



Introduction
Paromomyidae Simpson, Reference Simpson1940 is a family of plesiadapiforms known from North America (Silcox and Gunnell, Reference Silcox and Gunnell2008; Silcox et al., Reference Silcox, Rose and Bown2008), Europe (Russell et al., Reference Russell, Louis and Savage1967; Godinot, Reference Godinot1984; Estravís, Reference Estravís2000; Aumont, Reference Aumont2003; Marigó et al., Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012, Reference Marigó, Susanna, Minwer-Barakat, Madurell-Malapeira, Moyà-Solà, Casanovas-Vilar, Robles and Alba2014), and Asia (Tong and Wang, Reference Tong and Wang1998). Plesiadapiforms are herein considered stem primates (following Simpson, Reference Simpson1955; Van Valen and Sloan, Reference Van Valen and Sloan1965; Szalay, Reference Szalay1968, Reference Szalay1975a, Reference Szalay1975b; Gazin, Reference Gazin1971; Szalay et al., Reference Szalay, Tattersall and Decker1975, Reference Szalay, Rosenberger and Dagosto1987; Bown and Rose, Reference Bown and Rose1976; Radinsky, Reference Radinsky1982; Rose and Bown, Reference Rose and Bown1982; Silcox, Reference Silcox2001, Reference Silcox2007, Reference Silcox2008, Reference Silcox2017; Bloch and Boyer, Reference Bloch and Boyer2002, Reference Bloch and Boyer2007; Sargis, Reference Sargis2002; Bloch and Silcox, Reference Bloch and Silcox2006; Bloch et al., Reference Bloch, Silcox, Boyer and Sargis2007; Boyer and Bloch, Reference Boyer and Bloch2008; Silcox et al., Reference Silcox, Benham and Bloch2010, Reference Silcox, Sargis, Bloch and Boyer2015, Reference Silcox, Bloch, Boyer, Chester and López-Torres2017; Silcox and López-Torres, Reference Silcox and López-Torres2017). We acknowledge that some authors consider plesiadapiforms to be more distantly related to primates, with some or all taxa sharing a closer relationship to dermopterans (e.g., Beard, Reference Beard1989, Reference Beard1990, Reference Beard1991, Reference Beard1993a, Reference Beard1993b; Kay et al., Reference Kay, Thorington and Houde1990, Reference Kay, Thewissen and Yoder1992; Ni et al., Reference Ni, Gebo, Dagosto, Meng, Tafforeau, Flynn and Beard2013, Reference Ni, Li, Li and Beard2016). The debate over the broader relationships of plesiadapiforms does not impact the current study as it deals with relationships internal to Paromomyidae. The oldest paromomyid is Paromomys farrandi Clemens and Wilson, Reference Clemens and Wilson2009, which is reported from the early Paleocene (Torrejonian 1 North American Land Mammal Age [NALMA]) of northeastern Montana. The early age of this species, coupled with the fact that all of the most likely ancestors of paromomyids (i.e., members of the plesiadapiform families Purgatoriidae and Palaechthonidae) are also known from North America, is consistent with the hypothesis that this family had its origins on that continent. However, the paromomyids from Asia (Tong and Wang, Reference Tong and Wang1998) have never been formally described, which complicates the study of dispersals between North America and Asia. On the other hand, the European paromomyids have received more attention in the literature (Russell et al., Reference Russell, Louis and Savage1967; Godinot, Reference Godinot1981, Reference Godinot1984; Marandat, Reference Marandat1991; Estravís, Reference Estravís2000; Aumont, Reference Aumont2003, Reference Aumont2004; Hooker, Reference Hooker2010; Smith et al., Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011; Marigó et al., Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012), and the dispersal of these primates from North America to Europe is better understood. Recently, Hooker (Reference Hooker2015) formulated a two-phase model to account for mammalian dispersal patterns during the Paleocene-Eocene transition. The first (Phase 1) occurred during the latest Paleocene and brought to Europe euprimates, artiodactyls, and perissodactyls from Asia; rodents, pantodonts, oxyaenids, herpetotheriids, apatemyids, palaeanodonts, condylarths, neoplagiaulacids, viverravids, tillodonts from North America; and hyaenodontids from Africa. However, Beard et al. (Reference Beard, Wang, Meng, Ni, Gebo and Li2010) pointed out that direct migration between Asia and Europe would have been rather difficult because the Turgai Strait would have posed a significant marine barrier to terrestrial mammals. The second wave of immigrant species (Phase 2) to Europe from North America was possible due to increased temperatures during the Paleocene-Eocene Thermal Maximum (PETM) that allowed high latitude dispersal. The second dispersal included the marsupial Peradectes Matthew and Granger, Reference Matthew and Granger1921 and the mesonychid Pachyaena Cope Reference Cope1874 and is thought to have included paromomyids (Hooker, Reference Hooker2015; but see the following).
European paromomyids were first reported from Pourcy, France (Paleocene-Eocene Mammal Zone [PE] III), by Louis and Michaux (Reference Louis and Michaux1962) in a faunal list as an indeterminate species and genus of the ‘Phenacolemuridae,’ a family name now considered a junior synonym to Paromomyidae (Simpson, Reference Simpson1955). They were later ascribed to the North American paromomyid genus Phenacolemur Matthew, Reference Matthew1915 by Louis (Reference Louis1966), but no species level identification was given. Soon after, Russell et al. (Reference Russell, Louis and Savage1967) wrote the first comprehensive taxonomic study of European paromomyids in which they described two species from France: Phenacolemur fuscus from Mutigny, and Phenacolemur lapparenti from Avenay (PE V). Later, Godinot (Reference Godinot1984) described a new species, Arcius rougieri, from Palette (PE II), France. In that paper, Godinot (Reference Godinot1984) transferred the other two known species of European paromomyid into the newly named genus Arcius. Paromomyids have also been described from multiple early Eocene sites in France (Azillanet [Marandat, Reference Marandat1986], Condé-en-Brie [Louis, Reference Louis1966; Aumont, Reference Aumont2003, Reference Aumont2004], Fordones [Marandat, Reference Marandat1991], Fournes [Marandat, Reference Marandat1991], Gland [Aumont, Reference Aumont2003], Grauves [Louis, Reference Louis1970], Meudon [Russell et al., Reference Russell, Galoyer, Louis and Gingerich1988, Reference Russell, de Broin, Galoyer, Gaudant, Gingerich and Rage1990], Prémontré [Dégremont et al., Reference Dégremont, Duchaussois, Hautefeuille, Laurain, Louis and Tetu1985], Rians [Godinot, Reference Godinot1981], Sézanne [Louis, Reference Louis1970], Sotteville-sur-Mer [Smith et al., Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011], St. Agnan [Louis and Laurain, Reference Louis and Laurain1983], Venteuil [Aumont, Reference Aumont2003]), the United Kingdom (Abbey Wood [Hooker and Insole, Reference Hooker and Insole1980; Hooker, Reference Hooker1991, Reference Hooker1996, Reference Hooker1998]), Portugal (Silveirinha [Antunes, Reference Antunes1981; Estravís, Reference Estravís1992, Reference Estravís2000; Antunes et al., Reference Antunes, Casanovas, Cuesta, Checa, Santafé and Agustí1997]), and Spain (Masia de l’Hereuet [Marigó et al., Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012]). Godinot (Reference Godinot1984) hypothesized that all paromomyids from Europe belong to a single genus, Arcius.
A full analysis of the phylogenetic relationships of the European paromomyids has never been published, although some ideas about possible relationships have been expressed. Russell et al. (Reference Russell, Louis and Savage1967) suggested that the European paromomyids were closely related to Phenacolemur; Godinot (Reference Godinot1984) proposed A. rougieri as a potential ancestor to Arcius fuscus Russell et al., Reference Russell, Louis and Savage1967 and Arcius lapparenti Russell et al., Reference Russell, Louis and Savage1967 and that Arcius would be more closely related to Ignacius Matthew and Granger, Reference Matthew and Granger1921; Estravís (Reference Estravís2000) advocated that A. zbyszewskii Estravís, Reference Estravís2000 was the most basal species of Arcius. The only cladistic analysis of the European paromomyids was included by Aumont (Reference Aumont2003) in her unpublished dissertation, using 12 taxa and 35 dental characters. She found that A. fuscus and A. lapparenti formed a clade, with A. zbyszewskii as its sister taxon and A. rougieri being the most primitive lineage of the genus. The sister taxon for Arcius was the North American paromomyid Acidomomys hebeticus Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002.
This paper has three aims: (1) to revisit the alpha taxonomy of the European Paromomyidae, (2) to analyze the phylogenetic relationships among European paromomyids and their relationships to other members of the family using a data set that samples both characters and taxa more comprehensively than Aumont (Reference Aumont2003), and (3) to discuss the biogeographic implications of the phylogenetic analysis.
Repositories and institutional abbreviations
British Museum of Natural History (BMNH), London, UK; Institut Català de Paleontologia Miquel Crusafont (IPS), Sabadell, Spain; Muséum National d’Histoire Naturelle (Avenay collection, AV; Condé-en-Brie collection, CB; Mutigny collection, MU; Rians collection, RI), Paris, France; Royal Belgian Institute of Natural Sciences (Mammal collection, MAM), Brussels, Belgium; Universidade Nova de Lisboa (Silveirinha collection, SV), Lisbon, Portugal; Université de Montpellier (Fondones collection, FDN; Fournes collection, FRN; Palette collection, PAT), Montpellier, France; University of Alberta Laboratory of Vertebrate Paleontology (UALVP), Edmonton, Canada; University of California Museum of Paleontology (UCMP), Berkeley, USA; University of Michigan Museum of Paleontology (UM), Ann Arbor, USA; United States Geological Survey collection (USGS), Washington, USA; United States National Museum (USNM) Washington, USA; Yale Peabody Museum (Princeton University collection, YPM-PU), New Haven, USA.
Systematic paleontology
Order Primates Linnaeus, Reference Linnaeus1758
Superfamily Paromomyoidea (Simpson, Reference Simpson1940)
Family Paromomyidae Simpson, Reference Simpson1940
Arcius Godinot, Reference Godinot1984
1962 ‘Phenacolemuridae’ gen. indet. Reference Louis and MichauxLouis and Michaux, p. 171.
1966 Phenacolemur Reference LouisLouis, p. 51.
1967 Phenacolemur; Reference Russell, Louis and SavageRussell et al., p. 8, 12.
1970 Phenacolemur; Reference LouisLouis, p. 114.
1980 Phenacolemur; Reference Hooker and InsoleHooker and Insole, p. 38.
1981 Phenacolemur; Reference SilcoxAntunes, p. 257.
1981 Phenacolemur; Reference Antunes and RussellAntunes and Russell, p. 1101.
1981 Phenacolemur; Reference GodinotGodinot, p. 77.
1983 Phenacolemur; Reference Louis and LaurainLouis and Laurain, p. 9.
1984 Arcius Reference GodinotGodinot, p. 85.
1985 Phenacolemur; Reference Dégremont, Duchaussois, Hautefeuille, Laurain, Louis and TetuDégremont et al., p. 16.
1986 Arcius; Reference MarandatMarandat, p. 88.
1988 Arcius; Reference Russell, Galoyer, Louis and GingerichRussell et al., p. 432.
1991 Arcius; Reference MarandatMarandat, p. 92.
1996 Arcius; Reference HookerHooker, p. 209.
1998 Arcius; Reference HookerHooker, p. 449.
2000 Arcius; Reference EstravísEstravís, p. 283.
2003 Arcius; Reference AumontAumont, pl. 1–24.
2004 Arcius; Reference AumontAumont, fig. 3.
2010 Arcius; Reference HookerHooker, p. 48.
2011 Arcius; Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and YansSmith et al., fig. 19A, B.
2012 Arcius; Reference Marigó, Minwer-Barakat, Moyà-Solà and López-TorresMarigó et al., p. 430.
Type species
Arcius rougieri.
Other species
A. fuscus, A. lapparenti, A. zbyszewskii, Arcius hookeri n. sp., Arcius ilerdensis n. sp. (see descriptions below).
Emended diagnosis (modified from Godinot, Reference Godinot1984)
Presence of a molariform P4 (i.e., metacone approaching the size of paracone, expanded distolingual basin), not observed in any other paromomyid. Upper molars with distolingual basins less expanded distally than in Phenacolemur. Incisor with a lingual and a buccal crest, in contrast to Ignacius. The computed two-dimensional (2-D) area (max length × width of the crown in occlusal view) of P4 is similar to that of M1, in contrast to Ignacius. Lower molar cusps relatively taller relative to the base of the crown than those of Ignacius. Lower molar trigonids not as mesially inclined relative to the level of the base of the talonid basin as in any other paromomyid genus. Large third molar hypoconulid with a simple rounded lobe, in contrast to Phenacolemur, Ignacius, Acidomomys, and some species of Paromomys, which exhibit a central invagination (evident from a distal view) that runs mesiodistally on the hypoconulid lobe.
Occurrence
Neustrian (early Eocene) of France, United Kingdom, Portugal, and Spain; Grauvian (middle Eocene) of France.
Remarks
Following the classification of fossils from Condé-en-Brie to the genus Phenacolemur by Louis (Reference Louis1966), Russell et al. (Reference Russell, Louis and Savage1967) subsequently described new fossils from Mutigny and Avenay and classified them under that genus based on their resemblance to some of the North American representatives known at that time, such as Phenacolemur jepseni Simpson, Reference Simpson1955; Phenacolemur praecox Matthew, Reference Matthew1915; and Phenacolemur citatus Matthew, Reference Matthew1915. Russell et al. (Reference Russell, Louis and Savage1967) referred to the development of the hypoconulid in M3 in the European forms as reminiscent of that in Phenacolemur citatus and Phenacolemur jepseni, and the lack of ectocingulum in upper molars as similar to Phenacolemur jepseni. However, Russell et al. (Reference Russell, Louis and Savage1967) also mentioned that the P4 in the European specimens is smaller in size than the M1, a trait that is characteristic of Ignacius rather than Phenacolemur.
Following this line of reasoning, Godinot (Reference Godinot1984), in his paper naming the genus Arcius, suggested that it was closely related to Ignacius instead, specifically to Ignacius graybullianus Bown and Rose, Reference Bown and Rose1976 from the early Eocene of Wyoming. One of the characters that Godinot (Reference Godinot1984) highlighted as diagnostic of the genus was that the mandible (or, more appropriately, the alveolar ridge) reaches its highest point in the area of P4 and M1 alveoli, but at that time, the only mandible known was that of Arcius rougieri (Fig. 1). Still today, Arcius gnathic fossils are extremely rare, with the only ones reported being the mandible and maxillary fragments of A. rougieri (PAT1, PAT1 bis, and PAT2), two mandibles of A. lapparenti from Condé-en-Brie (CBX1-ph and an uncatalogued specimen [for the uncatalogued specimen, see Aumont, Reference Aumont2003, appendix G, pl. 3, fig. 3]), a maxilla and a mandible of Arcius sp. from Fournes (FNR43 and FNR54), an uncatalogued mandible of Arcius sp. from Le Quesnoy (Aumont, Reference Aumont2003, appendix G, pl. 3, fig. 4), and a mandible with M2-3 from Abbey Wood. The only lower jaw specimen known to exhibit the character described by Godinot (Reference Godinot1984) is PAT2 (Fig. 1; Arcius rougieri), and it is not present in other species (Aumont, Reference Aumont2003, appendix G, pl. 3, figs. 3 and 4). Therefore, this character is an autapomorphy of A. rougieri, and should not be considered diagnostic of the genus, but rather solely diagnostic of A. rougieri (see the following).
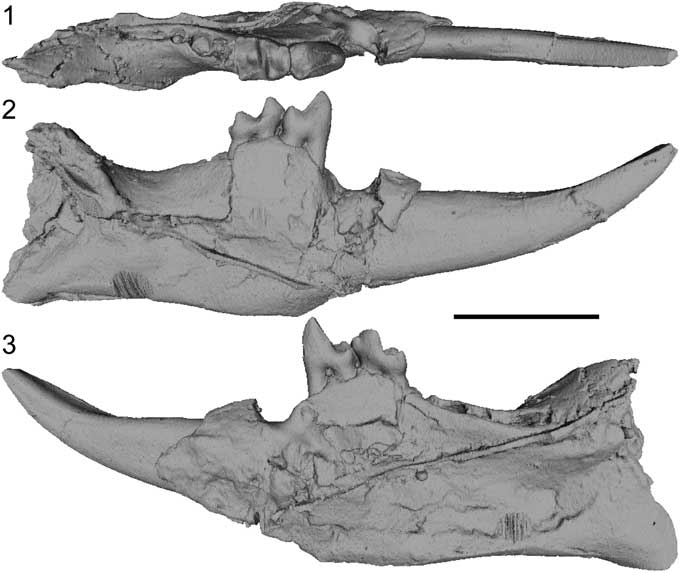
Figure 1 Micro-CT scan images of a cast of the mandible of Arcius rougieri Godinot, Reference Godinot1984 (PAT2) (1) in occlusal, (2) buccal, and (3) lingual views. Note that the strong line cutting through the mandible is a feature of the cast, not on the original specimen. Scale bar=0.5 cm.
Later, Robinson and Ivy (Reference Robinson and Ivy1994) suggested that Arcius was not monophyletic and classified this genus in the subfamily ‘Phenacolemurinae.’ According to Robinson and Ivy (Reference Robinson and Ivy1994), this subfamily contained the genera Phenacolemur, Ignacius, and Elwynella Rose and Bown, Reference Rose and Bown1982, along with their proposed genera ‘Simpsonlemur,’ ‘Pulverflumen,’ and ‘Dillerlemur,’ whose validity has been a source of debate (Silcox and Gunnell, Reference Silcox and Gunnell2008). By contrast, Aumont (Reference Aumont2003) recovered a monophyletic Arcius in her cladistic analysis, with the North American Acidomomys Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002 as its sister taxon.
Diagnostic characters for Arcius included here that have not been discussed previously in the literature include the simple enlarged hypoconulid lobe, the molariform P4, and the weak mesial inflection of the molar trigonids. With the exception of primitive representatives of the paromomyid clade (i.e., Paromomys farrandi), a markedly enlarged M3 hypoconulid lobe is typical of paromomyids. There are, however, two types of morphology of these lobes: simple and divided. Simple lobes, as observed in all species of Arcius, have a rounded shape in distal view, with a concave outline (Fig. 2.6–2.9). Divided lobes, as observed in Phenacolemur, Ignacius, Edworthia Fox et al., Reference Fox, Scott and Rankin2010, some species of Paromomys Gidley, Reference Gidley1923, and to a lesser extent, Acidomomys, exhibit an invagination of the occlusal surface that runs mesiodistally through the center of the lobe (Fig. 2.1–2.5). This gives the lobe a heart-shaped appearance in distal view. Arcius is also characterized by having a P4 that resembles a molar, more so than in other genera (Figs. 3.13, 3.17, 4.1). Although the P4 metacone is smaller than the paracone, it approaches the size of the paracone. This, combined with the similarity in size of P4 to M1 makes the premolar very molar-like. The similarity between the adult P4 to the M1 in Arcius parallels the resemblance in morphologies between the deciduous P4 and the M1 observed in other paromomyids, such as Phenacolemur (Rose, Reference Rose1981; Silcox et al., Reference Silcox, Rose and Bown2008) and Acidomomys (Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002). This might suggest that Arcius underwent a process of retention of a deciduous P4. Arcius rougieri shows a possible retention of the deciduous upper central incisor (see the following).
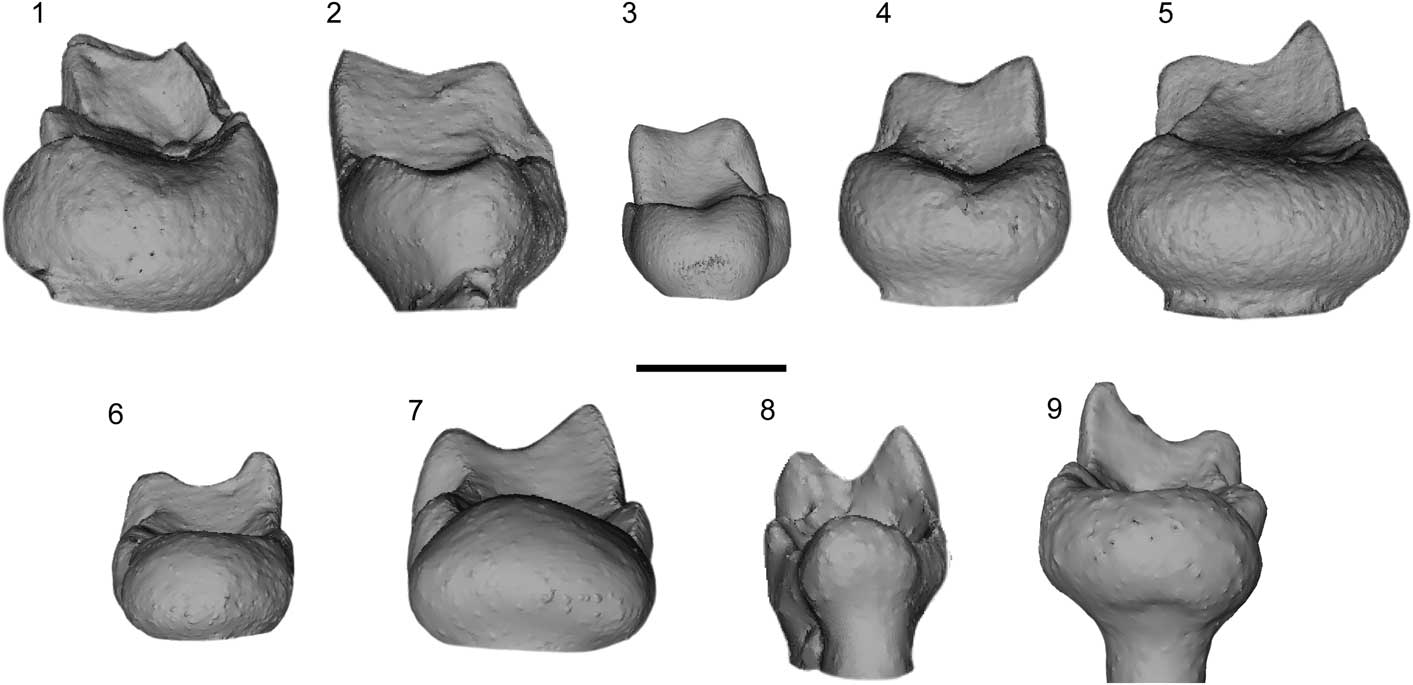
Figure 2 Distal view of micro-CT scan images of the M3 of various species of North American and European paromomyids. (1) Phenacolemur citatus Matthew, Reference Matthew1915, USGS 21712, right; (2) Paromomys depressidens Gidley, Reference Gidley1923, USNM 9677; (3) Edworthia lerbekmoi Fox et al., Reference Fox, Scott and Rankin2010, UALVP 50990, right; (4) Ignacius frugivorus Matthew and Granger, Reference Matthew and Granger1921, YPM-PU 20690, left; (5) Acidomomys hebeticus Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002, UM 108206, left; (6) Arcius fuscus Russell et al., Reference Russell, Louis and Savage1967, MU 6507, left; (7) Arcius lapparenti Russell et al., Reference Russell, Louis and Savage1967, AV 5849, left; (8) Arcius zbyszewskii Estravís, Reference Estravís2000, SV3-47, left; (9) Arcius hookeri n. sp., BMNH.M 44945, right. The paromomyids on the top row (1–5) have an invagination at the top central area of the hypoconulid lobe; all Arcius species (6–9) lack this feature. Scale bar=1 mm.
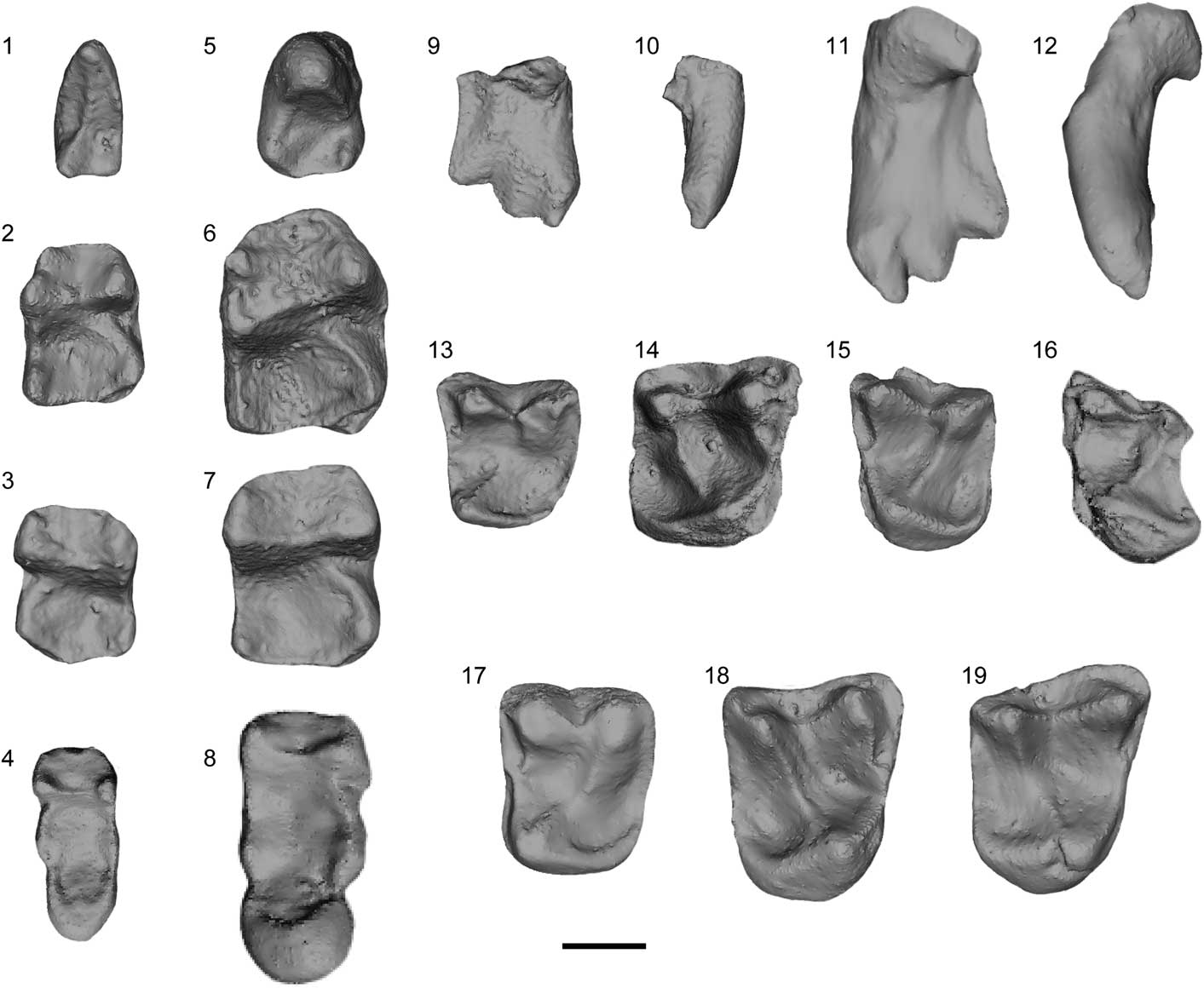
Figure 3 Micro-CT scan images; (1–4, 9, 10, 13–16) Arcius fuscus Russell et al., Reference Russell, Louis and Savage1967; (5–8, 11, 12, 17–19) Arcius lapparenti Russell et al., Reference Russell, Louis and Savage1967. (1) CB 2560, left P4, occlusal view; (2) MU 6458, right M1, occlusal view; (3) AV 422-L, left M2, occlusal view; (4) MU 6507, holotype, left M3, occlusal view; (5) CB 4162, left P4, occlusal view; (6) AV 7716, right M1, occlusal view; (7) AV 7707, right M2, occlusal view; (8) AV 5849, left M3, holotype, occlusal view; (9, 10) AV 6838, left I1, (9) lingual view, (10) mesial view; (11, 12) AV 7714, right I1, (11) lingual view, (12) mesial view; (13) CB 1914-L, left P4, occlusal view; (14) UCMP 71976, left M1, occlusal view; (15) AV 610-BN, left M2, occlusal view; (16) UCMP 71982, left M3, occlusal view; (17) CB 232-BN, left P4, occlusal view; (18) AV 1306-Ph, right M1, occlusal view; (19) AV 1092-BN, right M2, occlusal view. Scale bar=1 mm.
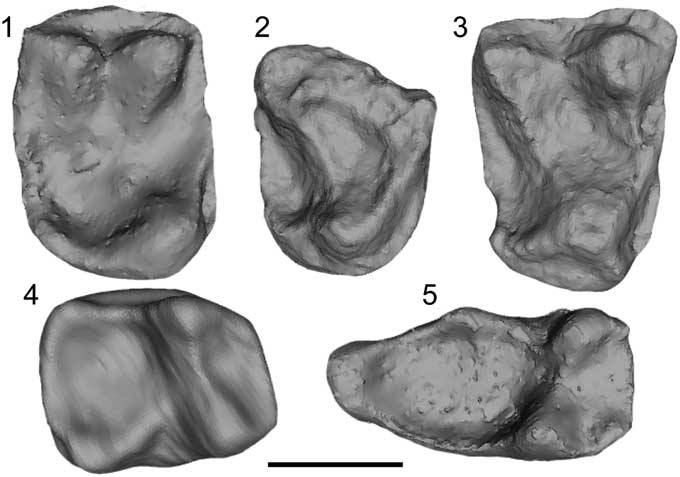
Figure 4 Micro-CT scan images of Arcius zbyszewskii Estravís, Reference Estravís2000. (1) SV2-99, right P4, occlusal view; (2) SV3-125, right M1 fragment, occlusal view; (3) SV1-29, right M2, holotype, occlusal view; (4) SV3-317, right M1, occlusal view; (5) SV3-47, left M3, occlusal view. Scale bar=1 mm.
Based on the two most complete specimens that belong to the genus Arcius, PAT1 and PAT2, the dental formula for this genus would be
 ${{2.1.2.3} \over {1.0.1.3}}$
. See also Table 1 for estimated body masses for all the species in the genus.
${{2.1.2.3} \over {1.0.1.3}}$
. See also Table 1 for estimated body masses for all the species in the genus.
Table 1 Estimated average body masses of all species of Arcius Godinot, Reference Godinot1984 based on data from Aumont (Reference Aumont2003) and the current study (for A. hookeri n. sp. and A. ilerdensis n. sp.). The regression equation used for Conroy’s (Reference Conroy1987) estimates was the one based on the prosimian sample.

Arcius rougieri Godinot, Reference Godinot1984

Figure 5 Micro-CT scan images of Arcius rougieri Godinot, Reference Godinot1984. Maxilla (PAT1, holotype) in (1) occlusal, (2) oblique occlusal, (3) oblique frontal views; (4) PAT5, right M2, occlusal view; (5) PAT3, left M3, occlusal view. (1–3) Scale bar=0.5 cm; (4, 5) scale bar=1 mm.
1981 Arcius cf. A. fuscus Godinot, Reference Godinot1981, p. 77.
1984 Arcius rougieri Godinot, Reference Godinot1984, p. 85.
1991 ?Arcius rougieri; Marandat, Reference Marandat1991, p. 92.
2003 Arcius rougieri; Aumont, Reference Aumont2003, pl. 3, fig. 5, pl. 6, fig, 5, 6. Pl. 18, fig. 1, pl. 19, 23.
Holotype
PAT1, partial maxilla with right I1–C and left I1–M1.
Emended diagnosis (modified from Godinot, Reference Godinot1984)
Smaller M2 than Arcius lapparenti, A. hookeri n. sp., and A. ilerdensis n. sp., but larger M1 than A. zbyszewskii. Further differs from A. fuscus and A. lapparenti in having a more apically extended (taller) anterocone than mediocone on I1, with a significantly smaller posterocone, and laterocone proximally shifted relative to the anterocone. Further differs from all other species of Arcius in having a more buccally extended parastylar region on M1 and a stepped postvallid on M1 and M2. Further differs from A. fuscus in having a more distally expanded distolingual basin on M3. Further differs from A. fuscus in having two crests, instead of three, on the P4 postvallid. Paraconid of M1 is smaller than in A. fuscus and A. lapparenti but larger than in A. zbyszewskii. Mesial inflection of M1 and M2 trigonids weaker than in A. ilerdensis n. sp. but stronger than in the other known species of Arcius. Further differs from A. zbyszewskii and A. ilerdensis n. sp. in having a cingulid on the buccal half of the distal aspect of M1 that runs up to the hypoconulid. Highest point of the alveolar ridge between P4 and M1; this has not been observed in any other species of Arcius.
Horizon and locality
Palette (type locality, PE II, see Biostratigraphy in the following), Bouches-du-Rhône, France; Fordones (PE II, see Biostratigraphy), Aude, France; Rians (PE II; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Var, France.
Biostratigraphy
There is some disagreement as to the age of Palette and Fordones. Marandat et al. (Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012) assigned Palette and Fordones to Paleocene-Eocene biozone I (PE I) and considered them intermediate in age between Silveirinha (older) and Rians/Fournes (younger). However, there seems to be a consensus that Silveirinha is very close in age to Sotteville-sur-Mer, probably slightly younger (Smith et al., Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012; Hooker, Reference Hooker2015), and Hooker (Reference Hooker2015) assigned Sotteville-sur-Mer to PE II, close to the onset of the carbon isotope excursion (CIE). Rians and Fournes have been calibrated to be of similar age to the site of Meudon (Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), and Hooker (Reference Hooker2015) suggested PE II as the age for Meudon as well. Therefore, based on the correlations suggested by Hooker (Reference Hooker2015), Palette and Fordones would be considered to be PE II in age.
Materials
PAT2, mandible with right I1–M1; PAT3, left M3; PAT5, right M2.
Remarks
In this paper, all diagnoses for the six species of Arcius include diagnostic features of the referred material along with the holotype. The reason behind this decision is that the holotypes for A. fuscus and A. lapparenti are M3 specimens (Russell et al., Reference Russell, Louis and Savage1967), and this tooth position is probably the most variable in paromomyids, in particular with respect to the relative proportions of the hypoconulid lobe with the rest of the tooth. This would make the diagnosis for these species solely based on M3 morphology very limited and not very diagnostic.
The holotype of Arcius rougieri, along with PAT1 bis and PAT2, represent the first described gnathic fossils with associated teeth in a European paromomyid. The I1 of A. rougieri differs greatly from those of A. fuscus and A. lapparenti (Rose et al., Reference Rose, Beard and Houde1993) and is more similar to the deciduous I1 of A. fuscus and A. lapparenti (Fig. 6) in that the anterocone is the tallest cusp rather than the mediocone (tallest in adult specimens attributed to A. fuscus and A. lapparenti). In addition, the laterocone is more proximally located to the mediocone in juveniles of A. fuscus and A. lapparenti than in adult representatives of the tooth, as is observed in the adult form of A. rougieri. There are some differences, however, between the I1 of A. rougieri and the deciduous I1 of A. fuscus and A. lapparenti in that the mediocone is better developed in juveniles of A. fuscus and A. lapparenti while the posterocone is less so, although the posterocone is still poorly expressed in A. rougieri compared to the condition in most adult paromomyid I1s. PAT1 is inferred to pertain to an adult because all of its adult teeth have erupted. The second upper incisor and the fourth upper premolar erupt after I1 in the North American paromomyid Acidomomys (Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002), and both teeth are fully erupted in PAT1. Although it is possible that the dental eruption sequence varied in Arcius from that observed in Acidomomys, it would be surprising for it to vary so much that I1 would erupt after all the adult teeth, particularly since no primate species is known to have I1 as the last tooth to erupt. Indeed, I1 is usually one of the first teeth to erupt (López-Torres et al., Reference López-Torres, Schillaci and Silcox2015, table 1). Therefore, it seems that the upper central incisors of A. rougieri were retained. Interestingly, as noted, the genus Arcius also has an adult P4 that resembles a deciduous P4 (or an adult molar).

Figure 6 Drawing of a decidious upper central incisor of Arcius lapparenti Russell et al., Reference Russell, Louis and Savage1967 (GR10029) in (1) medial view, (2) lingual view, (3) oblique (mediolingual) view. Original photograph in Aumont (Reference Aumont2003, appendix G, pl. 9, fig. 8). Scale bar=1 mm. Drawing by V. Bodo.
Previous to the description of Arcius, Godinot (Reference Godinot1981) described a paromomyid M1 from Rians (RI225) that he tentatively ascribed to Phenacolemur cf. P. fuscus. Godinot (Reference Godinot1981) highlights some differences between the Rians specimen and the specimens from Mutigny (type locality of Arcius fuscus), such as the Rians specimen having a paraconid closer to metaconid, a paracristid curved downward, and a greater mesiodistal length of the whole tooth. Godinot (Reference Godinot1981) admits that high intraspecific variability for A. fuscus could allow this tooth to be attributed to that species but suggests that new fossils could shed light on “l’espèce de Rians” (“the species of Rians”; Godinot, Reference Godinot1981, p. 78). The paraconid on RI225 is weak, which contrasts with the fairly strong paraconids on the M1 of A. fuscus and A. lapparenti. The only Arcius species with poorly expressed paraconids on M1 and a size comparable to A. fuscus is A. rougieri. Therefore, we suggest that the RI225 belongs to A. rougieri.
Marandat (Reference Marandat1991) tentatively ascribed to A. rougieri fossils found in the locality of Fordones. The Fordones fossils include the only known M3 for this species (FDN28). This specimen is missing the hypoconulid lobe, which is particularly relevant in determining which species of Arcius it belongs to. Aumont (Reference Aumont2003) suggested that the only way to know the correct ascription of FDN28 is to find an M3 from the type locality for A. rougieri (Palette), but the size, the metric proportions, and the fact that other A. rougieri fossils have been found in Fordones support the inference that FDN28 is the first M3 known of A. rougieri (Marandat, Reference Marandat1991; Aumont, Reference Aumont2003).
Arcius fuscus Russell, Louis, and Savage, Reference Russell, Louis and Savage1967
Figures 2.6, 3.1–3.4, 3.9, 3.10, 3.13–3.16, 7.2, 7.5
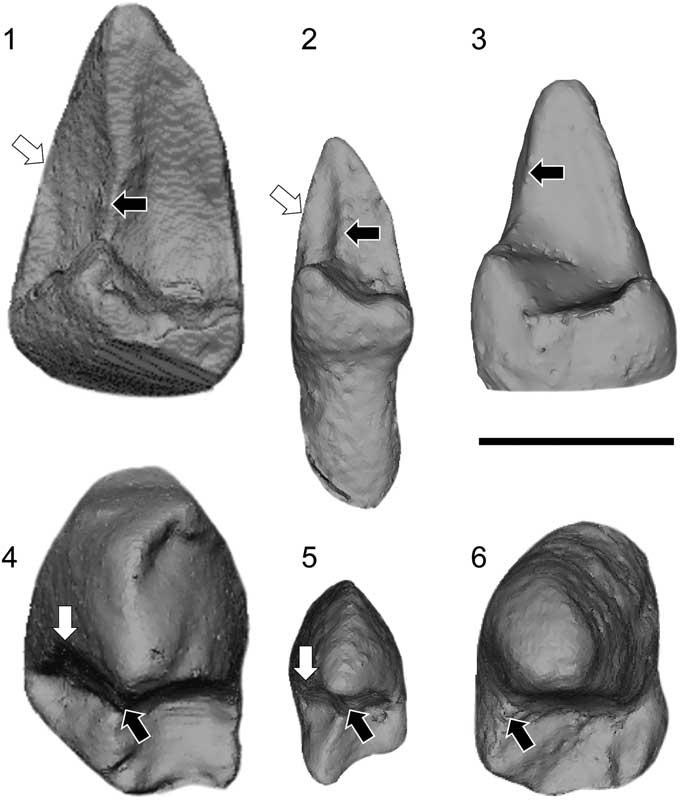
Figure 7 Micro-CT scan images of three paromomyid P4s. (1, 2, 4, 5) The ‘prehypoflexid cristid’ (white arrow) as seen on the P4 of (1, 4) Paromomys maturus Gidley, Reference Gidley1923 and (2, 5) Arcius fuscus Russell et al., Reference Russell, Louis and Savage1967. (3, 6) Arcius lapparenti Russell et al., Reference Russell, Louis and Savage1967 is shown as an example of a paromomyid with no prehypoflexid cristid. When the prehypoflexid cristid is present, the postprotocristid (black arrow) is shifted to a central position across the postvallid. The main difference between the prehypoflexid cristid of Paromomys maturus and that of A. fuscus is that in (4) Paromomys maturus it is located more mesially, contrary to (5) A. fuscus, in which the three cristids are in the same plane on the postvallid. Scale bar=1 mm.
1962 ‘Phenacolemuridae’ gen. indet. sp. indet. Reference Louis and MichauxLouis and Michaux, p. 171.
1966 Phenacolemur sp. Reference MatthewLouis, p. 51.
1967 Phenacolemur fuscus (in part) Reference Russell, Louis and SavageRussell et al., figs. 2a–j, l, 3d, g.
1981 Phenacolemur cf. P. fuscus Reference GodinotGodinot, p. 77.
1984 Arcius fuscus Reference GodinotGodinot, fig. 2b.
1991 Phenacolemur cf. P. fuscus Reference MarandatMarandat, p. 92.
2003 Arcius fuscus; Reference AumontAumont, pl. 1, figs. 3, 4, 8, pl. 2, figs. 9–12, pl. 4, figs. 4, 5, pl. 5, figs. 5, 6, 10, pl. 7, figs. 8–15, pl. 8, figs. 3, 8, 9, 13, 14, pl. 10, figs. 6–8, pl. 11, fig. 2, pl. 12, figs. 4, 6, pl. 13, figs. 3–5, 10, pl. 14, fig. 3.
2004 Arcius fuscus; Reference AumontAumont, fig. 3.5–3.9, 3.14–3.16.
Holotype
MU 6507, left M3.
Emended diagnosis (modified from Russell et al., Reference Russell, Louis and Savage1967 and Aumont, Reference Aumont2003)
Smaller M2 than those of Arcius lapparenti, A. hookeri n. sp., and A. ilerdensis n. sp. but larger M1 than those of A. rougieri and A. zbyszewskii. Further differs from A. rougieri in having a more apically extended (taller) mediocone than anterocone on I1, no expansion of the parastylar area of M3, presence of ‘prehypoflexid cristid’ on the P4, a concave mesial border of P4 in lateral view, stronger mesial inflection of molar trigonids, and lacking a stepped postvallid on M1. Further differs from A. zbyszewskii and A. ilerdensis n. sp. in having a cingulid on the buccal half of the distal aspect of M1 that runs up to the hypoconulid. Further differs from A. zbyszewskii in not having a paraconid on M3.
Occurrence
Mutigny (type locality, PE IV; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Marne, France; Fournes (PE II; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Hérault, France; Var, France; Gland (PE V; Aumont, Reference Aumont2003), Aisne, France; Avenay (PE V; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Marne, France; Sézanne (PE V; Aumont, Reference Aumont2003), Aisne, France; Condé-en-Brie (PE V; Hooker and Russell, Reference Hooker and Russell2012), Aisne, France; St. Agnan (MP10; Hooker, Reference Hooker1996), Aisne, France.
Materials
AV 422-L, left M2; AV 610-BN, left M2; AV 6838, left I1; CB 1914-L, left P4; CB 2560, left P4; MU 6458, right M1; UCMP 71976, left M1; UCMP 71982, left M3.
Remarks
Arcius fuscus, along with A. lapparenti, is one of the most common species of the genus found in Europe. When it was first described by Russell et al. (Reference Russell, Louis and Savage1967), A. fuscus was thought to come just from its type locality, Mutigny (Marne, France). Arcius lapparenti was also described for the first time by Russell et al. (Reference Russell, Louis and Savage1967) and at that time was thought to come only from its type locality, Avenay (Marne, France). However, Aumont (Reference Aumont2003, Reference Aumont2004) argued these two species were sympatric across many sites in France. Aumont (Reference Aumont2003) also interpreted the Arcius fuscus sample from Russell et al. (Reference Russell, Louis and Savage1967) to be a mixture of both A. fuscus and A. lapparenti and considered the only two P4 specimens (MU5627 and MU6294) found at Mutigny to belong to A. lapparenti, which is significant since P4 is generally the most diagnostic tooth for paromomyids. Aumont (Reference Aumont2003) came to that conclusion after studying the large sample from Condé-en-Brie (Aisne, France), where there are two distinct P4 sizes and morphs.
Russell et al. (Reference Russell, Louis and Savage1967) did not choose a P4 as the holotype for Arcius fuscus but instead selected an M3. They used differences in the inclination of the M3 postvallid, and in the degree of lateral projection of the entoconid, to differentiate A. fuscus from A. lapparenti. However, after the study of the large collection from Condé-en-Brie, it seems as though there is too much variability in these characters for them to be considered diagnostic of species of Arcius. Although form of the M3 hypoconulid is diagnostic for the genus as a whole, variability in this tooth position implies that distinguishing among species in the genus Arcius also requires consideration of characters from the premolars and other molars.
There are two genera of paromomyids that have three distinct crests on the postvallid of the P4 of some species: Arcius and Paromomys. These crests are interpreted here as a metacristid (this term used in preference to premetacristid or postmetacristid because of the absence of a metaconid; in this case, ‘metacristid’ is used for a crest in the metaconid position), a postprotocristid, and a third crest that we are referring to here as a ‘prehypoflexid cristid.’ These crests can be distinguished in the following way: (1) the metacristid runs lingually from the protoconid to the entocristid or to the lingual aspect of the bottom of the postvallid (if no entocristid is present); (2) the postprotocristid runs buccally or centrally from the protoconid to the cristid obliqua; and (3) the ‘prehypoflexid cristid’ runs buccally from the protoconid to the hypoflexid. The main difference between these two taxa is that all three crests are in the same plane in Arcius fuscus, but the ‘prehypoflexid cristid’ is less well defined and more distally located in Paromomys (Fig. 7). Only two crests are visible on the postvallid in all other known paromomyid species.
Arcius lapparenti Russell, Louis, and Savage, Reference Russell, Louis and Savage1967
Figures 2.7, 3.5–3.8, 3.11, 3.12, 3.17–3.19, 6, 7.3, 7.6
1967 Phenacolemur lapparenti (in part) Reference Russell, Louis and SavageRussell et al., figs. 2k, 3a–c, e, f.
1984 Arcius lapparenti Reference GodinotGodinot, fig. 2c–f.
2003 Arcius lapparenti; Reference AumontAumont, pl. 1, figs. 1, 2, 5–7, pl. 2, figs. 1–8, pl. 3, figs. 1–3, pl. 4, figs. 1–3, 6–8, pl. 5, figs. 1, 2, 7–9, pl. 6, figs. 1–4, pl. 7, figs. 1–7, pl. 8, figs. 1, 2, pl. 9, pl. 10, figs. 1–5, pl. 11, figs. 1, 9, pl. 12, figs. 1–3, 5, 7–12, pl. 13, figs. 1, 2, 6–9, 11–13, pl. 14, figs. 1, 2, 4–8, pl. 15, figs. 1, 2, pl. 16, 17, pl. 18, figs. 3, 4.
2004 Arcius lapparenti; Reference AumontAumont, fig. 3.1–3.4, 3.10–3.13.
Holotype
AV 5849, left M3.
Emended diagnosis (modified from Russell et al., Reference Russell, Louis and Savage1967 and Aumont, Reference Aumont2003)
Largest species of Arcius. Further differs from A. rougieri in having a more apically extended (taller) mediocone than anterocone on I1, no expansion of the parastylar area of M3, and stronger mesial inflection of molar trigonids and lacking a stepped postvallid on M1. Further differs from A. fuscus in lacking a ‘prehypoflexid cristid.’ Well-developed paraconid on M1, but proportionally smaller relative to the metaconid than in A. fuscus. Further differs from A. zbyszewskii and A. ilerdensis n. sp. in having a cingulid on the buccal half of the distal aspect of M1 that runs up to the hypoconulid. Further differs from A. zbyszewskii in not having a paraconid on M3.
Occurrence
Avenay (type locality, PE V; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Marne, France; Fournes (PE II; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Hérault, France; Meudon (PE II; Hooker, Reference Hooker2015), Hauts-de-Seine, France; Pourcy (PE III [55.12 mya]; Hooker, Reference Hooker2010), Marne, France; Mutigny (PE IV; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), Marne, France; Gland (PE V; Aumont, Reference Aumont2003), Aisne, France; Sézanne (PE V; Aumont, Reference Aumont2003), Aisne, France; Condé-en-Brie (PE V; Hooker and Russell, Reference Hooker and Russell2012), Aisne, France; Venteuil (MP10; Aumont, Reference Aumont2003), Marne, France; St. Agnan (MP10; Hooker, Reference Hooker1996), Aisne, France; Prémontré (MP10 [48.4 mya]; Franzen, Reference Franzen2005), Aisne, France.
Materials
AV 1092-BN, right M2; CB 232-BN, left P4; AV 7707, right M2; AV 7714, right I1; AV 7716, right M1; CB 3583, right M3; CB 4162, left P4; CBX 2-Ph, I1; AV 1306-Ph, right M1.
Remarks
Along with A. fuscus, A. lapparenti is one of the most common species of this European genus. While Russell et al. (Reference Russell, Louis and Savage1967) described A. fuscus as appearing at Mutigny, A. lapparenti was argued by the same authors to have been present in Avenay, very close geographically to Mutigny.
When Aumont (Reference Aumont2003, Reference Aumont2004) suggested the sympatry of these two species, she also reported that the samples from the type localities of both species were mixed. In addition, the molariform morphology of Arcius fourth upper premolars led to misidentification in the original Russell et al. (Reference Russell, Louis and Savage1967) paper. AV 5775, reported in Russell et al. (Reference Russell, Louis and Savage1967) as an M1 of A. lapparenti, is interpreted here to be a P4 based on its poorly developed postprotocrista, a feature of P4 in other species of Arcius. These two teeth can be distinguished mainly by the lack of a postprotocrista and by the slightly convex mesial aspect of the tooth on P4. Whereas relative size of the metacone to the paracone is often used to discriminate between M1 and P4 in primate species, the paracone of the P4 is so close in size to the metacone in A. lapparenti that it is quite challenging to see a size difference between these two cusps.
Arcius zbyszewskii Estravís, Reference Estravís2000
1981 Phenacolemur sp. Reference AntunesAntunes, p. 257.
1981 Phenacolemur cf. P. fuscus Reference Antunes and RussellAntunes and Russell, p. 1101.
2000 Arcius zbyszewskii Reference EstravísEstravís, p. 283.
2003 Arcius zbyszewskii; Reference AumontAumont, pl. 24.
2011 Arcius sp.; Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and YansSmith et al., fig. 19A, B.
Holotype
SV1-29, right M2.
Emended diagnosis (modified from Estravís, Reference Estravís2000)
Smallest species of Arcius, approaching the size of A. rougieri. Distolingual basin of M3 somewhat expanded distally, more than in A. rougieri but less than in A. fuscus. Small but patent paraconid on M1, stronger paraconid on M3 than in other species of Arcius. The hypoconulid lobe on M3 is enlarged, as typical of paromomyids, but is the smallest lobe in proportion to the rest of the tooth relative to any other paromomyid, with the exception of Paromomys farrandi.
Horizon and locality.—
Silveirinha (type locality, PE II; see Biostratigraphy), Baixo Mondego, Portugal; Sotteville-sur-Mer (PE II; Hooker, Reference Hooker2015), Normandy, France.
Biostratigraphy
Silveirinha has been considered to belong to MP 7 (PE I) (Estravís, Reference Estravís1992; López-Martínez et al., Reference López-Martínez, Smith, Peláez-Campomanes and Smith2006; Badiola et al., Reference Badiola, Checa, Cuesta, Quer, Hooker and Astibia2009; Smith et al., Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), but also to be very close in age to Sotteville-sur-Mer, probably slightly younger (Smith et al., Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012). Recently, Hooker (Reference Hooker2015) assigned Sotteville-sur-Mer to PE II, so Silveirinha is here considered to be PE II in age following his temporal framework.
Material
SV2-99, right P4; SV3-47, left M3; SV3-125, right M1; SV3-317, right M1; SV3-565, right M3.
Remarks
Antunes (Reference Antunes1981) first reported the presence of paromomyids in Portugal by including Phenacolemur in his faunal list of Silveirinha. It was not until Estravís’s work (1992, 2000) that the Portuguese fossils were included in the genus Arcius and assigned to a new species. Arcius zbyszewskii was suggested by Estravís (Reference Estravís2000) to represent a primitive lineage of European paromomyids due to its mosaic of Arcius characters together with primitive paromomyid characters.
Our interpretation of the Portuguese fossils differs from Estravís (Reference Estravís2000) in that SV1-24 is interpreted as an M1, instead of an M2. We argue that the paraconid on SV1-24 is no smaller than that of the other M1 (SV3-125), which is typical for paromomyids. There is also no difference in the mesiodistal length of the trigonid between the two specimens, while it is a common trait for paromomyids to have a shorter M2 trigonid.
Smith et al. (Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011) described a diminutive upper tooth of a paromomyid from Sotteville-sur-Mer (Normandy, France) that they considered to be a broken M1. The authors acknowledged the similarities of this tooth to the Portuguese fossils but did not ascribe it to any particular species of Arcius. The poor state of preservation of the tooth and the fact that Arcius is characterized by having molariform upper premolars makes it difficult to assign the specimen to a tooth position, but its very weak postprotocrista suggests it is a P4 rather than an M1. Arcius zbyszewskii is the only species of the genus in which the P4 has a very weak postprotocrista instead of it being completely absent. This trait, paired with the similarity in the small size of the specimen from Sotteville-sur-Mer compared to the collection from Silverinha, supports attribution of the Normandy specimen to A. zbyszewskii. Sotteville-sur-Mer and Silveirinha are considered to be of very similar age (Smith et al., Reference Smith, Dupuis, Folie, Quesnel, Storme, Iacumin, Riveline, Missiaen, Ladevèze and Yans2011) and are the oldest sites in Europe that yield paromomyid fossils, which would be consistent with grouping the Arcius from both sites under the same species.
Arcius hookeri new species
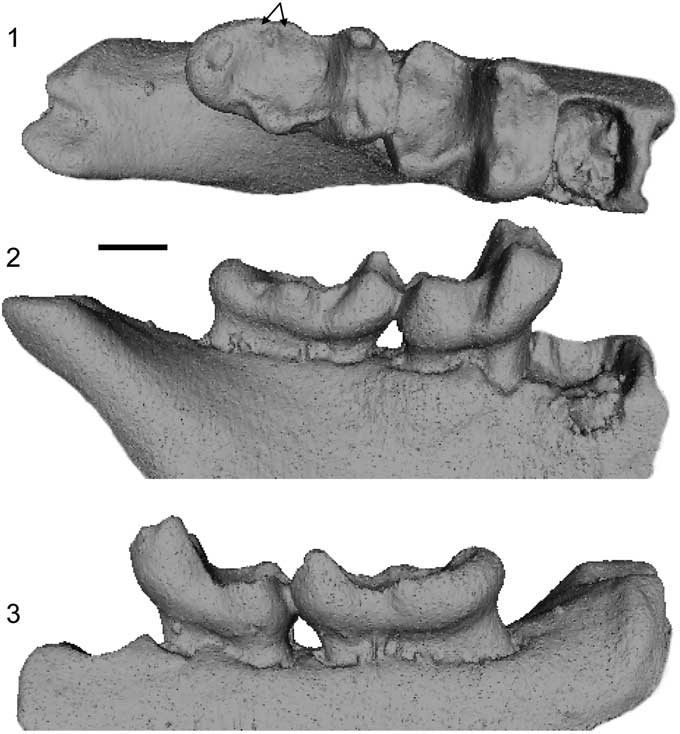
Figure 8 Micro-CT scan images of the holotype of Arcius hookeri n. sp. (BMNH.M 44945) in (1) occlusal, (2) buccal, (3) lingual views. Arrows indicate the presence of a double entoconid. Scale bar=1 mm.
1980 Phenacolemur cf. P. fuscus Reference Hooker and InsoleHooker and Insole, p. 38.
1996 Arcius fuscus Reference HookerHooker, p. 209.
1998 Arcius fuscus Reference HookerHooker, p. 449.
2003 Arcius lapparenti (in part) Reference AumontAumont, appendix A, tables 3, 4.
2010 Arcius lapparenti Reference HookerHooker, p. 48.
Holotype
BMNH.M 44945, right dentary with M2–M3.
Diagnosis
Buccal cingulids present on M2, but weaker than those found in A. fuscus and A. lapparenti and not extended onto the talonid. Trigonid wider, with a greater difference in height between the metaconid and the protoconid, than in other species of Arcius. Mesial inflection of the trigonid not as pronounced as in species of other paromomyid genera, but not as vertical as in A. rougieri and A. ilerdensis n. sp. Differs from all other species of Arcius, except for A. ilerdensis n. sp., in having a taller metaconid than the protoconid on M2 and M3. Differs from A. zbyszewskii in not having a paraconid on M3. Double entoconid present on M3, unlike all other species of Arcius.
Horizon and locality
Blackheath Beds from Abbey Wood (type locality, PE III), England, UK.
Biostratigraphy
Abbey Wood is a reference locality for PE III using mammalian biostratigraphy (Hooker, Reference Hooker1996).
Description
BMNH.M 44945 is the only specimen known of this species. It is composed of a partial dentary with an associated M2 and M3 in place. The complete distal alveolus of M1 is preserved, as well as the distal aspect of the mesial alveolus of M1. The mandibular ramus is partially preserved, and it retains the anterior edge of the masseteric fossa (Hooker, Reference Hooker2010). Arcius hookeri n. sp. shows typical paromomyid characteristics such as reduced paraconids, a protoconid-metaconid notch that is obscured by a fold of enamel, and an expanded hypoconulid on M3 that forms an additional lobe (Silcox and Gunnell, Reference Silcox and Gunnell2008). It shares with other Arcius species features such as taller cusps than in Ignacius, trigonids not as inclined as in other paromomyid genera, and the presence of a simple rounded hypoconulid lobe on M3 (instead of the bilobed hypoconulid lobe seen in other paromomyids; Fig. 2).
Etymology
Named after Dr. Jeremy J. Hooker of the British Museum of Natural History for his unparalleled contributions to the study of the mammalian paleontology of England.
Remarks
The record of paromomyids from England is very limited, with BMNH.M 44945 being the only specimen of a paromomyid to have ever been found in northwestern Europe. Hooker and Insole (Reference Hooker and Insole1980) tentatively reported BMNH.M 44945 as pertaining to Phenacolemur cf. P. fuscus, making this specimen the first known representative of Arcius outside continental Europe. Since then, the specific classification of this specimen has been problematic; it has been suggested to pertain to either A. fuscus (Hooker, Reference Hooker1996, Reference Hooker1998) or A. lapparenti (Aumont, Reference Aumont2003, Hooker, Reference Hooker2010). Hooker (Reference Hooker2010) noted that the English specimen has a small M3 that falls within the size range of A. fuscus for that tooth. He also noted that the M2, although not strictly falling within the size range for that tooth reported by Aumont (Reference Aumont2003, Reference Aumont2004), is closer in size to that of A. lapparenti than to that of A. fuscus. This suggests differences in terms of the relative proportions of these two teeth, but there are no specimens known for either A. lapparenti or A. fuscus in which the M2 and M3 are preserved in series to assess this possibility. Hooker (Reference Hooker2010) also pointed out the presence of two entoconids on the M3, a trait never seen in paromomyids before. When considered alongside the contrasts newly noted here (listed in the diagnosis), it seems as though the best solution is to consider the English specimen to pertain to a new species. See also Table 2 for diagnostic comparisons.
Table 2 Diagnostic characters for Arcius Godinot, Reference Godinot1984 species. Question marks indicate nonobservable characters.

Arcius ilerdensis new species
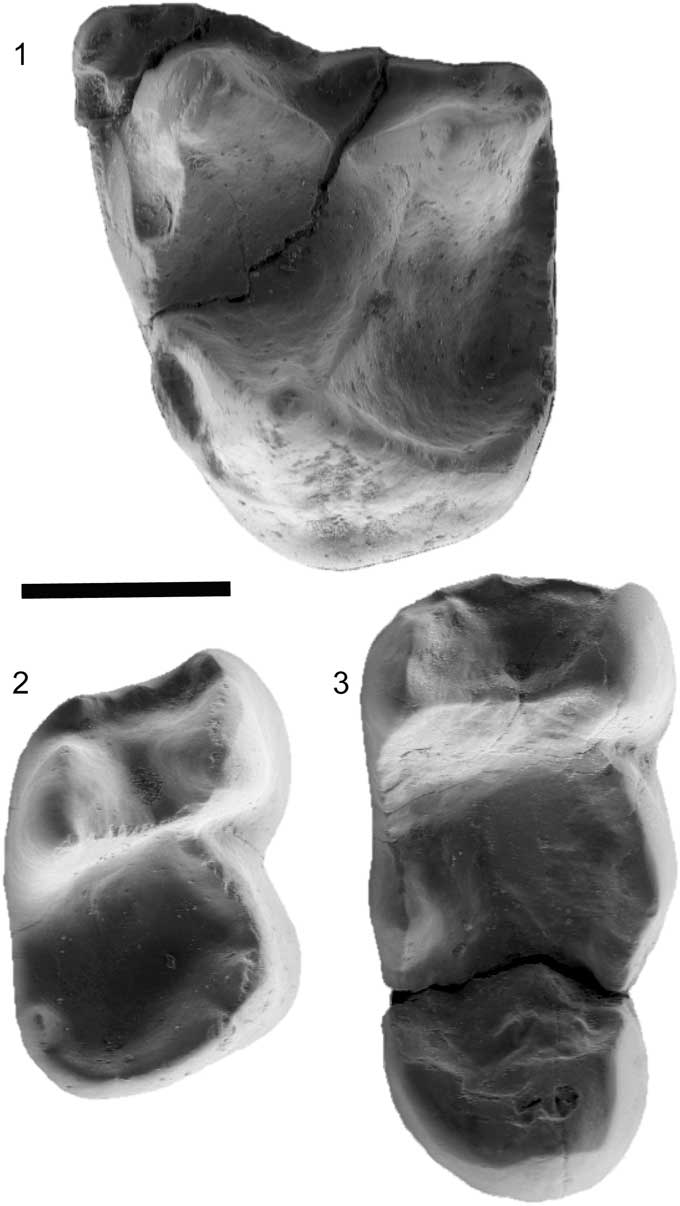
Figure 9 Micro-CT scan images of Arcius ilerdensis n. sp. (1) IPS 57508, left M2, occlusal view; (2) IPS 57510, lower right M2, holotype, occlusal view; (3) IPS 57511, right M3, occlusal view. Scale bar=1 mm..
2012 Arcius sp. Reference Marigó, Minwer-Barakat, Moyà-Solà and López-TorresMarigó et al.
Holotype
IPS 57510, right M2.
Diagnosis
Larger M2 than A. rougieri, larger M2 than A. zbyszewskii, similar areas of M2 to A. hookeri n. sp., and smaller M2 than A. lapparenti. Absence of distal cingulid on M2, unlike A. fuscus, A. lapparenti, and A. rougieri. Differs from all other species of Arcius in lacking a buccal cingulid on M2 and having a short buccal cingulid on the M3 trigonid. Paraconid present on M2, but weakly expressed. Mesial inflection of the trigonid on M2 weaker than any other species of Arcius, with the trigonid being completely vertical. Differs from all other species of Arcius, except A. hookeri n. sp., in having a taller metaconid than the protoconid on M2 and M3. Further differs from A. zbyszewskii in not having a paraconid on M3.
Horizon and locality
Masia de l’Hereuet (MP10?), Lleida, Spain.
Biostratigraphy
Marigó et al. (Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012) described this site as MP8+9, but most genera found in Masia de l’Hereuet usually appear in younger deposits (e.g., Eogliravus, MP10-13 [Escarguel, Reference Escarguel1999]; Lophiodon, MP 10 and later [Cuesta, Reference Cuesta1994], although it is present but rare in MP8+9 [Checa, Reference Checa1997]).
Description
Arcius ilerdensis n. sp. is represented only by four isolated teeth: two left M2 (one of which is damaged; IPS57509), a right M2, a right M3. It shares with other Arcius species features such as taller cusps than in Ignacius, trigonids not as inclined as in other paromomyid genera, and the presence of a simple, rounded hypoconulid lobe on M3 (instead of the bilobed hypoconulid lobe seen in other paromomyids; Fig. 2). The M2 (IPS57508) has a strong postparacrista and premetacrista, with deep basins. A very small paraconule and metaconule are present, typical of paromomyids (Silcox and Gunnell, Reference Silcox and Gunnell2008). The distal margin of the tooth is straight, with a distolingual basin that is fairly expanded, but less so than in Phenacolemur, a typical Arcius trait. The buccal cingulum is broad. The parastylar region is expanded, forming a parastyle, but no metastyle is present. A precingulum is present, but short (Marigó et al., Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012). The M2 (IPS57510) exhibits a trigonid and talonid of similar width and a concave postvallid on the trigonid. The protoconid is lower than the metaconid, and the paraconid is reduced, as is typical of paromomyids (Silcox and Gunnell, Reference Silcox and Gunnell2008). On the talonid, the hypoconulid and the entoconid are poorly distinguishable (Marigó et al., Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012). The M3 (IPS57511) has a more buccally positioned cristid obliqua than on the M2, with more clearly demarcated entoconid and hypoconid (Marigó et al., Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012). It lacks a paraconid and has a very broad hypoconulid lobe, which are typical features of paromomyids (Silcox and Gunnell, Reference Silcox and Gunnell2008).
Etymology
From the Latin Ilerda (ancient name of the city of Lleida), in allusion to its province of origin.
Materials
IPS 57508, left M2; IPS 57511, right M3.
Remarks
The Masia de l’Hereuet specimens were described by Marigó et al. (Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012) as members of the genus Arcius, but those authors did not give them a specific ascription due to the scarcity of the material. Although we agree with Marigó et al. (Reference Marigó, Minwer-Barakat, Moyà-Solà and López-Torres2012) that it would be beneficial for more fossils to be recovered from that site, the few fossils known show enough distinctive traits to be discerned from other species.
Lower molars of paromomyid species that do not belong to the genus Arcius have trigonids that are strongly mesially inflected, while Arcius shows weak levels of inflection. However, Arcius ilerdensis has a uniquely vertical trigonid, even relative to other members of the genus. It is also uncommon among Arcius that the metaconid is clearly taller than the protoconid in lower molars, a feature also seen in A. hookeri n. sp. The lack of a distal cingulid on the buccal half of the distal aspect of the M1-2 also contrasts with the more derived members of the genus (A. fuscus, A. lapparenti, and A. rougieri), suggesting that A. ilerdensis still retains somewhat primitive characters in the Arcius lineage.
Phylogenetic relationships of Arcius
To assess the phylogenetic relationships among the different species of Arcius, and the position of the genus among Paromomyidae, we conducted a cladistic analysis. A character list was created to assess specifically the relationships of Arcius based on character diagnoses from Matthew (Reference Matthew1915), Gidley (Reference Gidley1923), Russell et al. (Reference Russell, Louis and Savage1967), Rose and Bown (Reference Rose and Bown1982), Godinot (Reference Godinot1984), Robinson and Ivy (Reference Robinson and Ivy1994), Estravís (Reference Estravís2000), Bloch et al. (Reference Bloch, Boyer, Gingerich and Gunnell2002), Silcox et al. (Reference Silcox, Rose and Bown2008), and Fox et al. (Reference Fox, Scott and Rankin2010) (Table 3). Some of the characters that relate to higher-level relationships among plesiadapiforms were taken from Silcox’s (Reference Silcox2001) matrix for plesiadapiforms. The primitive purgatoriid Purgatorius coracis Fox and Scott, Reference Fox and Scott2011 was chosen as the outgroup for Paromomyidae. The analysis also includes the oldest and most primitive members of every paromomyid genus: Paromomys farrandi, Edworthia lerbekmoi Fox et al., Reference Fox, Scott and Rankin2010, Phenacolemur archus Secord, Reference Secord2008, Acidomomys hebeticus, and Ignacius fremontensis Gazin, Reference Gazin1971. The genus Elwynella, which includes only the species Elwynella oreas Rose and Bown, Reference Rose and Bown1982, was excluded from this analysis because it is the most recent genus of paromomyid to appear in the fossil record, and although it exhibits the primitive character of retention of a P3, it shares many derived molar characters found in the late Wasatchian paromomyid Phenacolemur jepseni, which suggests that it may be nested within a higher-level grouping of paromomyids. A total of 53 dental characters were scored for 13 taxa (Table 3; matrix available on publication from the Dryad Digital Repository; see also Supplemental Data 2).
Table 3 Descriptions of dental characters.

The parsimony analyses were performed using TNT (Goloboff et al., Reference Goloboff, Farris and Nixon2008) with all characters equally weighted. Four of the 53 characters (1, 8, 13, and 39) were ordered, and the rest were left unordered. A heuristic search was implemented with 1,000 repetitions, with 1,000 trees saved per replication. Five cladistics analyses were run: (1) a basic analysis that excluded the poorly sampled taxa from Abbey Wood (UK), Sotteville-sur-Mer (France), and Masia de l’Hereuet (Spain); (2) an analysis that included the Arcius from Abbey Wood; (3) an analysis that included the Arcius from Sotteville-sur-Mer; (4) an analysis that included the Arcius from Masia de l’Hereuet; and (5) an analysis that included all European paromomyid taxa. Analysis 1 is meant to give a general understanding of the relationships of Arcius based on the well-preserved species, whereas analyses 2, 3, and 4 are meant to accurately place the poorly sampled taxa. Analysis 5 is also meant to give a general understanding of the relationships of the genus but including all taxa (both well and poorly sampled). Analyses 1, 2, and 3 yielded only one unequivocal tree each (Fig. 10.1–10.3). Analysis 4 yielded 6 equally parsimonious trees. A strict consensus tree was generated in TNT from these trees (Fig. 10.4). Analysis 5 also yielded 6 equally parsimonious trees, and a strict consensus tree was generated in TNT (Fig. 11).
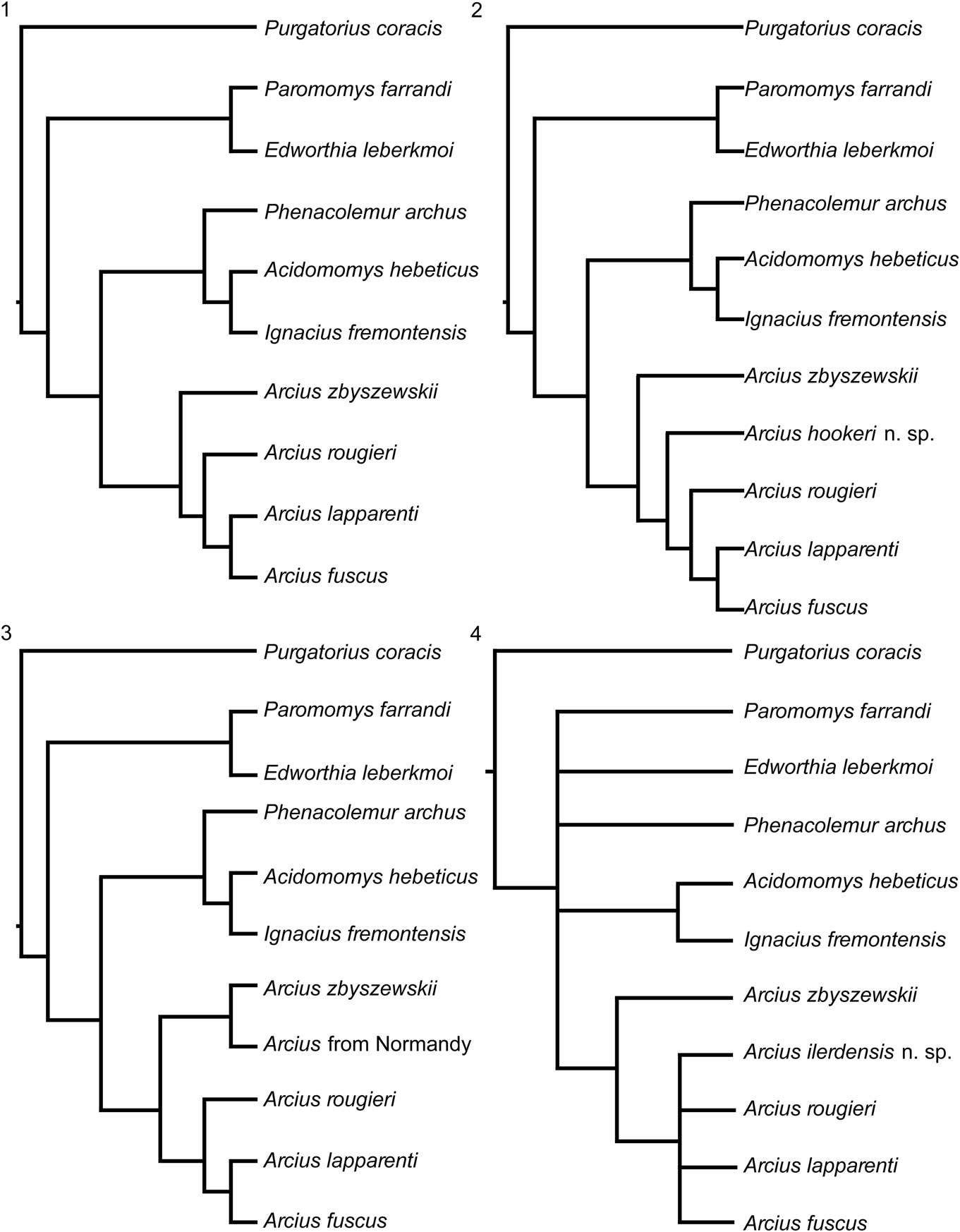
Figure 10 Hypotheses of phylogenetic relationship among selected genera of North American and European paromomyids based on cladistic analysis of 53 dental characters (Table 3, Supplemental Data 1, Figure S1). Cladistic analysis yielded a single most parsimonious cladogram for (1), (2), and (3). All cladograms rooted with Puercan Purgatorius coracis Fox and Scott, Reference Fox and Scott2011. All characters were unordered, except for characters 1, 8, 13, and 39. Cladogram (4) was generated by strict consensus of six trees (Figure S1).
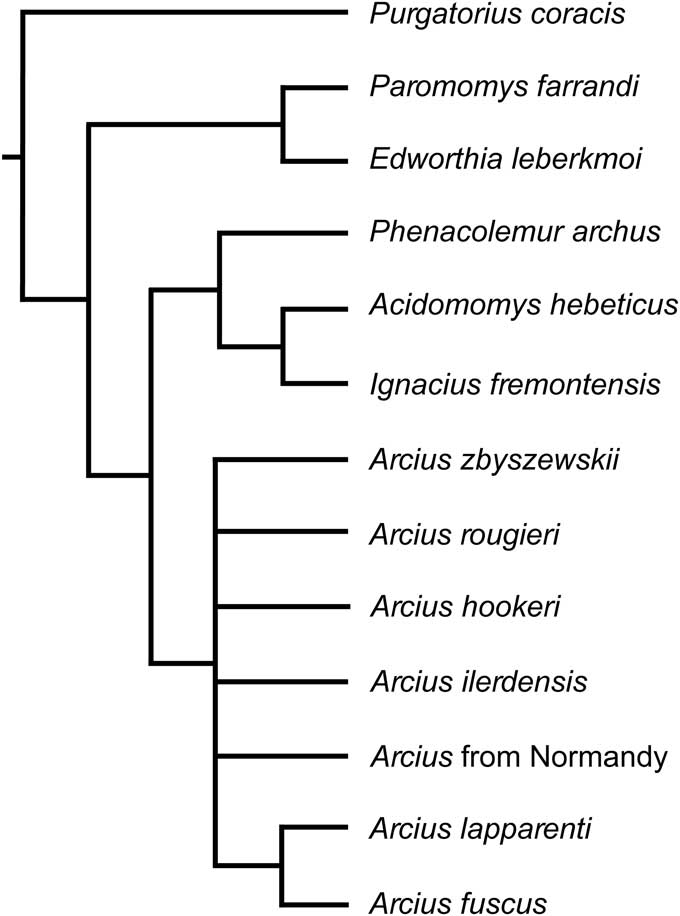
Figure 11 Cladogram generated by strict consensus of six trees. All European paromomyid taxa were included in this tree.
Results of the phylogenetic analysis
In the tree from Analysis 1, Arcius is found to be monophyletic, with A. fuscus and A. lapparenti forming a clade. Arcius rougieri appears as the sister group to the A. fuscus-A. lapparenti clade. Arcius zbyszewskii is placed as the most basal European paromomyid, which agrees with the inference by Estravís (Reference Estravís2000) that A. zbyszewskii is a primitive lineage of European paromomyids, opening up the possibility of it being an ancestral species to all European paromomyids. In terms of the rest of paromomyid relationships, all of the trees from this paper agree with Bloch et al. (Reference Bloch, Boyer, Gingerich and Gunnell2002) and Aumont (Reference Aumont2003) that Phenacolemur and Ignacius are closely related and that Paromomys is a very primitive member of the family. However, the position of Acidomomys seems more contentious. Whereas Acidomomys makes a clade with Ignacius in our trees, Bloch et al. (Reference Bloch, Boyer, Gingerich and Gunnell2002) found Acidomomys to be the sister taxon of the clade Ignacius+Phenacolemur, and Aumont’s (Reference Aumont2003) analysis resulted in Acidomomys being the sister taxon to Arcius. Aumont (Reference Aumont2003) stated that the two characters that support the Acidomomys-Arcius clade are “l’absence de la p3” (absence of P3) and “la presence d’une P3 uniradiculée” (presence of a single-rooted P3) (Aumont, Reference Aumont2003, p. 364). However, Acidomomys hebeticus does have a P3 (Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002), and therefore that trait seems to be incorrectly coded in Aumont’s (Reference Aumont2003) matrix. In addition, the other character (presence of a single-rooted P3) seems to be misreported because P3 is coded in the matrix as being double rooted for both Acidomomys hebeticus and Arcius rougieri and not observable (i.e., question mark) in the rest of Arcius species. While it is clear that Acidomomys hebeticus had a double-rooted P3 (Bloch et al., Reference Bloch, Boyer, Gingerich and Gunnell2002), as coded, the number of roots for P3 in Arcius is difficult to assess. Only a small piece of the distobuccal aspect of the P3 of A. rougieri is preserved (Godinot, Reference Godinot1984), and the alveoli for that tooth are not preserved. It is, therefore, impossible to determine how many roots would have been present. In light of these considerations, we would argue that there is no support for an Acidomomys-Arcius clade, which Aumont (Reference Aumont2003) tentatively named as the tribe Arciini (incorrectly reported by Aumont [Reference Aumont2003, p. 364] as ‘Arciusini’). In addition, the fact that Acidomomys appears as the sister taxon to Arcius might explain why Aumont’s phylogeny differs from those reported here in that Arcius rougieri represents the most primitive lineage of European paromomyids in her analysis, instead of Arcius zbyszewskii as found here.
When Arcius hookeri n. sp. is included in the analysis (Analysis 2; Fig. 10.2), it also yields one tree unequivocally. Arcius hookeri is placed as the sister taxon of a clade that includes A. fuscus, A. lapparenti, and A. rougieri. This result supports the inference that the Arcius specimen from England pertains to a separate lineage and therefore should be named as a distinct species. The clade that includes A. hookeri, A. fuscus, A. lapparenti, and A. rougieri is supported by the loss of the paraconid on the M3, which is clearly present in the more primitive A. zbyszewskii.
The addition of the paromomyid specimen from Sotteville-sur-Mer (Normandy, France) also results in a single most parsimonious tree (Analysis 3; Fig. 10.3). In this case, the Normandy specimen (MAM 6 STV 2008) forms a clade with Arcius zbyszewskii. This result is consistent with the morphological resemblance between the fossils from Portugal and the one from Normandy and supports the inference that all these specimens belong to the same species, A. zbyszewskii.
When including Arcius ilerdensis n. sp. in the analysis (Analysis 4), the clade A. fuscus-A. lapparenti-A. rougieri, present in the results from Analyses 1–3, collapses (Fig. 10.4). In three of the six trees recovered from this analysis (Fig. S1.1, S1.2, S1.6), A. ilerdensis falls out as being most closely related to A. rougieri. Arcius ilerdensis also appears in two other trees (Fig. S1.4, S1.5) as the sister group to the clade A. fuscus-A. lapparenti-A. rougieri, and as most closely related to A. lapparenti in another tree (Fig. S1.3).
When all species are included in the analysis (Analysis 5; Fig. 11), most Arcius lineages collapse, likely because of the large amounts of missing data for several taxa. However, it is important to note that the clade including A. fuscus and A. lapparenti still holds in this cladogram.
In terms of the other genera of paromomyids, our results show that the sister group of Arcius is not Acidomomys, as reported in Aumont (Reference Aumont2003), but the clade that includes Phenacolemur, Ignacius, and Acidomomys. The genus Acidomomys is most closely related to Ignacius. Paromomys and Edworthia appear as members of the most primitive lineage of paromomyids. These results also support the idea from Gunnell (Reference Gunnell1989) that the family Paromomyidae could be subdivided into two subfamilies: the Paromomyinae and the Phenacolemurinae. According to Gunnell (Reference Gunnell1989), based on the genera described at that time, paromomyines would only be composed of the genus Paromomys, and phenacolemurines would include Phenacolemur, Ignacius, and Elwynella. Here, Paromomyinae would be extended to include Paromomys and Edworthia, and Phenacolemurinae would include Phenacolemur, Ignacius, Arcius, and possibly Elwynella. However, to assess this question more fully, additional North American paromomyid species should be included in the analysis of the family.
Biogeographic implications
The results of the phylogenetic analyses, which consistently support the monophyly of Arcius, would suggest that there was only one dispersal event of paromomyids between North America and Europe. This dispersal would have been possible through land bridges between North America and Europe. The volcanic activity and regional uplift in eastern Greenland closed the Greenland Strait (or Denmark Strait) between Europe and Greenland, making it much narrower during the Paleocene and Eocene (Knox, Reference Knox1998). In addition, the English Channel was bridged, allowing dispersal between the British Isles and continental Europe, and in particular with France (King, Reference King2006).
One of the oldest occurrences of Arcius comes from the site of Sotteville-sur-Mer in Normandy, France. That region of Normandy could have been the entry point to continental Europe from the Greenland land bridge (Fig. 12), which would be consistent with finding the oldest representative of the genus in that area. However, Arcius zbyszewskii is also found at Silveirinha, which is inferred to be of nearly equivalent age to Sotteville-sur-Mer (i.e., PE II according to Hooker, Reference Hooker2015) and is quite distant in space. Iberia was isolated from continental Europe by the Carcassonne Strait already by the earliest Eocene (Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012), and the intercontinental dispersal of paromomyids is argued to have occurred during the PETM (Hooker, Reference Hooker2015), making the dispersal of A. zbyszewskii into southwestern Europe rather difficult. However, other sources (Plaziat, Reference Plaziat1981; Badiola et al., Reference Badiola, Checa, Cuesta, Quer, Hooker and Astibia2009) illustrate a land connection between Iberia and continental Europe during the early Eocene, suggesting that faunal dispersal from continental Europe into Iberia might have been possible by the earliest Eocene. In any case, the broad biogeographic range of Arcius zbyszewskii across western Europe suggests that this species might have undergone rapid dispersal across the continent in a short period of time. Even if the fossils from Silveirinha and Normandy did not belong to the same species, it would be one of the first cases of finding similar species in northern and southern Europe, which goes against the strong trend of north–south regionalism in this continent (Marandat, Reference Marandat1997).
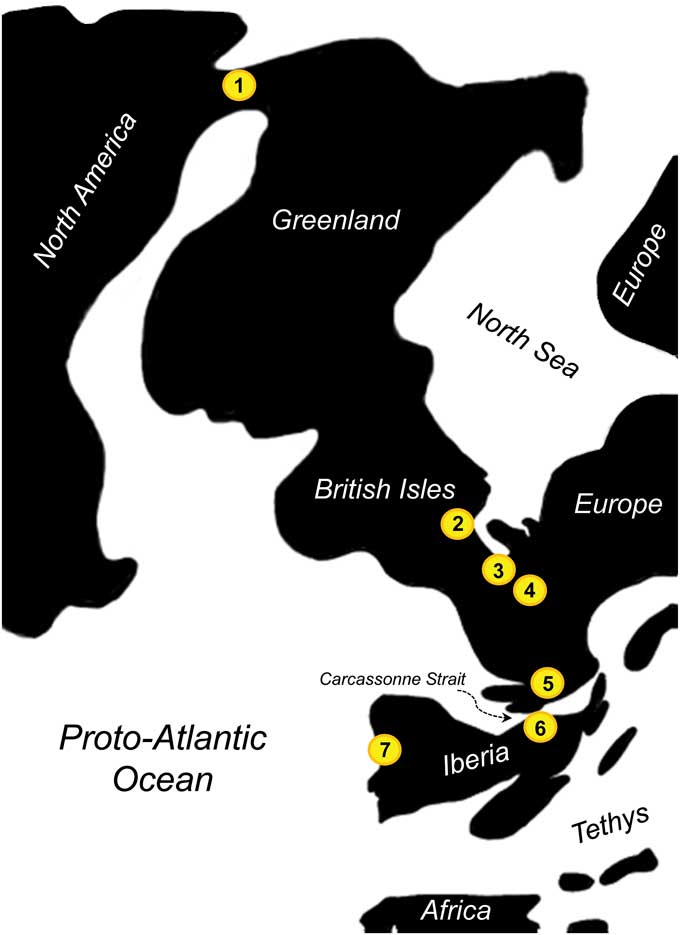
Figure 12 Paleogeography of Western Europe and Greenland during the earliest Eocene and location of some selected paromomyid-bearing locations (modified from Eberle and Greenwood, Reference Eberle and Greenwood2012, fig. 3; Marandat et al., Reference Marandat, Adnet, Marivaux, Martinez, Vianey-Liaud and Tabuce2012, fig. 1; and Hooker, Reference Hooker2015, fig. 8). (1) Eureka Sound Group, Canada; (2) Abbey Wood, UK; (3) Sotteville-sur-mer, France; (4) Paris Basin, France; (5) Palette, France; (6) Masia de l’Hereuet, Spain; (7) Silveirinha, Portugal.
In terms of the origins of Arcius, the phylogenetic results unequivocally support the existence of a long ghost lineage. The oldest member of the Phenacolemur-Ignacius-Acidomomys clade is Ignacius cf. I. fremontensis from Torrejonian 2 (To2; Rose, Reference Rose1981; Silcox and Williamson, Reference Silcox and Williamson2012). Therefore, the Arcius lineage is inferred to have branched off prior to To2. The absence of Arcius-like species in the well-sampled western North American record during the Torrejonian, Tiffanian, and Clarkforkian suggests that the evolution of this lineage might have happened in more northern latitudes and closer to the land bridges that would later connect North America with Europe. In fact, undescribed Ignacius-like paromomyids have been reported from the Eocene of Ellesmere Island, in the Canadian Arctic (West and Dawson, Reference West and Dawson1977; Eberle and Greenwood, Reference Eberle and Greenwood2012), hinting at the potential of this region, and of other areas in the Greenland land bridge (e.g., the east coast of Greenland; Larsen et al., Reference Larsen, Bjerager, Nedkvitne, Olaussen and Preuss2001, Reference Larsen, Piasecki and Stemmerik2002), to rewrite our understanding of paromomyid evolution and biogeography.
Conclusions
A new revision of the paromomyids from Europe (after Aumont, Reference Aumont2004) prompts the rediagnosis of the genus Arcius based on a significant collection of previously unpublished material, redefining Arcius fuscus and Arcius lapparenti after identifying the mix of specimens in the original taxonomic study by Russell et al. (Reference Russell, Louis and Savage1967), and the description of two new species: Arcius hookeri n. sp. and Arcius ilerdensis n. sp.
Our phylogenetic analysis shows that Arcius is monophyletic. This is consistent with previous work done by Aumont (Reference Aumont2003) but differs from her results in the precise relationships among species of Arcius. Our tree suggests that Arcius zbyszewskii is the most basal species of Arcius, in line with Estravís’s (Reference Estravís2000) inferences. Our results further differ from Aumont’s (Reference Aumont2003) in finding the clade Phenacolemur-Ignacius-Acidomomys as the sister clade to the European paromomyids, instead of just Acidomomys. When the more fragmentary taxa were added to the phylogenetic analysis, the results are consistent with the inference that the Normandy specimen belongs to Arcius zbyszewskii and that Arcius hookeri belongs to an independent lineage. The phylogenetic relationships of Arcius ilerdensis are less clear, but it appears basal to the clade of French species of Arcius. Finally, the monophyly of Arcius is consistent with a single dispersal for the family Paromomyidae from North America to Europe.
Acknowledgments
We thank M. Godinot and C. Argot (Muséum Nationale d’Histoire Naturelle), R. Rocha (Universidade Nova de Lisboa), J.J. Hooker (British Museum of Natural History), T. Smith (Royal Belgian Museum of Natural Sciences), J. Galkin, A. Davison, and R. O’Leary (American Museum of Natural History), P.A. Holroyd and W.A. Clemens (University of California Museum of Paleontology), R.C. Fox and M.W. Caldwell (University of Alberta), P.D. Gingerich, W.J. Sanders, and A. Rountrey (University of Michigan), E.J. Sargis, D. Brinkman, M. Fox, and C.A. Norris (Yale Peabody Museum), D.J. Bohaska and N. Pyenson (National Museum of Natural History – Smithsonian Institution), K.D. Rose (Johns Hopkins University), and J.I. Bloch (Florida Museum of Natural History) for access to specimens and/or casts. Thanks to T.B. Viola for fruitful discussions and to K.C. Beard, D.R. Begun, W.A. Clemens, and M.A. Schillaci for helpful comments. Viktoria Bodo prepared Figure 6; it is included with her permission. We are grateful to R. Minwer-Barakat and J. Marigó for permission to use original figures. We thank A. Aumont, M. Godinot, and R. Rocha for access to unpublished material. Thanks to E. Charles, R. Bhagat, and D. Lin for making relevant casts. We are very grateful to J.T. Gladman and C. Crawford for their assistance in CT-scanning the specimens of Arcius and to D.M. Boyer for access to resources. Thanks to K.R. Selig for further assistance during scanning. We are grateful to M.C. Torres Jové for access to data. We also thank M. Godinot, J.I. Bloch, and H.-D. Sues, whose comments significantly improved this paper. This research was supported by the Doris O. and Samuel P. Welles Research Fund, an American Museum of Natural History Collections Study Grant, and University of Toronto Department of Anthropology Research Travelling Grants to SLT, and an NSERC Discovery Grant to MTS.
Accessibility of supplemental data
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.873c4















