oval.
Contacyphon involutus Kirejtshuk & Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:EFE66094-44EF-45BA-9674-C86E46CF6A68
Derivation of name. Epithet of this new species means ‘dark', ‘tangled', ‘intricate', ‘involved', ‘difficult'.
Material. Holotype NHMUK I. 9466 (Figs 19b, 22a): ventral surface from inside (negative print), represented by almost all sclerites, except sides of metathorax and abdomen and apex of abdomen. The appendages are represented by left profemur, part of left mesofemur and mesotibia as well as left metafemur; Insect Limestone, NW Isle of Wight, Brodie Collection.
Paratype. NHMUK In. 17435: ventral surface from outside, represented by most sclerites of metaventrite and abdomen, and also head, outline of pronotum and right mesofemur (together with an elytron of Curculionidae); Insect Limestone, NW Isle of Wight, Smith Collection.
Additional specimen. NHMUK I. 9131 (Figs 19c, 22b): underside represented only by a part of mesoventrite, metaventrite with left metafemur and metatibia and abdomen; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from Contacyphon insularis sp. nov. in the more slender body shape, subcontiguous pro- and mesocoxae, longer elytra with less arcuate sides, narrower metepisterna. It also differs from Contacyphon kozlovi sp. nov. in the more slender body shape and subcontiguous mesocoxae. See also the diagnosis of previous species.
Description. Holotype I. 9466: length of specimen 4.6mm, width apparently 2.4mm. Body elongate oval and arcuately narrowed both anteriorly and posteriorly. Head transverse and fairly short, strongly declined ventrally. Anterior edge of head and mouthparts invisible. Pronotum transverse, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as a continuation of lateral sides from posterior angles; posterior angles distinct. Prosternum seemingly rather short; procoxae (sub)contiguous and rather transversely oblique. Mesoventrite obscure in the middle and very short, with obscure mesepisterna and mesepimera; mesocoxae (sub)contiguous, somewhat transversely oval and laterally acuminate. Metathorax with rather wide metepisterna (about 2.5 times as long as wide at its anterior edge), parametacoxal line scarcely visible, anterior edge of metacoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly. Elytra invisible. Abdominal ventrites 1–4 of comparable length, hypopygidium apparently somewhat longer. Legs moderately developed. Pro- and mesofemora somewhat narrower than mesocoxa is long; profemur about three times as long as wide. Metafemur slightly wider than pro- and mesofemora, apparently about 3.5 times as long as wide. Mesotibia rather narrow, about two fifths as wide as mesofemur.
Variations. Paratype In. 17435: length of print 3.2mm, width 1.6mm. Markedly smaller than the holotype and with hypopygidium somewhat shorter than each of 2–4 abdominal ventrites. Additional specimen I. 9131: differs from the holotype in the distinctly more oval (not obliquely elongate) mesocoxae, somewhat longer median part of femoral plate of metacoxae and wider metepisterna.
(?) Contacyphon kozlovi Kirejtshuk & Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:D2916CA6-E43E-4078-9A33-24E73F72F854
Derivation of name. The epithet of this new species is devoted to M. A. Kozlov, an outstanding entomologist and participant of the project on Bembridge Insect Beds, whose sudden death prevented him from continuing his valuable contribution to the Isle of Wight hymenopterous fauna (see Ross Reference Ross2014).
Material. Holotype NHMUK I. 9124 (Figs 19d, 22c): ventral surface from inside (negative print), represented by all sclerites, the lateral part of left elytron and complete right elytron; Insect Limestone, NW Isle of Wight, Brodie Collection.
Paratype. NHMUK I. 10291 (Fig. 22d); dorsum from inside, with unclear outlines of most sclerites, part of mesofemur and mesotibia; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from Contacyphon insularis sp. nov. in the somewhat more slender body shape, smaller eyes, subcontiguous procoxae and wider metepisterna. It also differs from Contacyphon involutus sp. nov. in the more robust body shape and separated mesocoxae. Among other characters, this new species is characterised by well-expressed copper lustre of its integument. This new species, in contrast to the previous one, has some untypical characters of the genus (comparatively long head with small eyes, subtruncate elytral apices and wider tibiae) and, therefore, its attribution to it is regarded here as questionable. As to elytral apices, it can be admitted that they could be rolled, while fossilised, because the integument of representatives of this family is not infrequently rather soft (poorly sclerotised). The same reason can be supposed, but with a smaller probability, for comparatively wider tibia. The head shape and comparative proportion of eyes are rather variable inside the genus. Therefore, the preliminary attribution of this new species to the genus Contacyphon (or allied genera) seems to be reasonable. See also the diagnosis of C. insularis sp. nov.
Description. Holotype I. 9124: length of specimen 2.6mm, width 1.5mm. Body elongate oval and arcuately narrowed both anteriorly and posteriorly; with strong copper lustre. Integument with dense, fine and fairly distinct punctures and alutaceous on interspaces between them. Head transverse and not quite short, strongly declined ventrally, with comparatively small eyes. Anterior edge of head and mouthparts invisible. Pronotum transverse, widest at base, arcuately narrowed anteriorly, anterior margin forming an arc as continuation of lateral sides from posterior angles; posterior angles almost distinct. Prosternum rather short; procoxae contiguous and rather transversely oblique. Mesoventrite obscure in the middle and very short, with distinct mesepisterna and mesepimera; mesocoxae narrowly separated, somewhat transversely oval and laterally acuminate. Metathorax with rather wide and subtriangular metepisterna (about twice as long as wide at widest part), parametacoxal line distinct, anterior edge of metacoxae slightly oblique; metacoxae with distinct femoral plate moderately expanded medially and narrowing exteriorly. Elytra seemingly nearly 1 and 1/4 as long as combined width, slightly widened to the middle and seemingly subtruncate at apex, with distinct subsutural line and steeply sloping laterally. Abdominal ventrites comparable in length, hypopygidium widely rounded at posterior edge. Legs invisible.
Variation. Paratype I. 10191: length at least 4.0mm (3.6mm within anterior edge of pronotum and elytral apices), width 2.2mm. Underside invisible, but dorsal thoracic sclerites (lying under elytra) and seven right abdominal laterosternites and tergites as well as right mesofemur and mesotibia partly visible. Scutellum subtriangular and with rounded apex. Tergites all of similar length.
Contacyphon sp.
Material. NHMUK I. 10303: ventral surface from inside (negative print), but the left lateral part of abdomen covered by the elytron and middle of pterothorax covered by the positive remains of the upper surface (concealing the characters important for species discrimination). Also visible are the profemora and basal parts of protibiae, as well as apex of left metafemur and base of metatibia, showing that this species belong to Contacyphon, but not to Scirtes; Insect Limestone, Isle of Wight, Brodie Collection.
Gen. & spp. indet.
(could be Scirtes or Contacyphon species described here)
Material. NHMUK ‘In. 26147': ventral surface of pterothorax and abdomen from inside (together with additional specimen Metacoxites ventritalis gen. & sp. nov. and remains of Curculionoidea); Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK In. 43418: upper surface of two elytra of the same specimen with distinct adsutural lines; Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK I. 9650: pterothorax and abdomen from outside with visible metepisterna and contiguous mesocoxae, and also scutellum, epipleura and apex of right elytron; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK In. 17432: inner side of pterothorax and abdomen with contiguous mesocoxae and prothoracic segment with head deep inside the limestone matrix, also left elytron with a distinct adsutural line and a part of right elytron; Insect Limestone, NW Isle of Wight, Smith Collection.
NHMUK I. 9109 (two specimens): (1) part of metacoxa and abdomen; (2) mixture of negative and positive prints, including scutellum, base of elytra, right epipleuron, and abdominal ventrites; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK In. 26156: dorsal surface of two elytra of the same specimen with distinct adsutural lines and transverse section of thorax; Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK I. 10167: dorsal surface of right elytron with distinct adsutural line and unusually sparse puncturation (together with four elytra of different specimens, including one specimen of Donaciinae); Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 10376 and NHMUK I. 9426 (part and counterpart): underside of body (together with two specimens of Staphylinidae); Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK In. 26118: abdomen with right metacoxa; Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK ‘I. 10090': head anteroventrally and side of body with a part of right elytron; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK In. 25686 (1), (2): two elytra of Scirtidae (together with two elytra of Plateumaris sp., 14 elytra incertae sedis, one metathorax incertae sedis and one abdomen incertae sedis); Insect Limestone, NW Isle of Wight, Hooley Collection.
NHMUK I. 8502: dorsal surface of body without elytral apices where abdominal segmentation exposed (together with one dorsum of coleopterous body incertae sedis of comparable size with excavation showing the transverse metacoxae without femoral plates, but without clear outline of other sclerites); Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 8705 and In. 17420 (part and counterpart): mixture of dorsal and ventral sclerites, without a part of pronotum and head, but with traces of oblique metacoxae with femoral plates; Insect Limestone, NW Isle of Wight, Brodie and Smith Collection.
NHMUK I. 8501: pterothorax and abdominal tergites dorsally and also a part of posterior wing, part of pronotum and sides of elytra; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 9608 (1): mostly left side of ventral surface with a deep cavity from mesofemur; Insect Limestone, NW Isle of Wight, Brodie Collection.
IWCMS 2018.43: underside represented by the left half of pterothorax and nearly entire abdomen, most sclerites are clearly outlined; Insect Limestone, Isle of Wight.
Discussion. The rather numerous specimens of this family (marsh beetles) in the Bembridge Marls clearly show that the places of deposition were connected to a swamp or shallow freshwater environment. The great number of Scirtidae found in the course of these studies can be compared with the proportion of this family in the Baltic amber fauna, while that in the amber of Oise is clearly smaller. The high number of Scirtidae in Baltic amber can be explained by the coincidence of the time of resin-production by coniferous trees and the time of flying of representatives of this family, which, like some Recent species, could have fed on spores from the male cones of Pinaceae. However, in the case of the Bembridge Marls such an explanation is not possible. A more probable explanation is that the group were entering sites of deposition near the places where they lived.
Family Eucinetidae Lacordaire, Reference Lacordaire1857

Figure 23 Beetles of families Eucinetidae, Elateridae, Eucnemidae, Chrysomelidae. (a) Eucinetes nikolaevae Kirejtshuk & Ponomarenko, sp. nov. (Eucinetidae): holotype, NHMUK In. 17286, body, ventral view; length of print = 2.8mm. (b) Macropunctum rossi Alexeev, sp. nov. (Elateridae): holotype, NHMUK I. 10085, body, ventral view; length of specimen = 7.7mm. (c) Plateumaris (Plateumaris) wightensis Kurochkin & Kirejtshuk, sp. nov. (Chrysomelidae): NHMUK In. 25605, elytron, dorsal view; length of print = 5.9mm. (d) Eucnemidae gen. & sp. indet.: NHMUK In. 17209, body, ventral view; length of print = 2.3mm.

Figure 24 Suborder Polyphaga, family Eucinetidae: holotype of Eucinetes nikolaevae Kirejtshuk & Ponomarenko, sp. nov., NHMUK In. 17286, body, ventral view.
Material. One specimen (identified by A. G. Kirejtshuk and A. G. Ponomarenko).
Plate-thigh beetles or Eucinetidae include only nine genera in the Recent fauna and are small- and medium-sized beetles (usually 0.8–4.0mm), more or less oval and rather convex dorsally. This family can be easily recognised in fossils due to distinct forms of the imaginal metathorax, meso- and metacoxae. The adults have the quite characteristic, strongly oblique and moderately large mesocoxae, very large metacoxae, very short prothoracic segment and very long elytra more or less acuminate at apices, head turned ventrally and simple tarsomeres (although sometimes tarsomeres are rather thick). Recent species are distributed worldwide (except for pre-polar areas); however, they occur in rather low density. Most of the species develop and feed on detritus and subcortical space with fungi. They are known to be associated with spores of slime moulds and fruiting bodies of some groups of Basidiomycetes. The group is a member of the rather archaic superfamily Scirtoidea, all families of which include a very small number of species in the Recent fauna and are poorly represented as fossils. This family is recorded in Upper Eocene Baltic amber (Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984) and the boundary of Lower/Upper Cretaceous Burmese amber (Rasnitzyn & Ross Reference Rasnitsyn and Ross2000; Grimaldi et al. Reference Grimaldi, Engel and Nascimbene2002). It seemed to have a rather early origin, and probably because of peculiarities of bionomy of its members (strict and close association with trees) it had a lower chance of entering depositional environments or it has never been abundant in both Recent and past faunas. Hong (Reference Hong1995) described two species as members of this family from the Lower Cretaceous of Xiangfanggou (China), although they need to be re-examined for confirmation of their attribution.
Genus Eucinetes Germar, Reference Germar1818
Type species. Scaphidium haemorrhoidale Germar, Reference Germar1818, Recent.
The specimen examined has no clear characters for generic discrimination, including broken inner part of the metacoxae. Nevertheless, it should be regarded as a member of the Eucinetes rather than any other genus of the family, because this specimen has the rather wide anterior edge of metacoxae and five visible abdominal ventrites, the ultimate ventrite (hypopygidium) is usually rather small. However, the metacoxae of the fossil examined have an outline very similar to those in Bisaya Reitter, Reference Reitter1884a, which is characterised by a very small body, widely truncate apex of metacoxae and five abdominal ventrites. It differs from Jentozkus Vit, Reference Vit1977 and Tohlezkus Vit, Reference Vit1977 in the much larger body, large metepisterna, very narrow epipleura in distal two-thirds and short median part of metacoxae. It differs from Eucilodes Vit, Reference Vit1985 and Euscaphurus Casey, Reference Casey1885 in the very short mesocoxae and rather short median part of metacoxae. In addition, the species of Euscaphurus also have a rather small body.
Eucinetes nikolaevae Kirejtshuk & Ponomarenko, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:AF6A1CDD-89CD-4722-B4F5-DD4584535CE3
Derivation of name. The epithet of this new species is devoted to Svetlana Vitalyevna Nikolaeva, a palaeontologist from the Palaeontological Institute of the Russian Academy of Sciences (Moscow) and International Commission of Zoological Nomenclature (London), who, in many ways, kindly assisted the participants of the INTAS project and many other entomologists and palaeontologists visiting London.
Material. Holotype In. 17286 (Figs 23a, 24): underside with strongly declined head, more or less clear thorax, ventrites, mesofemora and epipleura; Insect Limestone, NW Isle of Wight, Smith Collection.
Diagnosis. This new species is rather distinct in the shape of metacoxae and rather narrow epipleura in distal two-thirds. In particular, in contrast to the new species, the Recent species of the genus have the distal apices of metafemoral plates wider and lateral end of them much longer, and metepisterna markedly shorter. It also differs from other Recent species of Eucinetes and Jentozkus in having acute median parts of metacoxae, while in most species median apices of metacoxae are widely transverse to more or less emarginate.
Description. Holotype In. 17286: length of specimen 2.8mm, width 1.6mm, length of elytra 2.6mm. Body subelliptic, apparently rather convex dorsally and moderately convex ventrally; light brown in colouration. Thoracic lower sclerites, metacoxae and abdominal ventrites with smoothed sculpture as well as with coarsely and sparsely diffuse punctures. Head of usual shape (somewhat longer than wide at eyes), moderately elongate, rounded at sides, with antennal insertions close to anterior edge of frons, convex surface of frons rather narrow and slightly widened before subtruncate anterior edge, not reaching the posterior edge of mesoventrite. Labrum apparently subtruncate and nearly acute at apex, scarcely wider than long. Pronotum about three times as wide as long, apparently convex, widest at base and strongly narrowing to gently and widely arcuate anterior edge; anterior angles not expressed. Elytra about 1.6 times as long as their combined width; slightly narrowed at base, widely rounded along the middle and somewhat wider than pronotum, apparently complete and covering apex of abdomen. Mesoventrite moderately long. Metaventrite with parametacoxal lines. Metepisterna rather long, more than four times as long as wide. Mesocoxae moderately narrowly separated. Metacoxae strongly oblique medially, rather large, rather widely separated anteriorly and contiguous posteriorly, femoral plates rather large, but rather narrowing at rounded apices. Abdomen with only five visible ventrites, ventrite 1 about 1.5 times as long as ventrite 2, ventrites 3–5 subequal in length and each of them about half as long as ventrite 1. Mesofemur about 2.5 times as long as wide and reaching outer edge of epipleura. Mesotibia rather short and dilated, somewhat narrower than protibia.
Discussion. As this family is closely connected with rotten wood, finding it in the Bembridge Marls was possible due to it flying from forest adjacent to the swamps or shallow water where deposition was proceeding. Today, the specimens of this group sometimes occur in trunks and litter at water margins and this new species could have lived in wet wood near water basins.
Infraorder Elateriformia Crowson, Reference Crowson1960
Superfamily Elateroidea Leach, Reference Leach1815
Family Buprestidae Leach, Reference Leach1815
Material. A total of 14 specimens (identified by A. V. Alexeev, A. G. Kirejtshuk, A. G. Ponomarenko and A. S. Kurochkin).
This comparatively large and diverse family consists of species characterised by medium and large body length (2.2–60.0mm), usually somewhat flattened body and complete elytra, sinuate lateral edge of elytral base, short and transverse (‘vertical') head with mouthparts oriented ventrally, usually short and narrow antennae, rather separated and small pro- and mesocoxae, prosternal process nearly reaching anterior edge of metaventrite, metaventrite with a curved transverse suture, first two ventrites partly fused and rather widened tarsi. Bionomy of its members is also fairly diverse. Adults of Recent species feed primarily on the foliage of their host plants, but floricolous habits are known in many tribes of Buprestidae or jewel beetles. Larvae are mostly wood borers, attacking living or dead trees or shrubs. They may attack various parts of the host plant and hosts include gymnosperms, monocots and dicots. Larvae of Julodinae and Schizopodinae occur in the soil and probably feed on roots. Adults are usually good flyers and can cross considerable distances. Jewel beetles spread through all the world and are particularly abundant in areas with warm climate. They are known as fossils from the beginning of the Middle Jurassic; however, they are especially numerous in the Cretaceous and Palaeogene, where their remains compose a significant part of the oryctocenoses. The Mesozoic and Caenozoic faunas are rather different in generic and subfamily composition (Hong Reference Hong1982; Whalley & Jarzembowski Reference Whalley and Jarzembowski1985; Alexeev Reference Alexeev1994, Reference Alexeev2000; Pan et al. Reference Pan, Chang, Ren and Shih2011; etc.). Remains of this family in the European Middle Eocene represent more than 20 genera, although in the Oligocene they became less numerous (Pongracz Reference Pongracz1935; Piton Reference Piton1940; Haupt Reference Haupt1950, Reference Haupt1956; Spahr Reference Spahr1981; Weidlich Reference Weidlich1987; Hörnschemeyer & Wedmann Reference Hörnschemeyer and Wedmann1994; etc.).
Material. Among 12 specimens, including isolated elytra and one prothorax and paired elytra, 7–10 species are present: two Agrilinae Laporte, Reference Laporte1835 (including a member of the genus Agrilium Westwood, Reference Westwood1854: NHMUK I. 8626); four Buprestinae (including a member of the tribe Melanophilini Bedel, Reference Bedel1921: NHMUK I. 8772 and a member of the genus Buprestis Linnaeus, Reference Linnaeus1758: NHMUK In. 25603); two medium-sized jewel beetles; and one small-sized Anthaxia Eschscholtz, Reference Eschscholtz1829. Assemblage examined is temperate, but not tropical. There is no typically Eocene form from the Dicercini Gistel, Reference Gistel1848 in the specimens examined.
Family Elateridae Leach, Reference Leach1815
(Figs 23b, c, 25)

Figure 25 Suborder Polyphaga, family Elateridae: holotype of Macropunctum rossi Alexeev, sp. nov., NHMUK I. 10085, body, ventral view.
Material. A total of 27 specimens (identified by A. V. Alexeev, A. G. Kirejtshuk, A. G. Ponomarenko and A. S. Kurochkin).
Elateridae, or click beetles, are a rather large family characterised by the developed promesothoracic clicking mechanism, with the prosternal process ending with an acute apex, moderately to strongly overlapping mesoventrite, usually narrowly separated and rather small pro- and mesocoxae, depression along dorsum between pro- and pterothorax, antennae filiform or with more than three segments serrate and also comparatively narrow tarsi. Extant click beetles are widespread throughout the world. In the Mesozoic they were certainly more numerous than today. Elaterid remains appear in the fossil records at the boundary of the Triassic and Jurassic and represent great portion in many Mesozoic deposits, but in Cenozoic records their portion is comparatively smaller (Dolin Reference Dolin and Yaroshenko1973, Reference Dolin1975, Reference Dolin1976, Reference Dolin, Dolin, Panfilov, Ponomarenko and Pritykina1980; Ponomarenko 1995; Chang et al. Reference Chang, Kirejtshuk and Ren2010; etc.). Adults of Recent click beetles are often found on foliage or bark surfaces and may feed on plant shoots or tips, nectar or possibly epiphytic microflora. Their larvae are liquid feeders utilising extra-oral digestion, they may be either predaceous or phytophagous, and they occur under bark or rocks and in rotten wood, leaf litter or soil. This is a more or less common group in the Bembridge Marls, represented mostly by isolated elytra and two remains of incomplete beetle bodies. There are 17 species recognised, including four species of medium size and the rest are small ones. The remains of bodies belong to a member of the extinct genus Macropunctum Tröster, Reference Tröster1991, described from the Middle Eocene of Germany, but no other elytra of this genus with this characteristic structure were found among other remains. One elytron could belong to the genus Agrypnus Eschscholtz, Reference Eschscholtz1829, recorded with the generic name Lacon Laporte, Reference Laporte1836, also from Florissant (Upper Eocene) (Wickham Reference Wickham1916), Oeningen (Upper Miocene) (Heer Reference Heer1847) and Baltic amber (Spahr Reference Spahr1981).
Genus Macropunctum Tröster, Reference Tröster1991
Type species. Macropunctum messelensis Tröster, Reference Tröster1991, Middle Eocene, from the Messel Formation.
Macropunctum rossi Alexeev, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:2AC4120F-E588-4853-9408-581E9E991C28
Derivation of name. The name of this new species is devoted to the palaeontologist A. J. Ross.
Material. Holotype NHMUK I. 10085 (Figs 23b, 25): print of underside with missing head, legs and abdomen; Insect Limestone, NW Isle of Wight, Brodie Collection.
Additional specimen. NHMUK I. 9114: print of underside with obscure characters; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from most of the 11 described species of the genus (Tröster Reference Tröster1991, Reference Tröster1992, Reference Tröster1994) in the markedly smaller body size, more narrowly separated pro- and particularly mesocoxae, and also in the shape of mesoventrite, posterior edge, of which is twice excised and in the middle with wedge-shaped projection, reaching the middle of mesocoxae. This character of the new species is somewhat similar to that in Macropunctum densepunctum Wappler, Reference Wappler2003, which has such a projection, reaching the posterior edge of mesocoxae.
Description. Holotype I. 10085: length of specimen 7.7mm, width of print 3.1mm, length of elytron 4.9mm, width of elytron 1.5mm. More than 2.5 times as long as wide in the middle. Pronotum nearly as long as wide, slightly narrowing anteriorly and apparently with slightly arcuate lateral edges in distal two-thirds and strongly arcuately narrowing in anterior third. Entire surface of meso- and metaventrites with large and deep punctures filled with white wax-like matter. Prosternum with sutures forking in anterior two-sevenths. Underside of mesothorax without clear characters, except median trapezium-shaped mesoventrites with excised posterior edge and wedge-shaped process projecting posteriorly and reaching the middle of mesocoxae. Longitudinal median depression with a fossa reaching anterior edge of mesocoxae. Metaventrite transverse with distance between bases of metepisterna about 1.9 times as great as long in the middle and narrowing anteriorly. Metepisterna narrow, in distal half with straight sides, slightly oblique and in anterior part deviating to mesocoxae. Procoxae oval, small, distance between them less than a half of diameter of procoxae. Mesocoxae oval, about 1.5 times as large as procoxae, distance between them about a third of their diameter. Metacoxae short. Femoral plates at inner part as long as metacoxae, along inner half twice shortening and continuing with the same length. Abdomen unclear. Elytra visible as narrow stripes along pterothorax and most part of abdomen, each about 3.2 times as long as wide. Side of elytra in anterior two-thirds slightly arcuate and gently narrowing both anteriorly and posteriorly. Elytron with nine longitudinal punctured striae. Second–fifth striae from lateral edge closely approaching in distal sixth of elytron, although their joining unclear. Pressed punctures on elytra large and with white wax-like matter.
Family Throscidae Laporte, Reference Laporte1840

Figure 26 Beetles of families Throscidae, Byrhidae, Ptilodactylidae and Cantharidae. (a) Trixagus barclayi Kirejtshuk, sp. nov. (Throscidae): holotype, NHMUK I. 10017, body, lateroventral view; length of specimen = 2.6mm. (b) Byrrhites bembridgensis Kirejtshuk, gen. & sp. nov. (Byrrhidae): holotype, NHMUK I. 10174, body, lateroventral view; length of specimen = 3.8mm. (c) Paralichas striatopunctatus Kirejtshuk, sp. nov. (Ptilodactylidae): holotype, NHMUK In. 17449, pterothorax (negative print) and elytra, dorsal view; length of elytra = 3.6mm. (d) Themus (?Telephorops) polyaki Kirejtshuk, sp. nov. (Cantharidae): holotype, NHMUK In. 25641, body with missing head, dorsal view; length of specimen = 14mm.

Figure 27 Suborder Polyphaga, family Throscidae: holotype of Trixagus barclayi Kirejtshuk, sp. nov., NHMUK I. 10017, body, ventral view.
Material. Two specimens (identified by A. G. Kirejtshuk).
This comparatively small family includes in the Recent fauna beetles with medium-sized body (2.0–5.0mm), usually fusiform, i.e., more or less elongate and rather convex dorsally and ventrally, with head subvertically declined and rather retracted in the prothoracic segment, frequently developed antennal club or antennae serrate to pectinate, prosternal process broad and flat, emarginations on the thorax for reception of the legs and antennae, posterior pronotal angles strongly projecting posteriorly. They have a well-developed interlocking device and prothoracic segment with reduced lateral carina, lateral borders of the prosternal process extended anteriorly as ridges reaching anterior edge of the segment. This family is distinct from Buprestidae, Elateridae and Eucnemidae in having the lateral borders of the prosternal process extended anteriorly as ridges reaching anterior edge and also keels on head as well as in the non-connate ventrites 1 and 2, dorsal surface of pronotum and elytra in the place of articulation lying along one plane. The fossil examined shows characteristic antennal sockets at anterior edge of head, typical pronotosternal sutures and submesocoxal lines. Throscid beetles are frequently collected by sieving ground litter and debris (often along banks of waterbodies) or sweeping the low vegetation layer or in rotting conifer wood. Their larvae occur in soil, where they feed on ectomycorrhizal fungi on the roots of various trees. The family is spread almost throughout the world, although has not been reported from New Zealand. This family appears in the fossil record from the Lebanese and Burmese amber (Lower and boundary of Lower/Upper Cretaceous, respectively) (Rasnitsyn & Ross Reference Rasnitsyn and Ross2000; Kovalev et al. Reference Kovalev, Kirejtshuk and Azar2013) and Khetana deposits (Lower Cretaceous) (Gromov et al. Reference Gromov, Dmitriev, Yu Zherikhin, Lebedev, Ponomarenko, Rasnitsyn, Sukacheva and Ponomarenko1993). Most species from different genera were described from Baltic amber (Iablokoff-Khnzorian Reference Iablokoff-Khnzoryan1962; Muona Reference Muona1993; Kubisz Reference Kubisz2000), although some records of this family are known from London Clay (Upper Palaeocene–Lower Eocene) (Britton Reference Britton1960), Oise amber (Lowermost Eocene) (Kovalev et al. Reference Kovalev, Kirejtshuk and Nel2012), Messel (Middle Eocene) (Wedmann Reference Wedmann1994), Florissant (Upper Eocene) (Wickham Reference Wickham1914a) and Bitterfeld amber (Eocene) (Barthel & Hetzer Reference Barthel and Hetzer1982). A doubtful identification of this family was made in Fushun amber (Lower Eocene) (Hong Reference Hong2002).
Genus Trixagus Kugelann, Reference Kugelann1794
Type species. Dermestes adstrictor Herbst, Reference Herbst1792 (=Elater dermestoides Linnaeus, Reference Linnaeus1766) (by subsequent designation by Gyllenhal Reference Gyllenhal1808), Recent.
This genus is characterised by clavate antennae and lack of long oblique furrows on metaventrite for receipt of mesotarsi. The appearance of the remains examined is very similar to that in many Recent congeners. Adults of the Recent species of this genus occur on flowers and leaves in meadows and forest, and sometimes in litter at the sea shore. Most species of the genus are now found in areas with temperate and subtropical climate.
Trixagus barclayi Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:44228633-F97A-4F44-93E6-C4FDED3C4C0F
Derivation of name. The epithet of this new species is devoted to Maxwell V. L. Barclay, an enthusiastic coleopterist from the NHMUK and friend of the author, thanks to whose generous help many scientists from many countries have had very effective and prompt assistance for their study.
Materal. Holotype NHMUK I. 10017 (Figs 26a, 27): probably female; ventral surface from inside (negative print) with the distinct right antennal club and partly present outline of profemora; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This genus is recorded from Oise, Baltic and Bitterfeld amber and two species of it were described from Oise and Baltic amber (Iablokoff-Khnzorian Reference Iablokoff-Khnzoryan1962; Barthel & Hetzer Reference Barthel and Hetzer1982; Muona Reference Muona1993). Muona (Reference Muona1993) wrote that the members of this genus from Baltic amber have entire eyes and lack of paramedian grooves on metaventrite (both characters invisible in the specimen from the Bembridge Marls). One species of the genus was described from the lowermost French amber of Oise (Kovalev et al. Reference Kovalev, Kirejtshuk and Nel2012). Trixagus (Palaeothroscus) sosnovskii (Iablokoff-Khnzorian Reference Iablokoff-Khnzoryan1962) differs from the new fossil species in the less curved grooves on metaventrite for receipt of mesotarsi and not so expressed antennal club (indeed, this species could be a member of Aulonothroscus Horn, Reference Horn, Godman and Salvin1890 rather than Trixagus).
Pactopus americanus Wickham, Reference Wickham1914, described from the Florissant shales (Upper Eocene), is rather similar to the new species, although differs from it in the smaller and not very clear antennal club, wider metepisterna, peculiar configuration of grooves on the metaventrite for receipt of mesotarsi and apparently wider epipleural base. Wedmann (Reference Wedmann1994) described one specimen of Throscidae without genus attribution and species name. The latter are much wider and more robust than the type specimen of the new species described here and with not arcuate pronotal sides.
This new fossil species differs from the medium-sized and small Recent members of the genus in the more compact and shorter antennal club (which, on the other hand, could be a certain evidence of sex of the holotype described), or in the pronotal sides more arcuately narrowing anteriorly and much wider head. It differs from Trixagus majusculus Kovalev et al., Reference Kovalev, Kirejtshuk and Nel2012 in the smaller and more slender body, less oblique metacoxae and longer abdominal ventrite 1.
Description. Holotype I. 10017: length of specimen 2.6mm, width apparently 1.1mm, length of elytra (from base of mesoventrite to abdominal apex) apparently 1.7mm. Body elongate oval, apparently rather convex dorsally and ventrally; very light brown in colouration. Integument of head, prosternum and abdomen coarsely sculptured and without clear puncturation. Head of usual shape and rather long (three-quarters as long as wide at eyes), with clear antennal sockets at anterior edge, eyes moderately large. Labrum palpi and antennae invisible. Prothoracic segment widest at base and gently narrowing anteriorly. Elytra apparently completely covering abdomen, about 1.5 times as long as wide combined; widest at base, widely rounded along the middle and apices forming a joint arc. Prothoracic antennal grooves following the posterior edge of procoxae and gently turning along prosternal median plate. Prosternum with median longitudinal isolated plate and distinct notosternal sutures, procoxal cavities invisible, its anterior edge shallowly 3-emarginate. Mesoventrite rather short and unclear. Mesocoxae moderately large, suboval, moderately separated (distance between them about half of the diameter of mesocoxae). Metaventrite moderately long, about 1.5 times as long as prosternum. Metepisterna moderately narrow (about half as wide as distance between mesocoxae). Metacoxae obliquely transverse and of usual outline, with moderately long femoral plate. Epipleura rather wide, at base nearly as wide as diameter of mesocoxae. Abdomen somewhat than twice as long as metaventrite, very widely rounded at apex; ventrites 1–4 somewhat longer than the antennomeres 2–5. Profemora apparently of usual shape and about as wide as distance between mesocoxae.
Gen. & sp. indet.
Material. NHMUK I. 8927; left part of body (about 7.0mm long) including negative view of the head, part of metaventrite and abdominal ventrites, and apparently positive view of the prothorax; Insect Limestone, NW Isle of Wight, Brodie Collection.
Note. It is impossible to find any character to determine the genus of this specimen.
Discussion. The presence of Trixagus in the Bembridge Marls suggests that T. barclayi could have lived in debris at the shore of a waterbody or flew to the place of deposition. Muona (Reference Muona1993) noted that in Baltic amber four of five Recent genera have already been represented, and he described one extinct genus from there. He also made a conclusion that this family has been ‘more diverse formerly with several successful generic lines'. The finding considered here can at least provide indirect support of this hypothesis.
Family Eucnemidae Eschscholtz, Reference Eschscholtz1829
(Fig. 23d)
Material. One specimen (identified by A. G. Ponomarenko and A. G. Kirejtshuk) – NHMUK In. 17209.
This comparatively small group consists of species variable in body size (2.0–20.0mm), characterised by the subelliptic body, rather convex both dorsally and ventrally, with the head subvertically declined and rather retracted in the prothoracic segment, labrum not exposed from under frons, antennal insertions close to one another, pedicel obliquely inserted in the scape. Recent Eucnemidae or false click beetles are distributed in areas with forestry landscapes of all zoogeographical regions. Adults of this family are usually collected on plant surfaces, at lights at night or in flight-intercept traps. Larvae of most taxa occur in dead wood and are capable of moving through fairly solid wood by forcing their bodies between wood fibres; however, some larvae are soil-dwelling. The larvae of false click beetles are liquid feeders, so their actual diet is difficult to ascertain. It is possible that they feed on plasmodium of Myxomycetes, or digest plant tissue extra-orally. The overwhelming majority of fossil records of this family are from Caenozoic Baltic and Bitterfeld amber (Iablokoff-Khnzorian Reference Iablokoff-Khnzoryan1962; Spahr Reference Spahr1981; Muona Reference Muona1993). This group was also found as inclusions in Burmese amber (boundary of Lower/Upper Cretaceous) (Rasnitsyn & Ross Reference Rasnitsyn and Ross2000) and in Huangbanjigou (Lower Cretaceous) (Chang et al. Reference Chang, Muona, Hanyong, Li, Chen, Teräväinen, Ren, Qiang, Xingliao and Songhai2016). Some species were also described from London Clay (Lower Eocene) (Britton Reference Britton1960), White River deposits (Middle Eocene) (Scudder Reference Scudder1890) and Messel Middle (Eocene) (Wedmann Reference Wedmann1994), and also from the Florissant (Upper Eocene) (Wickham Reference Wickham1914b, Reference Wickham1916). The Bembridge Marls specimen has characters reminiscent of the Recent species of Dromaeolus Kiesenwetter, Reference Kiesenwetter1858, which was also found in Baltic amber (Klebs Reference Klebs1910; Muona Reference Muona1993).
Superfamily Byrrhoidea Latreille, Reference Latreille1804
Family Byrrhidae Latreille, Reference Latreille1804

Figure 28 Suborder Polyphaga, family Byrrhidae: holotype of Byrrhites bembridgensis Kirejtshuk, gen. & sp. nov., NHMUK I. 10174, body, ventral view.
Material. One specimen (identified by A. G. Kirejtshuk and A. G. Ponomarenko).
Byrrhidae or pill beetles are present in the Recent fauna as a small family with oval, robust, rather convex dorsally and usually medium-sized body (in most cases 3.0–7.0mm, but ranging from 1.0 to 15.0mm), head declined ventrally with the antennae frequently clubbed, thoracic sclerites with distinct depressions for antennae and legs, pronotum frequently with emarginate sides and posterior angles strongly projecting posteriorly, elytra with striate sculpture or without striae and epipleura not extended behind metacoxae, pro- and mesocoxae rather separated and contiguous metacoxae with distinct femoral plates, more or less dilated tibiae. In most cases their appendages can be completely withdrawn, leaving no part exposed to a predator. This family can be easily recognised as fossils due to quite characteristic features in position of head, disposition and structures of coxae. Most Recent species are distributed in the areas with moderate and prepolar climate, but a few species live in arid landscapes and in tropical territories. Pill beetles feed on leaves and rhizoids of mosses and liverworts (Bryophyta). A few species are associated with lichens and higher plants. Some pill beetles live in soil and adults of them sometimes emerge in a great number. In the fossil record this family with an unclear subfamily attribution appears from the Jurassic (Ponomarenko Reference Ponomarenko and Rasnitsyn1990). However, if the isolated elytra described as a formal genus Dzeregia Ponomarenko, Reference Ponomarenko and Rasnitsyn1985a indeed belong to this family, this group could originate in the Triassic and was one of the most numerous coleopterous group in the Jurassic deposits. Their abundance in the Cretaceous appears to be markedly greater than now (Hong Reference Hong1981; Ponomarenko Reference Ponomarenko and Rasnitsyn1990; Huang & Zhang Reference Huang and Zhang1997; Soriano et al. 2007; Kirejtshuk et al. Reference Kirejtshuk, Ponomarenko, Prokin, Chang, Nikolajev and Ren2010b; Kirejtshuk & Nel Reference Kirejtshuk and Nel2016). During the Caenozoic remains of pill beetles occur more rarely; they are not known from the European Eocene, but are described from the North American Upper Eocene (Florissant) and Miocene. In the Caenozoic deposits the subfamily Byrrhinae has many described species from the Neogene (Heer Reference Heer1849; Heyden & Heyden Reference Heyden and Heyden1866; Matthews Reference Matthews1974; etc.). Pill beetles are rather common in Pleistocene deposits. Cytilus tartarinus Scudder, Reference Scudder1900a (Byrrhinae) and Amphicyrta inhaesa Scudder, Reference Scudder1900a (Amphicyrtinae) have been described from the Florissant shales (Upper Eocene) (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko2018). The records of the subfamilies Byrrhinae and Syncalyptinae from Baltic amber were published without description (Spahr Reference Spahr1981; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984).
Subfamily Byrrhinae Latreille, Reference Latreille1804
The specimen examined should be regarded as a member of the subfamily Byrrhinae, because, in contrast to representatives of Amphicyrtinae LeConte, Reference LeConte1861, it has a wide prothoracic segment, wide protibiae and also, in contrast to Syncalyptinae Mulsant & Rey, Reference Mulsant and Rey1869, it has no trace of depressions for receipt of appendages. The metacoxae of this fossil specimen nearly reach the epipleuran edge and, therefore, it can be linked with the tribe Simplocariini Mulsant & Rey, Reference Mulsant and Rey1869. Besides, the dorsoventrally compressed femora and tibiae differentiates the new genus from other Simplocariini. However, it is impossible to find some characters to clarify if it has a closer relationship to Byrrhini (including Byrrhus Linnaeus, Reference Linnaeus1767; Cytilus Erichson, Reference Erichson1847; Porcinolus Mulsant & Rey, Reference Mulsant and Rey1869) or Pedilophorini Casey, Reference Casey1912 (including Eusomalia Casey, Reference Casey1912 and Listemus Casey, Reference Casey1912). Its regularly oval and not elongate body make it possible to differentiate it from Arctobyrrhus Münster, Reference Münster1902, Liochoria Pascoe, Reference Pascoe1875, Morychus Erichson, Reference Erichson1847 and Pedilophorus Steffahny, Reference Steffahny1842.
The very large and transverse antennal club is distinct from that in other members of the subfamily. This new genus is similar to some genera of tribes Byrrhini and Pedilophorini with medium-sized and short oval body, particularly to Byrrhus, Cytilus or Porcinolus, although the characters available do not allow certainty in this. This new species is characterized by the apparently gently convex lateral edges of pronotum, rather wide metepisterna and very short metafemoral plates with the simple posterior edge. This combination of the characters provides good diagnostics for this new species and makes it possible to propose a separate genus. Recent species of the mentioned genera have a larger body, emarginate pronotal sides or rather longer metafemoral plates. The genus Byrrhus is registered in Baltic amber (Berendt Reference Berendt1845–1856; Larsson Reference Larsson1978). “Byrrhus” lucae Heyden, Reference Heyden1859 has an shorter metaventrite and much wider apical part of body than those in the new fossil species here described. The other fossil species of the family were described from Rott (Heyden & Heyden Reference Heyden and Heyden1866), Oeningen (Heer Reference Heer1862) and Florissant (Scudder Reference Scudder1900b). All species formerly described need to be re-examined to clarify their attribution and proper diagnosis.
Tribe Simplocariini Mulsant & Rey, Reference Mulsant and Rey1869
Type genus Simplocaria Stephens, Reference Stephens1929
Genus Byrrhites gen. nov.
Type species. Byrrhites bembridgensis sp. nov.
Derivation of name. The name of this new genus is formed from the generic name Byrrhus. Gender: masculine.
Notes. This genus is represented by one species.
Diagnosis. Body of medium size. Head rather deflected and invaginated into prothorax, anterior part of frons (clypeus) isolated by transverse suture and labrum exposed. Eyes large. Antennae with thin flagella and very widely transverse 3-segmented club. Mesocoxae widely separated. Metacoxae subcontiguous. Mesoventrite very short. Metaventrite somewhat less than twice as long as prosternum and mesoventrite combined, without both visible discrimen and trace of metakatepisternal suture. Metepisterna in anterior part more than twice as wide as in posterior part. All femora and particularly tibiae rather flattened. Outer edge of all tibiae gently convex.
Comparison. Elongate and dorsally compressed body of Byrrhites bembridgensis sp. nov. with protrudent humeri and “soft” appearance makes it possible to consider the new genus as a member of the tribe Simplocariini positioned near Arctobyrrhus Münster, Reference Münster1902. The new genus is also characterised within Simplocariini.by the large antennal club, dorsoventrally compressed femora and tibiae.
Byrrhites bembridgensis Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:4C88FDB9-516A-40C2-B19D-A1D075402695
Derivation of name. The epithet refers to locality of the collecting of the holotype.
Material. Holotype NHMUK I. 10174 (Figs 26b, 28): underside with strongly declined head, more or less clear left side of thorax, right metacoxa, ventrites, epipleura, left anterior and intermediate legs; Insect Limestone, Isle of Wight, Brodie Collection.
Description. Holotype I. 10174: length of specimen 3.8mm, width 2.1mm, length of elytra 2.8mm. Body elongate, apparently rather convex dorsally and moderately convex ventrally; dark brown to blackish. Head, epipleura, thoracic and abdominal ventrites, femora and tibiae with rather coarse and relief cellular sculpture (or with wide scales looking like cellular sculpture) as well as with coarsely and sparsely diffuse punctures. Head of usual shape (shorter than wide behind eyes) and rounded at sides. Pronotum almost twice as wide as long, apparently convex; widest at base and strongly narrowing to gently arcuate anterior edge; anterior angles not expressed and posterior angles with distinct apices. Elytra about 2.7 times as long as combined width; slightly narrowed at base, widely rounded along the middle, slightly wider than pronotum, complete and covering apex of abdomen, their apices invisible. Metaventrite with parametacoxal lines. Procoxae moderately and mesocoxae rather widely separated. Metacoxae subcontiguous, slightly oblique medially and their clear short femoral plate with simple posterior edge. Profemur moderately short and widest at base, more than three times as long as wide. Protibia moderately short, rather dilated and almost as wide as profemur. Mesofemur somewhat wider and somewhat shorter than profemur. Mesotibia rather short and dilated, somewhat narrower than protibia. Metatibia with gently convex outer edge.
Discussion. The probable bionomy of this new species should be similar to that in most other species of the family, Recent members of which are associated with leaves and rhizoids of mosses and liverworts (Bryophyta).
Family Ptilodactylidae Laporte, Reference Laporte1836

Figure 29 Suborder Polyphaga, family Ptilodactylidae: holotype of Paralichas striatopunctatus Kirejtshuk, sp. nov., NHMUK I. 17449, elytra, pterothorax and base of abdomen, dorsal view.
Material. Four specimens (identified by A. G. Kirejtshuk).
This family is not numerous, but rather diverse in the Recent fauna, consisting of small- and medium-sized beetles (2.5–16.0mm), with the pronotum rather wide and convex at base and overhanging over the declined head, rather projecting labrum, long palpi and long antennae, procoxal cavities more or less transverse, metacoxae (sub)contiguous and with comparatively large femoral plate, 5–6 abdominal ventrites and more or less widened lobed tarsomeres 1–4. Most Recent diversity is found in areas with tropical and subtropical climate. At present, no species of this family is represented naturally in Europe (although Ptilodactyla exotica Chapin, Reference Chapin1927 has been spread in greenhouses by human activity in many countries). Most Recent species live in different riparian, semiaquatic and aquatic localities, mycetophagous and with larvae inhabiting wet rotten wood, leaf litter, gravel and debris at banks of water basins or in submerged wood and rotten foliage. The Ptilodactylidae, or toed-winged beetles, are described as fossils from Baltic amber (Ptilodactyloides stipulicornis Motschulsky, Reference Motschulsky1856 and ‘Pseudodactylus': Klebs Reference Klebs1910; Larsson Reference Larsson1978; Alekseev & Jäch Reference Alekseev and Jäch2016), and the boundary of Lower/Upper Cretaceous Burmese amber (Chatzimanolis et al. Reference Chatzimanolis, Cashion, Engel and Falin2012a). They are also recorded from Lower Cretaceous Lebanese amber (Kirejtshuk & Azar Reference Kirejtshuk and Azar2008, Reference Kirejtshuk and Azar2013). This family seemed to be rather numerous in the past; for example, it is very probable that a part of the Jurassic and Cretaceous remains described in the genus Artematopodites Ponomarenko, Reference Ponomarenko and Rasnitsyn1990 could be regarded in composition of this family. Finally, this family is also in materials from some Spanish and Chinese Jurassic or Cretaceous sites (Soriano et al. Reference Soriano, Declòs and Ponomarenko2007; Kirejtshuk et al. Reference Kirejtshuk, Ponomarenko, Prokin, Chang, Nikolajev and Ren2010b).
The remains examined can be compared with Ptilodactylidae and Cneoglossidae Champion, Reference Champion, Godman and Salvin1897, but, in contrast to all members of Cneoglossa Guérin-Méneville, Reference Guérin-Méneville1843, they have a much larger body and different peculiarities of puncturation and sculpture of elytra. Members of Ptilodactylidae very often have unusual puncturation and sculpture of elytra, in which the puncturation frequently tends to become denser and finer and looking more like microsculpture. The specimen examined belongs to a species with the quite characteristic ptilodactylid puncturation on elytra, rather narrow epipleura, narrowly separated mesocoxae, oblique metacoxae and peculiar configuration of submesocoxal lines.
Genus Paralichas White, Reference White1859
Type species. Paralichas guerini White, Reference White1859, Recent.
The remains examined probably belong to a genus with not very acuminate elytral apices. Elytral puncturation and sculpture between striae look like those in Lachnodactyla Champion, Reference Champion, Godman and Salvin1897, but the latter are with a larger and rather more slender body. Among these groups with not very acuminate elytral apices, the studied remains differ from the species of the genus Aploglossa Guérin-Méneville, Reference Guérin-Méneville1849 in having wider elytra with wider intervals between striae, much less coarse sculpture between the punctural rows and other peculiarities of puncturation and sculpture of the elytra (besides, many species of Aploglossa are much larger).
The striate puncturation on elytra is present in representatives of genera Anchycteis Horn, Reference Horn1880; Ptilodactyla Illiger, Reference Illiger1807; Epilichas White, Reference White1859; Stirophora Champion, Reference Champion, Godman and Salvin1897; Chaetodactyla Champion, Reference Champion, Godman and Salvin1897; Octoglossa Guérin-Méneville, Reference Guérin-Méneville1843; Falsotherius Pic, Reference Pic1913; and Byrrocryptus Broun, Reference Broun1893. But this new fossil species differs from them in the slightly smaller body and not so acute elytral apices (however, body size of Ptilodactyla is rather variable and in Epilichas the elytral puncturation is reduced and distinctly striate). It also differs from species of Daemon Laporte, Reference Laporte1836 and Microdaemon Kolbe, Reference Kolbe1897 in the not-so-acute elytral apices, sparser puncturation and more distinctly punctured striae. Finally, it differs from Pseudoepilichas Armstrong & Nakane, Reference Armstrong and Nakane1956 and Therius Billberg, Reference Billberg1820 in having wider elytra, as well as peculiarities of puncturation and sculpture of elytra (and also usually with reduced punctural striae), although members of small-sized species of Stenactyla Fairmaire, Reference Fairmaire1896 show very narrow elytral apices with deep longitudinal furrows.
This genus Paralichas is characterised by the elytra distinctly striately punctured and with reduced epipleura, and mesocoxae narrowly separated. The Recent range of it includes the Palaearchearctic Province of the Palaearctic Region, the Indo-Malayan Region, the eastern part of North America reaching Mexico and El Salvador (Ivie Reference Ivie, Arnett, Thomas, Skelley and Frank2002).
Paralichas striatopunctatus Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:AEA74CE6-7753-4E1D-AE59-D7804AAC43BA
Derivation of name. The epithet of this new species is formed from the Latin ‘striatus' (striate) and ‘punctatus' (punctured).
Material. Holotype NHMUK In. 17449 (Figs 26c, 29): apical part of elytra from above with distinct sculpture and puncturation, basal part removed and part of metanotum, mesocoxae, right metacoxa, metepisterna and submesocoxal lines are visible; Insect Limestone, NW Isle of Wight, Smith Collection.
Additional specimen. NHMUK I. 26107, one elytron of Ptilodactylidae (very probably conspecific to the described species of Paralichas), together with one elytron of Curculionoidea and one elytron of Coleoptera incertae sedis; Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. This new species differs from the Recent Indo-Malayan members of the genus in the somewhat smaller body and more smoothed sculpture of elytral integument between striae. It is also rather similar to Paralichas trivittis (Germar, Reference Germar1824), but markedly smaller.
Description. Holotype In. 17449: length of elytra 3.6mm, width 2.4mm. Pterothorax, abdomen and elytra arcuately outlined at sides; apparently rather convex dorsally and moderately convex ventrally; light brown to straw reddish in colouration. Elytra with granular and somewhat smoothed sculpture and with shallow longitudinal furrows conjoining apically along which more or less distinct punctures disposed. Elytra about 1.5 times as long as wide combined; slightly narrowing towards base, widely rounded along the middle and gently narrowing apically, complete and covering apex of abdomen. Metaventrite apparently twice as long as mesoventrite, with curved submesocoxal lines, but without visible parametacoxal lines. Metepisterna rather wide, apparently more than twice as long as wide. Mesocoxae moderately narrowly separated. Metacoxae strongly oblique medially, rather large, rather widely separated anteriorly and contiguous posteriorly, femoral plates rather large, but rather narrowing at rounded apices. Epipleura at base nearly two fifths as wide as metepisterna.
Gen. & sp. indet.
Material. NHMUK I. 10363: positive print of the middle part of both elytra with removed basal parts, beneath which the negative print is exposed; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 25564: narrow elytron 14mm in length, showing characteristic venation; Insect Limestone, NW Isle of Wight, Hooley Collection.
Discussion. Most Recent ptilodactylids live in various riparian, semiaquatic and aquatic localities, mycetophagous and with larvae inhabiting wet rotten wood, leaf litter, gravel and debris at banks of water basins or in submerged wood and rotten foliage. Thus, this group with such bionomy had many chances to come into sedimentary deposition.
Superfamily Cantharoidea Latreille, Reference Latreille1802
Family Cantharidae Latreille, Reference Latreille1802

Figure 30 Suborder Polyphaga, family Cantharidae: holotype of Themus (? Telephorops) polyaki Kirejtshuk, sp. nov., NHMUK In. 25641, elytra, scutellum, part of prothorax, left antenna, fragments of left legs, dorsal view.
Material. Two specimens (identified by A. G. Kirejtshuk).
This family is moderately numerous and rather diverse. In most cases the beetles have a rather thin and soft cuticle, prognathous head with exposed mandibles and palpi, flattened pronotum, diffuse and relief sculpture of elytra (frequently without clear puncturation), mostly reduced elytra with very narrow epipleura, narrowly separated or contiguous coxae (procoxae transverse, more or less projecting and with exposed trochantin), 8–10 abdominal ventrites, long and narrow legs with rather wide tarsomeres. This family now occurs globally, although it is more common in areas with a warm climate. In general, their larvae prey on other insects, but they sometimes do damage by gnawing young oak and other shoots. Adults have a relatively short life span and not infrequently appear in large numbers in vegetation and flowers. Many soldier beetles or Cantharidae were recorded and described from Baltic amber (Klebs Reference Klebs1910; Kuška Reference Kuška1992, Reference Kuška1996; Kazantsev Reference Kazantsev2013; etc.) and other European Caenozoic deposits (Kazantsev Reference Kazantsev2010: Rovno amber (Eocene); Heer Reference Heer1847: Radoboj (Lower Miocene) and Oeningen (Upper Miocene); Heyden & Heyden Reference Heyden and Heyden1866: Rott (Lower Miocene); Piton Reference Piton1939: Cantal (Upper Miocene); Förster Reference Förster1891: Brunstatt (Oligocene), etc.). The family is known from the Caenozoic of other regions of the globe, namely from the Shanwan (Lower Middle Miocene) (Zhang Reference Zhang1989), Sunchal (Palaeocene or Eocene) (Cockerell Reference Cockerell1936), Florissant (Upper Eocene) (Wickham Reference Thomson1909, Reference Wickham1913, Reference Wickham1917; etc.) and Chiapas (Oligocene–Miocene) (Wittmer Reference Wittmer1963). This family has also been found also in the Lower Cretaceous Lebanese and Albian-Cenomanian Burmese amber (Kirejtshuk & Azar Reference Kirejtshuk and Azar2013; Fanti et al. Reference Fanti, Damgaard and Ellenberger2018; etc.).
Genus Themus Motschulsky, Reference Motschulsky1857
Type species. Themus cyaneipennis Motschulsky, Reference Motschulsky1857, Recent.
Subgenus Telephorops Fairmaire, Reference Fairmaire1886
Type species. Telephorops impressipennis Fairmaire, Reference Fairmaire1886, Recent.
The proportions of elytra and character of sculpture on elytra of the fossil specimen accessible for study show that these remains belong to a member of the genus Themus, although they are somewhat similar in elytral characters to Podabrus Westwood, Reference Westwood and Westwood1838. It is impossible to determine the subgeneric attribution of this fossil species. Nevertheless, it seems to be a member of Telephorops Fairmaire, Reference Fairmaire1886 rather than Themus sensu stricto or Haplothemus Wittmer, Reference Wittmer1973. This genus is found in the eastern Hemisphere, mostly in Asia, particularly in the southern areas.
Themus (? Telephorops) polyaki Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:AB6E135B-683D-428C-9DA7-88BDE9FCF2A0
Derivation of name. The epithet of this new species is devoted to Vladimir A. Polyak, a friend of the author from childhood, who has maintained our relationship for over 40 years, showing a great generosity, sympathy and warm-heartedness.
Material. Holotype NHMUK In. 25641 (2) (Figs 21d, 26d): most part of the left elytron and whole right elytron, partly exposed posterior wings, remains of prothoracic sclerites, apex of antennae, left profemur, fragments of left meso- and metafemora, meso- and metatibiae, meso- and metatarsi (together with body of Scirtes sp. and two other coleopterous remains – incertae sedis); Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. Most similar to Themus (Telephorops) coelestis (Gorham, Reference Gorham1889), but clearly with a more smooth sculpture on elytra and a less metallic shade.
Description. Holotype In. 25641: length of specimen 14mm, width 5mm, length of elytra 10mm. Rather slender with soft integument, light straw on prothorax and black on elytra. Pronotum coarsely alutaceous and without clear puncturation. Elytra with dense, diffuse and rather shallow indistinct punctures, interspaces between them about the same as a puncture diameter and alutaceous; at sides and at apex puncturation becoming reduced and sculpture coarser; elytral base without punctures and microgranulose, and along the suture two elongate and slightly raised convex lines. Antennae apparently reaching distal two-thirds of elytra, left antenna represented by seven distal segments, antennomere 5 clearly widened apically and the following ones subcylindrical or slightly widened to apex, ultimate antennomere about four times as long as wide. Pronotum apparently subquadrangular, with widely rounded anterior angles. Scutellum transversely subtriangular. Elytra subparallel-sided to slightly widened apically, about twice as long as wide, subflattened and gently sloping laterally, widely arcuate at apices, adsutural line expressed along the entire length. Distal parts of posterior wings exposed between elytra and at left half of pterothorax base (where elytron was removed). Legs rather long and narrow. Femora apparently of usual shape, in length and width, about one-quarter as wide as elytron. Tibiae apparently slightly widened distally, slightly and very gradually curved, in length and width, about half as wide as femora. Tarsi rather wide, tarsomeres 2–4 distinctly lobed, tarsomere 4 deeply lobed (and protarsomere 4 nearly transverse). Protarsomere 5 longer than protarsomere 4 and with simple claws (dorsally).
Discussion. Adults of the Recent soldier beetles can occur in different types of vegetation and in a variety of open and forested habitats, although they are not known as having a preference for very wet places. This species could enter into the site of deposition as an occasional flyer.
Family Lampyridae Latreille, Reference Latreille and Cuvie1817
(Fig. 31a)

Figure 31 Beetles of families Lampyridae, Malachiidae and Nitidulidae. (a) Lampyridae, gen. & sp. indet.: NHMUK I. 9608, body, dorsal view; length of specimen = 4.0mm. (b) Attalus flexus Kirejtshuk, gen. & sp. nov. (Malachiidae): holotype, NHMUK In. 25571, body, lateral view; length of elytra = 2.7mm. (c) Epuraea (Epuraea) kozlovi Kirejtshuk & Kurochkin, sp. nov. (Nitidulidae): holotype, NHMUK I. 8506, body, ventral view. (d) Phenolia (Lasiodites) vanescens Kirejtshuk & Kurochkin, sp. nov. (Nitidulidae): holotype, part, NHMUK I. 10181, body, ventral view. Scale in mm.
Material. One specimen NHMUK I. 9608 (2) (identified by A. G. Kirejtshuk), showing the dorsal surface of elytra, remains of the prothoracic segment and tergites uncovered by elytra; Insect Limestone, NW Isle of Wight, Brodie Collection.
This comparatively small family is quite diverse and consists of many genera of usually medium-sized beetles (4.0–30.0mm in length) characterised by the usually elongate and flattened body with rather soft integument, flat pronotum frequently more or less covering head, deflected head with antennal insertions, which are more or less narrowly separated above mandibles, coxae narrowly separated or contiguous; frequently females or both sexes brachelytrous or some females larviform; may be luminescent as adults and larvae. Recent representatives of this family occur in all major regions of the world. Adults live in different terrestrial habits, although they are more frequently inhabitants of forest ecosystems. Larvae of fireflies or Lampyridae are predaceous, feeding on a variety of soft-bodied invertebrates, such as earthworms, snails and slugs or arthropods. This family is known mostly as Caenozoic fossils, namely from Baltic amber (Klebs Reference Klebs1910; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984), Florissant (Upper Eocene) (Wickham Reference Wickham1912), Luzice (Luschitze) (Upper Oligocene) (Beier Reference Beier1952), Rott (Lower Miocene) (Heyden & Heyden Reference Heyden and Heyden1866) and Oeningen (Upper Miocene) (Heer Reference Heer1865) although one species is described from Albian-Cenomanian amber (Kazantsev Reference Kazantsev2015). The remains examined are reminiscent of species with the elytra strongly widened at base and rather strongly narrowing apically (related to the genera Tenaspis LeConte, Reference LeConte1881 and Aspisoma Laporte, Reference Laporte1833) with light ornamental pattern on their discs; although, in contrast to many Recent species, this new fossil species is rather small (only 4.0mm in length body without head) and has no longitudinal ribs on elytra.
Infraorder Cucujiformia Lameere, Reference Lameere1938
Superfamily Cleroidea Latreille, Reference Latreille1802
Family Malachiidae Fleming, Reference Fleming1821

Figure 32 Suborder Polyphaga, family Malachiidae: holotype of Attalus flexus Kirejtshuk, sp. nov., NHMUK In. 25571, body, lateral view.
Material. One specimen (identified by A. G. Kirejtshuk).
This family is moderately numerous and rather diverse. In most cases, the soft-winged flower beetles (Malachiidae) have a rather thin and soft cuticle and are brightly coloured. They are easy to recognise by the presence of pouches, which can be inflated and then protruded from the sides of their thorax. Like the previous family, the soft-winged flower beetles are usually characterised by the prognathous head with exposed mandibles and palpi, flattened pronotum, diffuse sculpture of elytra (but frequently without clear puncturation), more or less incomplete elytra with very narrow epipleura and narrowly separated or contiguous coxae (procoxae transverse, more or less projecting and with the exposed trochantin). In contrast to Cantharidae, they have six abdominal ventrites, narrow or slightly widened tarsomeres. The Recent range of this family includes all zoogeographical regions, although they are most diverse in warm areas (particularly in the Old Mediterranean, including the Middle and Central Asia, and South-East Asia, Australia, Africa and South America). Adults are phytophagous (frequently pollinophagous) and live on vegetation and usually on flowers, although sometimes can feed also on other insects. Larvae are primarily predaceous. This family is not frequent as fossils, although it is fairly common in Upper Eocene Baltic amber (Spahr Reference Spahr1981; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984; Kubisz Reference Kubisz2001; Tshernyshev Reference Tshernyshev2016) and lowermost Eocene Oise amber (Kirejtshuk & Nel Reference Kirejtshuk and Nel2008, Reference Kirejtshuk and Nel2013). It is also found in Florissant (Upper Eocene) (Wickham Reference Wickham1912, Reference Wickham1917; etc.) and Cantal (Upper Miocene) (Théobald Reference Théobald1937). It was also recorded in lists of taxa from the Lower Cretaceous Lebanese amber and boundary of Lower/Upper Cretaceous Burmese amber (Rasnitsyn & Ross Reference Rasnitsyn and Ross2000; Kirejtshuk & Azar Reference Kirejtshuk and Azar2013).
Subfamily Malachiinae Fleming, Reference Fleming1821
Genus Attalus Erichson, Reference Erichson1840
Type species. Attalus lusitanicus Erichson, Reference Erichson1840, Recent.
Attalus flexus Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:4EF916EE-1EFD-4E0A-8DA9-FD3B830D8092
Derivation of name. The epithet of this new species means ‘bend', ‘curve', ‘turn'.
Material. Holotype NHMUK In. 25571 (Figs 31b, 32): left side of positive part with visible pronotum, most part of right elytron and abdominal apex; negative part of head with a scape, pro- and metafemora, part of protibia, most part of mesotibia; negative part of incomplete metaventrite with visible metacoxa and abdominal base, with some remains of integument in negative part; Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. This new species is similar to the modern members of Attalus but distinct from all of them with the larger head and width of pronotum. Klebs (Reference Klebs1910) included the genus Attalus in the list of Coleoptera from Baltic amber, however, this record should be confirmed by further finding.
Description. Holotype In. 25571: female; length of elytra 2.7mm, height 1.1mm. Head rather large and much longer than pronotum, with eyes rather large. Visible maxillary palpomere almost twice as long as wide, oblong, pointed. Pronotum narrow, about twice as wide as long and with gently curved edges. Elytra apparently almost covering abdomen. Metaventrite moderately convex. Pro- and mesocoxae subconical and rather projecting. Metacoxae transverse and apparently contiguous. Abdominal ventrite 1 apparently longest and ventrite 2 somewhat shorter than each of ventrites 3 and 4. Pro- and mesofemora moderately wide and apparently wider than pro- and mesocoxae.
Discussion. Adults of most Recent species of Attalus and other genera of soft-winged flower beetles live on different kinds of vegetation and, therefore, the specimen examined could be expected on parts of Typha plants growing above the water level or it may have flown into the area of deposition.
Superfamily Cucujoidea Latreille, Reference Latreille1802
Family Nitidulidae Latreille, Reference Latreille1802
(Figs 31c, d, 33–35)
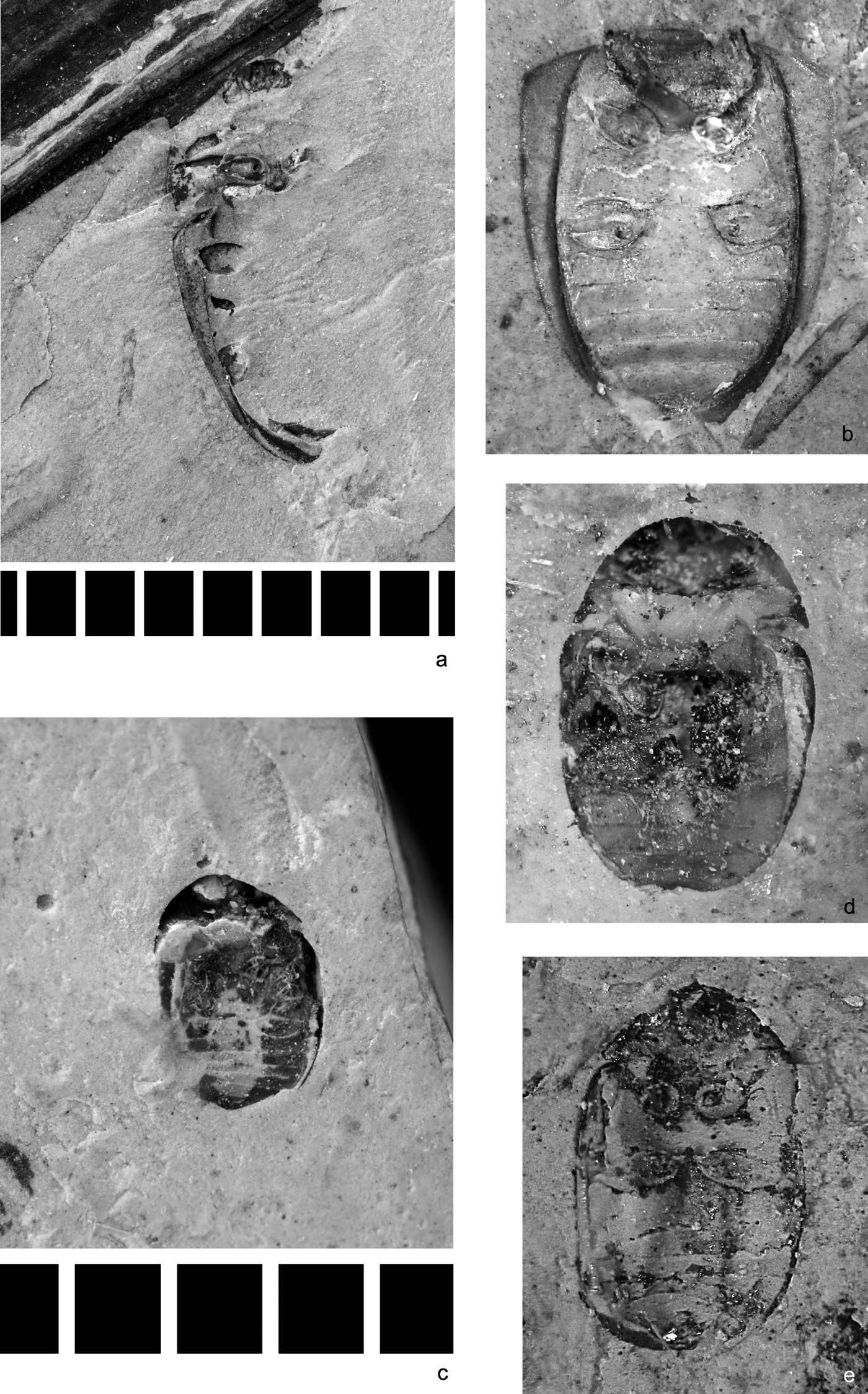
Figure 33 Beetles of family Nitidulidae. (a) Phenolia (Lasiodites) vanescens Kirejtshuk & Kurochkin, sp. nov.: holotype, counterpart, CAMSM X.50140.73, left part of body and legs, ventral view. (b) Prometopia europaea Kirejtshuk & Kurochkin, sp. nov.: NHMUK In. 17438, pterothorax, abdomen and epipleura, ventral view; length of specimen = 3.13mm. (c) Cyllodes argutus Kirejtshuk & Kurochkin, sp. n.: holotype, NHMUK I. 8595, body, ventral view. (d) Cyllodes sp.: NHMUK I. 10305, body, ventral view; length of specimen = 1.98mm. (e) Coxollodes palaeogenicus Kirejtshuk & Kurochkin, sp. nov.: holotype, NHMUK I. 10264, body, ventral view; length of print = 1.34mm.

Figure 34 Suborder Polyphaga, family Nitidulidae. (a) Holotype of Epuraea (Epuraea) kozlovi Kirejtshuk & Kurochkin, sp. nov., part, NHMUK I. 8506, body, ventral side. (b) Holotype of E. (E.) kozlovi sp. nov., counterpart, NHMUK I. 8492, body, dorsal view. (c) Holotype of Phenolia (Lasiodites) vanescens Kirejtshuk & Kurochkin, sp. nov., NHMUK I. 10181, body ventral with traces of sculpture of right elytron. (d) Holotype of P. (L.) vanescens sp. nov., counterpart, CAMSM X.50140.73 (TN 124), left side of body, ventral view.

Figure 35 Suborder Polyphaga, family Nitidulidae. (a) Holotype of Prometopia europaea Kirejtshuk & Kurochkin, sp. nov., NHMUK In. 17438, pterothorax and abdomen, ventral view. (b) Holotype of Cyllodes argutus Kirejtshuk & Kurochkin, sp. nov., NHMUK I. 8595, body, ventral view. (c) Cyllodes sp., NHMUK I. 10305, ventral view. (d) Holotype of Coxollodes palaeogenicus Kirejtshuk & Kurochkin, sp. nov., NHMUK I. 10264, body, ventral view.
Material. Nine specimens (identified by A. G. Kirejtshuk and A. S. Kurochkin).
Nitidulidae or sap beetles are a rather numerous and diverse family in the Recent fauna, characterised by small to medium body size (0.9–15.0mm), usually with clavate antennae, transverse pro- and mesocoxae, strongly transverse (never oblique) metacoxae. Nitidulidae in the Recent epoch is rather diverse and variable in bionomy and found in many ecological circumstances. However, this group is not present in water basins and in very wet substrates. It is also quite rare in arid areas. Soil is also very seldom inhabited by sap beetles. Most representatives are mycetophagous and also many groups are associated with flowers. Rare species of this family are phyllophagous or predaceous; and some groups live on colonies of scale insects and whiteflies (Cychramptodini Kirejtshuk & Lawrence, Reference Kirejtshuk and Lawrence1992 and Cybocephalinae Jacquelin du Val, Reference Jacquelin du Val1858). The most diverse representation of sap beetles is recorded in forest communities. The sap beetles (Nitidulidae) appear as fossils from the Lower Cretaceous and all earlier records are erroneous (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko1990). In this paper, the Carpophiline lineage of the family was recorded somewhat earlier (Baissa, Middle Neocomian) than the Nitiduline lineages (Obeshtshayushtshiy Creek, Cenomanian), and some groups of both carpophiline and nitiduline lineages were also recognised among specimens from Montsec (Barremian, Lower Cretaceous) (Soriano et al. Reference Soriano, Declòs and Ponomarenko2007) and Albian-Cenomanian Burmese amber (Kirejtshuk & Chetverikov Reference Kirejtshuk and Chetverikov2018). During the Caenozoic, this family is mostly known from the Upper Eocene Baltic amber (recorded chiefly in lists without description by Klebs Reference Klebs1910; Larsson Reference Larsson1978; Spahr Reference Spahr1981; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984), where all subfamilies of Nitidulidae have been found, except Amphicrossinae Kirejtshuk, Reference Kirejtshuk1986a and Cillaeinae Kirejtshuk & Audisio Kirejtshuk, Reference Kirejtshuk1986b. The latter was recorded in Florissant shales (Upper Eocene) (Wickham Reference Wickham1913), together with other sap beetles from Carpophilinae Erichson, Reference Erichson1842 and Nitidulinae. The oldest Caenozoic nitidulids were recorded in Menat (Palaeocene), which are represented there with different groups (Nabozhenko & Kirejtshuk Reference Nabozhenko and Kirejtshuk2013, Reference Nabozhenko and Kirejtshuk2017; Kirejtshuk & Nel Reference Kirejtshuk and Nel2018; etc.). Recently, various nitidulid groups were described from Baltic and Bitterfeld amber (Kirejtshuk & Poinar Reference Kirejtshuk and Poinar2007; Kirejtshuk & Kurochkin Reference Kirejtshuk and Kurochkin2010; Kurochkin & Kirejtshuk Reference Kurochkin and Kirejtshuk2010; Kirejtshuk Reference Kirejtshuk2011) and some were found in the lowermost Oise amber (Kirejtshuk & Nel Reference Kirejtshuk and Nel2008, Reference Kirejtshuk and Nel2013). From the Caenozoic there are Phenolia (Lasiodites) caucasicus (Kirejtshuk & Ponomarenko, Reference Kirejtshuk and Ponomarenko1990) from Wishnevaya Balka beds (North Caucasus, Karaganian: Middle Miocene); Amphotis depressa Théobald, Reference Théobald1937 and Oligamphotis irenae Théobald, Reference Théobald1937 from the Kleinkembs (Pays de Bade) deposits (Lower Oligocene); Amphotis bella Heer, Reference Heer1847 from the Lower Miocene Radoboj deposits; and ‘Cybocephalus cf. californicus Horn, Reference Horn1879' from Mojave Desert deposits (Middle Miocene) (Palmer et al. Reference Palmer, Carvalho, Cook, O'Neill, Petrunkevitch and Sailer1957).
Subfamily Epuraeinae Kirejtshuk, Reference Kirejtshuk1986a
Tribe Epuraeini Kirejtshuk, Reference Kirejtshuk1986a
This tribe has been recorded somewhat earlier than the other groups of the family (Baissa, Middle Neocomian). Almost all Recent groups of this subfamily live in forest, feeding on tree fungi or substrates attacked by them. There are exceptions among some hundreds of species: a few species can breed in other decaying remains of plant origin, like grassy vegetation and fruits of Angiosperms. Most representatives of the tribe in the Recent fauna are concentrated in the eastern Hemisphere, some groups related to the Palaearctic ones occur in North America, but in South America the only members are central-American Amedanyraea Kirejtshuk & Pakaluk, Reference Kirejtshuk and Pakaluk1996 and Pantropical Haptoncus Murray, Reference Murray1864.
Genus Epuraea Erichson, Reference Erichson1843
Type species. Nitidula decemguttata Fabricius, Reference Fabricius1792, Recent.
(?) Subgenus Epuraea (Epuraea) Erichson, Reference Erichson1843
Type species. As for genus.
The subgenus includes species with small- and medium-sized body (with few exceptions larger than 4mm), more or less elongate oval, moderately convex dorsally and flattened ventrally. Their dorsum always has diffuse puncturation. As a rule, the elytra leave at least a part of pygidium uncovered, but very rarely the whole pygidium and part of the previous segment are left uncovered. Pro- and mesocoxae narrowly separated, but distance between metacoxae usually much greater than that between pro- and mesocoxae. The subgeneric attribution is not quite clear, although most probably the new fossil species should be regarded in the composition of the nominative subgenus because of the absence of characters used for the discrimination of most subgeneric groups (Kirejtshuk Reference Kirejtshuk1998). The distance between metacoxae in the new fossil beetle is comparatively narrow and the posterior edge of its metaventrite between coxae is angularly excised. If the posterior edge of the metaventrite of this species is quite characteristic of Epuraeinae, the complete elytra of it, rather sloping at apex, are not very characteristic of Epuraea and Epuraeinae at all, while among members of Nitidulinae such a condition of the elytra is more usual. Therefore, there is some doubt about the subfamily attribution of the specimen examined.
Almost all Recent representatives of the genus Epuraea inhabit different types of forests and rarely occur anywhere else. Their trophics and development are connected with fungi, oozing tree sap and similar substrates. It is highly probable that the same environment existed in the Isle of Wight in the late Eocene.
Epuraea (? Epuraea) kozlovi Kirejtshuk & Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:785F3F17-C45B-45A3-A91F-0BC240595D58
Derivation of name. The epithet of this new species is devoted to M. A. Kozlov, an outstanding entomologist and participant in the project on Bembridge Insect Beds, whose sudden death prevented him from continuing his valuable contribution to the Isle of Wight hymenopterous fauna.
Material. Holotype NHMUK I. 8506 (Figs 31c, 34a): part with the underside showing clear outlines of most sclerites, except of any appendages, and epipleura. Counterpart: NHMUK I. 8492 (Fig. 34b): dorsum with clear outlines of all sclerites, except of latero-apical part of right elytron and appendages; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from most Palaearctic consubgeners in its shape of pronotum rather narrowing to posterior angles. It is diagnosed due to a peculiar combination of characters, such as arcuate sides of the pronotum (narrowing posteriorly and anteriorly); complete elytra rather sloping at apex; comparatively large eyes; large, sparse and rather distinct punctures on dorsum (especially punctures on elytra).
Description. Holotype I. 8506 (part) and I. 8492 (counterpart): length of part 2.1mm, width 0.9mm, length of elytra 1.5mm (counterpart: length of print 2.1mm, width 0.9mm, length of elytra 1.5mm). Rather elongate with pronotum and elytra gently narrowing apically; comparatively convex both dorsally and ventrally; remains of integument unicolorous brownish. Pronotum with round, shallow and distinct punctures, separated by about a puncture diameter; abdominal ventrite 1 with similar puncturation, but punctures less distinct. Elytra and metaventrite with interspaces between punctures greater, up to two puncture diameters; head and prosternum with somewhat smoothed puncturation, looking almost like coarse granulation. Head a little shorter than distance between eyes, with almost straight anterior edge, with large eyes, seemingly composed of comparatively large facets. Pronotum about 2.13 times as wide as length, with very gently sloping to (sub)explanate sides, widest at middle and almost equally narrowing anteriorly and posteriorly, with widely rounded anterior angles and posterior angles distinct and somewhat projecting posteriorly. Prosternal process narrow and subrhomboid, widely rounded to subangular at apex, moderately extending beyond procoxae, somewhat curved along procoxae. Elytra 1.7 times as long as wide combined, with gently sloping sides, and conjointly rounded apices (without sutural angle). Distance between mesocoxae about 1.25 times and that between metacoxae about 1.75 times as great as that between procoxae. Metaventrite transverse, with straight anterior edge and angularly excised posterior edge between coxae. Hypopygidium with widely rounded apex.
Notes. The complete elytra of the fossil species under consideration slope at apices and also the shape of its pronotum and character of puncturation are reminiscent of some representatives of the family Cryptophagidae. However, the distinctly transverse procoxae with exposed protrachantin, in addition to the other characters mentioned here, support the attribution of the new fossil species to Nitidulidae.
? Epuraea sp.
Material. NHMUK I. 8596: positive print of the dorsum, with clear outlines of all sclerites, without latero-apical part of right elytron and appendages; Insect Limestone, NW Isle of Wight, Brodie Collection.
Notes. Length 2.7mm, width 1.1mm. This specimen shows pronotum widest at base and elytra longest at suture.
Subfamily Nitidulinae Latreille, Reference Latreille1802
Tribe Nitidulini Latreille, Reference Latreille1802
This group is the most numerous and diverse in the Recent fauna. Recent representatives of this tribe show high diversity in bionomy; however, most genera are characteristic of Recent forest assemblages, usually developing in association with tree fungi and substrates attacked by them. The geographic range of the Recent fauna is almost worldwide, but the highest diversity and number of members are distributed in the Neotropical Region. This group appears in the published fossil record no earlier than Baltic amber (Schaufuss Reference Schaufuss1892; Klebs Reference Klebs1910; etc.). Nevertheless, in the Oligocene and later, this tribe became common in oryctocenoses of different sites.
Genus Phenolia Erichson, Reference Erichson1843
Type species. Nitidula grossa Fabricius, Reference Fabricius1801 (by monotypy), Recent.
Subgenus Lasiodites Jelínek, Reference Jelínek1999
Type species. Nitidula picta Maclay, Reference Macleay1825 (original designation by Jelínek, Reference Jelínek1999), Recent.
This subgenus differs from the nominative one in the less-developed antennal grooves at the sides of the median part of head, somewhat flattened dorsal surface of head, narrower prosternal process, sexual dimorphism in the tibiae and distinctly truncate apex of the male pygidium. It consists of comparatively large species with elongate oval body, moderately convex dorsally and flattened ventrally with more or less raised costae on complete elytra. Such general appearance also occurs in some groups of the Soronia-complex (Kirejtshuk Reference Kirejtshuk2003), although the moderately narrow prosternal process of the new fossil species makes it possible to separate it in the prints examined (larger species of the Soronia-complex usually have prosternal process widened apically). Phenolia (Lasiodites) vanescens sp. nov. seemed to also have a great similarity with extant members of the subgenus Lasiodites in both structure and bionomy. The Recent species of Lasiodites are mostly known to feed and to breed in decaying fruits and some other decaying plant remains, and are associated with all types of forests and even savannahs. The highest diversity of it in the Recent fauna was observed in mountains and tropical forests. Distribution of the extant fauna of the subgenus is restricted mostly to the eastern Hemisphere, mainly in the Afro-Malagasy and Indo-Malayan regions, but Phenolia (Lasiodites) picta (Macleay, Reference Macleay1825) are also found in the Australian Region, and the latter, together with some other species, increased their range due to mankind's activities (Kirejtshuk & Kvamme Reference Kirejtshuk and Kvamme2002). The genus is known from Miocene deposits of Stavropolsky Kray (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko1990) and Baltic amber (Kirejtshuk & Kurochkin Reference Kirejtshuk and Kurochkin2010). The new fossil species considered could have some association with broad-leaved forests.
Phenolia (Lasiodites) vanescens Kirejtshuk & Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:BD9124CD-CAC9-43C1-BD77-D863968B141A
Derivation of name. The epithet of this new species is formed from the Latin ‘vanesco' (to disappear, to vanish).
Material. Holotype. NHMUK I. 10181 (Figs 31d, 34c) (part): combination of positive part of the underside (head, prosternum, right part of metaventrite and abdomen, and also epipleura) and negative print of dorsum (mesonotum and elytra) (with large pieces of Typha remains); Insect Limestone, NW Isle of Wight, Brodie Collection. CAMSM X.50140.73 (TN 124) (Figs 33a, 34d) (counterpart): inner side (combination of negative and positive prints) of head, left part of prosternum, left epipleuron, apex of left elytron, left part of abdominal apex, left anterior leg, intermediate leg and part of metafemur; Insect Limestone, Isle of Wight, Smith Collection.
Diagnosis. This new species has the same body outline and shape of thoracic sclerites, character of sculpture of elytral integument and, in particular, shape of prosternal process as those in many extanct species of the subgenus Lasiodites. It is rather similar to Phenolia (Lasiodites) caucasicus Kirejtshuk, 1990 from the Miocene deposits of the Western Caucasus (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko1990) in its general body outline, shape of pronotum, presence of longitudinal rows on elytral surface, similar character of puncturation on prosternal process and shape of profemora, but differs from it in the posterior angles of pronotum more projecting posteriorly, somewhat narrower epipleura, narrower prosternal process with probably arcuate or subangular apex, much smaller punctures on elytra surface and more expressed elytral rows. The species under description also differs from Phenolia (Lasiodites) angustitibialis Kirejtshuk & Kurochkin, Reference Kirejtshuk and Kurochkin2010 in the larger body, coarser puncturation on head, narrower pronotum with the posterior angles more projecting and shorter elytra. The new species is also very similar to many Recent species of the subgenus Lasiodites, particularly Phenolia (Lasiodites) bakkei Kirejtshuk & Kvamme, Reference Kirejtshuk and Kvamme2002, P. (L.) chevrolati (Reitter, Reference Reitter1873), P. (L.) costipennis (Boheman, Reference Boheman1851), P. (L.) intermixta Kirejtshuk & Kvamme, Reference Kirejtshuk and Kvamme2002, P. (L.) picta and other consubgeners, due to its general body outline, although the elytral apices of the new fossil beetle seem to be more acute, like those in representatives of the subgenus Aethinodes Blackburn, Reference Blackburn1891. Some Recent narrow species of Lasiodites have acute elytral apices, but their bodies are certainly much narrower than that in the new species. The prosternal process of this new species is very similar to that in the Recent members of the subgenus.
Description. Holotype I. 10181 (part) and X.50140.73 (TN 124) (counterpart): length of part 7.2mm, width 4.1mm, length of left elytron 4.7mm, length of right elytron 4.8mm. Elongate oval, probably strongly convex dorsally and slightly ventrally; remains of integument brownish. Head with large and oval distinct punctures at base, separated by half of a puncture diameter; prosternum with shallow, but quite distinct, sparse and large irregular punctures in the median part and becoming very indistinct and shallower laterally and towards prosternal process, interspaces between them about as broad as one puncture diameter; epipleura with more clearly visible distinct punctures about as large as those on prosternal median part but somewhat sparser; elytra with at least seven rather well-expressed elevated longitudinal rows with traces of shallow punctures, intervals between rows with traces of sparse, rather shallow and large punctures, separated by about one puncture diameter or slightly more. Head transverse and with almost straight anterior edge. Scape of usual shape, about as broad as pedicellum is long. Pronotum about 2.5 times as wide as long, about a third as long as elytra, widest at base, gently sloping at sides, its posterior side moderately emarginate at almost distinct posterior angles. Prosternum slightly medially convex; its process moderately widened before seemingly widely rounded apex and very slightly curved along procoxae. Elytra 1.16 times as long as wide combined, evenly sloping at sides, narrowly rounded at apices, longest at suture, with a shallow sutural angle; epipleura at base about 1.6 times as wide as prosternal process. Protibia with slightly convex outer and almost straight inner edges. Profemora gently convex along anterior and posterior edges.
X.50140.73 (TN 124) (counterpart): length 6.8mm, width 3.6mm, length of left elytron 4.6mm. Antennal grooves subparallel-sided behind mentum. Prosternal process rather widened at widely rounded apex and with clear top of lateral angles, about as wide as procoxa is long. Femora somewhat wider than procoxa is long. Tibiae about as wide as prosternal process in the narrowest place (pro- and mesotibiae not curved at apex).
(?) Phenolia sp.
Material. NHMUK In. 43409: inner side of elytra, scutellum, base of pronotum and also a part of ventral surface of abdomen; Insect Limestone, NW Isle of Wight, Hooley Collection.
Notes. These remains, with length of elytra 3.0mm and their combined width 2.7mm, show outlines of pronotal base, scutellum and elytra, including the lateral basal angle somewhat similar to Phenolia species. The surface of elytra, with longitudinal furrows, is like those in many species of the genus.
Genus Prometopia Erichson, Reference Erichson1843
Type species. Nitidula sexmaculata Say, Reference Say1825, Recent.
Notes. This genus is regarded as consisting of two subgenera, Prometopia sensu stricto and Parametopia Reitter, Reference Reitter1884b, which are distinguished only by the shape of antennal club.
? Subgenus Prometopia Erichson, Reference Erichson1843
Type species. As for genus.
The new species described here resembles the extant species of the genus Prometopia and also species of Palaeometopia Kirejtshuk & Poinar, Reference Kirejtshuk and Poinar2007 from Dominican amber (Kirejtshuk & Poinar Reference Kirejtshuk and Poinar2007). The principal characters for the discrimination of these genera are located in the head and prothoracic segment. Unfortunately, both these tagmas are missing in the specimen examined. The news species has an oval body, so is considered to belong to the widespread extant genus Prometopina, rather than the extinct genus (Palaeometopia) with species that are elongated with long elytra. The genus consists of two subgenera, which can scarcely be separated without the study of the antennal club: species of Prometopia sensu stricto have a symmetric three-segmented club, while those of Parametopia Reitter, Reference Reitter1884b have an asymmetric four-segmented club. Thus, the subgeneric attribution of the new fossil species described here should be regarded as provisional, although elytral shape and other characters accessible in the specimen examined look like those in members of Prometopia sensu stricto. Many species of Prometopia, such as the new species considered, have a narrow isolated plate at the middle of the outer edge of metepisterna, while the known species of the extinct genus demonstrate no trace of such a plate.
The Recent species of the subgenus Prometopia sensu stricto mostly inhabit subcortical space of trees and visit fermenting tree sap and decaying tree fungi in subtropical and tropical forest. The extant species of the subgenus Prometopia sensu stricto are spread almost throughout the world, but no species is present in the Western Palaearctic (including Europe). Species of Parametopia are restricted in their range by the East Chinese (Palaearchearctic) Province and Indo-Malayan Region. The only previously known fossil species, Prometopia depiles Scudder, 1877, was described from a partial specimen (head missing, thorax broken) collected at the left bank of the Fraser River at Quesnel, British Columbia, and was dated as Lower Oligocene.
Prometopia (? Prometopia) europaea Kirejtshuk &
Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:7E2C6323-C876-4F86-A7FB-EE66B26236A5
Derivation of name. The epithet of this new species refers to the continent of its origin.
Material. Holotype NHMUK I. 17438 (Figs 33b, 35a): positive print of the underside of the pterothorax, abdomen and epipleura; Insect Limestone, NW Isle of Wight, Smith Collection.
Diagnosis. This new fossil species is characterised by a certain width of elytra, which can rather restrict the number of species for comparison (Prometopia (Parametopia) concolor Grouvelle, Reference Grouvelle1891, comb. nov.; Prometopia (Prometopia) aequalis Sharp, Reference Sharp, Sharp, Matthews and Lewis1890; P. (Prometopia) bidentata Schaeffer, Reference Schaeffer1909; P. (Prometopia) confluens Erichson, Reference Erichson1843; P. (Prometopia) cryptarchoides Reitter, Reference Reitter1873; P. (Prometopia) marginata Grouvelle, 1902; P. (Prometopia) quardimaculata Motschulsky, Reference Motschulsky1863; P. (Prometopia) quadrinotata Grouvelle, Reference Grouvelle1896; P. (Prometopia) rauca Sharp, Reference Sharp, Sharp, Matthews and Lewis1890; P. (Prometopia) sexmaculata (Say, Reference Say1825)); among the species with similar width of epipleura, the new fossil species can be diagnosed by the peculiar configuration of submesocoxal and submetacoxal lines, and also, in many cases, by the comparatively narrower epipleura. The American fossil P. depiles (see Scudder Reference Scudder1890), according to the original description and drawing, is rather similar to P. (? Prometopia) europaea sp. nov., although in comparison with the type specimen of the species described here it is slightly larger (length of elytra 3.75mm), with a less arcuate outline of elytra.
Description. Holotype I. 17438: length of specimen (without head and prothorax) 3.1mm, width of elytra 2.9mm, width of epipleura at base 0.4mm. Body elongate oval, subflattened ventrally; remains of integument reddish. Elytra 1.21 times as long as combined width, probably with similarly rounded apices. Elytral shoulders very strongly developed and wide. Epipleura very wide at base and probably almost reach apices of elytra, anterior epipleural angle with a distinct angle. Distance between metacoxae about 1.79 times as great as that between mesocoxae. Mesocoxae oblong oval and metacoxae transverse. Mesoventrite with posterior edge shallowly emarginate between mesocoxae, base of scutellum moderately wide and with probably missing apex. Metaventrite about as long as mesoventrite, with rather emarginate posterior edge. Metepisterna moderately narrow and with one isolated plate at their outer edge before the middle. Submetacoxal line distinct, running close to submetacoxal cavity and slightly arcuately deviating along middle of it. Ventrite 1 with rather shallow, round and sparse punctures separated by 2–3 puncture diameters. Ventrite 2 with rather shallow, round and denser punctures separated by 1.0–1.5 puncture diameters and gradually becoming smaller and denser apically. Hypopygidium with widely rounded apex.
Notes. This specimen shows some similarity to species of the family Coccinellidae because of the general oval body outline and lines behind coxal cavities. Behind each of the meso- and metacoxal cavities, two curved lines can be seen. However, in both cases the anterior one is a true structural line (submeso- and submetacoxal lines), while the distal ‘line' behind both coxal cavities appeared as a result of mineralisation of the insect and conditions of concretion of salt, which formed an irregular ‘line' when it was cracked by the palaeontologist who found it. This print specimen has the transverse mesocoxae, quite characteristic of Nitidulidae in general, while Coccinellidae are characterised by smaller and oval mesocoxae. The Recent species of Coccinellidae with wide epipleura have elytra more rigid and with anterior external angle somewhat rounded (but not with as distinct a top as represented in the new fossil beetle).
Prometopia depiles seems to be a true member of the genus because of its quite characteristic contour of epipleura, although the drawn pronotum looks like that in the members of Lobiopa Erichson, Reference Erichson1843 rather than in Prometopia (it is widest almost at the middle). Scudder (Reference Scudder1890, p. 500) pointed out that the pronotum of it is ‘proportionally shorter than in Prometopia'. It is difficult to compare these fossil species, because the print of P. depiles represents the dorsal surface of body, while that of P. (? Prometopia) europaea sp. nov. includes the underside sclerites of pterothorax with epipleura and abdomen.
Tribe Cyllodini Everts, Reference Everts1898
This tribe is rather uniform in structure, but quite diverse in bionomy. Its members are mostly mycetophagous, feeding on tree fungi and substrates infested by them or rarely on fungal fruit bodies growing on soil, although some are anthophagous, feeding on flowers, including decaying flowers (species of Camptodes Erichson, Reference Erichson1843 develop on large flowers of cacti, Amaryllis, etc.). It had a rather early origin, with one extinct genus described from the Upper Cretaceous sediments of Obeshtshayushtshiy Creek (Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko1990) and another found from the Lower Cretaceous of Spain.
Genus Cyllodes Erichson, Reference Erichson1843
Type species. Strongylus ater Herbst, Reference Herbst1792 (designated by Parsons Reference Parsons1943), Recent.
The representatives of this genus always have a short oval body, strongly convex dorsally and flattened ventrally; head somewhat protracted into prothorax and declined downwards; pronotum transverse, widest at base, somewhat covering base of elytra and with sharply pointed posterior angles; deeply excavate mesothorax; comparatively widely separated metacoxae; posterior edge of metaventrite widely emarginate or almost straight; prohypomera and epipleura steeply sloping downwards. In the fossil record this genus is known only from Baltic amber (Klebs Reference Klebs1910); however, the rather similar form Cyllolithus mirandus Kirejtshuk in Kirejtshuk & Ponomarenko (Reference Kirejtshuk and Ponomarenko1990) was described from Obeshtshayushtshiy Creek (Upper Cretaceous). This genus now has a world range, but most species of it are concentrated in the Indo-Malayan Region. Species of this genus have more or less uniform bionomy associated with tree fungi; most species occur in soft decaying fruit bodies of fungi from Agaricaceae.
Cyllodes argutus Kirejtshuk & Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:64B1BF1C-61BC-4F3C-96A9-0CD9D758EB0C
Derivation of name. The name of this new species is formed from the Latin ‘argutus' (sharp, pointed) due to the very distinct posterior angles of the pronotum in the new species.
Material. Holotype NHMUK I. 8595 (Figs 33c, 35b): ventral surface from inside (negative print) represented by most sclerites, except for appendages; head and prothoracic segment deep inside the limestone matrix; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. The new species resembles some Recent species with somewhat elongate oval, but not (sub)semicircular body, such as in Cyllodes angustipes Grouvelle, Reference Grouvelle1908, Cyllodes dubius (Reitter, Reference Reitter1877) and Cyllodes marginatus Grouvelle, Reference Grouvelle1908; however, the new species is distinct from them in having a much smaller body. It is also characterised by the rather narrowly separated procoxae and comparatively widely separated metacoxae.
Description. Holotype I. 8595: length of print 2.4mm, width 1.6mm, length of elytra 2.0mm. Oval and somewhat elongate body, probably strongly convex dorsally and very slightly ventrally; remains of integument brownish. Head rather small, narrower than half of pronotal base. Pronotum about 2.95 times as wide as long, steeply sloping at sides, widest at base, with very distinct posterior angles. Elytra 1.49 times as long as wide combined, epipleura rather wide and steeply sloping laterally. Prosternum seemingly (sub)carinate with rather narrowly separated procoxal cavities. Distance between mesocoxae about two-thirds of that between metacoxae. Metafemora with gently convex anterior and posterior edges. Mesoventrite distinctly carinate and with convex posterior edge between mesocoxae. Mesoventrite, epipleura and abdomen with very indistinct, sparse and shallow puncturation, interspaces between punctures superficially microshagreened. Metaventrite strongly transverse, with straight posterior edge between metacoxae. Abdominal ventrite 1 nearly as long as ventrites 2–4 combined and about 1.5 times as long as hypopygidium. Hypopygidium with widely rounded apex.
Cyllodes sp.
Material. NHMUK I. 10305: positive print of the underside, although not all sclerites clearly outlined and area between prosternum and mesoventrite particularly obscured because of mineralised substrate; Insect Limestone, Isle of Wight, Brodie Collection.
Description. Length of print 2.0mm, width 1.3mm, length of elytra 1.4mm. Distance between metacoxae about 2.5 times as great as that between procoxae. Elytra apices widely rounded and probably forming very shallow sutural angle. Epipleura reaching base of ventrite 2.
Notes. The specimen examined is rather similar to Cyllodes argutus sp. nov., but somewhat smaller and has no character to discriminate it. Therefore, the authors prefer to leave this form without species epithet.
Genus Coxollodes Kirejtshuk, 1987
Type species. Pallodes cyrtusoides Reitter, Reference Reitter1884, Recent.
This genus is rather similar to other genera of Cyllodini with oval and convex bodies. It is represented by some Recent species, which, in contrast to other genera, are characterised by the small body, very short prosternum, submesocoxal lines slightly deviating from the posterior edge of coxae, extremely narrowly separated procoxae and comparatively narrowly separated metacoxae. The specimen examined has most of these characters. The range of this genus is now restricted to the Palaearchearctic Province of the Palaearctic Region, as well as the Indo-Malayan and Australian regions. Bionomy of the Recent members of this genus seems to be very similar to other mycetophagous groups of the tribe and they are associated with tree fungi.
(?) Coxollodes palaeogenicus Kirejtshuk & Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:66F15959-11B4-4BA7-8FC6-580F0E032439
Derivation of name. Epithet of this new species is formed from the name of the stratigraphical period, dating its holotype.
Holotype. NHMUK I. 10264: negative print of poorly preserved beetle without visible remains of appendages (except for traceable contour of left metafemur); Insect Limestone, Isle of Wight, Brodie Collection.
Diagnosis. This new species seems to be more elongate than the Recent congeners and its posterior coxae somewhat more narrowly separated.
Description. Holotype I. 10264: length of specimen 1.3mm, width 0.9mm, length of elytra 1.1mm. Body elongate oval, rather convex dorsally and flattened ventrally. Head comparatively small and declined ventrally. Pronotum strongly transverse, widest at base, gradually narrowing anteriorly, with blunt to rounded posterior angles. Elytra complete, about 1.24 times as long as combined width, widest at anterior quarter, arcuately narrowing both anteriorly and posteriorly, with very gently sloping sides, slightly wider than pronotum; probably widely rounded at apex forming a joint arc. Epipleura moderately wide at base, probably rather sloping laterally (subvertical). Metepisterna long and gradually narrowing posteriorly. Mesocoxae transversely oval and rather moderately separated. Pro- and metafemora somewhat flattened and with moderately convex outer and inner edges. Metaventrite rather short and with apparently emarginate posterior edge between coxae. Metafemur nearly reaching lateral edge of elytra. Abdominal ventrite 1 with a narrow anterior median intercoxal projection, longer than rest of ventrites; hypopygidium somewhat shorter than ventrite 1 and much longer than ventites 2–4. Pygidium and apparently hypopygidium very widely rounded at apex and somewhat covering hypopygidium.
Discussion. Most representatives of this family considered and described here belong to the groups associated with forest assemblages and are characterised by mycetophagy and xylomycetophagy, although some Recent members of the genus Epuraea live in meadows and other grasslands feeding on decaying grass. No Recent group of sap beetles lives in water basins or under very wet conditions, except inhabitants of exuding tree sap and wet decaying fruits, or rarely in interstices of tropical rainforest trees and plants filled by water (Kovac et al. Reference Kovac, Jelínek, Hashim and Wiwatwitaya2007). Therefore, at least most sap beetles were unlikely to have been living under swamp conditions, where deposition of the Bembridge Marl Beds was proceeding. Thus, they are thought to have flown to these swamps from adjacent forests. The most Recent diversity of the genera Phenolia, Prometopia and Coxollodes are present in areas with a comparatively warm climate; these groups are absent in Recent Europe.
Family Cryptophagidae Kirby, Reference Kirby and Richardson1837

Figure 36 Beetles of families Cryptophagidae, Latridiidae, Corylophidae, Ciidae and Anthicidae. (a) Telmatophilus britannicus Kirejtshuk & Kurochkin, sp. nov. (Cryptophagidae): holotype, NHMUK I. 10117, body, dorsal; length of specimen = 1.63mm. (b) Corticariites kozlovi Kirejtshuk, sp. nov. (Latridiidae): holotype, NHMUK I. 10297, body, ventral view; length of specimen = 1.6mm. (c) Orthoperites antiquus Kirejtshuk & Kurochkin, sp. nov. (Corylophidae): holotype, NHMUK I. 9520, body ventral view; length of specimen = 0.9mm. (d) Octotemnites sepultus Kirejtshuk, gen. & sp. nov. (Ciidae): holotype, NHMUK I. 8774, body, dorsal view; length of specimen = 1.3mm. Cyclodinus efficax Kirejtshuk, sp. nov. (Anthicidae): (e) holotype, NHMUK I. 9457, body, dorsal view; length of print = 2.0mm; (f) paratype, NHMUK I. 10259, body, dorsal view; length of specimen = 2.1mm.

Figure 37 Suborder Polyphaga, family Cryptophagidae: Telmatophilus britannicus Kirejtshuk & Kurochkin, sp. nov., NHMUK I. 10117, body, ventral view.
Material. One specimen (identified by A. G. Kirejtshuk and A. S. Kurochkin).
This uncommon family includes, as a rule, rather small-sized beetles (usually 1.0–3.0mm and not larger than 6.0mm), very diverse in both bionomy and structure. They are usually elongate in shape, although sometimes rather short and oval, more or less convex dorsally, with clavate antennae, comparatively small and round to slightly transverse procoxae, mesocoxal cavities closed externally and narrowly separated, diffuse puncturation on almost complete elytra. Both active instars of the living circle are mycetophagous and associated with decaying substrates of plant origin, including under bark space and hollow trees, fungi and decaying grass, vertebrate and invertebrate animals' nests, cowsheds, stores, cellars, etc. The Recent range of this family covers all zoogeographical regions. Most of its representatives inhabit forest ecosystems. This coleopterous family was also recorded in the Upper Eocene Baltic and Bitterfeld amber (Klebs Reference Klebs1910; Barthel & Hetzer Reference Barthel and Hetzer1982; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984) and also in Rovno amber (Lyubarsky & Perkovsky Reference Lyubarsky and Perkovsky2010, Reference Lyubarsky and Perkovsky2012). The oldest record of the family is dated to the Lower Cretaceous (Cai & Wang Reference Cai and Wang2013: China, Yixian Formation), although cryptophagid-like specimens were also found in Lower Cretaceous Spain (Peris et al. Reference Peris, Lyubarsky and Perkovsky2017), Lebanese amber (Kirejtshuk & Azar Reference Kirejtshuk and Azar2013) and Upper Cretaceous Taimyr amber (Zherikhin Reference Zherikhin, Arnoldi, Zherikhin, Nikritin and Ponomarenko1977). Fossil Atomariinae are recorded from the Neogene (Heer Reference Heer1862; Zhang Reference Zhang1989), and fossil representatives of Cryptophagidae are known from Florissant shales (Upper Eocene) and Green River beds (Scudder Reference Scudder1890; Wickham Reference Wickham1913, Reference Wickham1914a) and Sunchal deposits (Palaeocene–Eocene) (Cockerell Reference Cockerell1926). Here, one species of the genus Telmatophilus Heer, Reference Heer1841 is recorded from the Bembridge Marls and also from Baltic amber (Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984). Recent congeners of it are associated with vegetation near water basins, including species of Typha.
Subfamily Cryptophaginae Kirby, Reference Kirby and Richardson1837
This subfamily is most numerous and most diverse among other groups of the family Cryptophagidae.
Tribe Cryptophagini Kirby, Reference Kirby and Richardson1837
Genus Telmatophilus Heer, Reference Heer1841
Type species. Telmatophilus typhae Fallen, Reference Fallén1802, Recent.
All characters of the remains examined (shape of head, antennal club, all pairs of coxae, outline of pronotum, elytra and abdomen) correspond with those in the Recent members of the genus Telmatophilus, although unusually the tarsi are missing. This genus was recorded also in Baltic and Bitterfeld amber (Klebs Reference Klebs1910; Barthel & Hetzer Reference Barthel and Hetzer1982; Hieke & Pietrziniuk Reference Hieke and Pietrzeniuk1984). Recent Holarctic species are associated with aquatic plants, particularly with Typha and Sparganium.
Telmatophilus britannicus Kirejtshuk & Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:0A524A9F-E8D9-483E-B05A-E6D771BD5090
Derivation of name. The epithet of this new species refers to the Isle of Wight in the British Isles.
Material. Holotype NHMUK I. 10264 (Figs 36a, 37): negative print of poorly preserved beetle (print of ventral side of body from inside) with antennae, but without any other appendages; Insect Limestone, Isle of Wight, Brodie Collection.
Diagnosis. This new fossil species differs from the Recent congeners in combination of the small-sized body, apparently light colouration and comparatively short part of body behind prothoracic segment. The lateral edges of pronotum of the new species are more arcuate, and the posterior edge of metaventrite between coxae seems to be straight rather than excised or emarginate. It looks most similar to Telmatophilus schoenherri (Gyllenhal, Reference Gyllenhal1808) because of its comparatively short posterior part of body and comparatively wide antennomere 9, but except for lighter colouration and more arcuate pronotal sides, the new species is distinct from the latter in the less projecting posterior angles of pronotum and much shorter prosternal process.
Description. Holotype I. 10264: length of specimen 1.6mm, width 0.6mm, length of elytra 1.0mm. Body elongate oval, apparently moderately convex dorsally and subflattened ventrally. Head with slightly convex anterior margin, transversely abrupt at anterior edge, with comparatively large and prominent eyes and with transverse line (frontoclypeal suture) at level of anterior edge of eyes. Antennae about one and a third as long as head wide and about 2.5 times as long as antennal club, almost reaching base of pronotum; club 3-segmented, elongate, slightly loose and consisting of comparatively large transverse segments. Pronotum about 1.95 times as wide as long, widest at middle and arcuately narrowing both anteriorly and posteriorly, gently sloping at sides, with almost pointed anterior and posterior angles, and probably slightly concave posterior margin. Elytra about 1.60 times as long as wide combined, slightly arcuate at sides, slightly wider than pronotum, apparently gently sloping at sides, apparently complete and covering apex of abdomen, their apices seeming to form a conjoint arc. Epipleura moderately narrow, slightly narrowing apically and almost reaching apices of elytra. Prosternum moderately long and with process comparatively wide, somewhat narrower than antennal club, subparallel-sided, truncate and slightly widened at apex. Distance between metacoxae about twice as wide as that between procoxae. Procoxae closed posteriorly, oval and slightly transverse. Metaventrite about as long as prosternum and mesoventrite combined, its posterior edge between coxae subtruncate. Abdominal ventrite 1 almost as long as ventrites 2–4 combined; hypopygidium widely rounded at apex.
Discussion. This finding is not unexpected, because the new species from the Bembridge Marls is similar to that of Recent ones; it could have easily come into the depositional swamp environment where Typha grew.
Family Latridiidae Erichson, Reference Erichson1842

Figure 38 Suborder Polyphaga, family Latridiidae: Corticariites kozlovi Kirejtshuk, sp. nov.; NHMUK I. 10297, body, ventral view.
Material. One specimen (identified by A. G. Kirejtshuk).
Latridiidae or mould beetles form a moderately numerous family of small beetles (0.8–3.0mm), usually more or less elongate and slightly or moderately convex dorsally. They have quite a characteristic combination of characters, including the comparatively large head with rather wide anterior part and clavate 10–11-segmented antennae placed closely anterior edge (at distance from eyes), coxae in all pairs more or less separated, very coarse sculpture of dorsum, 5–6 abdominal ventrites and three-segmented tarsi. Recent mould beetles are, at present, a completely mycetophagous group associated with spores and mycelium of different fungi (both lower and higher), particularly moulds and other Ascomycetes, living in branches of trees, leaf litter, fruit bodies of macrofungi and decaying substrates of plant origin. Recent species are distributed worldwide, except for pre-polar areas. The earliest records of this group are findings in Lebanese amber (Lower Cretaceous) (Kirejtshuk & Azar Reference Kirejtshuk and Azar2008, Reference Kirejtshuk and Azar2013; Kirejtshuk et al. 2009a, b) and Burmese amber (boundary of Lower/Upper Cretaceous) (Rasnitsyn & Ross Reference Rasnitsyn and Ross2000), and later in Taymyr amber from Yantardakh (Upper Cretaceous) (Zherikhin Reference Zherikhin, Arnoldi, Zherikhin, Nikritin and Ponomarenko1977). The highest number of members of this family (both Latridiinae and Corticariinae) were recorded from Baltic amber (Upper Eocene) (Klebs Reference Klebs1910; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984; Borowiec Reference Borowiec1985; Kubisz Reference Kubisz2000; Hawkeswood et al. Reference Hawkeswood, Makhan and Turner2009; Bukejs et al. Reference Bukejs, Kirejtshuk and Rücker2011, Reference Bukejs, Rieke and Rücker2012a, Reference Bukejs, Rücker and Kirejtshukb; Rieke Reference Rieke2012; etc.), although some species were also recovered from the Florissant shales (Upper Eocene) (Wickham Reference Wickham1913, Reference Wickham1914a, Reference Wickhamb), Aix deposits (Heer Reference Heer1856), Rott (Lower Miocene) (Schlechtendal Reference Schlechtendal1894) and Durham (Pliocene) (Lesne Reference Lesne1920).
Subfamily Corticariinae Curtis, Reference Curtis1829
This subfamily can be mis-identified as Hydraenidae because of its small body size and six ventrites; however, it can be distinguished due to the more separated coxae in each pair of legs, larger head, very short mesoventrite and different structure of the prothoracic segment.
Genus Corticariites Kirejtshuk, gen. nov.
Type species. Corticariites kozlovi sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:AC268FB1-B49D-44FE-9E60-706F2311787D
Derivation of name. The name of this new genus is formed from the generic name Corticaria. Gender: masculine.
Notes. This new genus is represented by the only species and, therefore, the description of it overlaps with the description of species.
Diagnosis. This fossil specimen belongs to the subfamily Corticariinae because of six clearly visible ventrites, as in species of Corticarina Reitter, Reference Reitter1880 and Cortinicara Johnson, 1975. However, the outline of the body in members of the latter is quite different from that in the fossil species under consideration. This new fossil species, in contrast to the species of the subfamily Corticariinae, has the pronotum widened anteriorly and somewhat reminiscent of that in the subfamily Latridiidae rather than Corticariinae. It has a quite short and narrow prothoracic segment, which is shorter than the head (Recent species of the genus Migneauxia Jacquelin du Val, Reference Jacquelin du Val1857 have a prothoracic segment with a somewhat similar outline, but less regularly hexagonal, markedly wider and longer than that in the new fossil species). Procoxae of the fossil specimen described are rather large, but they can be observed from inside, while in the Recent specimens they are usually observed from outside. Succinimontia infleta Zherichin, 1977 has pronotum widened anteriorly, but its abdomen includes only five ventrites and its head is clearly smaller than those in the new species described here.
It is difficult to compare the new fossil species with Corticaria aeterna Wickham, Reference Wickham1914, because the latter presents its dorsal surface in the holotype (drawn by the descriptor), but the latter is markedly larger (2.9mm in length), with smaller proportion of the head, and longer and narrower elytra (Wickham wrote that the species described by him is larger than most modern species). As to other fossil latridiids described (see the list in Kirejtshuk & Ponomarenko Reference Kirejtshuk and Ponomarenko2018), all of them belong to the genera with five ventrites. The underside of some species described from Baltic amber was not studied (Borowiec Reference Borowiec1985; Kubisz Reference Kubisz2000); however, the latter are also rather different from the new fossil species in general outline of the more slender body, markedly larger head and shorter prothoracic segment.
Corticariites kozlovi Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:A923E3CC-FFED-4186-A3DC-511029137851
Derivation of name. The epithet of this new species is devoted to M. A. Kozlov, an outstanding entomologist and participant of the project on Bembridge Insect Beds, whose sudden death prevented him from continuing his valuable contribution to the Isle of Wight Hymenoptera fauna.
Material. Holotype NHMUK I. 10297 (Figs 36b, 38): demonstrating the underside from inside (negative print) with the partly present profemora and tibiae, base of left mesofemur; elytral sides missing; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Holotype I. 10297: length of print 1.6mm, width apparently 0.8mm, length of elytra (from base of mesoventrite to abdominal apex) apparently 1.1mm. Body elongate oval, apparently moderately convex dorsally and ventrally; very light brown in colouration. Integument of mesoventrite and left metepisternum smoothed and without clear puncturation. Head of usual shape and rather long (somewhat shorter than width at eyes), subhexagonal, eyes moderately large. Labrum palpi and antennae invisible. Prothoracic segment apparently hexagonal. Elytra apparently completely covering abdomen, apparently about 1.4 times as long as wide combined; slightly narrowed at base, widely rounded along the middle and apices forming a joint arc. Prosternum apparently not carinate, its anterior edge shallowly emarginate. Procoxae transverse (from inside and apparently circular from outside) and narrowly separated. Mesoventrite rather short and unclear. Mesocoxae comparatively large (from inside), suboval and slightly transverse, moderately separated (distance between them about twice that of between procoxae). Metaventrite moderately long, about as long as prosternum and mesoventrite combined. Metepisterna moderately narrow, width at anterior edge somewhat greater than the distance between procoxae and somewhat less than the distance between mesocoxae, and gradually narrowing posteriorly. Metacoxae transverse and of usual outline, apparently moderately or rather widely separated. Abdomen somewhat longer than metaventrite, very widely rounded at apex; each of ventrites 1 and 6 somewhat longer than ventrites 2–5 taken separately. Pro- and mesofemora of usual shape and about as wide as distance between mesocoxae. Mesotibiae rather narrow, somewhat shorter and somewhat less than two-fifths as narrow as femora, narrowing at base and at apex. Protibiae rather narrow.
Discussion. This family has a rather early origin (not later than Lower Cretaceous) and it seems that at around the time of the formation of Bembridge Marls this group was common and its members were rather diverse. The specimen examined could have flown to the place of deposition, although this new species is more likely associated with decaying grass in swamps.
Family Corylophidae LeConte, Reference LeConte1852

Figure 39 Suborder Polyphaga, family Corylophidae: holotype of Orthoperites antiquus Kirejtshuk & Kurochkin, sp. nov., NHMUK I. 9520, body, ventral view.
Material. One specimen (identified by A. G. Kirejtshuk).
Corylophidae are uncommon, very small and short oval beetles (0.8–3.0mm), usually weakly convex both dorsally and ventrally (rarely elongate oval or rather convex), with the head small and frequently hidden under pronotum, mesocoxal cavities closed, widely separated metacoxae, abdomen with six ventrites (first is longest), elytra (sub)truncate and tarsi 4-4-4 or apparently 3-3-3. Recent species of this group occur in all zoogeographical regions. Corylophidae live in decaying plant matter and on surfaces of bark or leaves, and in various accumulations of rotting vegetation, such as hay stacks and grass piles; some of them can be collected in ant nests. Larvae and adults feed on spores and hyphae of moulds. This family was recorded as fossils only from lowermost Eocene Oise amber (Kirejtshuk & Nel Reference Kirejtshuk and Nel2013) and Upper Eocene Baltic amber (Klebs Reference Klebs1910; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984).
Subfamily Orthoperinae Jaquelin du Val, 1857
Genus Orthoperites Kirejtshuk & Kurochkin, gen. nov.
Type species. Orthoperites antiquus sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:DB9D14CB-2084-4714-A169-256CBEB49B77
Derivation of name. The name of this new genus is formed from the generic name Orthoperus. Gender: masculine.
Diagnosis. The rather small remains examined show the declined head, rather separated transverse metacoxae, truncate posterior edge of the metaventrite between coxae and epipleura somewhat sloping downwards laterally. This combination of characters gives enough ground to regard these remains as belonging to a species of the family Corylophidae or minute fungus beetles. However, it is impossible to carry out a proper comparison of the fossil species with members of the Recent genera and, therefore, the new genus is proposed here as a formal one or morphogenus and needs further clarification. Nevertheless, it can be provisionally linked with Orthoperus Jaquelin du Val, 1857, because the species of the latter have the pronotal base usually markedly narrower than elytra, but the new genus differs from Orthoperus in the less widely separated metacoxae.
Orthoperites antiquus Kirejtshuk & Kurochkin, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:45B73566-13CC-4416-ABE1-8CED1FE10E94
Derivation of name. The epithet of this new species means ‘ancient', ‘antique', ‘very old'.
Material. Holotype NHMUK I. 9520 (Figs 36c, 39): negative print of poorly preserved beetle with visible underside and dorsal surface of declined head as well as basal parts of epipleura; head and thorax, borders of ventrites and epipleura with remains of chitin; Insect Limestone, Isle of Wight, Brodie Collection.
Description. Holotype I. 9520: length of print 0.9mm, width 0.6mm. Body elongate oval and apparently twice as long as wide, rather convex dorsally and subflattened ventrally; head and thorax dark brown. Integument of head and thorax apparently finely and sparsely punctured and finely sculptured. Head moderately large and strongly declined ventrally. Pronotum strongly transverse, apparently widest at base, gradually narrowing anteriorly. Elytra not visible, although epipleura slope steeply downwards laterally. Procoxae invisible. Mesocoxae somewhat transverse and rather widely separated (distance between them greater than transverse diameter of mesocoxae). Metaventrite moderately long and with truncate posterior edge between coxae. Metepisterna moderately wide and subparallel-sided (with width apparently half the distance between mesocoxae). Distance between metacoxae markedly greater than that between mesocoxae. Abdominal ventrite 1 about as long as ventrites 2 and 3 combined; hypopygidium somewhat shorter that ventrite 1 and arcuate at apex.
Discussion. As the Recent Corylophidae are associated with decaying plant matter in terrestrial assemblages, the species examined would only enter the site of deposition of the Bembridge Marls by chance.
Superfamily Tenebrionoidea Latreille, Reference Latreille1802
Family Ciidae Leach in Samouelle, Reference Samouelle1819

Figure 40 Suborder Polyphaga, family Ciidae: holotype of Octotemnites sepultus Kirejtshuk, gen. & sp. nov., NHMUK ‘I. 8774', body, ventral view.
Material. One specimen (identified by A. G. Kirejtshuk).
This comparatively small family includes small beetles (usually 2.5–15.0 and rarely up to 1.2 or 19.0mm) with a dark (sub)cylindrical body, deflected head, 8–10-segmented antennae with three-segmented loose club, procoxae slightly transverse to conical and projecting, complete elytra with diffuse or seriate puncturation, tibiae without spurs and tarsi rather narrow. Recent minute tree fungus beetles are found in many, but not pre-polar, areas, mostly in forest landscapes. Both larvae and adults live in tree fungi, including very hard bracket fungi, Polyporaceae and other Basidiomycetes, often in large numbers. This family is rare as fossils apparently because of the comparatively small body size and lack of distinctive structural characters, making identification difficult, although it was recorded from Baltic amber (Upper Eocene) (Klebs Reference Klebs1910; Spahr Reference Spahr1981) and in Burmese amber (boundary of Lower/Upper Cretaceous) (Rasnitsyn & Ross Reference Rasnitsyn and Ross2000), and some species were described from Rott (Lower Miocene) (Heyden & Heyden Reference Heyden and Heyden1866). One specimen of this family, which is covered by a ‘milky' coating, is in the piece of Baltic amber deposited in the Paris museum (Muséum National d'Histoire Naturelle).
The remains of the specimen examined have some resemblance to those from some groups of the families Tenebrionidae and Scolytidae, although they could not belong to any of those groups because of the outline of the visible sclerites of dorsum, characteristic sloping of head, pronotum and elytra, including apices of the elytra, as well as details of the sculpture of integument.
Subfamily Orophiinae Thomson, Reference Thomson1863
Genus Octotemnites Kirejtshuk, gen. nov.
Type species. Octotemnites sepultus sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:C9D5D17C-0554-4FC6-92A0-FBBCD988FAD8
Notes. This genus is represented by the only species and, therefore, the description of it overlaps with the description of species.
Derivation of name. The name of this new genus is formed from the generic name Octotemnus. Gender: masculine.
Diagnosis. This new genus is proposed as formal, because it is impossible to find any strong similarities of the new species described here with species of the genera Octotemnus Mellié, Reference Mellié1847 or Rhopalodontus Mellié, Reference Mellié1847 from the Orophiinae, nor Sulcacis Dury, Reference Dury1917 from the Ciinae. The remains of the new species resemble representatives of the mentioned genera in the body size, proportions of evenly sloping head, pronotum and elytra. However, the outline of apparent elytral apices, puncturation and comparatively elongate frons of the new genus are more like those in Octotemnus than the other genera. Sculpture of the interspaces between punctures on the dorsum looks rather coarse, not smooth as in most Recent species of Octotemnus. Finally, the remains examined show extremely short scutellum, although this sclerite is usually longer in most Recent representatives of the family.
Octotemnites sepultus Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:E9264FB4-36D9-468A-9C48-A0B4FE9A5893
Derivation of name. The epithet of this new species means ‘buried', ‘fossil'.
Material. Holotype NHMUK I. 8774 (Figs 36d, 40): negative print showing dorsal surface (head, pronotum and elytra with remains of integument) embedded in the limestone matrix; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. Holotype I. 8774: length of print 1.3mm, width 0.5mm, length of elytra 0.8mm. Elliptical and rather convex dorsally; dark brown in colouration. Integument on elytra with moderately large and not dense, not quite distinct, punctures and apparently alutaceous to microreticulated on interspaces; that on pronotum with rather fine and dense, not quite distinct, punctures and apparently alutaceous to microreticulated on interspaces. Head not very wide, subtriangular, evenly sloping anteriorly, mandibles moderately developed and simple. Pronotum subquadrangular and with rounded angles, gradually and steeply sloping at sides and gently sloping anteriorly. Scutellum transverse, subtriangular to widely rounded at apex. Elytra about 1.5 times as long as combined width; subparallel-sided along most of length; their sides gradually and very steeply sloping; apices also evenly and gently sloping.
Discussion. The specimen examined is unlikely to enter the place of deposition of the Bembridge Marls, because all Recent species of minute tree fungus beetles, and probably this new fossil, are associated with forestry landscapes.
Family Anthicidae Latreille, Reference Latreille1819
(Figs 36e, f, 41a, 42)
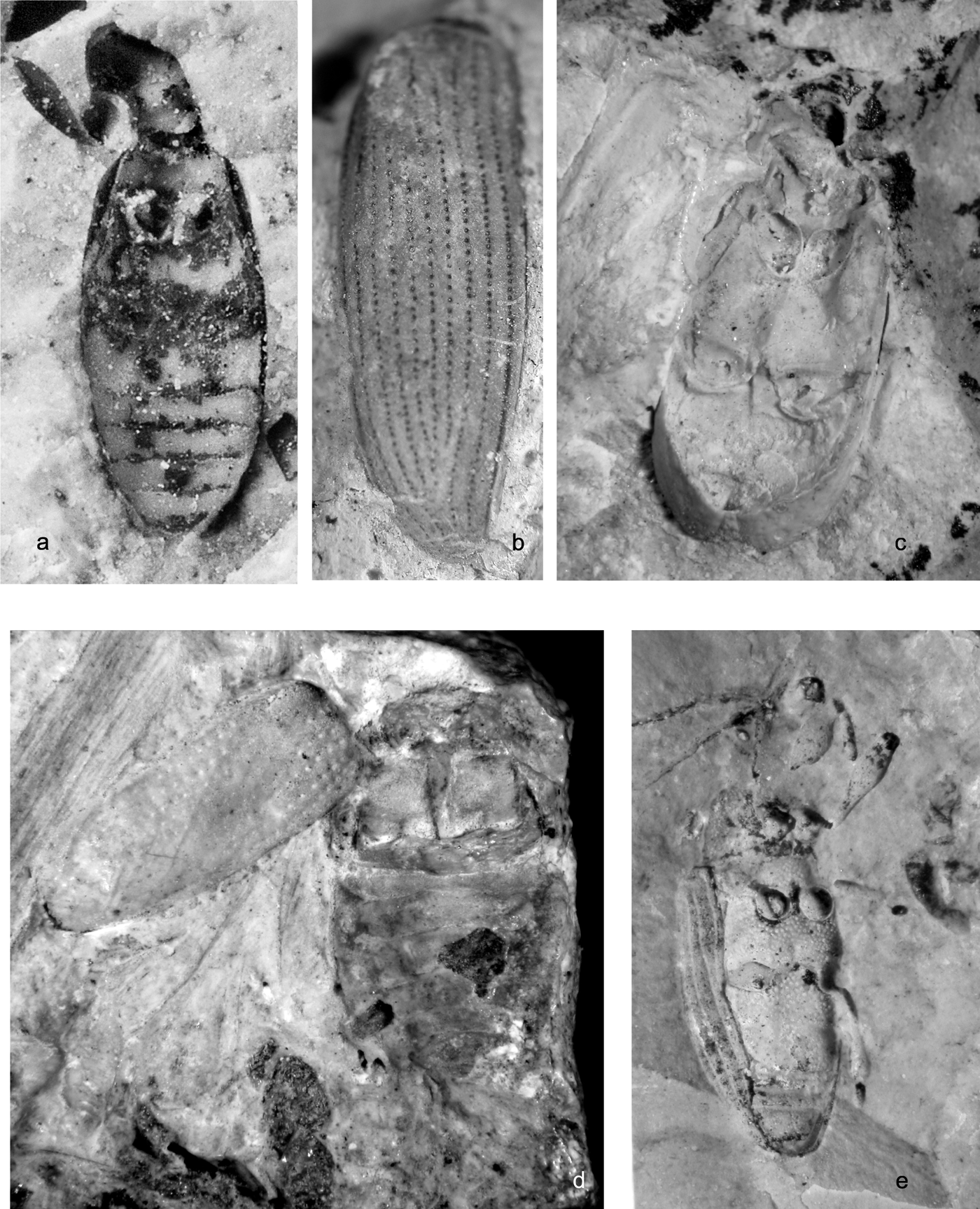
Figure 41 Beetles of families Anthicidae, Chrysomelidae and Curculionidae. (a) Anthicidae, gen. & sp. indet.: NHMUK I. 8500, body, prothoracic segment and anterior leg, lateral view, pterothorax and abdomen, ventral view; length of specimen = 2.5mm. (b) Plateumaris (Plateumaris) wightensis Kurochkin & Kirejtshuk, sp. nov. (Chrysomelidae): holotype, NHMUK I. 10259, elytron, dorsal; length of specimen = 5.8mm. (c) Plateumaris (Necrodexis) rubiconis Kurochkin & Kirejtshuk, subgen. & sp. nov. (Chrysomelidae): holotype, NHMUK I. 10326, body, ventral; length of specimen = 5.1mm. (d) Plateumaris (Plateumaris) robustus Kurochkin & Kirejtshuk, sp. nov. (Chrysomelidae): holotype, In. 25763, body, dorsal view; length of specimen = 4.25mm. (e) Curculionidae, gen. & sp. indet. (probable member of the subfamily Molytinae or Cleoninae Schönherr, Reference Schönherr1826): NHMUK I. 10182, body, ventral view; length of specimen = 5.4mm.
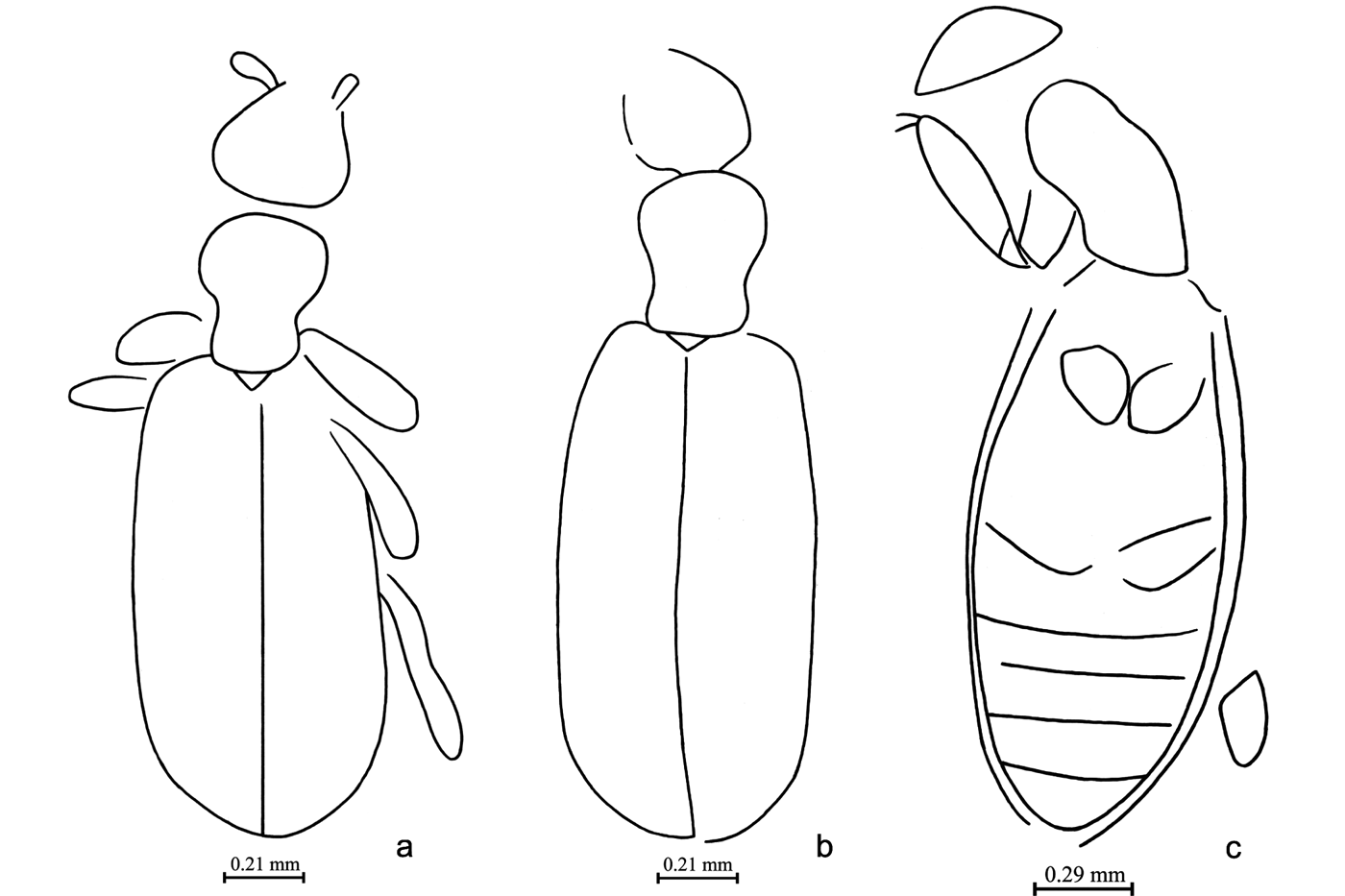
Figure 42 Suborder Polyphaga, family Anthicidae. (a) Holotype of Cyclodinus efficax Kirejtshuk, sp. nov., NHMUK I. 9457, body, dorsal view. (b) Paratype of C. efficax sp. nov., NHMUK I. 10259, body, dorsal view. (c) Gen. & sp. indet., NHMUK I. 8500, body, ventral view.
Material. Five specimens (identified by A. G. Kirejtshuk).
This small group consists of small- and medium-sized beetles (usually 1.5–5.5mm, sometimes up to 17.0mm), usually with a narrow pronotum and very narrow ‘neck' of the head, and, therefore, its representatives frequently have an appearance reminiscent of ants; pronotum generally tapers strongly towards the back; antennae are rather long; legs are long and thin, with only the femora relatively stout; tarsi more widened (at least pro- and mesotarsomeres 4 as well as metatarsomeres 3 are lobed). Recent representatives of the family are found in all zoogeographical regions. Larvae and adults of most species are mycetophagous (although a few are predaceous) and live in decaying matter of plant origin, frequently in leaf litter and other kinds of debris; some of them are characteristic of the edges of lakes and other bodies of water; adults of them sometimes emerge in great number. Fossils of this family are known mostly from Baltic amber (Upper Eocene) (Klebs Reference Klebs1910; Abdullah Reference Abdullah1964; Larsson Reference Larsson1978; Spahr Reference Spahr1981; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984; etc.), where the subfamilies Macratriinae LeConte, Reference LeConte1862, Steropinae Jacquelin du Val, Reference Jacquelin du Val1863, Tomoderinae Ranadona, 1961 and Anthicinae (including Notoxini Stephens, Reference Stephens1829) have been recorded. The oldest records of this family are found in Lebanese amber (Lower Cretaceous) (Kirejtshuk & Azar Reference Kirejtshuk and Azar2008) and Burmese amber (boundary of Lower/Upper Cretaceous) (Cockerell Reference Cockerell1917a: Eurygeniinae LeConte, Reference LeConte1862). Caenozoic records came from Sunchal (Palaeocene or Eocene?) (Cockerell Reference Cockerell1926), Oise amber (lowermost Eocene) (Kirejtshuk & Nel Reference Kirejtshuk and Azar2008), Florissant shales (Upper Eocene) (Wickham Reference Wickham1910), Izarra beds (Oligocene) (Arillo & Ortuno Reference Arillo and Ortuno1997) and Lower Oligocene of Aix (Oustalet Reference Oustalet1874).
Notes. Young (Reference Young1984) described the member of Pedilinae Lacordaire, Reference Lacordaire1859 sensu lato (as Pyrochroidae Latreille, Reference Latreille1807) genus Anisotria Young, Reference Young1984. He also transferred Lithomacratria mirabilis Wickham, Reference Wickham1914 from the Florissant shales, which has the same type of antennae as in the Pedilinae. However, this transfer is not reasonable, because according to the characters in the description, Lithomacratria mirabilis clearly does not have an elongate oval pronotum as in Macratria LaFerté-Senectère in Guérin-Méneville, Reference LaFerté-Sénectère1849 and Anisotria (in Macratriinae, pronotum is cylindrical or distinctly elongate oval; in Anisotria, pronotum strongly resembles that of Pedilus – short and broad), but the pronotum in this fossil species is widest at base and gently narrowing anteriorly, which means that Lithomacratria can be scarcely put into either Anthicidae or Pyrochroidae. Therefore, it should be regarded as Tenebrionoidea incertae sedis until further re-examination of the type specimen.
Subfamily Anthicinae Latreille, Reference Latreille1819
Genus Cyclodinus Mulsant & Rey, Reference Mulsant and Rey1866
Type species. Anthicus humilis Germar, Reference Germar1824, Recent.
This genus includes more than 90 species in the Recent fauna, of which 64 occur in the Palaearctic Region. The specimens examined show only the characters of the dorsal surface; however, the configuration of their visible sclerites, particularly the outline of the pronotum, is characteristic of species of the genus Cyclodinus. Recent members of the genus Cyclodinus are easily recognised by their distinctive dorsal habitus: glossy body with sparse setae, pronotum distinctly constricted in basal half and often with two small tubercles on the base (all from the dorsal view), pronotal disc is punctured increasingly densely toward the base of pronotum, tuft of setae is presented on the linear mesepisternum. The bionomy of Recent members of this genus seems to be similar to that in other groups formerly included in the genus Anthicus Paykull, Reference Paykull1798. Species of Cyclodinus frequently occur on marine beaches or near inland saline lakes.
Cyclodinus efficax Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:13801F96-214A-4653-AA30-2D2E28318BCC
Derivation of name. The epithet of this new species means ‘efficient', ‘successful', ‘convincing' and ‘persuasive'.
Material. Holotype NHMUK I. 9457 (Figs 36e, 42a): dorsal surface of body with somewhat detached and rather declined head; the head, anterior part of pronotum and left elytron are partly embedded in the limestone and with indistinct sides; both scapes, right meso- and metafemora as well as all left femora exposed; Insect Limestone, Isle of Wight, Brodie Collection.
Paratype. NHMUK I. 10259 (Figs 36f, 42b): dorsal surface of body without appendages; Insect Limestone, NW Isle of Wight, Brodie Collection.
Diagnosis. This new species differs from many Recent species in the very wide head, widely transverse posterior edge of head before the ‘neck' and comparatively dark colouration of the head and pronotum (while the elytra are unicolorous). The elytral apices of the new species are conjointly widely rounded. There are some European species (e.g., Cyclodinus minutus (La Ferté-Sénectère, 1842)), Nearctic and Centroasiatic species of Cyclodinus with widened posterior edge of head and comparatively dark colouration of head and pronotum (while the elytra are unicolorous). However, in all of these Recent species the head is not wider than the pronotum.
Description. Holotype I. 9457: length of specimen 2.0mm, width 0.6mm, length of elytra 1.4mm. Body elongate, apparently rather convex dorsally and moderately convex ventrally; reddish with somewhat darker pronotum and head. Head and pronotum apparently with smoothed integument; elytra with rather fine and densely diffuse punctures. Head rather elongate (somewhat longer than wide and strongly depressed in limestone) and rounded at sides (gently arcuate at base), rather convex and very narrowed at very short neck (scarcely traceable). Scape rather long (about three times as long as wide). Pronotum somewhat narrower than head, slightly longer than wide (taking into consideration the part embedded in limestone), strongly convex; widest at anterior half and narrowing before lateral sinuations at base (widest place about one and a third as wide as narrowest place); anterior edge moderately rounded and posterior edge subtransverse; anterior and posterior angles gently rounded. Scutellum widely subtriangular. Elytra about twice as long as wide combined; rather narrowed at base, subrectilinear along the middle and gently arcuate at apices, almost twice as wide as pronotum, apparently steeply sloping at sides, complete and covering apex of abdomen, their apices seeming to form a conjoint arc. Protibia moderately long and apparently widest in distal half, about as long as pronotum, and right femur about three times as long as wide. Mesofemur somewhat longer than profemur and metafemur somewhat longer than mesofemur.
Variation. Length of print 2.1mm, width 0.7mm, length of elytra 1.3mm. Head about as wide as pronotum and with neck apparently about a third as wide as head.
Gen. & sp. indet.
Material. NHMUK I. 8500 (Figs 41a, 42c): underside from inside, including the head oriented inside the limestone, prosternum obliquely deepened in the limestone, right procoxa, profemur and base of protibia, negative print of inner side of meso- and metaventrites, also abdomen and a part of left metafemur; Insect Limestone, NW Isle of Wight, Brodie Collection. NHMUK I. 9953 (counterpart of I.8500): upperside from inside, including the pronotum, scutellum and elytra, base of both mesofemora, profemur and base of protibia and left metafemur; Insect Limestone, NW Isle of Wight, Brodie Collection.
NHMUK I. 8770, underside from inside, including the metaventrite and abdomen, and also some parts of mouthparts, apex of right protibia and protarsomeres 1–2, part of prosternum, procoxa, parts of both mesofemora, part of right mesotibia and right metafemur; Insect Limestone, NW Isle of Wight, Brodie Collection.
Description. I. 8500: length of print 2.5mm (apparently 2.9mm), width 0.8mm, length of elytra 1.6mm. Body elongate, apparently rather convex dorsally and moderately convex ventrally; reddish. Ventral sclerites of meso- and metaventrites and abdomen apparently with rather fine and densely diffuse punctures. Head apparently elongate, gently convex dorsally and flattened ventrally. Pronotum markedly longer than wide, strongly convex; widest at anterior half and narrowing before lateral sinuations at base. Elytra about twice as long as wide combined; widest at anterior two-fifths, narrowing anteriorly and posteriorly, almost twice as wide as pronotum, apparently steeply sloping at sides, complete and covering apex of abdomen, their apices seeming to form a nearly conjoint arc. Procoxa rather projecting. Mesocoxae almost contiguous. Profemur moderately long, of usual shape and somewhat shorter than pronotum. Base of protibia rather narrow. Metafemur somewhat wider than profemur and of usual shape. I. 9953: sculpture of integument seems to be more or less smoothed and without distinct punctures. Scutellum nearly subsemicircular. Elytral apices conjointly acuminate. Mesofemora about as wide as pro- and metafemora. I. 8770: protarsomeres 1–2 subconical and almost twice longer than wide at apex.
Notes. The remains examined seem to belong to a species of Anthicidae because of the characteristic shape of prothoracic segment, outline of cross-section of head base, outline of the elytra and strongly projecting procoxae. The rest of the characters available in this specimen are also similar to those in members of the subfamily Anthicinae. The comparatively long pronotum of the specimen under consideration is reminiscent of some genera in the Recent fauna, like Stenidius La Ferté-Sénectère, 1849; however, the visible characters are not enough to positively identify the genus and species. These remains are similar to species of the genus Cyclodinus, but it is impossible to be certain. The specimen cannot belong to Cyclodinus efficax sp. nov. because of the longer pronotum and different outline of elytra.
Discussion. Adults of the Recent Anthicidae have an open mode of life on vegetation and, therefore, the remains examined could enter the depositional area of the Bembridge Marls occasionally. Species of Cyclodinus frequently occur on marine beaches or near inland saline lakes and could indicate this type of palaeoenvironment and landscape surrounding the places of deposition.
Superfamily Chrysomeloidea Latreille, Reference Latreille1802
Family Chrysomelidae Latreille, Reference Latreille1802

Figure 43 Suborder Polyphaga, family Chrysomelidae. (a) Holotype of Plateumaris (Plateumaris) wightensis Kurochkin & Kirejtshuk, sp. nov., NHMUK, In. 25518, elytron, dorsal view. (b) Holotype of Plateumaris (Plateumaris) robustus Kurochkin & Kirejtshuk, sp. nov., NHMUK In. 25763, pterothorax, abdomen, elytron and posterior wing, dorsal view. (c) Paratype of P. (P.) robustus sp. nov., NHMUK, In. 24439, elytron, dorsal view. (d) Holotype of Plateumaris (Necrodexis) rubiconis Kurochkin & Kirejtshuk, subgen. & sp. nov., NHMUK I. 10326, body, ventral view.
Material. A total of 64 specimens (identified by A. G. Ponomarenko, A. S. Kurochkin and A. G. Kirejtshuk).
Leaf beetles form one of the most diverse and numerous families in the Recent fauna, but they are not very common as fossils. They are characterised by the usually oval and convex body, (sub)hypognathous head with antennal insertions located at dorsal surface of frons, antennae filiform to serrate and not disposed ventrally, oval pro- and mesocoxal cavities, pseudotetramerous wide tarsi. Most Recent species feed on green parts of plants and are distributed worldwide. The oldest remains of leaf beetles are from the Upper Jurassic or Lower Cretaceous (Kirejtshuk et al. Reference Kirejtshuk, Moseyko and Ren2015). However, the published data on fossils of Chrysomelidae in general are quite ambiguous (Moseyko et al. Reference Moseyko, Kirejtshuk and Nel2010; Moseyko & Kirejtshuk Reference Moseyko and Kirejtshuk2013; Valentin et al. Reference Valentin, Gomez, Daviero-Gomez, Licht, Kirejtshuk, Ferchaud, Neraudeau, Vullo and Garcia2014; etc.). In general, this family seems to represent one of the rather old cucujiformian groups (Kirejtshuk Reference Kirejtshuk1994). Wolf-Schwenninger & Schawaller (Reference Wolf-Schwenninger, Schawaller, Martill, Bechly and Loveridge2007) recorded leaf beetle remains of the subfamily Eumolpinae Hope, Reference Hope1840 from the Santana Formation of Brazil (Lower Cretaceous). Kirejtshuk et al. (Reference Kirejtshuk, Ponomarenko, Prokin, Chang, Nikolajev and Ren2010b) recorded one chrysomeloid group from Yixian (Lower Cretaceous). Valentin et al. (Reference Valentin, Gomez, Daviero-Gomez, Licht, Kirejtshuk, Ferchaud, Neraudeau, Vullo and Garcia2014) recorded the subfamily Chrysomelinae from compression deposits of Jaunay-Clan (near Poitiers, SW France; Upper Cretaceous). A more detailed review of the fossil record is given in the catalogue by Kirejtshuk & Ponomarenko (Reference Kirejtshuk and Ponomarenko2018). In the Caenozoic, different chrysomelid groups were found in lowermost Eocene Oise amber (Moseyko et al. Reference Moseyko, Kirejtshuk and Nel2010; Kirejtshuk & Nel Reference Kirejtshuk and Nel2013) and Upper Eocene Baltic amber (Klebs Reference Klebs1910; Spahr Reference Spahr1981; Hieke & Pietrzeniuk Reference Hieke and Pietrzeniuk1984; Bukejs & Nadein Reference Bukejs and Nadein2013; Moseyko & Kirejtshuk Reference Moseyko and Kirejtshuk2013) and Rovno amber (Nadein & Perkovsky Reference Nadein and Perkovsky2010). This family is present in some Caenozoic sedimentary sites, and particularly abundant in Geiseltal near Halle (Middle Eocene) (Haupt Reference Haupt1956) and Florissant shales (Upper Eocene) (Wickham Reference Wickham1910, Reference Wickham1912, Reference Wickham1913, Reference Wickham1914a). The subfamily Donaciinae Kirby, Reference Kirby and Richardson1837 was already recorded in the Upper Cretaceous (Prokin & Ponomarenko Reference Prokin, Ponomarenko, Prokin, Petrov, Zhavoronkova and Tuzovskij2013: Altan-Ula, Southern Mongolia) and it seems to be more common in compression deposits of the second half of the Palaeogene and later outcrops (Heer Reference Heer1847; Förster Reference Förster1891; Wickham Reference Wickham1912; Théobald Reference Théobald1937; Goecke Reference Goecke1943; Reference Goecke1960; Haupt Reference Haupt1956; Askevold Reference Askevold1990a; Hayashi Reference Hayashi1998, Reference Hayashi1999, Reference Hayashi2000; etc.). Of more than 60 remains of leaf beetles found in the Bembridge Marls Beds, the majority belong to the subfamily Donaciinae (four species according to the size and elytral structure, the most common is Plateumaris wightensis sp. nov., the possible host plants of which are Typha), while four specimens seem to belong to one species of Chrysolampra Baly, Reference Baly1859 (Eumolpinae). Such predominance of only one subfamily is characteristic of the Pliocene–Pleistocene swamp deposits in Japan (Hayashi Reference Hayashi1998, Reference Hayashi1999, Reference Hayashi2000). Today, this group occurs mostly in areas of Eurasia with moderate and subtropical climate, and a small number of them occur in other areas, except in South America where they are not registered. Thus, leaf beetles of the examined sites are characteristic of the moderate zone rather than the tropical one.
Subfamily Donaciinae Kirby, Reference Kirby and Richardson1837
This subfamily occurs as fossils from the Late Cretaceous (Prokin & Ponomarenko Reference Prokin, Ponomarenko, Prokin, Petrov, Zhavoronkova and Tuzovskij2013); they are uncommon in the Palaeogene and became quite common in the Neogene. There are several dozen Recent species known, mostly in Eurasia, and several species in other continents, except South America. Some species of this group sometimes inhabit the same locality.
Tribe Plateumarini Askevold, Reference Askevold1987
Genus Plateumaris Thomson, Reference Thomson1859
Type species. Donacia nigra Fabricius, Reference Fabricius1792 (original designation by Thomson, Reference Thomson1859), Recent.
Remarks. Forty-eight isolated elytra are placed in the genus Plateumaris of the subfamily Donaciinae due to the characteristic punctato-striae elytra with arcuate apices, well-expressed adscutellar striola with more or less distinct punctures and smooth platform along the sutural margin just before the apices. Their elytra are convex and narrow, with ten longitudinal rows of more or less deep punctures. The length of elytra is 3–4 times as long as wide. The elytral apices are without interlocking tongue. The epipleura is absent or incomplete. The lateral elytral margin is weakly convex. Procoxae are contiguous, pro- and mesocoxae globose, metacoxae transverse. The Recent representatives of this tribe are characterised by the sutural edge at the elytral apices clearly explanate innerly, inner sutural bead sinuate far from apex and joined to outer bead, and elytral apices rounded in most species, sutural interval narrowing subapically; mesoventrite narrow between mesocoxae; prohypomera are glabrous or with few scattered setae. Beetles usually feed on Cyperaceae, but also on a few other emergent plant types.
Subgenus Plateumaris Thomson, Reference Thomson1859
Type species. As for the genus.
Plateumaris (Plateumaris) wightensis Kurochkin & Kirejtshuk, sp. nov.
Derivation of name. This new species is named after the toponym of the Isle of Wight.
Zoobank ID. urn:lsid:zoobank.org:act:A951526B-1CD9-40F4-B4B2-89FC382D1DFF
Material. Holotype NHMUK I. 25518 (Figs 41b, 43a): positive print of the left isolated elytron; Insect Limestone, NW Isle of Wight, Hooley Collection.
Paratypes (separate elytra). All from the NHMUK collection, Insect Limestone, NW Isle of Wight: I. 8606, Brodie Collection; I. 10286, isolated elytron of Plateumaris wightensis sp. nov. together with one elytron of Chrysomelidae incertae sedis and four elytra incertae sedis, Brodie Collection; In. 25583, In. 25615, In. 25639, In. 25667, In. 25681 and In. 26085, isolated elytra, Hooley Collection.
Diagnosis. This new species has elytra very similar to Donacia primaeva Wickham, Reference Wickham1912 from Upper Eocene Florissant, USA (Wickham Reference Wickham1912), which was later re-examined and transferred to the genus Plateumaris by I. S. Askevold (Reference Askevold1990b), because it shows a rather characteristic shape of elytra and comparatively long scutellary striola, but the new species differs from the American one in the wider elytra and more widely rounded elytral apices. Plateumaris (Plateumaris) wightensis sp. nov. is rather similar to P. (P.) robustus sp. nov. but differs from the latter in the character mentioned in its diagnosis. Both are also similar to P (P.) fallax Haupt, Reference Haupt1956 from Geiseltales (Eocene) of Germany, but differs from the latter in its elytral ratio, more narrow curve at elytral apices. Plateumaris (Plateumaris) wightensis sp. nov. is also somewhat similar in its elytral characters to some Recent species, such as the following:
Plateumaris (Plateumaris) constricticollis (Jacoby, Reference Jacoby1885), but differs in its smaller and much denser, oval and shallower punctures on the elytra, which are separated by about a puncture diameter, smooth interspaces between the elytral punctures and expressed striae at apices of elytra;
Plateumaris (Plateumaris) flavipes Kirby, Reference Kirby and Richardson1837, but differs in its smaller and much denser round and shallower punctures on elytra, which are separated by about a puncture diameter, and smoother interspaces between these punctures;
Plateumaris (Plateumaris) rufa (Say, Reference Say1826), but differs in its smaller and much denser round and shallower punctures on elytra, which are separated by about a puncture diameter, more clearly rounded elytral apices;
Plateumaris (Plateumaris) sericea (Linnaeus, Reference Linnaeus1761), but differs in its smooth interspaces between punctures on elytra, lack of clear transrugosity on elytral integument and distinct striae at elytral apices.
Description. Holotype I. 25518: length of elytron about 5.8mm, width 2.0mm. Elytra wide, tapering and rather widely rounded at apices. Ratio of elytral length to width about 3:0. Three rows along sutural edge and two along outer edge, longitudinally orienting and disposing along nearly entire elytra, fourth row from the sutural edge meeting with the fifth, fifth with sixth, sixth with seventh and seventh with eighth. Striae not deep, the strial punctures round, very distinct, rather deep and becoming finer apically. Distance between strial punctures range from almost confluent punctures to being separated by about three puncture diameters; intervals between striae 2–6-puncture diameters. Epipleura narrow.
Variations (paratypes). Length of elytra 5.8–7.5mm, width of elytron 2.0–2.5mm.
Probable bionomy. Plateumaris is a group of phytophagous water beetles; Recent species live in stagnant freshwater bodies, larvae commonly found on roots of water plants. Its adults are primarily floricolous.
Plateumaris (Plateumaris) aff. wightensis sp. nov.
Material (mostly separated elytra). Insect Limestone, NW Isle of Wight: NHMUK I. 10176 (isolated elytron of Plateumaris species together with one isolated elytron of Curculionoidea); NHMUK I. 8609, Brodie Collection; NHMUK In. 24389; NHMUK In. 25596; NHMUK In. 26060; In. 25685 (isolated elytron of Plateumaris species together with elytron of Curculionidea; two elytra Coleoptera incertae sedis and one isolated elytron of Dytiscidae); NHMUK In. 25532 (isolated elytron of Plateumaris species together with three elytra of Coleoptera incertae sedis); NHMUK In. 25590 (one isolated elytron of Plateumaris species together with two isolated elytra of Coleoptera incertae sedis); NHMUK In. 24851 (1), (2) (two isolated elytra of Plateumaris species together with two isolated elytra of Curculionidea and one isolated elytron of Coleoptera incertae sedis); NHMUK In. 25686 (3), (4) (two isolated elytra of Plateumaris species together with two negative prints of bodies of Scirtidae, 14 isolated elytra of Coleoptera incertae sedis, one metathorax of Coleoptera incertae sedis and one isolated abdomen of Coleoptera incertae sedis); NHMUK In. 26155 (one isolated elytron of Plateumaris species together with one isolated elytron of Coleoptera incertae sedis); NHMUK In. 26076 (1), (2) (two isolated elytra of Plateumaris species together with isolated pterothorax and abdomen of Curculionidea); NHMUK In. 25605 (Fig. 23c, one isolated elytron of Plateumaris species together with one isolated elytron of Curculionoidea and one isolated elytron of Carabidae); NHMUK In. 25643; NHMUK In. 26132 (one elytron of Plateumaris species together with one elytron of Coleoptera incertae sedis); NHMUK In. 25588; NHMUK In. 26160; NHMUK In. 25545; NHMUK In. 25514, Hooley Collection; NHMUK In. 2580; NHMUK In. 20552 (one body of Plateumaris species together with one body of Curculionidea, four isolated elytra of Coleoptera incertae sedis and one body of Coleoptera incertae sedis), Smith Collection.
Notes. Most specimens listed above were preliminarily studied by the authors. They seem to belong to Plateumaris (Plateumaris) sp. nov., although they need to be additionally examined to check their species attribution. They could belong to other species of the genus Plateumaris, different from the species considered here, because they differ slightly from the elytra of the type series of the new species in size and shape.
Plateumaris (Plateumaris) robustus Kurochkin & Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:541E1084-F4CE-4145-89E5-1647342A616D
Derivation of name. The epithet of this new species means ‘robust' and refers to its comparatively wide and subtruncate elytra.
Material. Holotype NHMUK In. 25763 (Figs 41d, 43b, c): (underside) elytron (inner view), remains of meso- and metaventrites, abdomen, remains of posterior wing with some veins; Insect Limestone, Isle of Wight, Hooley Collection.
Paratypes. NHMUK In. 25669, positive prints of isolated elytra; Insect Limestone, NW Isle of Wight, Hooley Collection.
Diagnosis. This new species is very close to Plateumaris wightensis sp. nov. (see description and diagnosis in previous section), but can be distinguished from it by its more robust and wider elytra (elytral ratio) with subtruncate and more widely rounded apex, larger, deeper and less numerous punctures on elytra. Plateumaris robustus sp. nov. bears some similarity to Plateumaris primaeva and Plateumaris fallax, but differs from both these fossil species in its more robust and wider elytra (elytral ratio), with subtruncate and more widely rounded apex, and from the first also in the much less convex anterior edge of metafemur.
Description. Holotype In. 25763: length of elytron 3.6mm, width 1.4mm; length of body 4.3mm, width 2.3mm. Elytron with ten subparallel striae and widely rounded at apex. Inner edge of elytron very slightly emarginate before its apex. Ratio of elytral length to width apparently about 2:6. Punctures of elytra round, distinct and separated by 1.5–3.0 puncture diameters, intervals between striae about 2–4 puncture diameters. Distal part of posterior femur demonstrating slightly convex anterior and apparently straight posterior edges. Ventrites strongly transverse and subequal in length. Hypopygidium apparently widely rounded at apex.
Paratype In. 24439: elytron rather wide and subparallel-sided, with very gently sloping sides, subtruncate and widely rounded at apex. Strial punctures large, distinct, round and very deep, becoming finer apically. Punctures variable in size, punctures of first and second rows from outer edge as well as of first and second rows from inner edge slightly smaller than the remainder. Distance between strial punctures from almost confluent to four puncture diameters; intervals between striae about 2–4 puncture diameters.
Subgenus Necrodexis Kurochkin & Kirejtshuk, gen. nov.
Zoobank ID. urn:lsid:zoobank.org:act:650EBF77-96B1-4E33-BABF-7674D597C578
Type species. Plateumaris (Necrodexis) rubiconis sp. nov.
Derivation of name. The name of this new subgenus is formed from the Greek ‘necros' (dead) and ‘dexis' (increase, increment, meeting, reception). Gender: masculine.
Note. This genus is represented by the only species and, therefore, the description of it overlaps with the description of species.
Diagnosis. The new species under consideration (type species of the new subgenus) is rather similar to other species of the genus Plateumaris, although, in contrast to the latter, it has contiguous mesocoxae, close metacoxae, very deep and comparatively narrow depression along the middle of metaventrite and abdominal base. In addition, the punctures in elytral striae of this new species are rather shallow and much finer than those in the species of Plateumaris. Contiguous or subcontiguous mesocoxae are a unique character of this new subgenus among members of the subfamily Donaciinae. Therefore, the authors consider it quite reasonable to regard this new species as a member of a separate taxon, closer to the Plateumaris than any other groups within the subfamily Donaciinae, and to give a subgeneric rank to it. Moreover, this new subgenus is also quite distinct from the second genus, Poecilocera Schaeffer, Reference Schaeffer1919, of the tribe Plateumarini, in having widely arcuate apices of elytra and contiguous mesocoxae.
The examined print of Plateumaris (Necrodexis) rubiconis sp. nov. is characterised by the peculiar shape of elytra and metaventrite and elytral punctures arranged in longitudinal rows. However, it has some degree of similarity with the Recent leaf beetles of the subfamilies Criocerinae Latreille, Reference Latreille1804 and Orsodacninae Thomson, Reference Thomson1859. In contrast to the new genus under consideration, the members of the first subfamily have moderately separated meso- and metacoxae, strongly developed shoulder tubercles of the elytra and lack the secondary sexual dimorphism in the structure of metaventrite. The puncturation of the elytra in species of this subfamily (Criocerinae) are arranged in longitudinal rows with very sparse punctures (markedly sparser than in Plateumaris and other Donaciinae). Also, members of the second subfamily (Orsodacninae) have very narrowly separated to almost contiguous pro-, meso- and metacoxae, their metaventrite is flat to slightly convex and elytra have irregular puncturation.
Notes. Sexual dimorphism in the structure of metaventrite, base of abdomen and hypopygidium is quite characteristic of Donaciinae; however, the level of expression of such dimorphism in Plateumaris (Necrodexis) rubiconis subgen. & sp. nov. is much greater than in any group of the subfamily.
Plateumaris (Necrodexis) rubiconis Kurochkin & Kirejtshuk, sp. nov.
Zoobank ID. urn:lsid:zoobank.org:act:1C7C2221-54A0-41CB-A067-1EC085E27D9D
Derivation of name. The name of this new species is formed from an ancient Latin name for a small river Rubicon in northern Italy, known from the legendary phrase of Julius Caesar, and refers to the boundary between the Eocene and Oligocene.
Material. Holotype NHMUK I. 10326 (Figs 41c, 43d): negative print (inner view) of the body of Plateumaris species with missing head, but with well-visible apices of elytra (dorsal view), together with one pterothorax and abdomen of a small Curculionoidea, print of underside of Coleoptera incertae sedis, two pairs of elytra of Coleoptera incertae sedis; Insect Limestone, Isle of Wight, Brodie Collection.
Description. Holotype (male) I. 10326: length of specimen 5.1mm, width 2.3mm. Elytra with apparently gently sloping sides and with longitudinal rows consisting of round, small, rather shallow and quite distinct punctures, separated by 2.0–2.5 puncture diameters, intervals between rows about as great as 4–5 puncture diameters. Elytra with rather narrowly rounded apices and forming a distinct sutural angle, their maximum length at suture. Epipleura rather narrow. Pro- and mesocoxae moderately large, subovoid and oblique. Mesocoxae about 1.4 times as large as procoxae. Both pro- and mesocoxae contiguous. Metacoxae strongly transverse, narrowly separated, and with almost straight anterior and moderately convex posterior margin. Metepisterna distinctly narrowing posteriorly. Metaventrite deeply and narrowly depressed along the entire length, its posterior edge angularly and rather deeply excised. Visible ventrites subequal in length, basal ventrites with one clear depression looking like a continuation of that in the middle of metaventrite. Abdominal apex apparently narrowly rounded (assuming that the fossil continues inside the limestone, where apparently the carinate apex of hypopygidium can be seen from inside a hole in the limestone).
Notes. Plateumaris (Necrodexis) rubiconis gen. & sp. nov. is externally somewhat similar to Plateumaris primaeva, but easily recognisable due to its unseparated pro- and mesocoxae, much narrower separated metacoxae and more narrowly rounded apex of abdomen. This new species is different from the Eocene Plateumaris fallax in its smaller size of body and elytra, more narrowly rounded apex of elytra and much smaller and shallower punctures on elytra.
Plateumaris spp.
Identified material (mostly separated elytra). All from the Insect Limestone, NW Isle of Wight: NHMUK In. 25550 (a part of isolated elytron of Plateumaris species); In. 43404 (one isolated elytron of Plateumaris species together with one abdomen of Hydrophilidae, large ventrites of Coleoptera incertae sedis and two isolated elytra of Curculionoidea); NHMUK In. 25662 (one isolated elytron of Plateumaris species); NHMUK In. 25674 (one isolated elytron without base and apex apparently larger than in Plateumaris wightensis sp. nov.); NHMUK In. 25078 (one isolated elytron of Plateumaris species without base and apex, together with one large elytron of Coleoptera incertae sedis); NHMUK In. 25680 (one basal part of elytron of large Donaciinae (could be Plateumaris species), which should be about twice as large as that in Plateumaris (Plateumaris) wightensis sp. nov.); NHMUK In. 25648 (one lateral part of elytron of large Donaciinae (could be of Plateumaris species), which could be about twice as large as that of Plateumaris (Plateumaris) wightensis sp. nov.); NHMUK In. 26170 (one separated elytron of Plateumaris species, together with three separated elytra of Coleoptera incertae sedis and one elytron of Curculionoidea), Hooley Collection.
Discussion. All Recent representatives of the subfamily Donaciinae are known to occur near rivers, lakes, swamps, etc., where they feed on the emergent leaves of semiaquatic and aquatic plants (such as Brasenia spp., Caltha spp., Carex spp., Phragmites spp., Potamogeton spp., Ruppia spp., Sagittaria spp., Scirpus spp., Sparganium spp., Typha spp., Nuphar spp., etc.), and frequently reach a very high density. Their larvae develop on leaves and underwater rhizomes of these plants as well. Therefore, it is very likely that the extinct new species of Plateumaris described herein could live under lake or swamp conditions in or close to the depositional environment of the Bembridge Marls. Numerous findings of the Typha remains, together with the remains of Plateumaris species, support this hypothesis. Most likely, both larvae and adults of the new species were strictly associated with Typha plants. A rather peculiar feature of the Bembridge Marls is the dominance of Plateumaris among Donaciinae, while other Caenozoic sites with Donaciine fossils include mostly species of the following genera: Donacia Fabricius, Reference Fabricius1775; Eodonacia Haupt, Reference Haupt1956; Hemidonacia Haupt, Reference Haupt1956; Donaciella Reitter, Reference Reitter1920; and Macroplea Samouelle, Reference Samouelle1819.
Superfamily Curculionoidea Latreille, Reference Latreille1802
(Fig. 41e)
Material. A total of 226 specimens (identified by V. G. Gratshev, A. G. Kirejtshuk, A. G. Ponomarenko and A. S. Kurochkin).
This superfamily is the largest among Recent Coleoptera, with a very wide distribution, and it is extremely diverse in ecology and bionomy. In most cases, Recent species have a small- to medium-sized and very heavily sclerotised body, head characteristically with a rostrum, distinctively shaped elytra usually with longitudinal rows of punctures on them, consolidated ventrites with first ones very frequently fused, pro- and mesocoxae narrowly separated, oval and rather small. Most of them are phytophagous, feeding on living and dead parts of plants, although some are mycetophagous. Curculionoids live in green vegetation, soil, leaf litter, wood and other habitats. They are more numerous in dry localities, and have a comparatively small representation in water basins. This superfamily seemed to originate in the Upper Triassic or Upper Jurassic (depending on systematic placement of the family Obrienidae: among Archostemata or Polyphaga). The true weevils or Curculionidae sensu lato appeared at the end of the Lower Cretaceous (although Scolytinae are known from the early Lower Cretaceous: both beetles – Cockerell Reference Cockerell1917b; Kirejtshuk et al. Reference Kirejtshuk, Azar, Tafforeau, Boistel and Fernandez2009b – and holes – Brongniart Reference Brongniart1877; Jarzembowski Reference Jarzembowski and Boucot1990; Zherikhin & Gratshev Reference Zherikhin, Gratshev, Pakaluk and Slipinski1995; etc.), and in the Upper Cretaceous they became dominating in most oryctocenoses. In later deposits, the number of sites with weevils keeps the first place almost always, except deposits from the Miocene. In the Bembridge Marls, Curculionidea are represented mainly by isolated elytra measuring 1.2–6.0mm, and a few entire beetles are entombed in the limestone with conchoidal structure. Most isolated elytra are deposited on the surface of laminae. There could be more than 35 species present, belonging to four families: Anthribidae Billberg, Reference Billberg1820 (Anthribinae, Zygaenodini Lacordaire, Reference Lacordaire1866), Apionidae = Brentidae Billberg, Reference Billberg1820 and Curculionidae Schönherr, Reference Schönherr1823 (e.g., I. 10182; Brodie Collection – Fig. 41e), Curculioninae (Tychiini Thomson, Reference Thomson1859, Cryptorhynchinae Schönherr, Reference Schönherr1826, Molytinae Schönherr, Reference Schönherr1826, Erirhininae Schönherr, Reference Schönherr1826) and Barididae Schönherr Reference Schönherr1826 (Ceutorhynchinae Thomson Reference Thomson1859). The examined assemblage is very probably temperate and consists of forms with different types of phytophagy. Aquatic groups are not clearly represented in the materials examined.
2. General discussion
Beetles are one of most common orders of insects collected from the Bembridge Marls of the Isle of Wight (near to 15%). They are represented by more than 35 families and probably more than 200 species. A total of 44 species were established throughout the course of this study, and 42 of them are described as new. The specimens examined make it possible to propose eight new fossil genera and one subgenus.
There are presently more than 200 specimens of water beetles or beetles whose larvae could develop in fresh water, among approximately 662 specimens identified to family level; i.e., near 20%. This proportion is rather high for the Eocene in general and many sites of Oligocene age. Localities with numerous water beetles are more characteristic of the Upper Oligocene and Miocene. Dominance of swamp Scirtes species is unique among fossil records in general, although a rather great number of marsh beetles (Scirtidae) have also been found in Baltic amber (Upper Eocene) as well as in lowermost Eocene French amber and Fushun amber (Lower Eocene). Aquatic larvae of Recent species of the genus Scirtes live in shallow water in the layer of decaying plant remains or in floating duckweed, liverworts, floating moss (Salvinia natans) and other small floating plants (Azolla in the case of the Bembridge Marls water basin; see Hayes & Collinson Reference Hayes and Collinson2014). Recent populations of Scirtes species can reach a high density, particularly in shallow lakes with reed mace (Typha) and sedge. Larvae of Scirtidae could occur together with species of Spercheus among floating Azolla. Many rove beetles examined in the Bembridge Marl Beds could be shore beetles. An outstanding feature of taphocenoses examined is the high number of Hydraenidae, also characteristic of shallow water today. Among other probable coleopterous indicators of the palaeoenvironment are species of the genus Cyclodinus, whose Recent representatives frequently occur on marine beaches or near inland saline lakes, and species of the Telmatophilus, whose Recent members live on freshwater vegetation above the water level. Leaf beetles of the subfamily Donaciinae, represented mostly by one dominant species, were a rather important component of the coleopterous fauna of Bembridge Marls. It was possible that they fed mostly on Typha. Soft submerged vegetation seemed to be absent or rare because of the lack of Haliplidae, Hydrochinae and some other characteristic Hydrophilidae. It can be supposed that such type of vegetation was absent because of the dominance of Berosus species (but not Enoplurus Hope, 1838) and the presence of Hygrotus (but not representatives of the subgenus Coelambus Thomson, 1860) as well as the presence of Laccophilus and Methlini could point to slightly saline conditions. Finally, because about a quarter of the beetles in the Bembridge Marls were aquatic or near-water inhabitants, phytophagous (including algophagous), mycetophagous and carnivorous feeding strategies were probably taking place.
Thus, the water basin or basins where the deposition of the Bembridge Marls was taking place were rather shallow and with a fairly stable coastline (because of the lack of the waterside ground beetles). The coastal areas and, probably, inner shallow waters were overgrown by Typha (because of plenty of minute moss beetles (Hydraenidae), Scirtes and Telmatophilus), but the areas of exposed water surface with plenty of floating plant leaves (like Potamogeton species and Nymphaceae) seemed to have a smaller area (because of the presence of a comparatively small number of leaf beetles from Donaciinae and the absence of Gyrinidae and water weevils). Surfaces of beds, perhaps, were formed at periods of drying up of algo-bacterial mats. The water was rather hard, although it scarcely had a high percentage of sodium chloride (because of the presence of Anostraca, there was a high diversity of predaceous diving beetles, but the Mesozoic Anostraca lived under conditions exclusionary of salinity). This water basin was most probably stagnant. The finding of ant-like flower beetles from Cyclodinus could be treated as an indirect indicator of salinity. The composition of water beetles is unique for the Palaeogene (because of the high diversity of predaceous diving beetles) and partly for geological history as a whole (because of the many marsh beetles, particularly Scirtes).
Forest beetles were not so numerous as in other Eocene localities such as Green River, Messel and Geiseltal. The most common forest beetles in the Bembridge Marls are jewel beetles (Buprestidae) and sap beetles (Nitidulidae), but, here, at least the jewel beetles are much rarer than in other typical Eocene localities. These families of forest beetles are good fliers and could fly a very long distance (particularly jewel beetles). Certain forest reticulated beetles (Cupedidae) can also be good flyers. The absence, or at least scarcity, of Cerambycidae is rather strange for this age. It is thought that the areas with forest could not be near to the place of deposition. The comparative dominance of the rove beetles over ground beetles is quite unusual and more characteristic of cold or volcanic situations (as in the composition of Biamo (Oligocene) in the Russian Far East). There were no elytron of typical coastal carabids found (like Elaphrus Fabricius, Reference Fabricius1775; Bembidion Latreille, Reference Latreille1802; Blethisa Bonelli, Reference Bonelli1810). The terrestrial beetles are more often entombed as entire beetles in the hard limestone with conchoidal fracture, whereas the water beetles are more often located in laminae with numerous plant remains (Typha). Reconstruction of the environment of deposition as a low-lying herbaceous flood plain seems to be quite probable.
Coleopterous faunas of the Lower–Middle Eocene and Oligocene differ, as deduced from comparing the numbers of families from different localities. The most numerous group in all faunas is the superfamily Curculionoidea. However, the next dominant families are different in the different localities. A generalised dominance scale of families for Eocene sites is as follows: Elateridae–Buprestidae–Carabidae–Chrysomelidae. For the Oligocene, it is Carabidae–Chrysomelidae–Cerambycidae–Staphylinidae. Of course, this is based on the statistics for only some dozens of localities, and the dominance of coleopterous families in each locality could be greatly dependable on the circumstances of taphonomy and peculiarities of the landscape during the time of deposition. The dominance scale in the Isle of Wight Bembridge Marls fauna (Curculionidae–Staphylinidae–Hydrophilidae–Scirtidae–Chrysomelidae) seems to be intermediate between the Eocene and Oligocene types, but it is closer to Oligocene ones (e.g., the dominance scale for the Russian Far East site Biamo, of comparable age, is Elateridae–Staphylinidae–Hydrophilidae–Carabidae–Chrysomelidae).
There are very few taxonomic similarities in Middle Eocene beetles among the oryctocoenoses of the Insect Bed: extinct genera Neothanes from Carabidae, Macropunctum from Elateridae and Necrodexis subgen. nov., and also the genera Byrrhites gen. nov., Davidraenites gen. nov., Eolimnebius gen. nov., Hydraenites gen. nov., Metacoxites gen. nov. and Octotemnites gen. nov. are the only extinct genera. The main resource of knowledge on the Upper Eocene Coleoptera is Baltic amber, but the latter formed in a different environment to the Insect Bed so there are corresponding differences in taphonomy. There is not any beetle among the collections of the Insect Bed that could be treated as conspecific with taxa from the lowermost Oligocene or uppermost Eocene localities, such as those from Florissant in the USA and Biamo in the Russian Far East of comparable age. The most probable stratigraphical position of the Bembridge Marl Beds on the basis of the beetle fauna is uppermost Eocene or lowermost Oligocene, so this corresponds well with the currently believed stratigraphical age of latest Eocene for the Bembridge Marls.
Composition of the Bembridge Marls beetles, as with the Baltic amber fauna, demonstrates some links of similarity with the Recent fauna of South East Asia (Kirejtshuk Reference Kirejtshuk2005b; Kirejtshuk & Kurochkin Reference Kirejtshuk and Kurochkin2010). This feature can be seen partly in the peculiar diversity of Scirtidae in the Bembridge Marls and particularly in the generic attribution of the Bembridge Marls representatives of the families Ptilodactylidae, Cantharidae and Nitidulidae. Also, like Baltic amber Coleoptera, the coleopterous fossils considered here demonstrate some boreal (not subtropical) connection, particularly in the high representation of Donaciinae in remains from the Bembridge Marls and, to a certain degree, the comparable representation of Scirtidae also there. However, the Bembridge Marls representative of Lampyridae shows more resemblance to some groups in the western Hemisphere than those in the eastern one. Earlier faunas do not show such a clear selectivity in the trend of links between fossil and Recent faunas (e.g., if we consider the Palaeocene fauna of Menat or the lowermost Eocene fauna of Oise: Kirejtshuk & Nel Reference Kirejtshuk and Nel2013; Nabozhenko & Kirejtshuk Reference Nabozhenko and Kirejtshuk2013; Kirejtshuk & Nel Reference Kirejtshuk and Nel2018; etc.).
The end of the Eocene was the time of the main climatic shift from the ‘greenhouse' mode of climate of the Middle Eocene (‘thermo-era') to the Recent ‘icehouse' one (‘glaciothermo-era'). Climate of this time was not as equable as in the Eocene, but much more equable than that in the Recent epoch. The equable climate was disappearing, so it is difficult to compare the results in terms of Recent climatology. Faunas at equable times usually consist of a mixture of groups characteristic of Recent tropical and Recent temporate climates (see Archibald & Farrell Reference Archibald and Farrell2003). However, the beetle fauna of the Insect Bed of the Isle of Wight includes mainly temperate components. The authors did not find any specific tropical or clear subtropical elements among the beetles and did not find any similarity of the Insect Bed to a Recent tropical rainforest (unlike McCobb et al. Reference McCobb, Duncan, Jarzembowski, Stankievicz, Wills and Briggs1998). Most older deposits demonstrate more obvious links with the Recent subtropical fauna. Today, the Recent ant species of Oecophylla Smith, 1861 are tropical insects, but Oecophylla remains have been found not only in the Bembridge Marls but also in the temperate Oeningen deposits (Upper Miocene). The Recent species of the genus Mastotermes Froggatt, 1897 are also tropical insects today; however, the Tertiary Mastotermes have been found in many temperate localities as well.
3. Conclusion
The above discussion leads to the following conclusions:
(1) Landscape: low-lying wetland flood plain, mainly open landscape with insular forests or several waterbodies, shallow stagnant water plentifully overgrown by reed mace (Typha), with floating and submergent plants with hard water, sedimentation – mainly palustrine limestone (Platt & Weight Reference Platt and Weight1992).
(2) Climate: more or less equable, probably without frosts, warm temperate.
(3) Most probable stratigraphical position: uppermost Eocene or lowermost Oligocene.
4. Acknowledgements
This paper contributes to the INTAS project 03-51-4367, which was initiated by A. J. Ross. In general, the research of beetles from the Bembridge Marls would have been scarcely possible without encouragement and friendly assistance from A. Ross (formerly NHMUK, now at the National Museum of Scotland in Edinburgh), and the opportunity to use laboratory equipment and other facilities in London. Some pictures were prepared in the specialised laboratory of the NHMUK. The authors also greatly appreciate M. V. L. Barclay, R. Booth and other colleagues in the Entomology Department of the NHM for their assistance to this study in various ways. B. A. Korotyaev, A. V. Kovalev and N. N. Yunakov (Zoological Institute of the Russian Academy of Sciences), D. Telnov (NHM), I. Löbl (Muséum d'Histoire Naturelle, Geneva), F. Vitali (Luxembourg), A. A. Prokin (Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences, Borok, Russia), R. Constantin (Saint-Lô, France), S. E. Tshernyshev (Institute of Systematics and Ecology of Animals, Siberian Branch, Russian Academy of Sciences, Frunze Street, 11, Novosibirsk) and A. V. Kompantzev (Institute of Evolution and Ecology of Animals of the Russian Academy of Sciences, Moscow) gave valuable advice on the identification and description of coleopterous remains. The authors wish to express their thanks to Phil Crabb (formerly NHMUK) for taking some of the photos used for preparation of the figures. Thanks also go to Bill Crighton (NMS volunteer) for checking the paper's English. The studies of the first author were carried out within the framework of the Russian state research project No. AAAA-A17-117030310210-3. This study was also partly supported by the Programme of the Presidium of the Russian Academy of Sciences, ‘Origin and Evolution of Biosphere' and ‘Evolution of the organic world. Significance and influence of planetary processes', and also the Russian Foundation of Basic Research (grant numbers 12-04-00663-a, 18-04-00243-a and 19-04-00465-a). A. Yu. Solodovnikov's stay at the NHMUK was partially supported by US NSF PEET (grant number 00118749).











































