Introduction
Tomato (Solanum lycopersicum L.) is an important member of the solanaceae family and is cultivated extensively across the globe for consumption. In 2019, global tomato production was 180.76 Mt from an estimated area of 5.30 Mha, with an average yield of 35.93 t/ha (FAOSTAT, 2019). It has become a popular vegetable in recent years due to its nutritional value, processing traits and nutraceutical values. It is a vital source of several nutrients such as vitamins and minerals that play a significant role in the growth and development of human beings. Interest in tomatoes has grown since the discovery that lycopene has anti-oxidative, anti-cancer properties. The development of plant biotechnology and genetic engineering tools has opened several doors of opportunities for engineering tomato plants (Gerszberg et al., Reference Gerszberg, Hnatuszko-Konka, Kowalczyk and Kononowicz2014). Tomatoes grown for salad must possess deep red colour, flavour and texture to satisfy consumer's preferences and at the same time, be able to withstand transportation, marketing and post-harvest handling. Tomato is in continuous demand throughout the year for its fresh and processed products in the international market (Hannan et al., Reference Hannan, Biswas, Ahmed, Hossain and Islam2007a). It is grown worldwide by farmers with small landholdings grow tomato as a high-value cash crop. Extensive tomato cultivation offers an opportunity to farmers to change from sustenance to commercial farming and considerably boost marginal farmer's income (Fan et al., Reference Fan, Brzeska, Keyzer and Halsema2013).
However, several major constraints including biotic (bacterial, fungal and viral pathogens) and abiotic stresses greatly reduce the yield, fruit quality, nutritional content and shelf-life of tomato fruits (Hanson et al., Reference Hanson, Lu, Wang, Chen, Kenyon and Tan2016). In the past, commercial tomato growing areas have witnessed 100% yield losses worldwide due to tomato leaf curl virus (ToLCV) disease (Singh et al., Reference Singh, Rai, Singh, Singh and Srivastava2014). Furthermore, there exists wide genetic diversity among begomoviruses. So far, more than 100 species have been identified and among which, ToLCV is a prominent disease, usually transmitted by whitefly (Bemisia tabaci). In response to ToLCV infection, plants produce typical symptoms such as arrested growth, reduction in the size of the leaf and internodes, severe curling, twisting and rolling of the leaves that goes together with dark green outgrowth or vein enation on the undersurface of the leaflets (Smith, Reference Smith1935; Vasudeva and Samraj, Reference Vasudeva and Samraj J1948).
To date, there are no effective control measures present to manage the ToLCV disease. It has become practically impossible to control its vector viz., whitefly by chemical means. Moreover, the vector has developed strong resistance against various groups of insecticides in the past few years due to various reasons (Horowitz et al., Reference Horowitz, Kontsedalov, Khasdan and Ishaaya2005) due to which breeding of varieties and hybrids resistant to ToLCV has gained scientific attention (Shankarappa et al., Reference Shankarappa, Sriharsha, Rangaswamy, Aswathanarayana, Prameela, Kulkarni, Muniyappa, Mohanrao and Maruthi2008). The area under tomato cultivation comprises more than 90–95% hybrids as compared to varieties. Hybrids are known to be highly uniform, high yielding, better in terms of quality and possessing a higher level of resistance to biotic and abiotic stresses. Despite the high seed cost, farmers still prefer hybrid cultivars, for increasing their profits. This made the breeders direct their attention more towards hybrid development to satisfy the increased demands of growers and consumers.
The phenomena of heterosis in tomato hybrids have been supported by several investigators in the past (Bai and Lindhout, Reference Bai and Lindhout2007; Solieman et al., Reference Solieman, El-Gabry and Abido2013). The ability of a hybrid to resist the disease also depends upon the degree of resistance present in both the parents. Heterosis is well known to boost the yield and also to enhance other important qualitative and quantitative traits. Heterosis is a natural phenomenon where the F1 hybrid is superior at least over an average of two parents. Two factors i.e. appropriate inbred selection and identification of superior heterotic combinations are extremely decisive in exploiting the heterosis in crop improvement (Singh et al., Reference Singh, Dey, Bhatia, Kumar, Sharma and Behera2019). Combining ability analysis acts as a tool for identification of desirable parent and also specific cross combination for further exploitation (Munshi and Verma, Reference Munshi and Verma1999; Singh et al., Reference Singh, Dey, Bhatia, Kumar, Sharma and Behera2019). Combining ability analysis provides a useful insight into the selection of diverse parents about the performance of the resistant hybrids. The involvement of various types of gene effects in different magnitudes to govern the quality traits which are generally quantitative in nature and also the lack of superior combiners have limited the production of resistant hybrids (Ahmad et al., Reference Ahmad, Quamruzzaman and Nazimuddin2009). Genetic analysis paves the way for assessment of the breeding potential of parental lines or identification of the best combiners in crop (Singh et al., Reference Singh, Rai, Singh, Singh and Srivastava2014), which can be effectively exploited to develop hybrids or accumulate fixable genes to develop a variety.
The choice of effective breeding method and selection of suitable parents for the improvement of any crop depends upon the nature and magnitude of gene action controlling the inheritance of yield and its contributing traits (Rattan and Chadha, Reference Rattan and Chadha2009). Furthermore, the selection of desirable mating designs such as line × tester or half diallel depending upon the number of parents and availability of resources is highly important to unravel the genetic basis of heterosis (Bansal et al., Reference Bansal, Banga and Banga2012). Knowledge about gene action also helps to predict whether one can go for recombination and selection or heterosis breeding. Keeping in view the above facts, the current investigation was undertaken to evaluate the heterotic performance of testcross progenies, developed through half diallel mating design for yield and quality traits as well as for resistance to ToLCV disease.
Materials and methods
Planting material and experimental design
The current study was carried out in the experimental plot (block-8), Division of Vegetable Crops, Indian Institute of Horticultural Research (IIHR), Hesaraghatta (coordinates: 130.58′N latitude and 770.37′E longitude), Bengaluru-89 (Karnataka). The experimentation materials consist of 11 prescreened lines, which were having different Ty genes (Chandel et al., Reference Chandel, Sadashiva and Singh2019), were used as parents to develop 55 F1 hybrids in a half diallel fashion. The crossing was performed in the months of September–October 2018; red ripe fruits were harvested in the last week of December. Seeds were extracted through the fermentation method and dried in sun for 2–3 d. The details about the parental line and their sources of origin are presented in online Supplementary Table S1. Parents, hybrids along with one resistant check Abhinav and one susceptible check Punjab Chhuhara were sown in RCBD design with three replication. Furthermore, evaluation for ToLCV resistance and other horticultural traits was performed from mid-February 2019 onwards. Each replication consists of 15 plants in a single row with a spacing of 100 cm × 45 cm. Beds were covered with 25 μm thick black plastic mulch and fertigation was applied as per package of practices recommended by ICAR-IIHR for the cultivation of tomato. Protective insecticidal sprays were restricted against sucking pest (whiteflies) which serve as a vector for virus transmission and a variety that is susceptible to ToLCV (Punjab Chhuhara) was also planted around the main growing field to develop the viral load.
Agro-morphological characterization
The data for various morphological and horticultural traits were collected from 10 randomly tagged plants per replication. For plant height (cm) data were collected at final harvest, whereas data for the number of flowers per cluster were recorded at 50% blooming stage. The fruit set percentage was calculated by the ratio of number of fruits per cluster to the number of flowers per cluster. All fruits from each plant were weighed and counted respectively to measure yield per plant (kg), average fruit weight (g) and the number of fruits per plant. Five fruits from 10 randomly tagged plants in each replication were collected for recording data of fruit characteristic traits such as the number of locules, total soluble solids (TSS) (using hand refractometer model ERMA 0–10 °Brix FT 327), fruit firmness (kg/cm2) using a needle-type pocket penetrometer, pericarp thickness (cm) using a digital vernier caliper. Disease symptoms and severity of ToLCV were recorded from 15 plants of each genotype in each plot on 30 d after transplanting, 50–60 d after transplanting and 90 d after transplanting (fruit maturation stage) based on plant phenotype. No protection measures were taken during crop growth against whitefly and additional plants of ToLCV susceptible variety Punjab Chhuhara were sown in infector row to maintain a sufficient load of virus inoculum. Plants were examined for symptoms that appeared at 30, 60 and 90 d after transplanting (Muniyappa et al., Reference Muniyappa, Jalikop, Saikia, Channarayappa, Ishwarabhat and Ramappa1991).
DNA extraction and polymerase chain reaction (PCR) reaction conditions
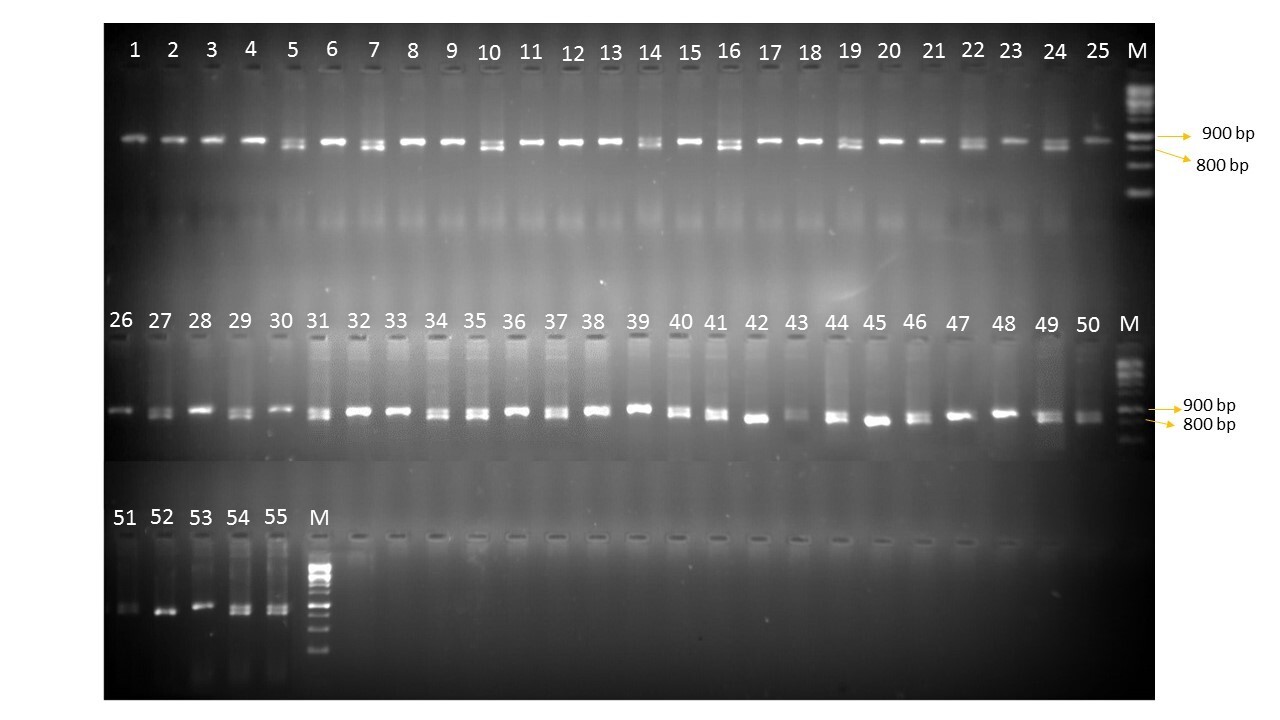
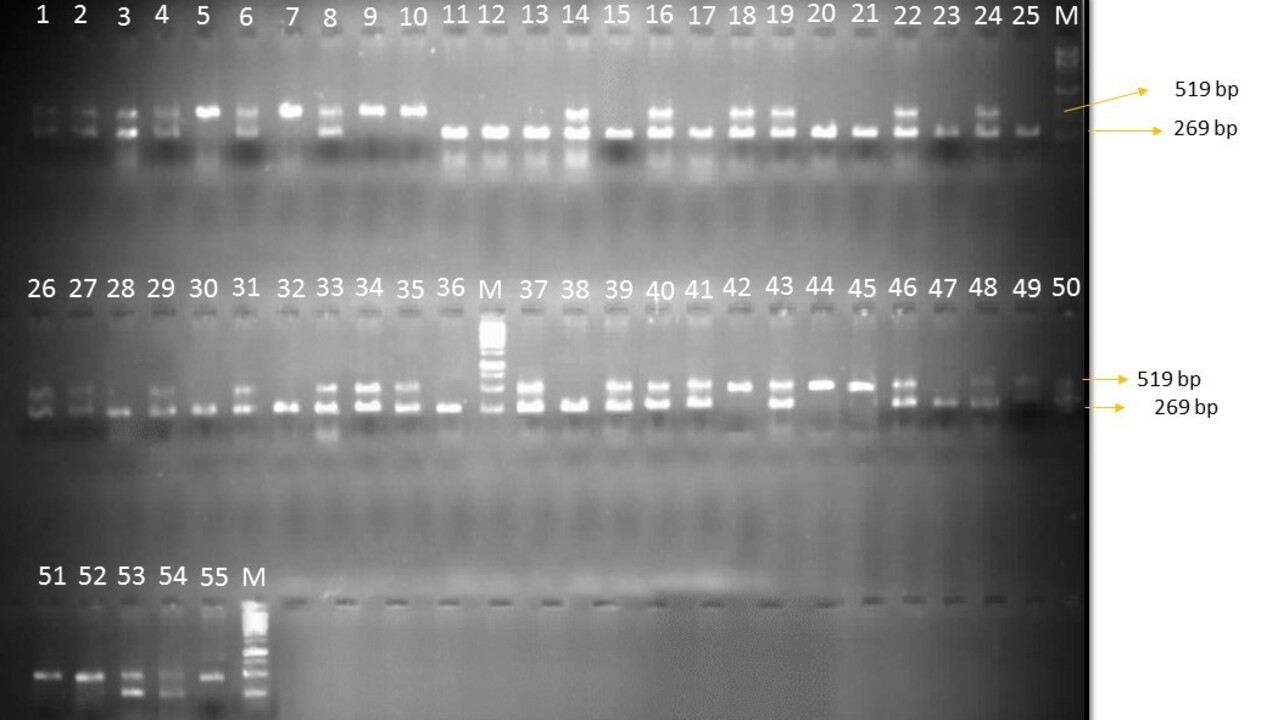
DNA was extracted using the CTAB method from the parents and hybrids at cotyledonary leaves stage of 3–5 d old tomato saplings using a method as suggested by Murray and Thompson (Reference Murray and Thompson1980) with some minor modifications. Total genomic DNA was quantified by using a spectrophotometer at 260 nm; the concentration was calculated against the standard value of 1 OD260 = 50 μg/ml and the integrity was examined by gel electrophoresis on 0.8% (wt/vol) agarose gel. Two SCAR markers, namely TG0302 linked to Ty-2 allele and SCAR1 linked to Ty-3 allele, were used in the current study. PCR was carried out in a thermocycler (Mastercycler Gradient; Eppendorf). PCR amplification was performed in a volume of 10 μl in 0.2 ml thin-walled PCR tube strips containing 1 μl of template DNA, 0.5 μl of each forward and reverse primers, 5 μl of nuclease-free water and 3 μl master mix. Separation and visualization of PCR products were performed on 1.5% agarose gel. Optimal conditions for amplification of SCAR marker linked to Ty-2 were used following Yang et al. (Reference Yang, Caro, Hutton, Scott, Guo and Wang2014). PCR conditions for the SCAR1 marker linked to Ty-3 performed as described by Dong et al. (Reference Dong, Han, Siddique, Kwon, Zhao and Wang2016). A gel documentation system (Alpha imager 2200 of Alpha Innotech Corporation, California) was used to visualize and capture the photograph of PCR products.
Statistical analysis
Heterosis for various traits under consideration was calculated in terms of percent decrease or increase of the F1 hybrids over its better parent, mid-parent and standard check (Hayes et al., Reference Hayes, Immer and Smith1965). Significance was tested at a probability of P < 0.05 and P < 0.01 through the t-test. The diallel analysis for estimation of general combining ability (GCA), specific combining ability (SCA), heritability, variance and mean performance was performed by using the computer software program Windowstat 9.2 (Indostat Services, Hyderabad, India).
Results
Analysis of variance (ANOVA) and genetic components of variance
ANOVA revealed that all the treatments and the parents were significant for all the characters. Similarly among the F1, most of the traits were found to be significant (online Supplementary Table S2). However, the interaction between parents and hybrids was found to be significant for all the traits except for TSS. The diallel ANOVA for combining ability showed the existence of significant variation among the treatments and parents for all the studied traits (Table 1). Highly significant variation due to GCA as well as SCA indicated the importance of additive as well as non-additive types of gene action of inheritance for all the traits. Variance for GCA, SCA and GCA to SCA ratio, additive and dominance variance for 13 characters were estimated and presented in Table 1. The magnitude of SCA variance is more as compared to GCA variance for all the characters under study except for PDI. The GCA to SCA ratio is <1 for all the characters except for PDI. Also, the degree of dominance has shown values greater than unity for all the traits indicated dominant nature of these traits except for PDI. Dominance variance is more than additive variance for all the characters except for percent disease incidence.
Table 1. Estimates of genetic components of variance for yield and quality traits in Tomato

Mean performances of parental lines along with hybrids for horticultural traits
The results for the mean performance of parents and hybrids used in this study are presented in Table 2 and online Supplementary Table S3. Mean values obtained from the parental line indicate wide ranges of genetic variability among them for most of the studied characters. The parental line IIHR-2919 was found to produce the highest mean values for the traits such as plant height, number of flowers per cluster, number of fruits per cluster, lowest number of locules and TSS. For traits concerning earliness such as days to 50% flowering significant lowest mean values were reflected by the parental line IIHR-2898; whereas for yield and yield related traits such as fruit set percentage, number of fruits per plant and yield per plant, parent IIHR-2852 produced the significant highest mean values. Among the used parental lines IIHR-2853 gave the significant highest mean values for fruit diameter and average fruit weight. For traits such as pericarp thickness and fruit firmness, parent IIHR-2907 revealed the significant highest mean values. Furthermore, developed hybrids also reflected wide variations for all traits studied. Results obtained from hybrids showed that most of them produced average values that tended to be either more than their respective mid-parental values or exceeded the better-parental values. Out of 55 hybrids, the highest mean value were found to be those of the F1 hybrids; IIHR-2902 × IIHR-2852 for plant height and average fruit weight; IIHR-2907 × IIHR-2898 for days to 50% flowering; IIHR-2902 × IIHR-2919 for number of flowers per cluster; IIHR-2898 × IIHR-2853 for number of fruits per cluster, TLBR-6 × IIHR-2919 for fruit set percentage; IIHR-5-3-7-5 × IIHR-2853 for number of fruits per plant; IIHR-2919 × IIHR-2898 for number of locules; IIHR-2886 × IIHR-2853 for TSS; IIHR-2913 × IIHR-2853 for pericarp thickness; IIHR-2919 × IIHR-2852 for fruit firmness and IIHR-2913 × IIHR-2898 for yield per plant.
Table 2. Mean performance of parental lines and checks for different yield and quality traits in tomato

Molecular validation of Ty genes
In this experiment, only two markers linked to Ty-2 and Ty-3 alleles have been employed to validate the resistance source (online Supplementary Figs S1 and S2). These upon amplification with TG0302 primers (Ty-2) showed a band size of 900 bp (R) confirming the presence of the Ty-2 gene and SCAR1 marker with a band size of 519 (R) confers the presence of the Ty-3 gene whereas all the other genotypes showed a susceptible reaction with the absence of the resistance governing genes. IIHR-2852 and IIHR-2902 are the only parents which have shown the presence of both Ty-2 and Ty-3 alleles. Fifteen hybrids have shown the presence of Ty-2 alone whereas only three hybrids have shown Ty-3 allele amplification alone in the homozygous state. Only one hybrid namely IIHR-2902 × IIHR-2852 has shown the presence of both alleles i.e. Ty-2 and Ty-3 in the homozygous state.
Estimation of GCA and SCA
The basic approach towards selecting parents for hybrids development based on their mean performance does not always provide fruitful results. Therefore, before drawing any conclusion, we have determined the combining ability, gene action and heterotic potential for all the traits under study. The estimates of desirable GCA and SCA effects of the top three parents and hybrids for various traits under study are presented in Table 3. Nature and magnitude of combining ability effects provide guidelines to identify the better parents and their effective utilization of hybrids development. The obtained results of GCA effects revealed that none of the parents were found to be a good general combiner for all the characters as evident from Table 4. An inclusive assessment of GCA effects showed that among all the parents, IIHR-2919 was a good general combiner for five traits namely number of flowers per cluster, number of fruits per cluster, number of fruits per plant, number of locules and yield per plant whereas for traits such as fruit set percentage, TSS and percent disease incidence parental line IIHR-2852 was found to be the most suitable general combiner. Among all the parental lines, IIHR-2907 was found to be a good general combiner for traits such as pericarp thickness and fruit firmness. Parents IIHR-2853 for plant height; IIHR-2888 for days to 50% flowering and TLBR-6 for average fruit weight have shown the significant GCA effect in the desirable direction. Dominance and performance of specific cross combination can be assessed through SCA of hybrids. In this study, SCA effect of all the 55 hybrids for different traits is represented in online Supplementary Table S4. Plant height is an important growth parameter to support yield and its component traits. The estimate of SCA effect for this trait revealed that crosses IIHR-2902 × IIHR-2852 (33.43) had a significant and higher magnitude of SCA effects. For days to 50% flowering, the highest negative SCA effect is observed in cross TLBR-6 × IIHR-2853 (−3.12). For the number of flowers per cluster, the highest positive SCA effect was observed in cross IIHR-2902 × IIHR-2919 in which parents are high × high GCA combiners. Hybrid IIHR-2898 × IIHR-2853 had the highest SCA effect for the number of fruits per cluster, in which parents are high × low GCA combiners. IIHR-5-3-7-5 × IIHR-2853 exhibited the highest SCA effect for the number of fruits per plant in which parents are L × H type combiners. For average fruit weight, IIHR-2902 × IIHR-2852 had the highest positive SCA effect. SCA effect was found to be minimum in hybrid IIHR-2902 × IIHR-2853 for the number of locules and parents were H × H negative combiners. For TSS IIHR-2886 × IIHR-2852 had the highest positive SCA effect and it has L × H combiners for the traits. IIHR-2913 × IIHR-2853 exhibited the highest positive SCA effect for pericarp thickness and it has L × L combiners. For fruit firmness, IIHR-2919 × IIHR-2853 exhibited the highest positive SCA effect with H × L combiners. The hybrid combinations IIHR-2913 × IIHR-2898, IIHR-2902 × IIHR-2886 and IIHR-2913 × IIHR-2888 were found to be best on the basis of both per se performance and SCA effect for the character namely yield per plant. For percent disease incidence, the highest negative SCA effect was observed in cross IIHR-5-3-7-5 × IIHR-2853 in which parents are H × H combiners.
Table 3. Top three desirable parents and hybrids with their GCA and SCA effects for different yield and quality traits in Tomato

*, **Significant at 5% and 1% probability levels, respectively.
Table 4. Estimates of GCA effects for yield and quality traits in tomato

*, **Significant at 5% and 1% probability levels, respectively.
Estimation of heterosis
The heterotic response of all the 55 testcross progenies varied in magnitude and highly significant heterosis was observed for most of the traits in both directions. Based on heterosis percentage over the standard check for 12 traits along with their better parent heterosis and mid parent heterosis, top five best hybrid combinations have been selected (online Supplementary Table S5). For plant height, 44 hybrids showed significant positive heterosis over the standard check (Abhinav) whereas only 13 hybrids were found to produce significant positive better parent heterosis. Also, 24 hybrids have exhibited significant positive mid-parent heterosis for plant height. Early flowering is generally an indication of early yield (Yordanov, Reference Yordanov1983). With respect to the days to 50% flowering, 20 crosses showed significant negative heterosis over the standard check (Abhinav). Hybrid IIHR-2907 × IIHR-2898 followed by IIHR-2886 × IIHR-2852 and IIHR-2902 × IIHR-2852 were considered as top three best hybrids based on standard heterosis whereas 31 and 22 hybrids have exhibited significantly negative better parent and mid-parent heterosis for days to 50% flowering, respectively. For higher productivity, the number of flowers per cluster is an important factor where greater number of flowers per cluster is preferred. Among 55 testcrosses, 25 and 33 hybrids have shown significant positive better parent and mid-parent heterosis, respectively. However, IIHR-2902 × IIHR-2919 followed by IIHR-2919 × IIHR-2898 and IIHR-2913 × IIHR-2853 were considered as top three best hybrids based on significant positive economic heterosis over the standard check for the number of flowers per cluster. The number of fruits per cluster is an important component that contributes to productivity. Twenty-four hybrids have shown significant positive mid-parent heterosis whereas 15 hybrids showed significant positive better parent heterosis. Only one hybrid IIHR-2898 × IIHR-2853 has shown significant positive economic heterosis over the standard check for the number of fruits per cluster. The yield of fruits per plant is dependent on the fruit-related characters namely, number of fruits per plant and average fruit weight. Thus for this character, one has to select the hybrids exhibiting positive heterosis. Among 55 hybrids, 44 and 28 crosses showed significant positive mid-parent and better parent heterosis for the number of fruits per plant respectively whereas only five hybrids have shown significant positive standard check heterosis. However, for average fruit weight, 20 hybrids have shown significant positive economical heterosis over the standard check. Nineteen hybrids showed significant positive mid-parent heterosis whereas 12 hybrids showed significant positive better parent heterosis. Concerning fruit set percentage, 21 and 13 crosses have exhibited significant positive mid-parent and better parent heterosis, respectively. However, none of the hybrids have exhibited significant positive heterosis over the standard check. In tomato, quality parameters such as a smaller number of locules per fruit, TSS, pericarp thickness and firmness determine the quality related to taste, flavour and utility to processing or fresh market. Higher TSS are desirable for higher recovery of processed products. In this study, 36 hybrids showed significant negative mid-parent heterosis for the number of locules per fruit. IIHR-2919 × IIHR-2898 followed by IIHR-2898 × IIHR-2852 and IIHR-2902 × IIHR-2919 were considered to be the top three best hybrids on the basis of standard heterosis. For TSS, 14 and nine hybrids showed significant positive mid-parent and better parent heterosis respectively. None of the hybrids were found to show significant positive heterosis for TSS. Pericarp thickness plays a much important role in longer storability. Eight hybrids have shown significant positive heterosis over the standard check. However, 21 crosses showed significant positive mid-parent heterosis for pericarp thickness. For fruit firmness out of 55 hybrids, 14 hybrids exhibited significant positive mid-parent heterosis whereas six hybrids have shown significant positive heterosis better parent. Hybrid IIHR-2919 × IIHR-2853 have shown highest significant positive heterosis over the standard check. Further for yield per plant, only one hybrid IIHR-2913 × IIHR-2898 exhibited the significant highest positive standard check heterosis whereas 39 and 27 hybrids have shown significant positive heterosis over mid-parent and better parent for yield per plant.
Discussion
The results pertaining to the mean performance of parents and their hybrids showed significant differences for all the traits under study. Substantial variations for all the studied characters pointed towards the presence of ample genetic variability among the parents and their resultant hybrids. The results concerning plant height, as observed in the current investigation are in close agreement with the findings of Singh and Asati (Reference Singh and Asati2011) and Solieman et al. (Reference Solieman, El-Gabry and Abido2013), where they found significant differences among the studied genotypes of tomato for this character. Ample variations for yield and its contributing traits were also previously reported by Dhatt and Singh (Reference Dhatt and Singh2008), Naveen et al. (Reference Naveen, Cheema and Dhatt2008), Chattopadhyay et al. (Reference Chattopadhyay, Dutta, Dutta and Hazra2011), Solieman et al. (Reference Solieman, El-Gabry and Abido2013), Pandiarana et al. (Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015) and Kaushik and Dhaliwal (Reference Kaushik and Dhaliwal2018).
In any hybrid development programme selection of parents only based on mean performance does not necessarily give useful results (Allard, Reference Allard1960). Therefore, per se performance along with GCA effect of the parents is equally important to select any parent for hybridization programme. Furthermore, estimation of the genetic components of variance namely additive and non-additive is crucial for a fruitful crop improvement programme. The results illustrated that estimates of σ SCA2 were higher in magnitude with respect to σ GCA2 for all of the characters except for PDI. Therefore, the non-additive gene action especially dominance variance played an important role of inheritance in such characters. For such traits, heterosis breeding is an appropriate strategy for their improvement. The outcomes showed that tomato hybrids may perform better in several aspects than either of their parents or commercial varieties. The more consistent and relative contribution estimates of these effects in the genetic control of various traits are governed by several genes of small effect provided by the predictability ratio (Baker, Reference Baker1978). These results indicated the dominance of non-additive gene effects for most of the traits, as the predictability ratios were <0.50 except for the trait percent disease incidence. Earlier studies also showed that non-additive gene action controls fruit yield per plant along with most of the other studied fruit characters (Chattopadhyay et al., Reference Chattopadhyay, Dutta, Dutta and Hazra2011; Shende et al., Reference Shende, Seth, Mukherjee and Chattopadhyay2012; Pandiarana et al., Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015; Kaushik and Dhaliwal, Reference Kaushik and Dhaliwal2018). In this study, the low to intermediate level of h ns2 was observed for the majority of traits suggesting non-additive genetic control of these traits, which might be due to large epistatic effects. The early generation selection for these yield and quality traits would be difficult due to dominance effects in the expression of phenotypic variance. Hence, selection must be done in later generations. The involvement of both additive and non-additive gene actions in the control of average fruit weight and PDI of ToLCV has also been documented previously (Chattopadhyay et al., Reference Chattopadhyay, Dutta, Dutta and Hazra2011; Shende et al., Reference Shende, Seth, Mukherjee and Chattopadhyay2012; Singh et al., Reference Singh, Rai, Singh, Singh and Srivastava2014). Therefore, restricted recurrent selection by intermating the most desirable segregants or diallel selective mating in early segregating generation could be useful breeding strategies for improving such traits which are governed by additive gene action. However, some studies have shown contrary findings including non-additive gene effects for PDI (Pandiarana et al., Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015). Such differences in these studies on the genetic control of this trait may occur due to the use of different parents, the crossing pattern used, the precision of the study and variations in environmental conditions, which can affect estimates of gene actions. The quantitative characters and ToLCV resistance are controlled by a large number of genes and are strongly influenced by the environmental factors (Ahmad et al., Reference Ahmad, Quamruzzaman and Nazimuddin2009). Therefore, it is desirable to make a strict individual plant selection on quantitative characters (Singh et al., Reference Singh, Rai, Singh, Singh and Srivastava2014).
Breeding for enhancing the yield is very difficult as yield is a complex trait and it is governed by several genes with a specific interaction. General performances of the 55 F1 hybrids showed superiority on their performances for the characters such as the number of fruits per plant and yield per plant. Different degrees of gene effects were found to be involved in the inheritance of the number of flowers per cluster, number of fruits per plant and yield per plant. The results were in agreement with the findings of Solieman (Reference Solieman2009), Đorđević et al. (Reference Đorđević, Zečević, Zdravković, Živanović and Todorović2010) and Pandiarana et al. (Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015). None of the parental lines or cross combinations were found to have high GCA or SCA for all the traits studied. However, IIHR-2919 exhibited the maximum GCA for traits such as the number of flower per cluster, number of fruits per cluster, number of fruits per plant, yield per plant (kg), number of locules and PDI for ToLCV significantly in the desired direction. The selected top three cross combinations based on significant SCA effects in a desirable direction for all the traits revealed that no single F1 hybrid exhibited the significant SCA effects for all the studied traits. The SCA effect governs the performance of test cross combination and the role of non-additive gene action in the identification of heterotic hybrids (Das et al., Reference Das, Yadav, Choudhary, Singh, Khade and Chandel2020).
Analysis of the different cross combinations showed that the crosses involved three types of combinations: H × H, H × L or L × H and L × L, where L for non-significant GCA effect either negative/positive in direction and H stands for a significant GCA effect in the desired direction (Pandiarana et al., Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015). In this study, the H × H cross combinations were found in hybrids IIHR-2902 × IIHR-2852 for plant height, IIHR-2902 × IIHR-2919 for number of flower per cluster and number of locules IIHR-2902 × IIHR-2853 for the number of locules, therefore additive and additive × additive interactions were involved in such cross combination. These hybrids would be very desirable in tomato, as promising segregants would be isolated in the early advanced generation whereas hybrids such as IIHR-2913 × IIHR-2898 for yield per plant, IIHR-2898 × IIHR2853 for the number of fruits per cluster, TLBR-6 × IIHR-2919 for fruit set percentage and IIHR-5-3-7-5 × IIHR-2853 for the number of fruits per plant which are showing L × H or H × L cross combinations. This indicates the presence of additive effect predominantly in good combiner and possibly a complementary epistatic effect in the poor combiner, to maximize gene expression, these two gene actions are acting in a complementary fashion (Salimath and Bahl, Reference Salimath and Bahl1985). In hybrids IIHR-2913 × IIHR-2853 for pericarp thickness involving L × L combinations, SCA effects seemed to play a prominent role and show the presence of non-additive gene action (Pandiarana et al., Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015). Significant and positive SCA effects for all of the economic characters of tomato involve these three types of combinations of GCA effects in the parents (Kumar et al., Reference Kumar, Srivastava, Singh, Vasistha, Singh and Singh2013; Solieman et al., Reference Solieman, El-Gabry and Abido2013).
Heterosis breeding is one of the most significant tools to exploit genetic diversity in tomato (Mohanty and Mishra, Reference Mohanty and Mishra1999). The phenomenon of heterosis was reported by Shull (Reference Shull1948) and since then, several researchers have worked on different crops to exploit this phenomenon. The nature and magnitude of heterobeltiosis helps in the identification of promising cross combinations and their exploitation to obtain better transgressive segregants. Earlier studies conducted by using a different set of parents and environments have also reported significant positive mid-parent heterosis for traits such as plant height, number of flower per cluster, number of fruits per plant and yield per plant (Bhatt et al., Reference Bhatt, Biswas and Kumar2001; Shende et al., Reference Shende, Seth, Mukherjee and Chattopadhyay2012; Solieman et al., Reference Solieman, El-Gabry and Abido2013; Pandiarana et al., Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015; Vijeth et al., Reference Vijeth, Dhaliwal, Jindal and Sharma2018). However, significant negative heterosis for characters such as days to 50% flowering and number of locules per fruit were also reported in studies of Sekhar et al. (Reference Sekhar, Prakash, Salimath, Hiremath, Sridevi and Patil2010) and positive heterosis for TSS content by Dhadde et al. (Reference Dhadde, Patil, Dharmatti and Bhat2009). An absence of significant heterosis in most of the cross combinations in the current study may be due to the internal cancellation of heterosis components, as was observed in other previous studies (Shende et al., Reference Shende, Seth, Mukherjee and Chattopadhyay2012; Pandiarana et al., Reference Pandiarana, Chattopadhyay, Seth, Shende, Dutta and Hazra2015).
Biotic and abiotic stresses are major limiting factors in the successful cultivation of tomato (Vijeth et al., Reference Vijeth, Dhaliwal, Jindal and Sharma2018). This study focused on the identification of tomato hybrids resistance to ToLCV along with improved horticultural traits. The parental lines namely IIHR-2852, IIHR-2919, IIHR-2913, IIHR-2902 and IIHR-2907 were symptomless until fruit-harvesting stage whereas the remaining parents developed mild to severe symptoms. The results showed that the presence of Ty-2 and Ty-3 in the parental lines influenced the host response against ToLCV (Prasanna et al., Reference Prasanna, Kashyap, Krishna, Sinha, Reddy and Malathi2015; Hanson et al., Reference Hanson, Lu, Wang, Chen, Kenyon and Tan2016). Parents with Ty-3 resistant alleles either under homozygous or heterozygous conditions remained symptomless. Similar results were also obtained in the presence of a combination of both Ty-2 and Ty-3 genes. Parents or hybrids having Ty-2 alleles alone have shown the development of mild to severe symptoms. These results suggested that Ty-2 had become ineffective against ToLCV which was known to impart good resistance against the leaf curl-causing monopartite virus. Therefore a combination of Ty-2 and Ty-3 genes in the hybrids either in heterozygous for one (Ty-2 or Ty-3) or homozygous for both (Ty-2 and Ty-3) exhibited highly resistant reaction against ToLCV compared to Ty-2 alone carrying hybrids. The results are also in harmony with the previous researchers (Barbieri et al., Reference Barbieri, Acciarri, Sabatini, Sardo, Accotto and Pecchioni2010; Prasanna et al., Reference Prasanna, Kashyap, Krishna, Sinha, Reddy and Malathi2015; Ohnishi et al., Reference Ohnishi, Yamaguchi and Saito2016; Prabhandakavi et al., Reference Prabhandakavi, Kumar, Palicherla, Ramchandran and Pinnamaneni2018; Kaushal et al., Reference Kaushal, Sadashiva, Krishna, Srinivasa, Singh, Sriram, Dhananjay, Venugopalan and Ravishankar2020).
The current investigation revealed that a single parental line or hybrid cannot be considered as a good performer for all the characters. However, line IIHR-2919 was found to show maximum significant GCA in the desirable direction for all the characters except for plant height, days to 50% flowering, fruit set percentage and average fruit weight. Evaluation of heterosis% revealed that hybrid IIHR-2913 × IIHR-2898 can be considered as best performing hybrid for yield and average fruit weight over the standard check Abhinav which also exhibited highest positive and significant SCA effect for these traits. Furthermore, the presence of Ty-3 gene under heterozygous conditions and Ty-2 gene in the homozygous state could result in higher level of resistance against ToLCV monopartite virus at the field level. Therefore, parental line IIHR-2919 could be utilized as a good combiner and isolated to obtain desirable segregates for improving different traits. Also, the hybrid IIHR-2913 × IIHR-2898 can be released for commercial cultivation after further evaluation in multiple locations for multiple years.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262121000630
Acknowledgements
The authors are highly thankful to Division of Vegetable Crops, ICAR-IIHR, Bengaluru and Department of Vegetable Science, COH, Bengaluru (UHS, Bagalkot) for providing all sorts of facilities and financial support for this study. The first author also wants to acknowledge the Indian Council of Agricultural Research (ICAR) for awarding ICAR-PG fellowship during this period of research work. The authors are highly thankful to Dr Neeraj Sankhyan, Assistant Professor (English) Dr YSPUH&F, Nauni, Solan (HP) for assisting in editing of English language throughout the manuscript.