


Introduction
This paper has two main objectives: (1) to advance a paradigm change on the subject of life on Mars; and (2) to add substance to the topic of this conference, panspermia.
Panspermia is believed to have been first written about by the early Greek philosopher Anaxagoras, and was first scientifically addressed some 150 years ago by Louis Pasteur in his attempt to culture microorganisms from a piece of the Orgueil meteorite. It is my belief that a simple modification of the Labelled Release (LR) experiment may provide the first live evidence of panspermia.
Life is the most complex phenomenon known. Thus, the detection of any biochemical or biosignature is unlikely to be accepted as proof of extant life. For that reason, the LR, as sent to Mars aboard the Viking Mission in 1976 (Levin & Straat Reference Levin and Straat1979), was designed to detect active metabolism, which the author believes would be proof of extant life.
The Viking LR is depicted in Fig. 1. A 0.5 cc sample of Martian soil is placed into a 4.0 cc container under simulated Martian atmosphere (10 mb CO2), with helium added to effect a total atmospheric pressure of 85 mb. The soil is maintained at 10°C and inoculated with 0.15 ml of an aqueous solution of the 14C-labelled compounds shown in Table 1. Chromatographic movement of the liquid through the soil produces wet to barely moist conditions. Any microorganisms present that could metabolize any of the nutrients thus supplied would be expected to produce an end product of one or more labelled gases.

Fig. 1. Schematic of the Viking LR experiment.
Table 1. Labelled release nutrients
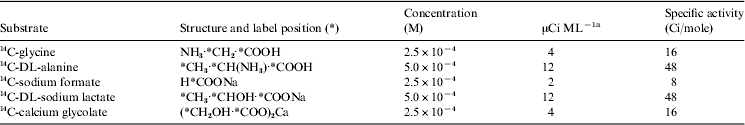
a Total=34 μCi, which yields 6.8×107 dpm ml−1.
The compounds chosen as nutrients were Miller–Urey compounds, because they are thought to have formed on the early Earth, to have participated in the origin of life, and to have been passed on to today’s life forms. In the many hundreds of tests performed in Earth laboratory and field tests it was demonstrated that the nutrient solution always produced positive responses from live cultures and from microbe-populated soils. The gas evolved quickly saturated the small amount of liquid, readily making the phase change from liquid to gas. The gas rose through the connecting tube that precluded aerosols or dust from making the journey into the headspace of the detector chamber. The amount of gas accumulating was measured by its radioactivity as counted by beta detectors. In the event of a positive response, a duplicate sample of the same soil, the control, was heated to 160°C for 3 hours to inactivate putative microorganisms. (This was the control procedure specified by NASA for all three Viking life detection tests.) The sample was allowed to cool and then tested. A negative control confirmed that the initial response was from living organisms. Were the control response positive, it would indicate that the test response had been caused by a chemical or physical agent rather than by living organisms.
The LR was honed for 20 years on thousands of tests on microbial species in pure cultures, mixed cultures, soils from exotic places, and in field tests in extreme environments, such as seen in Figs 2–4. Simple, very rapid, and producing essentially no noise because of the phase separation from the mother liquor, the LR proved sensitive to as few as ∼20 cells per sample (Cameron & Benoit Reference Cameron and Benoit1970). The LR earned a strong pedigree by going through ten years of development as ‘Gulliver’, as the ‘sticky string’ experiment was called prior to formation of the Viking Mission. Selection of Gulliver for Viking was made by four NASA-appointed committees. Renamed the ‘Labelled Release Experiment’, it was subjected to ten more years of development. This involved quarterly and annual reports, annual submittals for continued funding, frequent inspections by NASA and instrument manufacturer officials, design reviews, critical design reviews, and visits from the infamous ‘Tiger Teams’ that investigated any perceived problems.

Fig. 2. LR test at 12 000 ft (above timberline) on White Mountain, CA.

Fig. 3. LR test on Death valley sand dune. Despite only 0.9% moisture in the top 2 mm of sand, a strong positive response was immediately obtained.

Fig. 4. LR ‘sticky string’ test on Salton desert flats.
The LR on Mars
30 July 2006 marked the 30th Anniversary of the LR’s first experiment on Mars. The results of that first experiment were startling. Immediately upon injection of the labelled nutrients, gas began evolving from the soil. The experiment ran for 7 sols (Martian days, 26.4 hours each), with gas rapidly rising for 3 sols, and then approaching a plateau for the remaining time. A fresh culture chamber was then rotated into place, and the instrument was purged of gas. The control was then run. It produced a nil response. At that point, the LR had satisfied the pre-mission criteria for life.
Going beyond its Mission criteria, the LR went on to buttress its remarkable finding by obtaining strong additional evidence. Figure 5 shows the results of the first cycles of all tests at the Viking 1 landing site. The corresponding data for Viking 2 are shown in Fig. 6.

Fig. 5. Comparison of radioactivity evolved following the first injection of radioactive nutrient to each analysis cycle of VL-1. A fresh sample was used for the active sequences of cycles 1 and 3 whereas the sample used for active cycle 4 was stored for approximately 141 sols at 10–26°C prior to use. For cycle 2, a stored portion of the same sample used for cycle 1 was heated for 3 hours at 160°C prior to nutrient injection. All data have been corrected for background counts observed prior to nutrient injection.

Fig. 6. Comparison of radioactivity evolved following the first injection of radioactive nutrient to each analysis cycle of VL-2. A fresh sample was used for each cycle except cycle 5 which used a sample stored for approximately 84 sols at 7°C prior to injection. The sample used in cycle 3 was obtained from under a rock. Cycles 1, 3, and 5 were active sequences, whereas cycles 2 and 4 were control sequences in which the samples were heated for 3 hours at approximately 51.5°C and 46°C, respectively, prior to nutrient injection. Sample volumes were 0.5 cc except for cycle 5 which contained 2.2 cc. All data have been corrected for background counts observed prior to injection.
At both sites all of the test and control data were consistent with the criteria for life. An immediately proposed theory that the positive responses were caused by activation of the soil by ultraviolet light was dispensed with by VL2 Cycle 3, seen in Fig. 6, the results of a sample taken from under a rock moved just after dawn, which, nonetheless, showed strong activity.
The LR controls established that the ‘active agent’ detected in the Martian soil was destroyed at 160°C, was greatly impaired at 46°C, was essentially destroyed at 51°C, and was fully depleted after holding in the dark at approximately 10°C inside the sample distribution box for 3 and 4 months at the respective sites. All results were supportive or consistent with the detection of a biological agent. Still, the consensus remained: ‘chemistry not biology’.
Despite much evidence to the contrary over the past three decades, a ‘Standard Model’ has emerged for life on Mars:
The surface of Mars is inimical to extant life because of the absence of liquid water, the intense UV flux and a ubiquitous layer of highly oxidizing chemical(s). The absence of organic matter in the surface material is proof of the oxidizing layer and/or the effect of the UV flux, and of the absence of life. Life may have existed on the surface in the geological past when conditions were more hospitable. Extant life may inhabit underground oases where there is liquid water and environmental conditions provide a favorable habitat.
This paper will show that this model, along with its corollaries, is not supported in fact.
Each of the obstacles to biology posed by the Standard Model and its corollaries has been rebutted. All such obstacles publicly proposed are shown in Table 2. None has been sustained. The principal ones are discussed below.
Table 2. Non-biological theories put forth to refute Mars LR evidence for life

(a) Failure to detect organics
The Viking GCMS Experimenter disclaimed life detection ability for that instrument, pointing out that 109 cells in that instrument’s 60 mg sample were required to supply enough organic matter for detection (Biemann et al. Reference Biemann1977). (However, he contended that, were there any living microorganisms present in the soil, the numerous bodies of the dead ones would supply the needed amount of organic matter for detection.)
The Viking Pyrolytic Release (PR) experiment sought to detect photosynthetic life by detecting the fixation of C-labelled Martian atmosphere (CO and CO2) by organisms in the soil when placed under simulated sunlight. In terrestrial tests, the Experimenters reported the formation of labelled organic matter on sterile soil, stating ‘The amount of product formed could be considerable over geologic time’ (Hubbard et al. Reference Hubbard, Hardy and Horowitz1971). An optical filter was placed in the Viking PR instrument in an attempt to prevent false positives by screening out the UV flux below 300 nm which caused the reaction. However, this did not eliminate formation of organics in the PR on Mars (but at levels far below that required as evidence of life). The formation of organics occurred even in the controls, as demonstrated earlier (Hubbard et al. Reference Hubbard, Hardy, Voecks and Golub1973). Thus, organics are strongly indicated as being formed on the surface of Mars. The fact that the GCMS failed to find them indicates a problem with the Viking GCMS.
Since Viking, various other problems, mechanical and poor sensitivity, have been cited for the Viking GCMS (Levin et al. Reference Levin, Kuznetz and Lafleur2000; Glavin et al. Reference Glavin, Schubert, Botta, Kminek and Bada2001) and, recently, it has been shown that the GCMS technique failed to detect microorganisms in the hyperarid Atacama Desert which are thought to be a good analogue for Mars (Navarro-Gonzalez et al. Reference Navarro-Gonzalez2006).
(b) Strong oxidant
As already seen in Table 2, many theories have been advanced to propose hydrogen peroxide, other strong oxidants, and various chemical and physical agents on the surface of Mars as the source of the LR responses. However, there is much evidence discounting the oxidant theories (Levin Reference Levin2006). Viking, itself, provided the first refutation. The Viking Magnetic Properties experiment found a high percentage of the iron in the soil to be magnetic, thereby demonstrating that it was not fully oxidized. This is shown in Fig. 7.

Fig. 7. Reference test chart magnet image for (a) VL-1 on sol 31 and (b) for VL-2 on sol 422 mm to 4 mm of surface material were picked up by each magnet:
If there is a lot of material adhering to the magnet, it would certainly say that whatever the surface processes are on Mars, they are not innately highly oxidizing (Hargraves, Viking Magnetic Properties Experimenter (1975)).
The formation and preservation of organics by the PR mentioned above is also evidence against an oxidant. The fact that the organic matter formed in the PR instrument on Mars was not destroyed by the putative oxidant in the soil strongly argues against the presence of an oxidant.
The Viking GCMS itself produced evidence against an oxidant. The GCMS Experimenter stated the instrument was sensitive to several parts per billion of organic matter (produced by the pyrolysis of the sample, which was apparently less than the organic yield from the 109 bacterial cells he stated was required for the detection of organics (Biemann et al. Reference Biemann1977)). The GCMS’s detection of ‘a few tens of parts per billion’ of methyl chloride, a residual from stringent chemical cleaning of the instrument on Earth, was pointed to as evidence of the instrument’s sensitivity. However, were there even a few parts per billion of a strong oxidant in the soil sample, the methyl chloride would have been oxidized to methylene chloride vapour in amounts that would have been detected as such by the GCMS.
Two Earth-based observations found no peroxide in the entire column of the Martian atmosphere (Krasnopolsky et al. Reference Krasnopolsky, Bjoraker, Mumma and Jennings1997; Krasnopolsky & Feldman Reference Krasnopolsky and Feldman2001). Each established an upper limit of the oxidant below that capable of producing an LR response. A recent announcement reports hydrogen peroxide in the Martian atmosphere at a mixing ratio of 10−8, claiming anew that this accounts for the LR. However, this miniscule amount could not produce the LR active responses, or survive the UV flux, or contact with surface materials. Furthermore, it is less than the amount in the Earth’s atmosphere, under which life thrives.
The Rover Opportunity also confirmed that the surface of Mars is not highly oxidizing. Its analysis of the Martian soil, as seen in Fig. 8, shows that the majority of iron in the soil is not fully oxidized to the Fe2O3 state, but is in the ferrous state.

Fig. 8. Evidence of reduced surface material on Mars as analyzed by Rover opportunity.
Finally, even granting the blatantly difficult case for an oxidant coating Mars, all claims that this explains the LR result fail to address the fact that no such oxidants have exhibited the thermal sensitivity of the LR controls. None of the oxidant theories, or any of the other abiological theories, fit the data.
(c) ‘Too much too soon!’
Soon after the first positive LR response, it was stated that the response was ‘Too much too soon’ to have been produced by Martian microorganisms. Figure 9 shows the Martian VL1 Cycle 1 LR response in relation to responses from a variety of terrestrial field tests. The response on Mars is within the terrestrial range, and is at the lower end where it might be expected because of the low Martian temperature. The Mars response amplitude and kinetics are shown to be quite similar to those of Antarctic soils and endolithic microorganisms extracted from Antarctic rocks.

Fig. 9. Comparison of terrestrial and Mars LR active responses.
(d) Second injection
After the 7-sol LR experiments, second injections of nutrient were made onto soils that had given positive results. Thereupon, rapid re-absorption of approximately 20% of the headspace gas occurred. Although second injections were not part of pre-mission LR criteria, the fact that a new outpouring of labelled gas did not occur led to further doubt of the biological interpretation of the Mars LR. However, a search of the LR terrestrial test data found an Antarctic soil that showed a similar response to a second injection of nutrient. The Mars and Antarctic soil second injection responses are shown in Figs 10(a) and 10(b). The indication is that the microorganisms in these samples from both planets died during the latter part of first injection cycles. When the second injections wet the apparently alkaline soils, gas re-absorption occurred.

Fig. 10. Effect of second injection on (a) Antarctic soil and (b) Mars soil. In (a) the soil contained less than 10 cells by classical microbiological methods (Cameron et al. Reference Cameron, King and David1970).
The fact that active soils stored at 10°C for approximately 3 and 4 months (Figs 5 and 6) produced no gas evolution upon injection of the substrates is consistent with the ‘limiting factor’ being the death of soil microorganisms. The unlikely alternative is that the active agent in the soil was a chemical that became unstable when held in the dark at a temperature within the Mars ambient range.
(e) ‘No liquid water, no life’
This is now the primary surviving obstacle raised against a biological interpretation of the Mars LR experiment. However, there is much evidence for liquid water on the Martian surface, beginning with Viking. The Lander footpads had thermistors in them to determine the surface temperature. As the morning progressed on Mars, the surface temperature rose. Lander 2 recorded that the rise halted for several minutes at 273°K, the temperature at which ice melts after absorbing energy to supply the latent heat of fusion. Figure 11, an image taken by Viking Lander 2, shows that frost or snow was present.

Fig. 11. Heavy frost or snow at VL-2 lander site (Viking Lander image 211093).
Pathfinder measured the temperature of the Martian air. At 0.5 m above the ground, the daytime temperature exceeded 20°C. Figure 12 shows water exists close (perhaps within several centimetres) to much of the surface of Mars, including both Viking landing sites. Presumably, most of the time the water is in the form of ice. However, the daily rise in the temperature would produce liquid water. Furthermore, theoretical modelling and experiments have demonstrated that ice liquefies under Martian conditions (Levin & Weatherwax Reference Levin and Weatherwax2003). In developing the Astronaut space suits, the Hamilton Standard Company found that ice on the backpacks of Astronauts in a re-compression chamber melted when the chamber reached 0°C under as little as 4 mb total pressure. Figure 13 shows frost on a rock placed under Martian conditions. The frost melted into liquid water as the temperature was raised to 0°C. Figure 14 is an image taken by the Rover Opportunity, showing what appears to be mud. The same image is shown in Fig. 15 with its red, green, and blue channels combined give a colour image. A NASA image evidencing current liquid water on Mars is seen in Fig. 16.

Fig. 12. Most recent Odyssey findings.

Fig. 13. Mars in a jar: liquid water from frost on rock under Martian conditions. Courtesy of D. Gan and L. Kuznetz, University of California, Berkley.

Fig. 14. Mud puddles in Mars.

Fig. 15. Colour image of mud on Mars?

Fig. 16. Mars global surveyor image. NASA scientists propose that liquid water may currently seep from the walls of this unnamed crater in the planet’s southern hemisphere. Photo courtesy of NASA.
In response to this mounting evidence of liquid water on Mars, erstwhile proponents of a dry Mars now propose an ‘Oases Theory’, saying that there might be liquid water on Mars but only in underground oases. They then concede that life might exist in such niches, but maintain that the surface of Mars is sterile. However, such a theory seems anti-Darwinian. If microorganisms have evolved on Earth to occupy every niche, some as inhospitable as those on Mars, why would this not have occurred on Mars? Figure 17 is an electron microscope image of microorganisms found growing in Arctic permafrost in which the temperature is perpetually below freezing. This scenario is no worse than some scenarios on Mars.

Fig. 17. Microbes at the South Pole. Researchers have found evidence that microbes live in the ice at the South Pole (Whitehouse Reference Whitehouse2000).
In addition to rebutting obstacles, other developments have enhanced the prospect for life on Mars.
(a) Desert varnish
The possibility of desert varnish being found on Mars has just been revived as evidence for life (Sherriff Reference Sherriff2006). This issue was first raised in 1977 (Moore et al. Reference Moore, Hutton, Scott, Spitzer and Shorthill1977) and discussed in great detail over ensuing years (DiGregorio Reference DiGregorio2001), with much evidence produced that at least certain types, if not all, desert varnish is mediated by microorganisms. Figure 18 is an image of Mars that shows what may be desert varnish.

Fig. 18. Possible desert varnish on Mars. The front-lit rocks at the Viking landing site show a glistening sheen that may be desert varnish. NASA image, Barry DiGregorio.
(b) Circadian rhythm
All forms of life examined show circadian rhythm, a periodic, cyclic change in metabolism and/or activity. Kinetics of the evolution of gas in the LR Mars signal indicated a possible circadian rhythm. A paper examining possible circadian rhythm in the Mars LR data ‘strongly support(s) the hypothesis of a biological origin’ (Bianciardi Reference Bianciardi2004). Additional work is underway by the author of this method who has been supplied all of the LR response data for his analysis.
(c) Atmospheric indicators
Methane (Formisano et al. Reference Formisano, Atreya, Encrenaz, Ignatiev and Giuranna2004), formaldehyde (Howe Reference Howe2004), and, possibly, ammonia (Whitehouse Reference Whitehouse2004) have been reported in the Martian atmosphere. These gases are indicative of, or consistent with, the presence of life. The methane was associated with water vapour in the lower atmosphere that might reflect the presence of liquid surface water, or, itself, might be able to support life. Methane is a very short-lived gas, and, on Earth, is sustained in the atmosphere by biological activity. An abiological method proposed for Mars was that the gas was emitted by volcanic activity. No significant volcanism has been detected by whole-planet scans of Mars. In addition, a recent study of the amount of gas evolved by the Hawaiian volcano Mona Loa finds that volcanism is a very unlikely source of the Martian methane (Ryan et al. Reference Ryan, Dlugokencky, Tans and Trudeau2006).
The present situation
The present situation with respect to life on Mars may be summarized as follows:
● no finding made on Mars is inimical to life;
● some 40 attempts to explain away the LR findings over the past 30 years have failed scientific scrutiny;
● no abiological experiment has duplicated or realistically approximated the Mars LR positive and control data;
● a Biologic Imperative has been recognized on Earth;
● experimental evidence shows that some microorganisms can survive all the shocks and insults incident to interplanetary travel, and could have been transported there from Earth or elsewhere, as demonstrated in Figure 19 (Levin Reference Levin2005).
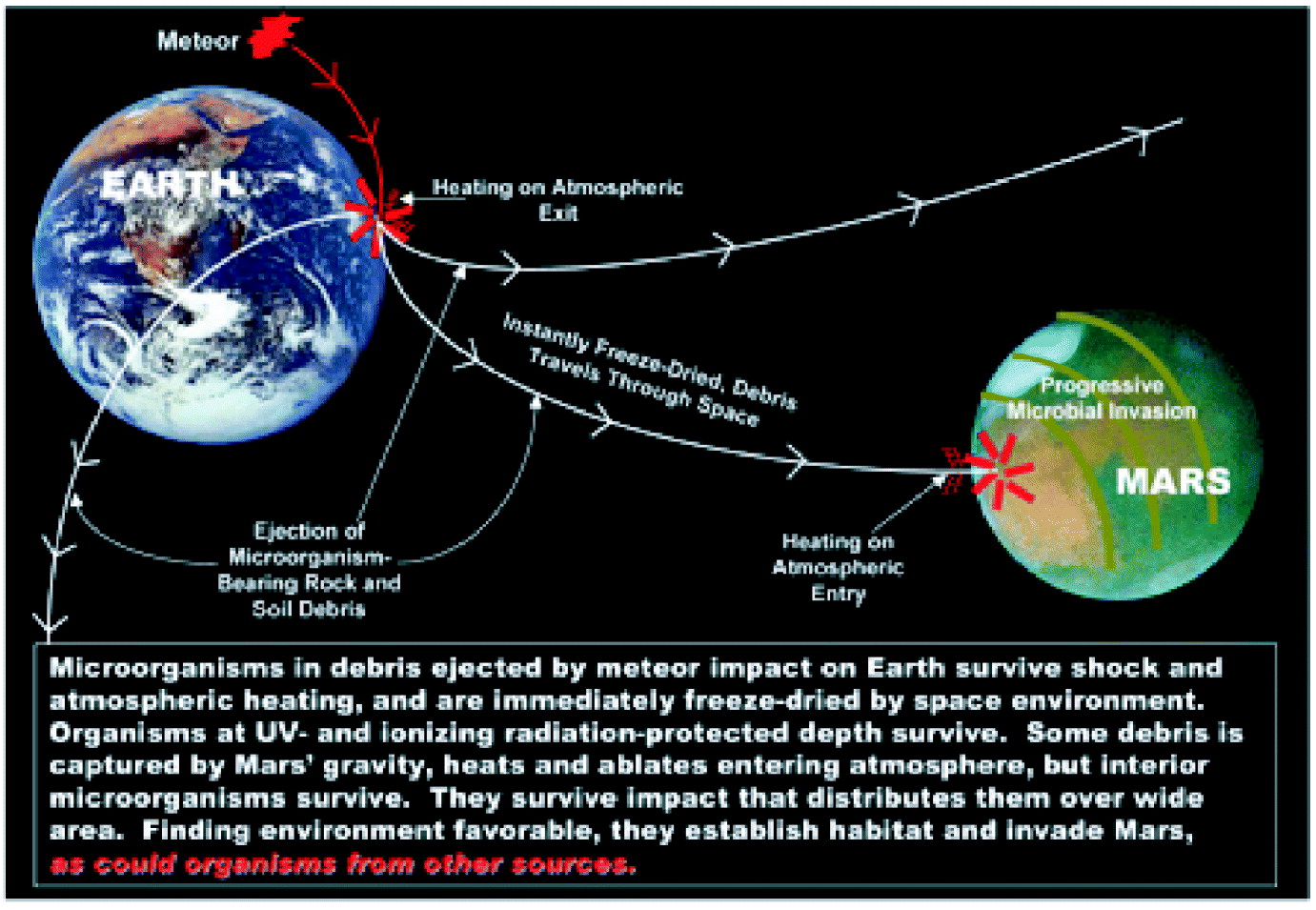
Fig. 19. How the Earth infects Mars.
Nonetheless, the consensus, although moving somewhat towards acceptance of the Viking LR results in recent years, remains negative.
It is now more difficult to propose that Mars is sterile than that it supports microbial life. Earth and Mars share a common biosphere, the extent of which remains unknown. However, science is not a democratic process. Paradigm-breaking discoveries have always been subject to years of scepticism.
I submit that the long-cited ‘Extraordinary claims require extraordinary evidence’ dictum has now been met for life on Mars. The evidence provided by the Mars LR data, now reinforced by pertinent findings on Mars and Earth, and by the continual failures to duplicate the Mars LR data abiologically, have elevated the evidence to ‘extraordinary’. During the same time interval, new knowledge gained concerning life’s habitats and pervasiveness, and its likely ability at interplanetary travel, has removed the LR’s claim from the category of ‘extraordinary’. What would be truly extraordinary in the light of present knowledge would be finding Mars to be sterile.
The time for the paradigm change concerning life on Mars and elsewhere is at hand.
Acknowledgements
The author wishes to thank NASA for its financial support of his life detection efforts from 1959 to 1979. Without the excellent scientific support of Dr Patricia Ann Straat, my Viking LR Co-Experimenter for ten years, the LR would never have made it to Mars. The moral support of Dr Richard B. Hoover, NASA Marshall Space Flight Center, in inviting me to present papers at the SPIE Astrobiology Conferences is gratefully acknowledged. Mrs Kathy Brailer, my long-time Executive Assistant, deserves enormous credit for her excellent assistance in assembling this and other papers by the author and his co-authors. The support of Spherix Inc., which permitted the author to continue his Mars pursuits after 1979, is gratefully acknowledged.





















