



Introduction
The Carboniferous is formally subdivided into two subseries by the International Commission on Stratigraphy as the Mississippian, comprising the Tournaisian, Visean, and Serpukhovian stages, and the Pennsylvanian with the Bashkirian, Moscovian, Kasimovian, and Gzhelian stages (Richards, Reference Richards, Lucas, DiMichele, Barrick, Schneider and Spielmann2013; Ogg et al., Reference Ogg, Ogg and Gradstein2016). The Mississippian-Pennsylvanian boundary corresponds to the lower-middle Carboniferous boundary in the Russian regional chronostratigraphic subdivisions, and according to these subdivisions, the Bashkirian and Moscovian stages are subdivided into regional substages or horizons (see Kagarmanov and Donakova, Reference Kagarmanov, Donakova and Donakova1990; Kulagina et al., Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001). The stages/substages or horizons and their boundaries are mainly determined in current studies by using the evolutionary lineages of the certain conodont taxa. The fusulinids have also been extensively evaluated as indices for the Mississippian-Pennsylvanian boundary; however, due to problems of provincialism, conodonts appear to be the best indicators. The fusulinid taxa of the Mississippian-Pennsylvanian boundary in the Eurasian-Arctic Faunal Realm (Ross, Reference Ross1967) differ from the Midcontinent-Andean Faunal Realm (Ross, Reference Ross1967) due to the closure of the seaway between Gondwana and Laurussia, which resulted in diachronous development of different phylogenetic lineages (Groves, Reference Groves1988; Groves et al., Reference Groves, Nemyrovska and Alekseev1999). The Mississippian-Pennsylvanian (Serpukhovian-Bashkirian) boundary has a GSSP at Arrow Canyon, Nevada, USA (Lane et al., Reference Lane, Brenckle, Baesemann and Richards1999) and is recognized by the FAD of conodont Declinognathodus noduliferus (Ellison and Graves, Reference Ellison and Graves1941) (sensu lato) in the Gnathodus girtyi simplex Dunn, Reference Dunn1966–Declinognathodus noduliferus sensu lato lineage.
The Hadim Nappe successions preserve suitable strata to determine the stage/substages or horizon boundaries by fusulinid assemblages, with an almost continuous succession, including stage/substages or horizons within shallow marine carbonates with some siliciclastic intercalations from the Middle (?)–Late Devonian to Late Cretaceous in the Central Taurides (Özgül, Reference Özgül1976, Reference Özgül1997; Monod, Reference Monod1977; Turan, Reference Turan1990). The shallow-marine and carbonate-rich Hadim Nappe successions and their fusulinid assemblages are very similar to those from other regions of the Tethyan Realm (Kalvoda, Reference Kalvoda1990, Reference Kalvoda2003), particularly the Russian Platform and the southern Urals. Some studies have established stage boundaries based on fusulinids (Okuyucu, Reference Okuyucu1997, Reference Okuyucu2002, Reference Okuyucu2008, Reference Okuyucu2009; Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007; Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011; Demirel and Altıner, Reference Demirel and Altıner2016) and conodonts (Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner and Özkan–Altıner2012; Atakul-Özdemir, Reference Atakul-Özdemir2015) corresponding to the Carboniferous units of the Hadim Nappe.
The aims of this study are: (1) to describe and to give the stratigraphic distribution and biostratigraphic characteristics of the fusulinid assemblages of the upper Serpukhovian–Bashkirian successions of the Hadim Nappe, (2) to correlate them with the coeval assemblages in Turkey and other regions, and (3) to delineate the Mississippian-Pennsylvanian boundary based on the fusulinid fauna. For this purpose, two fusulinid-rich carbonate sections (Yassıpınar and Gölbelen) of the Hadim Nappe were selected for detailed biostratigraphic studies from the Central Taurides, where the Hadim Nappe widely crops out with continuous successions. Two of these are the Yassıpınar (Hadim area) and Gölbelen (Bozkır region) sections in the Central Taurides (Fig. 1).
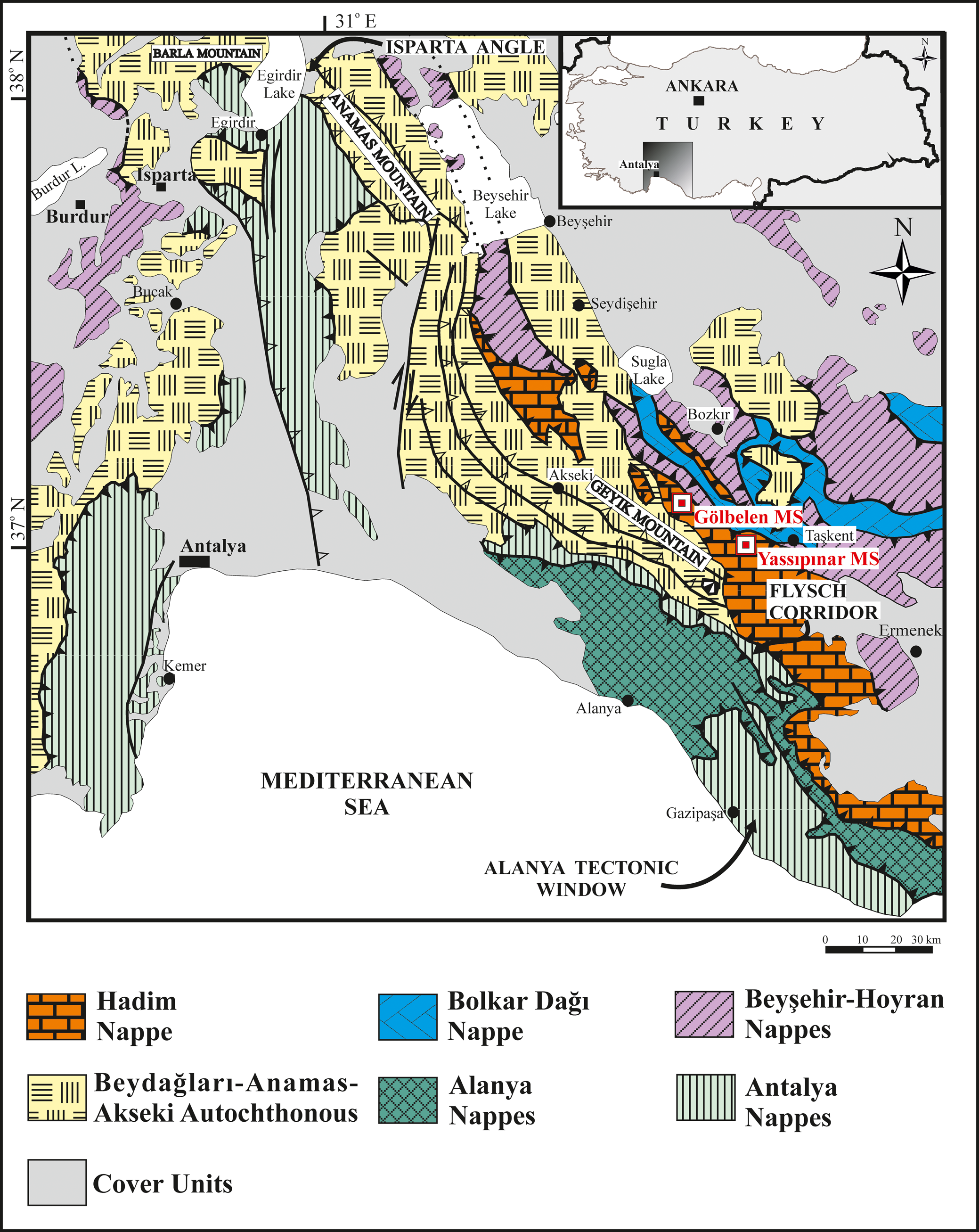
Figure 1. Schematic map showing the distribution of autochthonous and allochthonous sequences in the area between the Western and Central Taurides with the location of the Yassıpınar and Gölbelen measured sections (MS) (simplified and revised after Özgül, Reference Özgül, Tekeli and Göncüoğlu1984).
Geological setting
The Taurides, one of the major units of the Alpine-Himalayan Orogenic Belt, is geographically subdivided into three segments or parts by Özgül (Reference Özgül1976, Reference Özgül, Tekeli and Göncüoğlu1984) as: (1) the Eastern Taurides to the east of the Ecemis fault, (2) the Central Taurides between the Kırkkavak fault and the Ecemis fault, and (3) the Western Taurides from the Aegean coast to the Kırkkavak fault. The Tauride Belt is made up of a number of autochthonous and allochthonous units with distinct stratigraphic, structural, and metamorphic features (e.g., Blumenthal, Reference Blumenthal1944, Reference Blumenthal1951; Brunn et al., Reference Brunn, Dumont, De Graciansky, Gutnic, Juteau, Marcoux, Monod, Poisson and Campbell1971, Reference Brunn, Argyriadis, Marcoux, Monod, Poisson and Ricou1973; Özgül, Reference Özgül1976, Reference Özgül, Tekeli and Göncüoğlu1984, Reference Özgül1997) that have been regarded as nappes (Brunn et al., Reference Brunn, Dumont, De Graciansky, Gutnic, Juteau, Marcoux, Monod, Poisson and Campbell1971) or tectonostratigraphic units (Özgül, Reference Özgül1976, Reference Özgül, Tekeli and Göncüoğlu1984) (Fig. 1). Of these, the Beydağları-Anamas-Akseki Unit is autochthonous, whereas the allochthonous units are characterized by the Hadim (Aladağ Unit of Özgül, Reference Özgül1976), Bolkar Dağı, Beyşehir-Hoyran, Antalya, and Alanya nappes (Blumenthal, Reference Blumenthal1944, Reference Blumenthal1951; Gutnic et al., Reference Gutnic, Kelter and Monod1968, Reference Gutnic, Monod, Poisson and Dumont1979; Brunn et al., Reference Brunn, Graciansky, Gutnic, Juteau, Lefevre, Marcoux, Monod and Poisson1970, Reference Brunn, Dumont, De Graciansky, Gutnic, Juteau, Marcoux, Monod, Poisson and Campbell1971; Monod, Reference Monod1977; Turan, Reference Turan1990, Reference Turan2000, Reference Turan2010).
The Hadim Nappe, originally established by Blumenthal (Reference Blumenthal1941, Reference Blumenthal1951) in the Central Taurides, is made up of sedimentary rocks, including shallow-marine platform carbonates with some siliciclastic intercalations that range in age from the Middle (?)–Late Devonian to the Late Cretaceous (Güvenç, Reference Güvenç1965, Reference Güvenç1977a; Özgül, Reference Özgül1971, Reference Özgül1997; Monod, Reference Monod1977; Turan, Reference Turan1990, Reference Turan2000, Reference Turan2010) (Fig. 2). The Hadim Nappe corresponds to the “Siyah Aladag Nappe” (Blumenthal, Reference Blumenthal1941) and Belemedik Paleozoic Window (Blumenthal, Reference Blumenthal1947) of the Eastern Taurides, the “Aladağ Unit” (Özgül, Reference Özgül1976) and the “Bademli-Camlik Unit” (Monod, Reference Monod1977) of the Central Taurides, and the Karadag Serie (Graciansky, Reference Graciansky1968) of the Western Taurides.

Figure 2. Generalized columnar section of the Hadim Nappe (modified after Monod, Reference Monod1977; Turan, Reference Turan1990; Özgül, Reference Özgül1997). Key: U. C., Upper Cretaceous.
In previous studies, numerous formation names have been used for the Carboniferous deposits of the Hadim Nappe (e.g., Monod, Reference Monod1977; Güvenç, Reference Güvenç1980; Tekeli et al., Reference Tekeli, Aksay, Urgun, Işık, Tekeli and Göncüoğlu1984; Lengeranlı et al., Reference Lengeranlı, Celtek, Batik, Ayhan, Tuzun, Inan and Inam1986; Okuyucu and Güvenç, Reference Okuyucu and Güvenç1997; Özgül, Reference Özgül1997). In this study, the lithostratigraphic scheme of Güvenç (Reference Güvenç1977b, Reference Güvenç1980) is adopted, and the name Dikenli is used for the Carboniferous deposits. The Carboniferous Dikenli Formation consists of black shale, siltstone, and carbonates at the base, fusulinid-rich carbonates with sandstone intercalations in the central part, and carbonates and Girvanella Limestone facies (sensu Güvenç, Reference Güvenç1965) at the top (Fig. 2).
The upper Serpukhovian–Bashkirian successions of the Hadim Nappe from the Central Taurides were studied in two measured stratigraphic sections, namely the Yassıpınar section (UTM: 36S 444515.2mE, 4084999.6mN) to the south of Hadim town (Fig. 3) and the Gölbelen section (UTM: 36S 419446.7mE, 4100126.3mN) to the south of the Bozkır town (Fig. 4). The Mississippian-Pennsylvanian boundary interval was detected only at the Yassıpınar section (Fig. 5). In contrast, the base of the Gölbelen section begins with a base of the Bashkirian Stage (Syuranian Substage) (Fig. 6). The lower part of the Bashkirian Stage in both sections is made up of mainly oolitic limestone with rare quartz arenite sandstone interbeds, indicating a shallow-marine environment (Figs. 5, 6). The oolitic limestone beds are mainly gray- or yellow-colored, thin- to medium-bedded, and represented by the oolitic grainstone microfacies with a poor fauna and flora. Although the oolitic limestone and bioclastic limestone intercalations show continuation to the overlying strata of the Bashkirian Stage (Akavasian–Askynbashian substages) at the Gölbelen section, this interval is represented by yellow-colored and thick-bedded quartz arenite sandstone in the Yassıpınar section, except for 1 m thick limestone strata in the middle part. The upper part of the Bashkirian strata at both sections consists of mainly fusulinid-rich bioclastic limestone without any terrigenous input (Figs. 5, 6). The fusulinid-rich bioclastic limestone beds are gray-colored and medium- to thick-bedded, and characterized dominantly by bioclastic grainstone and locally by bioclastic grainstone-packstone microfacies with an abundant fusulinid fauna, with algae, echinoderms, bryozoans, and rare intraclasts.

Figure 3. Geological map of the study area showing the location of the Yassıpınar measured section (simplified and revised after Turan, Reference Turan1990).

Figure 4. Geological map of the study area showing the location of the Gölbelen measured section (simplified and revised after Metin, Reference Metin1994).

Figure 5. Lithostratigraphy and fusulinid distribution in the Yassıpınar section.

Figure 6. Lithostratigraphy and fusulinid distribution in the Gölbelen section.
Lithostratigraphic properties of the Serpukhovian and Bashkirian stages
The Serpukhovian strata of the Hadim Nappe are mainly composed of sandstone and rare limestone, with a poor fauna and flora. The lowermost Bashkirian is predominantly represented by oolitic limestone beds indicating a shallow-water depositional environment (Figs. 5, 6). The fusulinids in this interval were only recovered from the central parts of the ooids. The Akavasian and Askynbashian interval of the Yassıpınar section consists dominantly of sandstone without fusulinids. In contrast, at the Gölbelen section, this interval is represented by bioclastic limestone with rich and diversified fusulinid assemblages, and most characteristic fusulinids of the Akavasian-Askynbashian interval have been defined from this section. Although some terrigenous deposits and a poor fusulinid fauna occur in the boundary interval of the two sections, almost all fusulinid assemblages of this interval were recovered precisely based on the studied two sections (Fig. 7).

Figure 7. Total stratigraphic distribution of fusulinids from the upper Serpukhovian to Bashkirian in this study.
Biostratigraphy
Biostratigraphy of the Serpukhovian-Bashkirian boundary
Although the Serpukhovian-Bashkirian boundary is recognized by the FAD of conodont Declinognathodus noduliferus at its GSSP at the Arrow Canyon, Nevada, USA (Lane et al., Reference Lane, Brenckle, Baesemann and Richards1999), it is also known that the fusulinid foraminifera allow identification of the boundary. According to previous studies, there are many fusulinid foraminifera to distinguish the boundary between the Serpukhovian and the Bashkirian stages in various regions around the world (Brenckle et al., Reference Brenckle, Lane, Manger and Saunders1977, Reference Brenckle, Groves, Skipp, Ramsbottom, Saunders, Owens, Ramsbottom, Saunders and Owens1982, Reference Brenckle, Baesemann, Lane, West, Webster, Langenheim, Brand and Richards1997; Skipp et al., Reference Skipp, Baesemann and Brenckle1985; Vachard and Maslo, Reference Vachard and Maslo1996; Davydov, Reference Davydov1997; Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003; Richards et al., Reference Richards, Lane, Brenckle, Hills, Hills, Henderson and Bamber2002; Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011; Leven, Reference Leven2012; Kulagina et al., Reference Kulagina, Nikolaeva, Pazukhin and Kochetova2014). Despite many different index foraminiferal taxa and regions, due to the good correlation we prefer to focus on the Tethyan Realm, especially the Russian Platform and the southern Urals. The base of the Bashkirian along the Mississippian-Pennsylvanian transition is determined by the first occurrence of Plectostaffella jakhensis (Reitlinger, Reference Reitlinger and Rauzer-Chernousova1971), which is the one of the index foraminifera in the Russian Platform, southern Urals, Hadim Nappe (Central Taurides, Turkey), and Elburz (Iran) (Einor, Reference Einor, Wagner, Winkler Prins and Granados1996; Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001, Reference Kulagina, Nikolaeva, Pazukhin and Kochetova2014; Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011; Leven and Gorgij, Reference Leven and Gorgij2011). The Serpukhovian-Bashkirian boundary was determined between the Eostaffella ex gr. E. ikensis-Eostaffella postmosquensis Zone and the Plectostaffella jakhensis-Plectostaffella bogdanovkensis Zone in the Central Taurides based on foraminifers by Atakul-Özdemir et al. (Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011). Later, this boundary was studied by Atakul-Özdemir et al. (Reference Atakul-Özdemir, Altıner and Özkan–Altıner2012) based on conodonts and the conodont Declinognathodus noduliferus bernesgae Sanz-López et al., Reference Sanz-López, Blanco-Ferrera, García-López and Sánchez De Posada2006 is found just below the boundary, with the base of the Bashkirian stage determined by the occurrence of the conodont Declinognathodus inaequalis (Higgins, Reference Higgins1975).
The fusulinid assemblages of the upper Serpukhovian and Bashkirian strata permit a fine subdivision and comparison with coeval fusulinid assemblages of other Tethyan regions, such as the Taurides (Turkey), southern Urals, Russian Platform, and Donetz Basin. The biostratigraphic characteristics of the upper Serpukhovian and the Bashkirian stages are as follows:
Serpukhovian Stage, upper substage
The Serpukhovian interval is represented mainly by siliciclastic rocks and rarely carbonate strata in the Hadim Nappe. In this study, the uppermost part of the Serpukhovian stage has been defined in the Yassıpınar stratigraphic section, and it is characterized by thick sandstone layers with a thin limestone bed that contains Eostaffella postmosquensis acutiformis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951 (Figs. 5, 7, 8).
Bashkirian Stage, Syuranian Substage
Due to extensive tectonic deformation, the base of the Gölbelen section is bounded by a fault and the lower Bashkirian (lowermost part of the Syuranian Substage) succession could not be determined (Fig. 6). However, the basal part of the section includes some characteristic fusulinid faunas of the Syuranian Substage (lower Bashkirian) (Figs. 6, 7). In the Yassıpınar section, which has a continuous succession from Serpukhovian to Bashkirian, the base of the Bashkirian Stage is determined by the FAD of Plectostaffella jakhensis, and occurrence of the Plectostaffella bogdanovkensis Reitlinger, Reference Reitlinger and Menner1980 and Plectostaffella varvariensis (Brazhnikova and Potievskaya, Reference Brazhnikova and Potievskaya1948) from the above beds (Figs. 5, 7).
The Syuranian Substage of the Bashkirian Stage includes abundant and diverse fusulinid assemblages in the Hadim Nappe sections. These fusulinid assemblages are composed of Plectostaffella bogdanovkensis, P. jakhensis, P. varvariensis, Semistaffella minuscilaria (Reitlinger, Reference Reitlinger and Rauzer-Chernousova1971), and Semistaffella variabilis (Reitlinger, Reference Reitlinger1961), which are important index taxa for the Syuranian Substage (Figs. 8, 9).

Figure 8. Thin-section photomicrographs of the fusulinid assemblages of the Yassıpınar (YS) and Gölbelen (GS) sections. (1) Eostaffella acuta Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950, axial section, 15Y16A.08.02, YS. (2) Eostaffella designata (Zeller, Reference Zeller1953), subaxial section, 15Y07.04.02, YS. (3) Eostaffella kashirica Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, axial section, 18G25.12, GS. (4, 5) Eostaffella mirifica Brazhnikova in Brazhnikova et al., Reference Brazhnikova, Vakarchuk, Vdovenko, Vinnichenko, Karpova, Kolomiets, Potievskaya, Rostovtseva and Shevchenko1967, (4) axial section, 15Y15.05, YS; (5) axial section, 18G18.11.02, GS. (6, 7) Eostaffella ovoidea (Rauzer-Chernousova, Reference Rauzer-Chernousova1948), (6) axial section, 15Y12.08, YS; (7) axial section, 18G14.03.01, GS. (8, 9) Eostaffella postmosquensis postmosquensis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, (8) axial section, 15Y05.03.02, YS; (9) subaxial section, 18G21.04.03, GS. (10, 11) Eostaffella postmosquensis acutiformis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, (10) axial section, 15Y05.05.01, YS; (11) axial section, 15Y04.01.01, YS. (12) Eostaffella cf. E. ovoidea (Rauzer-Chernousova, Reference Rauzer-Chernousova1948), subaxial section, 18G16.03, GS. (13) Eostaffella ex gr. E. pseudostruvei pseudostruvei (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936), axial section, 15Y10.05.01, YS. (14, 15) Eostaffella pseudostruvei angusta Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, (14) axial section, 15Y05.06.01, YS; (15) axial section, 18G21.04.01, GS. (16, 17) Plectostaffella bogdanovkensis Reitlinger, Reference Reitlinger and Menner1980, subaxial sections, (16) 15Y07.04.01, YS; (17) 15Y08.02, YS. (18) Plectostaffella cf. P. evolutica (Rumyantseva, Reference Rumyantseva and Shayakubov1970), axial section, 15Y05.04.01, YS. (19, 20) Plectostaffella jakhensis (Reitlinger, Reference Reitlinger and Rauzer-Chernousova1971), (19) axial section, 15Y05.03.01, YS; (20) subaxial section, 15Y05.06.02, YS. (21, 22) Plectostaffella varvariensis (Brazhnikova and Potievskaya, Reference Brazhnikova and Potievskaya1948), axial sections, (21) 15Y08.03, YS; (22) 15Y08.06.02, YS. (23, 24) Ozawainella pararhomboidalis Manukalova, Reference Manukalova1950, axial sections, (23) 15Y16B.07, YS; (24) 18G20.04, GS. (25, 26) Semistaffella minuscilaria Reitlinger, Reference Reitlinger and Rauzer-Chernousova1971, axial sections, (25) 18G13.03, GS; (26) 18G15.05, GS. Scale bars = 250 μm (1–22, 25, 26); = 500 μm (23, 24).

Figure 9. Thin-section photomicrographs of the fusulinid assemblages of the Yassıpınar (YS) and Gölbelen (GS) sections. (1, 2) Semistaffella primitiva (Reitlinger, Reference Reitlinger1961), (1) subaxial section, 15Y13.03, YS; (2) axial section, 18G13.04.01, GS. (3, 4) Semistaffella variabilis (Reitlinger, Reference Reitlinger1961), (3) subaxial section, 15Y13.08.01; YS; (4) axial section, 18G13.04.02, GS. (5) Varistaffella eostaffellaeformis (Rumyantseva, Reference Rumyantseva and Shayakubov1970), subaxial section, 18G15.03, GS. (6, 7) Varistaffella ziganica (Sinitsyna in Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975), (6) subaxial section, 15Y10.04.01, YS; (7) axial section, 15Y16A.10, YS. (8, 9) Pseudostaffella antiqua (Dutkevich, Reference Dutkevich1934), (8) axial section, 18G18.08, GS; (9) subaxial section, 18G19.06, GS. (10, 11) Pseudostaffella composita Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950, axial sections, (10) 15Y15.04, GS; (11) 18G18.06, GS. (12) Pseudostaffella compressa (Rauzer-Chernousova, Reference Rauzer-Chernousova1938), axial section, 18G17.04, GS. (13) Pseudostaffella ex gr. P. gorskyii Dutkevich in Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950, axial section, 18G23.06, GS. (14, 15) Pseudostaffella grandis (Shlykova in Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950), axial sections, (14) 18G18.07.01, GS; (15) 18G19.03, GS. (16) Pseudostaffella aff. P. paracompressa Safonova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, axial section, 15Y16A.08.01, GS. (17) Pseudostaffella cf. P. proozawai Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, axial section, 15Y15.03, YS. (18, 19) Pseudostaffella sofronizkyi Safonova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, (18) subaxial section, 18G18.09, GS; (19) axial section, 15Y14.06, YS. (20–23) Pseudostaffella uralica Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, axial sections, (20) 15Y12.03, YS; (21) 15Y12.07, YS; (22) 18G14.03.02, GS; (23) 18G21.03, GS. (24–26) Schubertella ex gr. Sch. multiforme Villa in Villa and Merino-Tomé, Reference Villa and Merino-Tomé2016, axial sections, (24) 15Y16A.07, YS; (25) 15Y16A.27, YS; (26) 15Y16B.05, YS. (27, 28) Schubertella obscura obscura Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930, axial sections, (27) 15Y20.04, YS; (28) 15Y22.03.02, YS. (29, 30) Schubertella obscura mosquensis Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, (29) subaxial section, 15Y14.04, YS; (30) axial section, 15Y16B.08, YS. Scale bars = 250 μm (1–7, 19–30); = 500 μm (8–18).
Plectostaffella jakhensis was originally described by Reitlinger (Reference Reitlinger and Rauzer-Chernousova1971) from the South Urals (Jakhino village, Berdyaush River), and has been defined in coeval strata by many authors (Groves, Reference Groves1988; Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001; Leven et al., Reference Leven, Davydov and Gorgij2006; Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011; Leven and Gorgij, Reference Leven and Gorgij2011; Leven, Reference Leven2012). Plectostaffella bogdanovkensis, which is one of the most important taxa of this interval, was first described from the Bogdanovkian horizon of the Bashkirian Stage by Reitlinger (Reference Reitlinger and Menner1980), and then determined in coeval strata (Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001, Reference Kulagina, Nikolaeva, Pazukhin and Kochetova2014; Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011; Leven and Gorgij, Reference Leven and Gorgij2011) or from the upper substages of the Bashkirian Stage (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007; Leven, Reference Leven2012). The other characteristic taxon of this interval, Plectostaffella varvariensis, was determined at the base of the lower Bashkirian succession in Donbas (Ukraine) by Brazhnikova and Potievskaya (Reference Brazhnikova and Potievskaya1948). Plectostaffella varvariensis, widely distributed in the Tethyan Realm, is known in the southern Urals (Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001), Elburz Mountains (Iran) and central Iran (Leven et al., Reference Leven, Davydov and Gorgij2006; Leven and Gorgij, Reference Leven and Gorgij2011), Pamir Mountains (Tajikistan) (Leven, Reference Leven2012), and Taurides (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007; Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011) during the Syuranian–Askynbashian interval. The diverse fusulinid assemblages of the Syuranian Substage also comprise Eostaffella postmosquensis postmosquensis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; E. postmosquensis acutiformis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; E. designata (Zeller, Reference Zeller1953); E. ovoidea (Rauzer-Chernousova, Reference Rauzer-Chernousova1948); E. ex gr. E. pseudostruvei pseudostruvei (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936); E. pseudostruvei angusta Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Plectostaffella cf. P. evolutica (Rumyantseva, Reference Rumyantseva and Shayakubov1970); Semistaffella primitiva (Reitlinger, Reference Reitlinger1961); Varistaffella eostaffellaeformis (Rumyantseva, Reference Rumyantseva and Shayakubov1970); V. ziganica (Sinitsyna in Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975); and Pseudostaffella uralica Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951 (Fig. 7). The characteristic early Bashkirian fusulinids, including Eostaffella ovoidea, Semistaffella minuscilaria, S. variabilis, and Pseudostaffella uralica are also described within the Akavasian Substage of the Bashkirian in the Yassıpınar section (Figs. 5, 7).
Bashkirian Stage, Akavasian Substage
Besides the abundant and diverse fusulinid assemblages, advanced species of the genus Pseudostaffella Thompson, Reference Thompson1942 first appear in the Akavasian Substage of the Bashkirian Stage in the studied sections. The strata of the Akavasian Substage contain quite abundant fusulinid assemblages with Eostaffella postmosquensis postmosquensis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; E. ex gr. E. pseudostruvei pseudostruvei (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936); E. mirifica Brazhnikova in Brazhnikova et al., Reference Brazhnikova, Vakarchuk, Vdovenko, Vinnichenko, Karpova, Kolomiets, Potievskaya, Rostovtseva and Shevchenko1967; E. ovoidea (Rauzer-Chernousova, Reference Rauzer-Chernousova1948); Semistaffella primitiva (Reitlinger, Reference Reitlinger1961); S. variabilis (Reitlinger, Reference Reitlinger1961); Pseudostaffella antiqua (Dutkevich, Reference Dutkevich1934); P. composita Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950; P. compressa (Rauzer-Chernousova, Reference Rauzer-Chernousova1938); P. grandis (Shlykova in Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950); P. cf. P. proozawai Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; P. sofronizkyi Safonova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; P. uralica Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; and Schubertella obscura mosquensis Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951 (Fig. 7). The species Pseudostaffella antiqua, which is the index taxon of the Akavasian Substage, is only found in the Gölbelen section (Figs. 6, 9). This characteristic species was originally described by Dutkevich (Reference Dutkevich1934) in the lower Moscovian successions of the Urals. Later, it was also determined throughout the Bashkirian–lower Moscovian successions in the Russian Platform, Kolva-Vishera (Russia), Cantabrian Mountains (Spain), and the Nagaiwa Formation (Japan) (Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1954; Ginkel, Reference Ginkel1965; Kobayashi, Reference Kobayashi1973). Pseudostaffella antiqua is widely distributed and is known in the southern Urals (Russia) (Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001; Kulagina, Reference Kulagina2003), Elburz Mountains (Iran) and central Iran (Leven et al., Reference Leven, Davydov and Gorgij2006; Leven and Gorgij, Reference Leven and Gorgij2011), Pamir Mountains (Tajikistan) (Leven, Reference Leven2012), and Hadim Nappe. The other common taxa of the Akavasian Substage comprise Eostaffella mirifica, Pseudostaffella composita, and Pseudostaffella sofronizkyi, which are present in the Yassıpınar and Gölbelen sections of the Hadim Nappe (Figs. 5, 6).
Bashkirian Stage, Askynbashian Substage
The fusulinid assemblages of this substage are more abundant and more diverse than in the previous one, and the advanced fusulinid genus Depratina Solovieva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996 first appears (Fig. 10) with other fusulinid genera (Ozawainella Thompson, Reference Thompson1935, Staffellaeformes Solovieva, Reference Solovieva1986, and Schubertella Staff and Wedekind, Reference Staff and Wedekind1910) in the Yassıpınar and Gölbelen sections (Figs. 5, 6). The species Staffellaeformes staffellaeformis (Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951), index taxon of the base of the Askynbashian substage of the Bashkirian, is identified only in the Gölbelen section (Fig. 6). The Yassıpınar section also includes different species belonging to the genus Staffellaeformes (Fig. 5). Staffellaeformes staffellaeformis was first described from the Russian Platform by Kireeva (in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) in lower Moscovian units, later described from the Alai (Turkmenistan) (Dzhenchuraeva, Reference Dzhenchuraeva1979) and the Cantabrian Mountains (Villa, Reference Villa1995) in the upper Bashkirian–lower Moscovian successions. In accordance with this study, Staffellaeformes staffellaeformis was determined in coeval strata of the Bashkirian successions of the southern Urals (Russia) (Kulagina et al., Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001; Kulagina, Reference Kulagina2003, Reference Kulagina2008, Reference Kulagina2009), Elburz Mountains (Iran) and central Iran (Leven and Gorgij, Reference Leven and Gorgij2011), Pamir Mountains (Tajikistan) (Leven, Reference Leven2012), and Hadim Nappe (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007). The entire fusulinid assemblage of this interval contains Eostaffella acuta Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950; E. mirifica Brazhnikova in Brazhnikova et al., Reference Brazhnikova, Vakarchuk, Vdovenko, Vinnichenko, Karpova, Kolomiets, Potievskaya, Rostovtseva and Shevchenko1967; E. postmosquensis postmosquensis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; E. pseudostruvei angusta Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Ozawainella pararhomboidalis Manukalova, Reference Manukalova1950; Varistaffella ziganica (Sinitsyna in Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975); Pseudostaffella antiqua (Dutkevich, Reference Dutkevich1934); P. grandis (Shlykova in Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950); P. aff. P. paracompressa Safonova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; P. uralica Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Schubertella ex gr. Sch. multiforme Villa in Villa and Merino-Tomé, Reference Villa and Merino-Tomé2016; Sch. obscura mosquensis Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Depratina convoluta (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930) n. comb.; D. prisca prisca (Deprat, Reference Deprat1912); D. prisca sphaeroidea (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951); Staffellaeformes bona (Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1954); S. parva parva (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930); S. parva robusta (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936) n. comb.; and S. staffellaeformis (Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) (Fig. 7). Although the Yassıpınar section is characterized by more diverse and abundant fusulinid assemblages, the Gölbelen stratigraphic section includes index taxa of the Askynbashian Substage. Furthermore, despite the fact that two sections contain the same generic taxa, only two species, Ozawainella pararhomboidalis and Staffellaeformes parva parva, are mutual species.

Figure 10. Thin-section photomicrographs of the fusulinid assemblages of the Yassıpınar (YS) and Gölbelen (GS) sections. (1–3) Profusulinella pararhomboides Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936, subaxial sections, (1) 15Y20.03, YS; (2) 15Y26.03, YS; (3) axial section, 18G27.12, GS. (4–6) Profusulinella rhomboides (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930), (4) subaxial section 15Y19.06, YS; axial sections, (5) 15Y26.04, YS; (6) 18G31.07, GS. (7) Solovievaia ex gr. S. nuratavensis (Solovieva, Reference Solovieva1977), axial section, 18G26.07, GS. (8, 9) Depratina beppensis (Toriyama, Reference Toriyama1958), axial sections, (8) 15Y18.12, YS; (9) 15Y18.21, YS. (10–13) Depratina convoluta (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930) n. comb., axial sections, (10) 15Y17.04, YS; (11) 15Y18.03, YS; (12) 15Y23.09, YS; (13) 18G26.09, GS. (14, 15) Depratina prisca prisca (Deprat, Reference Deprat1912), axial sections, (14) 15Y16A.19.01, YS; (15) 15Y18.13, YS. (16–18) Depratina prisca sphaeroidea (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951), axial sections, (16) 15Y16A.12, YS; (17) 15Y16A.17, YS; (18) 18G24.03, GS. (19, 20) Depratina prisca timanica (Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951), axial sections, (19) 15Y18.11, YS; (20) 15Y21.13, YS. (21–23) Depratina turani Akbaş n. sp., (21) axial section, 15Y17.03, YS; (22) subaxial section, 15Y17.06, YS; (23) axial section, 15Y17.08 (holotype), YS. Scale bars = 500 μm (11–13); = 1 mm (1–10, 14–23).
Bashkirian Stage, Arkhangelskian Substage
The Arkhangelskian has an enormous diversity of fusulinid assemblages in the Hadim Nappe sections. In the studied sections, in addition to the previously noted genera, advanced fusulinids such as Solovievaia Vachard and Le Coze, Reference Vachard and Le Coze2018; Aljutovella Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Tikhonovichiella Solovieva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996; and Verella Dalmatskaya, Reference Dalmatskaya1951 first appear in the Arkhangelskian Substage. The species of the latter two genera, Tikhonovichiella tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) and Verella spicata Dalmatskaya, Reference Dalmatskaya1951, are the index taxa of the Arkhangelskian Substage. Although the species Tikhonovichiella tikhonovichi is present in both the Yassıpınar and Gölbelen sections, the species Verella spicata is only found in the Gölbelen section (Figs. 5, 6). One of the characteristic taxon of the Bashkirian-Moscovian boundary, Tikhonovichiella tikhonovichi, was described by Rauzer-Chernousova (in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) from the lower Moscovian succession of the Russian Platform. Subsequently, it has been reported in the Urals (Russia) (Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975; Kulagina et al., Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001; Kulagina, Reference Kulagina2003, Reference Kulagina2008), Pamir Mountains (Tajikistan) (Leven, Reference Leven2012), and in the Hadim Nappe (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007) successions. The other characteristic taxon of this interval, Verella spicata, was originally described by Dalmatskaya (Reference Dalmatskaya1951) at the base of the Moscovian Stage of the Russian Platform, then was reported in the upper Bashkirian successions of the South Gissar (Bensh, Reference Bensh1969; Saltovskaya, Reference Saltovskaya1974), Alai (Turkmenistan) (Dzhenchuraeva, Reference Dzhenchuraeva1979), Cantabrian Mountains (Villa, Reference Villa1995), Pamir Mountains (Tajikistan) (Leven, Reference Leven2012), and Hadim Nappe (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007). Except these two index species, the Arkhangelskian Substage in the Hadim Nappe sections contains following diverse fusulinid species; Eostaffella kashirica Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Ozawainella pararhomboidalis Manukalova, Reference Manukalova1950; Pseudostaffella ex gr. P. gorskyii Dutkevich in Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1950; Schubertella obscura obscura Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930; Profusulinella pararhomboides Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936; P. rhomboides (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930); Solovievaia ex gr. S. nuratavensis (Solovieva, Reference Solovieva1977); Depratina beppensis (Toriyama, Reference Toriyama1958); D. convoluta (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930); D. prisca prisca (Deprat, Reference Deprat1912); D. prisca sphaeroidea (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951); D. prisca timanica (Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951); D. turani Akbaş n. sp.; Staffellaeformes bona (Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1954); S. parva robusta (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936) n. comb.; S. staffellaeformis (Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951); S. tashliensis (Lebedeva in Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975); Aljutovella cf. A. pseudoaljutovica Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951; Tikhonovichiella praetikhonovichi Akbaş n. sp.; T. tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951); Verella normalis Rumyantseva, Reference Rumyantseva and Verkhov1962; and V. spicata Dalmatskaya, Reference Dalmatskaya1951 (Fig. 7).
Biostratigraphic correlation
Plectostaffella jakhensis, one of the important taxon for indication of the Mississippian-Pennsylvanian boundary, was reported by Reitlinger (Reference Reitlinger and Rauzer-Chernousova1971) for the first time in the lower Bashkirian strata of southern Urals. Later, it was reported in the southern Urals (Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001) and Taurides (Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011). In this study, the first appearance of Plectostaffella jakhensis is in the strata below the appearance of Plectostaffella bogdanovkensis and Plectostaffella varvariensis, at the base of the Bashkirian (Fig. 5). This is very typical and widespread at the Serpukhovian-Bashkirian boundary strata and makes it easier to correlate this boundary with other regions.
The Syuranian Substage of the Bashkirian Stage is characterized by Plectostaffella bogdanovkensis, P. jakhensis, and P. varvariensis in the southern Urals (Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003; Kulagina et al., Reference Kulagina, Pazukhin, Nikolaeva and Kochetova2000, Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001, Reference Kulagina, Nikolaeva, Pazukhin and Kochetova2014), Elburz Mountains (Leven and Gorgij, Reference Leven and Gorgij2011), and Taurides (Atakul-Özdemir et al., Reference Atakul-Özdemir, Altıner, Özkan–Altıner and Yılmaz2011). In this study, the Serpukhovian-Bashkirian boundary is also recognized by the occurrence of these index taxa, which is similar to the southern Urals based on similar fusulinid assemblages and oolitic facies (Kulagina et al., Reference Kulagina, Gorozhanina, Nikolaeva and Gorozhanin2018; Kulagina and Gorozhanina, Reference Kulagina and Gorozhanina2019). The Akavasian Substage of the Bashkirian Stage is represented by the appearance of diverse species of the genera Pseudostaffella Thompson, Reference Thompson1942 and Varistaffella Kulagina and Sinitsyna, Reference Kulagina and Sinitsyna2003 with a distinct three-layered wall structure (Kulagina and Sinitsyna, Reference Kulagina and Sinitsyna2003), and its base is characterized by FAD of the Pseudostaffella antiqua in the southern Urals (Kulagina and Sinitsyna, Reference Kulagina, Sinitsyna, Ross, Ross and Brenckle1997, Reference Kulagina and Sinitsyna2003). In the Hadim Nappe sections, the Akavasian Substage shows close similarity to the southern Urals fusulinid fauna in terms of the diversity and abundance of the Pseudostaffella assemblage, and the appearance of Pseudostaffella antiqua at the base of this substage. The Askynbashian Substage is characterized by the appearance of more advanced fusulinids with distinct chomata, three-layered wall, and subspherical or short inflated fusiform test, such as in the genera Staffellaeformes (at the base of this substage) and Depratina (at the top of this substage) in the Bashkirian successions of the Hadim Nappe (Fig. 7). The base of the Askynbashian Substage in this study is determined by the FAD of the species Staffellaeformes staffellaeformis, which is known in the coeval interval in the southern Urals (Kulagina et al., Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001; Kulagina, Reference Kulagina2003, Reference Kulagina2008, Reference Kulagina2009), Elburz Mountains (Leven and Gorgij, Reference Leven and Gorgij2011), Pamir Mountains (Leven, Reference Leven2012), and Taurides (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007). Tikhonovichiella tikhonovichi and Verella spicata indicate the uppermost Bashkirian (Arkhangelskian Substage) in the Bashkirian–Moscovian transition. The former is determined in the lower part, and latter is in the upper part of the Arkhangelskian Substage in the Hadim Nappe sections. Tikhonovichiella tikhonovichi is recognized in the uppermost Bashkirian successions of the Urals (Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975; Kulagina et al., Reference Kulagina, Pazukhin, Kotschetkova, Sinitsyna and Kochetova2001; Kulagina, Reference Kulagina2003, Reference Kulagina2008), Pamir Mountains (Leven, Reference Leven2012), and Taurides (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007). The other important taxon, Verella spicata, is detected in this interval and occurs in coeval successions of the South Gissar and Zerafshan (Bensh, Reference Bensh1969; Saltovskaya, Reference Saltovskaya1974), Alai (Dzhenchuraeva, Reference Dzhenchuraeva1979), Cantabrian Mountains (Villa, Reference Villa1995), Pamir Mountains (Leven, Reference Leven2012), and Taurides (Dzhenchuraeva and Okuyucu, Reference Dzhenchuraeva and Okuyucu2007).
The faunas of the studied Hadim Nappe sections share many common species, mainly with southern Urals and other Tethyan regions.
Materials and methods
We studied a total of 31 limestone samples from the Yassıpınar and Gölbelen sections, analyzing two randomly oriented thin sections per sample and >300 oriented thin sections. A transmitted light microscope was used for examination of the thin sections and taking microphotographs of the fusulinids.
In this study, the systematic and taxonomic descriptions of fusulinids follow the handbook on taxonomy of the Paleozoic foraminifera proposed by Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996), with addition of the genus Varistaffella Kulagina and Sinitsyna, Reference Kulagina and Sinitsyna2003 proposed by Kulagina and Sinitsyna (Reference Kulagina and Sinitsyna2003).
Repository and institutional abbreviation
All holotypes and paratypes are reposited in the collection of the MA-(HY15) at the Faculty of Engineering and Natural Sciences, Department of Geological Engineering, Konya Technical University, Turkey.
Systematic paleontology
Class Foraminifera d’Orbigny, Reference d'Orbigny1826
Superorder Fusulinoidea Fursenko, Reference Fursenko1958
Order Fusulinida Fursenko, Reference Fursenko1958
Family Profusulinellidae Solovieva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996
Genus Depratina Solovieva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996
Type species
Schwagerina prisca Deprat, Reference Deprat1912, from China (Indochina), eastern Yun–Nan.
Remarks
The dominantly small-sized and nearly globular or short-inflated, fusiform-shaped Profusulinella group was originally determined as the “prisca group” by Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951). Later, Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996) assigned the “prisca group” to the genus Depratina. The species belonging to this genus differ from species of the genus Profusulinella, which have relatively globular or short-inflated fusiform tests, smaller L/D ratio, and poorly developed chomata. Hence, in this study, the species belonging to the “prisca” group (Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) and the species that have the same features are assembled under the genus Depratina.
Depratina convoluta (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930) new combination
Figure 10.10–10.13
- Reference Lee, Chen and Chu1930
Fusulinella (Neofusulinella) parva convoluta Lee and Chen in Lee et al., p. 119, pl. 7, figs. 1, 2.
- Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951
Profusulinella convoluta; Safonova and Rauzer-Chernousova in Rauzer-Chernousova et al., p. 160, pl. 13, figs. 10, 11.
- Reference Bensh1969
Profusulinella convoluta; Bensh, p. 112, pl. 2, figs. 9, 10.
- Reference Dzhenchuraeva1979
Profusulinella convoluta; Dzhenchuraeva, pl. 11, fig. 3.
- Reference Leven1998
Profusulinella convoluta; Leven, p. 17, pl. 1, fig. 44.
- Reference Leven, Davydov and Gorgij2006
Profusulinella convoluta; Leven et al., fig. 12.6.
- ?Reference Leven, Davydov and Gorgij2006
Profusulinella ex gr. P. convoluta; Leven et al., fig. 12.14.
- Reference Dzhenchuraeva and Okuyucu2007
Profusulinella convoluta; Dzhenchuraeva and Okuyucu, pl. 3, figs. 2, 3, pl. 4, fig. 5.
- Reference Fohrer, Nemyrovska, Samankassou and Ueno2007
Profusulinella convoluta; Fohrer et al., p. 43, fig. 23.1–23.10.
- ?Reference Fohrer, Nemyrovska, Samankassou and Ueno2007
Profusulinella cf. P. convoluta; Fohrer et al., p. 43, fig. 23.11.
- ?Reference Leven and Gorgij2011
Profusulinella (Profusulinella) ex gr. P. convoluta; Leven and Gorgij, pl. 4, fig. 11.
Holotype
From the Huanglung Limestone, China (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930, p. 119, pl. 7, fig. 1).
Occurrence
The Yassıpınar and Gölbelen sections of the Hadim Nappe (Hadim/Konya, Bozkır/Konya, Central Taurides), late Bashkirian–early Moscovian (Askynbashian–Vereian substages).
Description
Test small and inflated-fusiform in shape. Proloculus globular and small, with 28–60 μm external diameter. Length of test (L) 0.79–1.13 mm; diameter (D) 0.53–0.78 mm; L/D ratio 1.31–1.49. Adult tests have 4.5–5 volutions. The three-layered microgranular wall consists of thin and dark tectum, and thin lower tectorium and upper tectorium. Septa weak and plane in the middle part, and slightly fluted at the poles. Chomata asymmetrical and relatively massive.
Materials
Four axial sections (15Y17.04, 15Y18.03, 15Y23.09, and 18G26.09), all are reposited in the collection of the MA-HY15.
Remarks
This species was first described by Lee and Chen in Lee et al. (Reference Lee, Chen and Chu1930) under the name Fusulinella (Neofusulinella) parva convoluta, and then included in the “parva group” of Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) under the genus Profusulinella as Profusulinella convoluta. In this study, according to the systematics proposed by Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996), this species is assigned as the genus Depratina based on its short-inflated fusiform test and weak chomata compared to the genus Profusulinella. This systematic revision is also supported by the correlation graph between the L/D ratio of the type species of Depratina and Profusulinella, and L/D ratio of Depratina convoluta n. comb. (Fig. 12).

Figure 11. Thin-section photomicrographs of the fusulinid assemblages of the Yassıpınar (YS) and Gölbelen (GS) sections. (1–3) Depratina turani Akbaş n. sp., (1) subaxial section, 18G26.08, GS, (2, 3) detail view of the wall structure of the Depratina turani Akbaş n. sp., (2) 15Y17.08 (holotype); (3) 18G26.08 (three-layered wall including tectum, lower and upper tectorium). (4, 5) Staffellaeformes bona (Grozdilova and Lebedeva, Reference Grozdilova and Lebedeva1954), axial sections, (4) 15Y16A.19.02, YS; (5) 18G25.08, GS. (6, 7) Staffellaeformes parva parva (Lee and Chen in Lee et al., Reference Lee, Chen and Chu1930), axial sections, (6) 15Y16A.04, YS; (7) 18G19.04.01, GS. (8, 9) Staffellaeformes parva robusta (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936) n. comb., axial sections, (8) 15Y16A.06, YS; (9) 18G24.04, GS. (10–12) Staffellaeformes staffellaeformis (Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951), (10) subaxial section, 18G19.08.01, GS; axial sections, (11) 18G22.03, GS; (12) 18G23.07, GS. (13) Staffellaeformes tashliensis (Lebedeva in Grozdilova et al., Reference Grozdilova, Lebedeva, Lipina, Malakhova, Mikhailova, Chermnykh, Postoyalko, Simonova, Sinitsyna, Krylova, Grozdilova, Pozner, Sultanaev, Stepanov, Krylova, Grozdilova, Pozner and Sultanaev1975), axial section, 18G23.12, GS. (14) Aljutovella cf. A. pseudoaljutovica Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, axial section, 18G26.05, GS. (15–19) Tikhonovichiella praetikhonovichi Akbaş n. sp., axial sections, (15) 15Y18.05 (holotype), YS; (16) 15Y19.07, YS; (17) 15Y21.38, YS; (18, 19) detail view of the wall structure of the Tikhonovichiella praetikhonovichi Akbaş n. sp., (18) 15Y18.05 (holotype); (19) 15Y21.38 (three-layered wall including tectum, lower and upper tectorium). (20, 21) Tikhonovichiella tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951), (20) axial section, 15Y18.09, YS; (21) tangential section, 18G26.10, GS. (22, 23) Verella normalis Rumyantseva, Reference Rumyantseva and Verkhov1962, axial sections, (22) 15Y17.07, YS; (23) 18G25.07, GS. (24, 25) Verella spicata Dalmatskaya, Reference Dalmatskaya1951, axial sections, (24) 18G25.05, GS; (25) 18G25.14, GS. Scale bars = 500 μm (2–7, 10, 11, 18, 19); = 1 mm (1, 8, 9, 12–17, 20, 21); = 2 mm (22–25).

Figure 12. L/D Correlation graph between the type species of the Depratina and Profusulinella, and L/D ratio of Depratina convoluta n. comb.
Depratina turani Akbaş new species
Figures 10.21–10.23, 11.1–11.3
Holotype
15Y17.08 (axial section), from Dikenli Formation of the Hadim Nappe, southern Turkey, reposited in the Faculty of Engineering and Natural Sciences, Department of Geological Engineering, Konya Technical University, Turkey (collection number: MA-HY15/15Y17.08).
Diagnosis
Species of Depratina with small and sub-spherical test, low L/D ratio, and asymmetrical chomata.
Occurrence
The Yassıpınar and Gölbelen stratigraphic sections of the Hadim Nappe in Central Taurides (Konya, Turkey). Carboniferous, late Bashkirian, Arkhangelskian Substage.
Description
Test small and sub-spherical in shape. The middle part of the test is inflated and widely rounded. The lateral sides are convex and peripheries narrowly arched in shape. The initial one or two volutions are spherical, the latter sub-spherical due to slightly elongate, with 90° change in axis of coiling in some individuals. The progress of the test is gradual with tight coiling. The proloculus small- to medium-sized, with 50–110 μm external diameter. Length of test (L) 1.35–1.42 mm; diameter (D) 1.11–1.20 mm; L/D ratio 1.17–1.28. Adult tests have 5.5–6.5 volutions. Wall calcareous, thin, microgranular, three-layered with a distinct dark tectum, dark lower and upper tectorium. Septa weak and plane. Chomata massive, asymmetrical, and dominantly rounded in shape, but sub-ribbon in initial volutions of some individuals, and the thickness relatively decreases toward the poles.
Etymology
This species is named after Dr. Ahmet Turan, Konya Technical University, Konya, Turkey, in honor of his contributions to the geology of the Taurides.
Materials
Two axial sections (15Y17.03, 15Y17.08) and two subaxial sections (15Y17.06, 18G26.08), all are reposited in collection number MA-HY15.
Remarks
The described new species is most similar to the Depratina prisca sphaeroidea (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) based on the shape of the test and wall structure, but differs from it by tighter coiling, asymmetrical and dominantly rounded chomata, which became thinner toward the poles.
Genus Staffellaeformes Solovieva, Reference Solovieva1986
Type species
Profusulinella staffellaeformis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, from Russian Platform, Russia.
Remarks
Staffellaeformes staffellaeformis, which was originally described in the composition of the genus Profusulinella under the “parva group” (Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951), is assigned to the genus Staffellaeformes, which was described by Solovieva (Reference Solovieva1986) with Profusulinella staffellaeformis Kireeva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951 as the type species. Later, the original form in Solovieva's (Reference Solovieva1986) description of the Staffellaeformes was included in the systematics of Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996) as a separate genus. In this study, the validity and characteristics of the genus Staffellaeformes are accepted in the sense of the systematics of Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996). The species belonging to the genus Staffellaeformes differ easily from the species of the genus Profusulinella based on their sub-spherical or nautiloid tests, almost equal length and diameter, weak developed septa, and types of chomata.
Staffellaeformes parva robusta (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936) new combination
Figure 11.8, 11.9
- Reference Rauzer-Chernousova, Belyaev and Reitlinger1936
Profusulinella parva robusta Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., p. 178, pl. 1, fig. 4.
- Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951
Profusulinella parva robusta; Safonova in Rauzer-Chernousova et al., p. 158, pl. 13, fig. 7.
- Reference Dzhenchuraeva and Okuyucu2007
Profusulinella parva robusta; Dzhenchuraeva and Okuyucu, pl. 2, fig. 26.
Holotype
From upper Paleozoic deposits of the Pechora region, Russia (Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936, p. 178–179, pl. 1, fig. 4).
Occurrence
The Yassıpınar and Gölbelen sections of the Hadim Nappe (Hadim/Konya, Bozkır/Konya, Central Taurides), late Bashkirian (Askynbashian–Arkhangelskian substages).
Description
Test small and subspherical to ovoid in shape with early spherical volutions. Proloculus globular and small, with 50–60 μm external diameter. The length of test (L) 1.25–1.30 mm; diameter (D) 0.84–0.98 mm; L/D ratio 1.33–1.49. Adult tests have 4–5 volutions. The three layered microgranular wall consists of a thin and dark tectum, lower tectorium, and thin upper tectorium. Septa weak and plane in the middle part, and slightly fluted at the poles. Chomata massive and rounded in shape.
Materials
Two axial sections (15Y16A.06 and 18G24.04), all reposited in the collection number MA-HY15.
Remarks
This species was described by Rauzer-Chernousova and Belyaev (in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Belyaev and Reitlinger1936) as a species of the genus Profusulinella, but with its subspherical to ovoid test and weakly developed septa, it is included in the genus Staffellaeformes Solovieva, Reference Solovieva1986 in this study. This systematic revision is also supported by the correlation graph between the L/D ratio of the type species of Staffellaeformes and Profusulinella, and L/D ratio of Staffellaeformes parva robusta n. comb. (Fig. 13).

Figure 13. L/D Correlation graph between the type species of the Staffellaeformes and Profusulinella, and L/D ratio of Staffellaeformes parva robusta n. comb.
Family Aljutovellidae Solovieva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996
Genus Tikhonovichiella Solovieva in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996
Type species
Aljutovella tikhonovichi Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951, from the Russian Platform, Russia.
Tikhonovichiella praetikhonovichi Akbaş new species
Figure 11.15–11.19
Holotype
15Y18.05 (axial section), from Dikenli Formation of the Hadim Nappe, southern Turkey, reposited in the Faculty of Engineering and Natural Sciences, Department of Geological Engineering, Konya Technical University, Turkey (collection number: MA-HY15/15Y18.05).
Diagnosis
Species of Tikhonovichiella with small and inflated fusiform test and weakly developed septa and chomata.
Occurrence
The Yassıpınar section of the Hadim Nappe in Central Taurides (Konya, Turkey). Carboniferous, late Bashkirian–early Moscovian, Arkhangelskian–Vereian substages.
Description
Test small and fusiform to inflated fusiform in shape, with nautiloid early volutions. The middle part of the test is narrowly rounded. Lateral sides straight and the poles arched to slightly pointed. Proloculus globular and small in size with 32–60 μm external diameter. Length of test (L) 1.10–1.25 mm; diameter (D) 0.55–0.61 mm; L/D ratio 1.93–2.08. Adult tests have four volutions. The three-layered microgranular wall consists of a thin and dark tectum, distinct lower tectorium, and thin upper tectorium. Septa weak and plane in the middle part, and slightly fluted at the poles. Chomata massive and squared in shape.
Etymology
Latin prae, previous, similarity with Tikhonovichiella tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951).
Materials
Three axial sections (15Y18.05, 15Y19.07, 15Y21.38) and one tangential section, all are reposited in collection number MA-HY15.
Remarks
Tikhonovichiella praetikhonovichi Akbaş n. sp. is very close to Tikhonovichiella tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., Reference Rauzer-Chernousova, Gryzlova, Kireeva, Leontovich, Safonova and Chernova1951) based on small sizes and shape of chomata, but differs by more elongate test and higher L/D ratio. Although Tikhonovichiella tikhonovichi is known as a primitive representative of the genus Tikhonovichiella, this new species is more primitive than the former based on development of the chomata and septa.
Conclusions
This study concerns two sections (Yassıpınar and Gölbelen) of the upper Serpukhovian–Bashkirian succession of the Hadim Nappe in the Central Taurides. The Mississippian-Pennsylvanian boundary beds consist mainly of oolitic grainstone with rare siliciclastic intercalations at both sections. Whereas the lower Bashkirian is represented by quartz arenite sandstone at the Yassıpınar section, the coeval interval at the Gölbelen section is composed of bioclastic grainstone or bioclastic grainstone-packstone with diverse and abundant fauna and flora. The upper part of each section is characterized by medium-thick bedded, bioclastic grainstone, rarely packstone-bearing abundant fusulinid assemblages associated with a rich fauna and flora.
The fusulinid-based biostratigraphy of the two studied stratigraphic sections from the Hadim Nappe in the Central Taurides of Turkey has resulted in the definition of the upper Serpukhovian and Bashkirian (Syuranian, Akavasian, Askynbashian, and Arkhangelskian substages). The Mississippian-Pennsylvanian (Serpukhovian-Bashkirian) boundary is defined by the first appearance of the fusulinid Plectostaffella jakhensis in the Yassıpınar section. Due to extensive tectonic deformation, this boundary could not be determined in the Gölbelen section.
Based on the systematics proposed by Rauzer-Chernousova et al. (Reference Rauzer-Chernousova, Bensh, Vdovenko, Gibshman, Leven, Lipina, Reitlinger, Solovieva and Chediya1996) and this study, the taxonomic position of two fusulinid species (Depratina convoluta n. comb. and Staffellaeformes parva robusta n. comb.) have been revised.
Depratina turani Akbaş n. sp. and Tikhonovichiella praetikhonovichi Akbaş n. sp. are described for the first time from the upper Bashkirian (Arkhangelskian Substage) successions of the Hadim Nappe.
The upper Serpukhovian–Bashkirian fusulinid assemblages found in the studied sections of Hadim Nappe display very similar compositions at the genus and species levels to their counterparts from the other parts of the Tethyan Realm, including the Russian Platform, Donetz Basin, Pamir Mountains (Darvaz), Spain, central Iran, and especially the southern Urals.
Acknowledgments
This study was financially supported by Scientific Research Projects Coordination Unit of the Selcuk University, Konya, Turkey (Project no: 17101003). We are thankful to K. Sayit (METU) for improving the English of the text. Many thanks to the reviewers, E. Kulagina and P. Brenckle, for their constructive remarks and very useful comments that significantly improved the manuscript, and G.P. Nestell for the editorial handling.













