


INTRODUCTION
Approximately one-third of the known copepod species are parasites or commensals of invertebrates or fish (Humes, Reference Humes1994; Ho, Reference Ho2001). Some parasitic copepods of fish species, such as the members of the family Caligidae, often referred to as sea lice, are responsible for serious disease problems in fish farming (Pike and Wadsworth, Reference Pike and Wadsworth1999). The biology of parasitic copepods of invertebrates is much less well known than that of fish (Kabata, Reference Kabata1981). The best-documented parasitic copepods of bivalve molluscs belong to the order Poecilostomatoida (Ho, Reference Ho2000). Most of them are ectoparasites that are usually found in the mantle cavity of their hosts, but some cases of endoparasites have also been described (Lauckner, Reference Lauckner and Kinne1983). It is presumed that the majority of these copepods are not really harmful to their hosts (Lauckner, Reference Lauckner and Kinne1983). The damage caused by some of these copepods to their bivalve hosts is thought to be mainly caused by their appendages used for attachment and crawling. Nevertheless, some parasitic copepods have been associated with high mortality rates in bivalve molluscs (Ho and Zheng, Reference Ho and Zheng1994). It is not known if the variation of resistance or tolerance to parasitic copepods in bivalves is under genetic control and relatively little is also known about the defence mechanisms of bivalves to parasitic copepods.
The parasitic copepod Myicola ostreae belongs to the family Myicolidae and was originally described as a parasite of the Pacific oyster Crassostrea gigas in Japan by Hoshina and Sugiura (Reference Hoshina and Sugiura1953). Several bivalve hosts have been reported in Europe for M. ostreae after the introduction of C. gigas namely, the Portuguese oyster C. angulata (Comps, Reference Comps1972), the European flat oyster Ostrea edulis (His, Reference His1979) and the razor clam Sinonovacula constricta (Kim, Reference Kim2004). Gill lesions caused by M. ostreae have been observed in C. angulata and C. gigas and it is presumed that high infestation intensity may have some impact on the host (Comps, Reference Comps1972; His, Reference His1979). This copepod is presumed to be native from Asia as it is known to be present in Japan and Korea, countries where C. gigas is endemic (Hoshina and Sugiura, Reference Hoshina and Sugiura1953; Kim, Reference Kim2004). M. ostreae was probably accidentally introduced into France with the mass introduction of C. gigas from Japan in the 1970's (His, Reference His1979). Since then this copepod has been observed in several European countries (Streftaris et al. Reference Streftaris, Zenetos and Papathanassiou2005). Little is known about the biology of M. ostreae, but it is presumed to share common features with other members of the family Myicolidae having 6 nauplial and 5 copepodid stages before they reach the adult stage (Ho, Reference Ho2000). The infective stage is the first copepodid where individuals seek a suitable host.
C. angulata and C. gigas are two closely related edible cupped oysters of high commercial importance. They are presumed to be native to the North West Pacific region and were introduced into many countries (Ruesink et al. Reference Ruesink, Lenihan, Trimble, Heiman, Micheli, Byers and Kay2005). C. angulata was accidentally introduced in Europe from Asia, probably soon after the genesis of global shipping routes (Boudry et al. Reference Boudry, Heurtebise, Collet, Cornette and Gérard1998). C. gigas was voluntarily introduced into Europe in the late 1960's and early 1970's (Grizel and Héral, Reference Grizel and Héral1991). C. angulata and C. gigas interbreed readily in the laboratory (Menzel, Reference Menzel1974) as they are genetically very closely related, as revealed by allozymic markers (Buroker et al. Reference Buroker, Hershberger and Chew1979). Hybrids are fully fertile and no hybridization barrier has yet been demonstrated in the laboratory (Huvet et al. Reference Huvet, Balabaud, Bierne and Boudry2001, Reference Huvet, Gérard, Ledu, Phélipot, Heurtebise and Boudry2002). There is also evidence that the two taxa can hybridize in the wild, namely in Ria Formosa lagoon, Portugal (Huvet et al. Reference Huvet, Fabioux, McCombie, Lapègue and Boudry2004) where the present study was performed. Nevertheless, genetic (O'Foighil et al. Reference O'Foighil, Gaffney, Wilbur and Hilbish1998; Huvet et al. Reference Huvet, Lapègue, Magoulas and Boudry2000; Leitão et al. Reference Leitão, Chaves, Santos, Guedes-Pinto and Boudry2007), morphological (Batista et al. Reference Batista, Ben-Hamadou, Fonseca, Taris, Ruano, Reis-Henriques and Boudry2008) and physiological (Haure et al. Reference Haure, Huvet, Palvadeau, Nourry, Penisson, Martin and Boudry2003; Batista et al. Reference Batista, Leitão, Fonseca, Ben-Hamadou, Ruano, Henriques, Guedes-Pinto and Boudry2007) differences have been observed between the two taxa. In the present work we performed an experiment in the wild that allowed us to describe the infestation level of M. ostreae in the closely related cupped oysters C. angulata, C. gigas and their F1 hybrids. The current study allowed us to propose a genetic basis for resistance mechanisms to M. ostreae in cupped oysters. Moreover, histopathological analyses were also performed to examine host response mechanisms and impact on the host.
MATERIALS AND METHODS
Experimental procedure
Crassostrea angulata and C. gigas adults were collected from wild populations in Sado river (Portugal) and Seudre estuary (France), respectively. Full reciprocal crosses were carried out using 20 C. angulata individuals (15 females and 5 males) and 18 C. gigas individuals (10 females and 8 males) in order to produce the following groups: C. angulata (AA), C. gigas (GG), hybrids derived from C. angulata females and C. gigas males (AG), and hybrids derived from C. gigas females and C. angulata males (GA). Larval rearing was carried out in the shellfish hatchery of IFREMER Genetic and Pathology Laboratory (La Tremblade, France) and the juvenile rearing was performed in the shellfish hatchery of IPIMAR/INRB in Tavira (Portugal) as described by Batista et al. (Reference Batista, Leitão, Fonseca, Ben-Hamadou, Ruano, Henriques, Guedes-Pinto and Boudry2007). Four mesh bags with 50 oysters each per group were transferred from the Tavira shellfish hatchery to the Cacela-Velha site in Ria Formosa Lagoon (Portugal) on 3 March 2004. At the beginning of the experiment, the oysters from the 4 groups had approximately the same mean shell height and live weight and were 204 days old. Ria Formosa is a tidal lagoon system located on the southern coast of Portugal with a mean depth of 4 m and does not receive permanent fresh water streams. The Cacela-Velha site is in a mid-intertidal zone and the sea floor is a mixture of sand and mud. Salinity at the Cacela-Velha site during the present study was generally high (around 35 ppt) and temperature ranged between 12 and 25°C. Oyster bags were placed off-bottom (ca. 40 cm) on iron frame tables. The oysters were collected on 26 November 2004. In total 280 oysters were examined by macroscopic observation (between 11 and 22 individuals per bag) for the presence of parasitic copepods in their soft tissues and the number of copepods present in each individual was counted for 84 of them. The copepods observed were identified based on the description by Hoshina and Sugiura (Reference Hoshina and Sugiura1953). The parasite level in the oysters was assessed using 2 parameters: prevalence (percentage of infested hosts) and intensity (number of parasites per infested host). The dry shell weight of both valves of the oysters analysed was recorded to a precision of 0·001 g. For histological examination, a transverse section (ca. 5 mm thick) through the body, containing mantle, gonad, digestive gland and gills was done for each specimen. The tissues were then fixed in Davidson's solution for 48 h and preserved in 70% ethanol. Sections were cut at a thickness of 3–4 μm, deparaffinized, and stained with haematoxylin and eosin following standard techniques.
Data analysis
Data were analysed using one-way analysis of variance (ANOVA) to determine if there were significant differences among groups for dry shell weight, prevalence and intensity of M. ostreae using the oysters bags as the replicates (n=4). The Cochran test was used to assess heteroscedasticity. Pair-wise comparisons between groups were performed using Student-Newman-Keuls (SNK) tests when significant differences were revealed by ANOVA.
RESULTS
The morphological features of the female copepods observed were consistent with the characters proposed by Hoshina and Sugiura (Reference Hoshina and Sugiura1953) for M. ostreae, namely: female body cylindrical, with prosome swollen in ovigerous specimens, caudal ramus slender, with setae much shorter than ramus, the first antenna with 7 segments, the second antenna with 3 segments and maxilliped absent. The egg sacs were multiseriate. The mean length of the specimens observed was 2·1 mm and ranged between 1·8 and 2·5 mm (not including setae on caudal rami). The main morphological features that allowed us to distinguish the copepods observed in the present study from other species of the genus Myicola were: (1) size of ovigerous females (between 1·8 and 2·5 mm); (2) number of eggs in egg sac (about 40 eggs); (3) caudal ramus of females (length/width=5·0); and (4) second segment of the legs 5 of females (length/width=1·9). The copepods were observed in the gills of the oysters and no other organ appeared to be affected. All copepods observed were females that were carrying eggs. Some of the females carried eggs that when disturbed hatched free-swimming nauplius with a length of approximately 150 μm.
Significant differences in the prevalence of M. ostreae were observed among groups (F(3, 12)=40·4, P<0·001). In the C. angulata group (AA), the mean prevalence of M. ostreae was 71% whereas in AG and GA hybrids a slightly lower prevalence was observed (60 and 59%, respectively), but not significantly different from the AA group (SNK test; P>0·05). A highly significantly lower mean prevalence (SNK test; P<0·001) was observed in C. gigas progeny (GG; 23%) compared to the AA, AG and GA groups (Fig. 1A). The number of M. ostreae per oyster in C. angulata progeny ranged from 1 to 13, and in both AG and GA hybrids the number of copepods ranged between 1 and 8. In C. gigas progeny the number of parasitic copepods per host ranged between 1 and 5. Significant differences in the mean intensity were observed among groups (F(3, 12)=11·1, P<0·001). Pair-wise comparisons revealed no significant differences in intensity between the AA, AG and GA groups (SNK test, P>0·05). However, the infestation intensity was significantly lower (SNK test, P<0·05) in C. gigas in comparison with the other groups (Fig. 1B).

Fig. 1. Mean prevalence (A) and intensity (B) of Myicola ostreae in Crassostrea angulata (AA), C. gigas (GG) and their reciprocal hybrids (AG and GA). The error bars represent the standard deviation (n=4). Groups with different letters are significantly different (SNK test, P<0·05).
The area in the gills where M. ostreae was attached had a yellow/green aureole which could be associated with a haemocyte infiltration. Histological examination revealed copepods in the gills that were interpreted as being M. ostreae based on the macroscopic observations. Occasionally, haemocytic infiltrations and gill lesions were observed near the region where the copepods were attached and tissue debris was observed surrounding the copepods (Fig. 2). Some M. ostreae attached to the gills of C. gigas specimens (in ca. 20% of the individuals analysed) were involved by a massive agglomerate of haemocyte-like cells (Fig. 3). The external part of this formation was covered by a thin layer of fibroblast-like cells, similar to a capsule. No fibres were observed in the agglomerate of cells surrounding the copepods. This encapsulation response was only observed in C. gigas individuals.
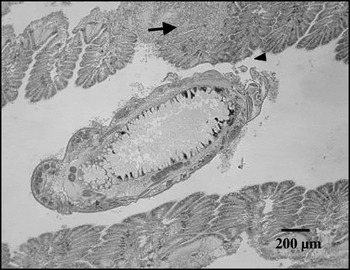
Fig. 2. A section of the gills of Crassostrea angulata showing a Myicola ostreae attached to gills (arrowhead) and haemocyte infiltration near that region (arrow). Haematoxylin-eosin staining.

Fig. 3. A section of (A) an adult female of Myicola ostreae observed in the gills of Crassostrea gigas involved with haemocyte-like cells, and (B) magnification of the cells surrounding the copepod. co, M. ostreae; eg, copepod eggs; ce, haemocyte-like cells; f, cell-like wall. Haematoxylin-eosin staining.
The shell dry weight (DSW) was significantly different among groups (F(3, 12)=52·7, P<0·001). The DSW of the GG group was significantly higher (SNK test; P<0·01) than the AA and AG groups, but not from the GA group (Fig. 4). The DSW of the AA group was significantly lower than the AG, GA and GG groups (SNK test; P<0·01). No significant differences were observed between the AG and GA groups (SNK test; P>0·05).
DISCUSSION
A higher prevalence and intensity of M. ostreae was observed in C. angulata than in C. gigas. We also reported that F1 C. angulata×C. gigas reciprocal hybrids had a pattern of susceptibility to M. ostreae that resembled the susceptible parent, which constitutes the first example to date of such a type of response (dominance for susceptibility) on hybrid animals (Fritz et al. Reference Fritz, Moulia and Newcombe1999; Wolinska et al. Reference Wolinska, Lively and Spaak2007). A putative defence mechanism, involving the encapsulation of the parasitic copepods at the gill surface, was only observed in C. gigas individuals. These results lead us to hypothesize that the differential susceptibility to M. ostreae between C. angulata and C. gigas may be ascribed to host defence factors.
Parasite level in C. angulata and C. gigas
The prevalence of M. ostreae in C. gigas observed in the present study (23%) was similar to that observed by Hoshina and Sugiura (Reference Hoshina and Sugiura1953), of 10–30%, and within the interval reported by His (Reference His1979) of 4–40%. We also observed an infestation intensity in C. gigas similar to that reported by His (Reference His1979) of 1 to 4 copepods per host. This suggests that the parasite levels observed in the Cacela-Velha site were within the values previously observed in C. gigas. However, parasite levels observed in C. angulata were significantly higher than in C. gigas. Since there are no published studies about the prevalence and intensity of M. ostreae in C. angulata it is not possible to assess whether the levels observed were abnormally high for this species or not.
Although the infestation mechanism of M. ostreae is not known, it may be assumed that it is similar to that observed in other parasitic copepods of bivalves in which a free-swimming copepodid enters the host by the inhalant current (Gee and Davey, Reference Gee and Davey1986). The infestation success is therefore generally associated with the amount of water filtered by the host and consequently by its size and age. At the end of the experiment, the dry shell weight of C. gigas individuals was significantly higher than C. angulata individuals (Fig. 4). Furthermore, it was shown that the mean filtration rate in C. gigas is equal or higher to that in C. angulata (His, Reference His1972; Haure et al. Reference Haure, Huvet, Palvadeau, Nourry, Penisson, Martin and Boudry2003). Hence, the higher parasite level observed in C. angulata when compared with C. gigas is unlikely to be due to differences in size and/or filtration rate. A possible explanation for the observed higher parasite level in C. angulata is that European populations of this taxon have not been in contact with M. ostreae since it was introduced into Europe from Asia (O'Foighil et al. Reference O'Foighil, Gaffney, Wilbur and Hilbish1998), and hence they could be seen as a naive host. Indeed, M. ostreae was not observed in Taiwan (Lin and Ho, Reference Lin and Ho1999) where C. angulata is known to be present and from where C. angulata is presumed to have been introduced into Europe (Boudry et al. Reference Boudry, Heurtebise, Collet, Cornette and Gérard1998).

Fig. 4. Mean dry shell weight of Crassostrea angulata (AA), C. gigas (GG) and their reciprocal hybrids (AG and GA). The error bars represent the standard deviation (n=4). Groups with different letters are significantly different (SNK test, P<0·05).
Parasite level in hybrids
Several studies have been conducted on parasitism of plant and animal hybrids (Fritz et al. Reference Fritz, Moulia and Newcombe1999; Moulia, Reference Moulia1999; Wolinska et al. Reference Wolinska, Lively and Spaak2007). Different patterns have been observed which imply different genetic mechanisms. Hybrids can be more susceptible (hybrid susceptibility pattern) or more resistance (hybrid resistance pattern) to parasites compared to parental taxa. Moreover, hybrids may resemble one of the parental taxa in resistance characteristic (dominance pattern). For this pattern, two different scenarios can be observed: (1) hybrids may resemble the susceptible parent (dominance for susceptibility); or (2) hybrids may resemble the resistant parent (dominance for resistance). Parasite levels in hybrids can also be intermediate between parental taxa (additive pattern). It seems that the hybrid susceptibility pattern is the most common one in animal hybrids and that the dominance pattern is the most unusual one (Fritz et al. Reference Fritz, Moulia and Newcombe1999). In the present study, the parasite level of reciprocal C. angulata×C. gigas F1 hybrids was similar to C. angulata but significantly higher than in C. gigas. These results suggest that susceptibility to M. ostreae is inherited as a dominant character in the F1 hybrids. To our knowledge all studies conducted with animal hybrids until present observed the dominance to resistant pattern (Fritz et al. Reference Fritz, Moulia and Newcombe1999; Moulia, Reference Moulia1999; Wolinska et al. Reference Wolinska, Lively and Spaak2007). On the contrary, the results of the present study support a hypothesis of dominantly inherited susceptible traits.
A study conducted with 2 closely related blue mussels revealed that susceptibility was related to a single parental genome (Coustau et al. Reference Coustau, Renaud, Maillard, Pasteur and Delay1991). It was observed that Mytilus edulis and introgressed individuals with a predominantly M. edulis genome were more susceptible to the trematode parasite Prosorhynchus squamatus than M. galloprovincialis. Since the putative different type of hybrids sampled in the wild (i.e., F1s, F2s, back-crosses or later-generation hybrids) were pooled together this prevents a direct comparison with our results. Nevertheless, a similar pattern was observed in the present study in which individuals with C. angulata genes had a higher parasite level than pure C. gigas individuals. One explanation for the lower parasite level observed in C. gigas in comparison with C. angulata and F1 hybrids might be that some host-specific factors required by M. ostreae are not present in C. gigas. Another explanation might be that putative anti-parasite factors produced by C. gigas are silenced or reduced in the hybrids leading to the higher parasite levels observed in the present study.
Effect on host and host response
In the present study, gill lesions and yellow/green aureoles were observed in the regions where M. ostreae copepods were attached. Comps (Reference Comps1972) also reported gill lesions and yellowish aureoles in the regions of attachment of M. ostreae in C. angulata individuals. In some cases histological examination revealed heavy haemocytic infiltration in the gills most probably caused by copepods. The gill lesions caused by M. ostreae were apparently not severe. Although the gill lesions caused by M. ostreae per se apparently have little impact on the host, maybe they can reduce host fitness and/or allow the introduction of pathogens especially when infestation intensity is high. Indeed, it has recently been suggested that ectoparasites can immunosuppress their invertebrate host (Yang and Cox-Foster, Reference Yang and Cox-Foster2005).
Phagocytosis and encapsulation are 2 major mechanisms of the cellular response in bivalve molluscs. Encapsulation is the mechanism by which haemocytes isolate or remove foreign intruders through the formation of a capsule-like envelope around them (Cheng and Rifkin, Reference Cheng, Rifkin and Snieszko1970). This host defence reaction is generally used against intruders that are too large to be phagocytosed. In the present study, single copepods were observed in the gills of C. gigas engulfed by a capsule-like structure composed by multiple cells but no fibres were observed. Five distinct types of encapsulation were described in bivalve molluscs in response to metazoan parasites by Cheng and Rifkin (Reference Cheng, Rifkin and Snieszko1970). The encapsulation complex observed in the present study resembles the ‘Leucocytic encapsulation’ type described by Cheng and Rifkin (Reference Cheng, Rifkin and Snieszko1970) that involves the aggregation of haemocytes to form a tunic surrounding the parasite.
Figueras et al. (Reference Figueras, Jardon and Caldas1991) observed in M. galloprovincialis mussels an unidentified copepod buried in the peripheral connective tissue of the digestive gland producing a strong haemocytic encapsulation response. A similar response was observed by Cáceres-Martínez and Vásquez-Yeomans (Reference Cáceres-Martínez and Vásquez-Yeomans1997) in M. galloprovincialis and M. californianus mussels described as a granuloma-like structure engulfing Pseudomyicola spinosus copepods in which haemocytes were involved. Copepods encapsulated by haemocytes were also observed in connective tissue surrounding the digestive gland of M. galloprovincialis (Olivas-Valdez and Cáceres-Martínez, Reference Olivas-Valdez and Cáceres-Martínez2002). The encapsulation response described in the 3 previous studies is similar to the one observed in the present study though the capsule-like structures were observed outside of oyster tissues at the gill surface. A similar host reaction was observed in the polychaete Spirographis spallanzani to the copepod Sabelliphilus sarsi (Carton, Reference Carton1967). Carton observed that the copepods were engulfed by an exudate of blood and coelomic cells leading to a reduction in the number of copepods infesting S. spallanzani. Other studies have also reported large differences in the encapsulation response between closely related taxa (Knopf and Mahnke, Reference Knopf and Mahnke2004; Prévost et al. Reference Prévost, Eslina, Dourya, Moreaub and Guillota2005).
The lower prevalence and intensity of M. ostreae observed in C. gigas than in C. angulata together with the observation of copepod encapsulation only in C. gigas lead us to hypothesize that the differential susceptibility to M. ostreae between the two oyster taxa may be ascribed to host defence factors. However, it remains unclear if, and how, the encapsulation response observed in C. gigas can eliminate adults of M. ostreae. Further insights into the putative cellular host response of C. gigas and C. angulata to M. ostreae, might be obtained by experimental infestation studies. It is known that resistance of parental taxa and hybrids to parasites may be influenced by genetic and environmental factors as well as by the physiological condition and age of the host (Fritz et al. Reference Fritz, Moulia and Newcombe1999). In the present study, the effects of these confounding factors were minimized since all animals were of the same age, were grown under identical conditions, were naturally infested by the parasite and the genetic status of the hybrids was known.
We thank Maurício Teixeira for his excellent assistance in maintaining spat in the nursery and progeny groups in the field. We also thank two anonymous referees for their comments that helped to improve a previous version of the manuscript. This work was made possible by an FCT grant (SFRH/BD/8972/2002) and an E.U. Marie Curie Training fellowship (PLUDAMOR, QLK5-CT-2000-60036) to F. M. B.




