


INTRODUCTION
Distinctive forest vegetation on tropical mountains that are frequently or persistently immersed in ground-level clouds often is described as tropical montane cloud forest (TMCF). These forests often exhibit a variety of ecological features that distinguish them from tropical forests that are more rarely exposed to ground-level clouds, especially a stunted and gnarled canopy with high cover of epiphytes (especially bryophytes and filmy ferns, Hietz Reference HIETZ, Bruijnzeel, Scatena and Hamilton2010). The overriding role of cloud immersion in shaping this vegetation's physiognomy is emphasized by its recurrence across sites for which other environmental factors (e.g. temperature, precipitation, altitude, wind, slope, soils) exhibit wide variation; and in tropical regions with different biogeographic affinities. The mechanisms contributing to distinctive TMCF physiognomy and ecology have received long and detailed study, beginning with Shreve (Reference SHREVE1914) and Brown (Reference BROWN1919), and including a recent synthesis volume detailing current advances (Bruijnzeel et al. Reference BRUIJNZEEL, SCATENA and HAMILTON2010). The objectives of this paper are to provide a concise overview of ecological interactions that characterize these distinctive forests and to suggest directions for future study. We define TMCF primarily on the basis of frequent cloud immersion, distinguishing these tropical montane forests from other tropical forests in drier and warmer environments.
A variety of schemes for classifying tropical montane forest vegetation zones has been devised, and the designation of TMCF generally coincides with tropical montane rain forest (Grubb Reference GRUBB1971) or upper montane rain forest and subalpine forest; this includes forests affected by frequent and/or persistent ground-level cloud (Grubb Reference GRUBB1977). Greater cloud immersion typically accompanies decreased temperatures with increasing altitude (Bruijnzeel & Hamilton Reference BRUIJNZEEL and HAMILTON2000). We emphasize at the outset that the designation of a discrete TMCF type can be misleading because most commonly the composition and structure of tropical montane vegetation exhibits more or less continuous variation across complex environmental gradients (Lieberman et al. Reference LIEBERMAN, LIEBERMAN, PERALTA and HARTSHORN1996). Moreover, precise delineation of the distribution of TMCF is problematic because direct observations of ground-level clouds are not available on an extensive basis. Mulligan (Reference MULLIGAN, Bruijnzeel, Scatena and Hamilton2010) demonstrated that limits of TMCF-classified vegetation coincide best with forested landscapes where greater than 70% of the time satellite-visible clouds and/or ground-level condensing conditions (modelled) occur. Also, strictly speaking tropical cloud forests are not restricted to mountains (Gradstein et al. Reference GRADSTEIN, OBREGON, GEHRIG, BENDIX, Bruijnzeel, Scatena and Hamilton2010), but they are most commonly observed between about 1000 m and 2500 m asl; they are found in sites with a range of temperatures. Jarvis & Mulligan (Reference JARVIS, MULLIGAN, Bruijnzeel, Scatena and Hamilton2010) demonstrated that the climate of TMCFs is significantly different from other tropical montane forests, especially in terms of lower Tmax and Tmean, mostly because they occur at higher altitude. TMCFs tend to be much closer to coasts, as the oceans provide a continuous source of atmospheric moisture to supply cloud formation. Also, cloud forests tend to occur more frequently on topographically exposed landscapes, ridges or peaks, than other montane forests. The geological settings of TMCFs are not distinctive and their soils exhibit a wide range of properties. A particularly distinctive feature of TMCF distribution is the so-called Massenerhebung (or mass-elevation) effect: the elevation of TMCF is much lower on smaller than on larger mountain massifs (Grubb Reference GRUBB1971); the smaller mountains tend to be closer to the sea. The tropical Massenerhebung effect reflects greater cloud immersion on smaller mountains resulting from lower temperatures and a steeper adiabatic lapse rate owing to proximity to the sea; hence, this effect further emphasizes the key role played by climate and cloud immersion in defining TMCF distribution.
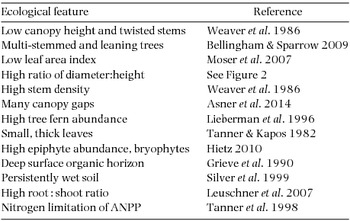
A suite of biotic features characterizes TMCF in contrast to other tropical forests (Table 1). These features vary among TMCFs as a result of differences in environments and floras; for example, not all TMCFs are stunted, and stunting and twisting are not confined to wet TMCF. Also, most research relating environment and vegetation in TMCF has been conducted in the neotropics and a few other locations (e.g. Hawaii), and wider study is needed to better characterize global relationships. Nevertheless, we would argue that certain ecological features comprise a useful, unifying set that defines and aids in analysing the TMCF phenomenon. The low height, or stunting, of the canopy is common in most TMCF. Although decreased forest canopy height accompanies declining temperature at high elevation in most of Earth's mountains, stunting in the TMCF can be extreme (Weaver et al. Reference WEAVER, MEDINA, POOL, DUGGER, GONZALEZ-LIBOY and CUEVAS1986). Moreover, the TMCF stems are often gnarled and twisted or leaning, and multi-stemmed trees are common (Bellingham & Sparrow Reference BELLINGHAM and SPARROW2009). The leaf area index (LAI) in most TMCFs is lower than for lowland forests (Moser et al. Reference MOSER, HERTEL and LEUSCHNER2007, Unger et al. Reference UNGER, HOMEIER and LEUSCHNER2012). The ratio of stem diameter to tree height increases with altitude on most tropical mountains (Girardin et al. Reference GIRARDIN, FARFAN-RIOS, GARCIA, FEELEY, JØRGENSEN, MURAKAMI, PÉREZ, SEIDEL, PANIAGUA, FUENTES CLAROS, MALDONADO, SILMAN, SALINAS, REYNEL, NEILL, SERRANO, CABALLERO, TORRE-CUADROS, MACIA, KILLEEN and MALHI2014a), and stem density is usually higher in TMCF than in lower montane forests. Canopy openings are often more frequent in TMCF (Asner et al. Reference ASNER, ANDERSON, MARTIN, KNAPP, TUPAYACHI, SINCA and MALHI2014) which together with the humid climate and low LAI favours high abundance of tree ferns (Lieberman et al. Reference LIEBERMAN, LIEBERMAN, PERALTA and HARTSHORN1996), and other ferns (Salazar et al. Reference SALAZAR, HOMEIER, KESSLER, ABRAHAMCZYK, LEHNERT, KRÖMER and KLUGE2013). Leaves of many TMCF species are classified as microphylls (Sugden Reference SUGDEN1985, Tanner & Kapos Reference TANNER and KAPOS1982), and leaf thickness is often high. A final canopy feature that is particularly distinctive in most TMCF is the high abundance of epiphytes, especially liverworts, mosses and filmy ferns (Hietz Reference HIETZ, Bruijnzeel, Scatena and Hamilton2010); at the extreme all stem surfaces can be clothed in a thick layer of epiphytes, and epiphyte-derived canopy soil (decaying organic matter) accumulates (Bohlman et al. Reference BOHLMAN, MATELSON and NADKARNI1995, Golley et al. Reference GOLLEY, McGINNIS and CLEMENTS1971).
Table 1. Key characteristics of tropical montane cloud forests.
Although soils of TMCF exhibit a wide range of physical and chemical properties, certain below-ground features appear to be common (Roman et al. Reference ROMAN, SCATENA, BRUIJNZEEL, Bruijnzeel, Scatena and Hamilton2010). Root biomass and the root : shoot ratio of TMCF are higher than in most other closed-canopy forests, and some evidence suggests distinctively high below-ground production (Girardin et al. Reference GIRARDIN, MALHI, ARAGÃO, MAMANI, HUASCO, DURAND, FEELEY, RAPP, SILVA-ESPEJO, SILMAN, SALINAS and WHITTAKER2010, Moser et al. Reference MOSER, LEUSCHNER, HERTEL, GRAEFE, SOETHE and IOST2011). Most TMCF soils exhibit high soil organic matter content and many have a deep organic horizon developed over the mineral soil; this organic horizon may be peat (due to waterlogging) or mor humus (due to acidity and phenolics) or intermediate (Roman et al. Reference ROMAN, SCATENA, BRUIJNZEEL, Bruijnzeel, Scatena and Hamilton2010). Perhaps most importantly TMCF soils are often wet because of the combination of high precipitation and low evaporative demand (cool and humid), and as a result of persistent saturation many TMCF soils are low in oxygen, exhibiting reducing conditions (Silver et al. Reference SILVER, LUGO and KELLER1999). Together with slow nutrient recycling through dead organic matter, these reducing conditions can contribute to chronic nutrient limitation of plant production, especially N limitation (Tanner et al. Reference TANNER, VITOUSEK and CUEVAS1998).
HYPOTHETICAL FRAMEWORK
The environmental and biotic factors that characterize the ecological interactions in TMCF are summarized in a conceptual diagram (Figure 1). Because the distinctive features of TMCF (Table 1) are expressed over a wide range of environments and to differing degrees among TMCFs, the contribution of various driving factors undoubtedly differs among TMCFs; indeed, this variation provides insights for understanding the causes of the TMCF phenomenon.

Figure 1. Hypothesized causal diagram for the effects of environmental factors (in red) shaping cloud forest ecosystem properties (blue) and cloud forest structure (green). Features in bold have been observed in all studies to date whereas other features occur only in some cases.
Following on early work of Shreve (Reference SHREVE1914), Brown (Reference BROWN1919) and many others, we hypothesize that the primary factor shaping the TMCF is the climatic driver: the combination of relatively low temperatures, high humidity and cloud immersion. This primary driver either directly influences, interacts with, or is augmented by a suite of secondary factors to limit the above-ground productivity of TMCF and contribute to their distinctive structure. Here we provide a brief summary of a hypothetical causal framework for the ecosystem features of TMCF followed by a detailed overview of current evidence. We again emphasize that TMCF actually exhibits a broad range of ecosystem features.
The key role played by cloud immersion in shaping the TMCF ecosystem seems clear, but the mechanisms by which cloud immersion leads to the distinctive physiognomy and functions of TMCF are not entirely obvious. Light availability undoubtedly limits net photosynthesis in TMCFs (Figure 1) compared with forests in less cloudy climates. For example, TMCF at 1550 m asl in Jamaica received 19% less shortwave radiation than a site in the drier lowlands near sea level (Aylett Reference AYLETT1985). The low air temperature in high-elevation TMCF limits C assimilation (Wittich et al. Reference WITTICH, HORNA, HOMEIER and LEUSCHNER2012) and net primary productivity (Figure 1) in much the same way as noted for temperate alpine tree-line environments (Tranquillini Reference TRANQUILLINI1979). Moreover, in the cloudy conditions of TMCF leaf temperatures are typically lower than for sunlit leaves, further constraining photosynthesis.
Nutrient limitation of above-ground production is suggested by the high root : shoot production ratio observed in TMCF (Moser et al. Reference MOSER, LEUSCHNER, HERTEL, GRAEFE, SOETHE and IOST2011), and current evidence points to low nitrogen availability as the most common cause of nutrient limitation in TMCFs (Cleveland et al. Reference CLEVELAND, TOWNSEND, TAYLOR, ALVAREZ-CLARE, BUSTAMANTE, CHUYONG, DOBROWSKI, GRIERSON, HARMS, HOULTON, MARKLEIN, PARTON, PORDER, REED, SIERRA, SILVER, TANNER and WIEDER2011, Fisher et al. Reference FISHER, MALHI, TORRES, METCALFE, VAN DE WEG, MEIR, SILVA-ESPEJO and HUASCO2013), though there is good evidence that both P and N are limiting in some TMCF (Homeier et al. Reference HOMEIER, HERTEL, CAMENZIND, CUMBICUS, MARAUN, MARTINSON, POMA, RILIG, SANDMANN, SCHEU, VELDKAMP, WILCKE, WULLAERT and LEUSCHNER2012). Chronic N limitation of above-ground productivity (Figure 1) results from some combination of low input, high losses and delayed recycling owing to suppressed microbial activity in the usually cool and wet TMCF environment.
High annual precipitation in TMCF, including the contribution of wind-driven rain or cloud drip, assures that most TMCFs are not much affected by soil drought. Quite the contrary, high rainfall and low evaporative demand associated with cool, humid atmospheric conditions result in soils that, in many sites, are often at or near saturation (Figure 1). The mechanisms whereby saturated soils contribute to TMCF dynamics are complex. First, the direct effect of low oxygen on root function probably plays a varying role, depending upon plant species and the frequency and extent of anaerobic conditions in soil. In some situations root penetration of soil may be restricted leading to shallow rooting and low resistance to uprooting during windstorms (Soethe et al. Reference SOETHE, LEHMANN and ENGELS2006). Also, nutrient uptake by roots may be further curtailed by low-oxygen conditions. Together these influences contribute further to chronic N limitation and low canopy C assimilation. Moreover, a higher proportion of assimilated C is allocated below ground to acquire soil nutrients and maintain wind-firmness, feeding back to lower above-ground net primary productivity (ANPP, Figure 1).
Another striking feature of TMCF ecosystems is the frequently high accumulation of epiphytes, especially bryophytes, made possible by the high atmospheric moisture (Figure 1). The effect of high epiphyte loads on the other features of TMCF ecosystems has received limited attention. It is possible that the high mass loading of water-saturated bryophytes might lead to structural stability problems in some TMCFs, thereby contributing to observed canopy stunting, high tree diameter : height ratios and greater allocation to root systems. The role of wind in shaping TMCFs (Figure 1) has been suggested (Lawton Reference LAWTON1982) and high winds can induce some of the canopy traits of cloud-forest trees (Cordero Reference CORDERO1999), but TMCF stunting is also expressed in low-wind environments (Martin et al. Reference MARTIN, SHERMAN and FAHEY2007). The high frequency of canopy openings in some TMCFs has been ascribed to low ANPP and slow closure of forest openings, thereby favouring high density of understorey stems (Asner et al. Reference ASNER, ANDERSON, MARTIN, KNAPP, TUPAYACHI, SINCA and MALHI2014).
A final consideration in the structure and function of TMCF is the contribution of phylogenetic vs phenotypic factors. Stunting of TMCF trees is seen in species that grow much taller at lower altitudes; for example, in Jamaica one of the most common species in the most stunted (mor ridge) forest, Clethra occidentalis, also grows to nearly 30 m tall in lowland wet limestone forest. Other species common in the most stunted TMCF in Jamaica are found only as epiphytes at lower altitude (e.g. Clusia havetioides) or absent at lower altitudes (e.g. Lyonia jamaicensis). Conversely, some TMCF, which show extreme stunting, are dominated by species which are rare or absent at lower altitudes, for example the trees dominating the elfin forest on Pico del Este in Puerto Rico (Howard Reference HOWARD1968). Thus, TMCF structure can result from inflexible species characteristics but also from flexible responses to the extreme environment in plastic species.
MECHANISMS AND EVIDENCE
Cloudiness and effects of cloud immersion
Persistent cloudiness and cloud immersion reduce insolation relative to clear sky in TMCFs (19% in Jamaican TMCF, Aylett Reference AYLETT1985, Hafkenscheid Reference HAFKENSCHEID2000; 40% in Puerto Rican TMCF, Baynton Reference BAYNTON1968; 62% in Bolivia, Schawe et al. Reference SCHAWE, GEROLD, BACH, GRADSTEIN, Bruijnzeel, Scatena and Hamilton2010), but surprisingly little research has been conducted to quantify the effects of reduced photosynthetic photon flux density (PPFD) on photosynthetic C gain in TMCF vegetation. In a subtropical montane cloud forest, eddy flux measurements demonstrated that whole-canopy CO2 uptake was reduced by 21% under foggy conditions that reduced insolation by 64% in comparison to non-foggy conditions (Mildenberger et al. Reference MILDENBERGER, BEIDERWIEDEN, HSIA and KLEMM2009; Figure 1). Moreover, the light climate in TMCF is one of very low PPFD in fog potentially followed by very high PPFD in bright sunlight; the switch between high and low light in TMCF could be deleterious. However, evidence to date does not suggest that TMCF species exhibit highly distinctive photosynthetic physiology in comparison with lowland trees. For example, an investigation of the photosynthetic characteristics of leaves from TMCF at 3025 m asl in the Peruvian Andes concluded that the maximum carboxylation capacity and the maximum rate of electron transport were not different from values for lowland rain forest leaves (on a leaf area basis, when calculated for a standard temperature of 25°C), but dark respiration was higher (van de Weg et al. Reference VAN DE WEG, MEIR, GRACE and RAMOS2012). Moreover, van de Weg et al. (Reference VAN DE WEG, MEIR, GRACE and ATKIN2009) observed that leaf optical properties of cloud-forest species were similar to those of rain-forest trees, with typical ranges of plasticity in leaf mass per area with depth in the canopy. In a pantropical survey, Wittich et al. (Reference WITTICH, HORNA, HOMEIER and LEUSCHNER2012) concluded that light-saturated photosynthesis may decline slightly with increasing altitude in tropical mountains, but the explanation of this observation was not clear and possibly is related to inorganic nutrition and low temperature; they suggested that low N and P availability might limit compensatory responses of enzyme activity that could otherwise balance effects of low temperature. In sum, the combination of low total PPFD and low temperatures appears to constrain net C gain in TMCF (Figure 1), but further research is needed to clarify the exact mechanisms. An additional influence of cloud immersion on photosynthesis may result from leaf wetness: water films on the abaxial leaf surface could constrain diffusion of CO2 into the leaf mesophyll and limit photosynthesis (Letts & Mulligan Reference LETTS and MULLIGAN2005). However, data on leaf wetness in cloud-forest trees is needed to confirm the importance of this mechanism.
Cloud immersion strongly affects the ecosystem water balance both by reducing evapotranspiration and through the enhanced precipitation inputs associated with wind-driven fog collected on canopy surfaces. Both potential and actual evapotranspiration rates are considerably lower in TMCF than for adjacent lower montane forests (McJannet et al. Reference McJANNET, WALLACE, REDDELL, Bruijnzeel, Scatena and Hamilton2010, Santiago et al. Reference SANTIAGO, GOLDSTEIN, MEINZER, FOWNES and MUELLER-DOMBOIS2000, Schawe et al. Reference SCHAWE, GEROLD, BACH, GRADSTEIN, Bruijnzeel, Scatena and Hamilton2010), but the magnitude of this difference varies with climatic features both among (Mulligan Reference MULLIGAN, Bruijnzeel, Scatena and Hamilton2010) and within (Lawton et al. Reference LAWTON, NAIR, RAY, REGMI, POUNDS, WELCH, Bruijnzeel, Scatena and Hamilton2010) montane regions. Recent advances in the measurement of fog interception have clarified the role of cloud immersion in augmenting precipitation in TMCF. Holwerda et al. (Reference HOLWERDA, BRUIJNZEEL, OORD, SCATENA, Bruijnzeel, Scatena and Hamilton2010) concluded that cloud-water interception exceeded 1 mm d−1 in a Puerto Rican elfin forest, and Häger & Dohrenbusch (Reference HÄGER and DOHRENBUSCH2011) reported that throughfall was 800 mm y−1 higher than rainfall in north-western Costa Rica. Giambelluca et al. (Reference GIAMBELLUCA, DELAY, NULLET, SCHOLL, GINGERICH, Bruijnzeel, Scatena and Hamilton2010) estimated over 1000 mm y−1 in a wet cloud forest in Hawaii; however, they concluded that most of the intercepted water was re-evaporated rather than reaching the ground as cloud drip. The combined effect of reduced evapotranspiration and enhanced precipitation associated with cloud immersion might increase the frequency and duration of soil saturation in TMCFs (Schawe et al. Reference SCHAWE, GEROLD, BACH, GRADSTEIN, Bruijnzeel, Scatena and Hamilton2010; Figure 1).
A final direct effect of cloud immersion contributing to the TMCF ecosystem is associated with the composition and abundance of epiphytic plants. The diversity and biomass of both vascular and non-vascular epiphytes is high in most TMCFs, and cloud immersion undoubtedly contributes to this pattern (Figure 1). The most distinctive feature of the epiphyte vegetation in many TMCFs is the high abundance of non-vascular species and filmy ferns (Hymenophyllum spp.) that often cover nearly all stem surfaces (Hietz Reference HIETZ, Bruijnzeel, Scatena and Hamilton2010, Horwath Reference HORWATH2011). The epiphytic bryophytes of TMCF are predominantly leafy liverworts (Jungermanniales) in contrast to lowland rain forest where true mosses can be equally common (Horwath Reference HORWATH2011). Among ferns, the genera Hymenophyllum and Elaphoglossum and the family Grammitidaceae are characteristic of TMCF (Hietz Reference HIETZ, Bruijnzeel, Scatena and Hamilton2010, Kessler et al. Reference KESSLER, PARRIS and KESSLER2001, Krömer et al. Reference KRÖMER, KESSLER, GRADSTEIN and ACEBEY2005). High atmospheric moisture particularly favours these taxa, and absorption of cloud water probably contributes to their water supply (Shreve Reference SHREVE1911, Tobón et al. Reference TOBÓN, BRUIJNZEEL, FRUMAU, CALVO-ALVARADO, Bruijnzeel, Scatena and Hamilton2010). The possible role of cloud-water in supplying growth-limiting mineral nutrients to epiphytic vegetation, or favouring epiphytic N2-fixing taxa also is notable (Hietz et al. Reference HIETZ, WANEK, WANIA and NADKARNI2002). Conversely, epiphyte vegetation including epiphylls could restrict tree photosynthesis by growing over leaves and reducing light availability (Grubb Reference GRUBB1977). To our knowledge, no measurements of this effect have been made for TMCF.
Nutrient limitation
The role of mineral nutrient limitation in contributing to TMCF ecosystem dynamics has been an enduring theme of TMCF studies. An early idea that reduced convective transport associated with low transpirational water flux limits nutrient acquisition (Odum Reference ODUM, Odum and Pigeon1970) has been largely discounted (Bruijnzeel & Veneklaas Reference BRUIJNZEEL and VENEKLAAS1998, Grubb Reference GRUBB1977), but relatively low foliar nutrient concentrations (on a mass basis) clearly pointed to the likelihood of nutrient limitation (Grubb Reference GRUBB1977). Tanner et al. (Reference TANNER, VITOUSEK and CUEVAS1998) summarized experimental evidence for nutrient limitation of TMCF productivity and noted that most, but not all, studies indicated nutrient limitation. Moreover, they concluded that N was usually the principal limiting nutrient (Figure 1), a conclusion that has been borne out in more recent fertilization studies (Cleveland et al. Reference CLEVELAND, TOWNSEND, TAYLOR, ALVAREZ-CLARE, BUSTAMANTE, CHUYONG, DOBROWSKI, GRIERSON, HARMS, HOULTON, MARKLEIN, PARTON, PORDER, REED, SIERRA, SILVER, TANNER and WIEDER2011, Fisher et al. Reference FISHER, MALHI, TORRES, METCALFE, VAN DE WEG, MEIR, SILVA-ESPEJO and HUASCO2013). These results agree with some observations of decreasing foliar N : P ratios with increasing altitude in tropical mountains (Soethe et al. Reference SOETHE, LEHMANN and ENGELS2008, van de Weg et al. Reference VAN DE WEG, MEIR, GRACE and ATKIN2009). Photosynthetic parameters of TMCF trees also support N rather than P limitation of C assimilation (van de Weg et al. Reference VAN DE WEG, MEIR, GRACE and RAMOS2012). However, colimitation by N and P may occur in some TMCF (Homeier et al. Reference HOMEIER, HERTEL, CAMENZIND, CUMBICUS, MARAUN, MARTINSON, POMA, RILIG, SANDMANN, SCHEU, VELDKAMP, WILCKE, WULLAERT and LEUSCHNER2012), depending upon local soil properties (Wittich et al. Reference WITTICH, HORNA, HOMEIER and LEUSCHNER2012). Moreover, recent measurements of nitrate leaching from a TMCF in Costa Rica (Brookshire et al. Reference BROOKSHIRE, HEDIN, NEWBOLD, SIGMAN and JACKSON2012) call into question the universal role of soil N availability in limiting TMCF productivity. A resolution to this apparent paradox – that tree growth is limited by N in soils from which water with low concentrations of nitrate is draining – may simply be a question of relative amounts, as nitrate is very soluble and it may be impossible for tree mycorrhizas to take it all up. Early studies in Puerto Rico reported nitrate in stream water (McDowell & Asbury Reference McDOWELL and ASBURY1994) in an area where fertilization of forest recovering from a hurricane resulted in about a doubling of leaf-litter production (Zimmerman et al. Reference ZIMMERMAN, PULLIAM, LODGE, QUIÑONES-ORFILA, FETCHER, GUZMÁN-GRAJALES, PARROTTA, ASBURY, WALKER and WAIDE1995).
Evidence about the mechanisms contributing to apparent N limitation of TMCF productivity is not entirely conclusive; any or all of low N inputs, delayed recycling or high N losses could contribute to chronic N limitation. Benner et al. (Reference BENNER, VITOUSEK, OSTERTAG, Bruijnzeel, Scatena and Hamilton2010) concluded that the generally low nutrient status of TMCF compared with other tropical forests probably is not associated with chronically low nutrient inputs but rather with differences in the rates of nutrient cycling or losses. However, low temperatures have been linked to limitation of N2 fixation in Earth's terrestrial biomes (Houlton et al. Reference HOULTON, WANG, VITOUSEK and FIELD2008), and although they are common in lowland tropical forests, symbiotic N2-fixing tree taxa are uncommon in TMCF. Nitrogen fixation by lichens (Forman Reference FORMAN1975) and other canopy epiphytes clearly occurs as indicated both by direct measurements (Benner et al. Reference BENNER, CONROY, LUNCH, TOYODA and VITOUSEK2007) and by isotopic evidence (Hietz et al. Reference HIETZ, WANEK, WANIA and NADKARNI2002). Free-living N2 fixation also occurs in forest floor and canopy soils, but the quantities remain highly uncertain and more research is needed (Matson et al. Reference MATSON, CORRE, BURNEO and VELDKAMP2015). Intriguingly, Benner & Vitousek (Reference BENNER and VITOUSEK2007) observed that the abundance of N2-fixing canopy epiphytes was stimulated by soil P fertilization in a Hawaiian TMCF, suggesting a possible interaction between P and N supply. The high accumulation of soil organic matter in TMCF is indicative of delayed recycling of nutrients and could contribute to chronic N2 limitation of NPP (Tanner et al. Reference TANNER, VITOUSEK and CUEVAS1998, Unger et al. Reference UNGER, HOMEIER and LEUSCHNER2012). Declining temperature appears to be the principal rate-limiting factor for declining litter decay with increasing altitude in tropical mountains (Salinas et al. Reference SALINAS, MALHI, MEIR, SILMAN, ROMAN CUESTA, HUAMAN, SALINAS, HUAMAN, GIBAJA, MAMANI and FARFAN2011, Schuur Reference SCHUUR2001), but a possible role of excess water and low O2 is described below; this factor also could contribute to increased losses of N (Figure 1).
Mycorrhizal associations undoubtedly play a key role in mediating nutrient relations in TMCF, but they have received limited attention to date. Although arbuscular mycorrhizas are associated with most TMCF tree genera (Brundrett Reference BRUNDRETT2009), many species of Ericaceae, with their distinctive ericoid mycorrhizal associations, are common as TMCF epiphytes (Rains et al. Reference RAINS, NADKARNI and BLEDSOE2003) and at environmental extremes in tropical mountains (Gentry et al. Reference GENTRY, CHURCHILL, BALSLEV, FORERO and LUTEYN1995). Moreover, in some tropical regions, tree species supporting ectomycorrhizal associates from the families Fagaceae and Pinaceae are common in TMCF. These mycorrhizal associations exhibit some systematic differences in nutrient acquisition mechanisms (Marschner & Dell Reference MARSCHNER and DELL1994); studies of their efficacy across TMCF ectones where taxa with the different associations are present or absent might be particularly informative for understanding nutrient relations of TMCF.
Nutrient limitation could contribute to TMCF ecosystem dynamics either directly by constraining photosynthetic activity or indirectly by diverting C from above-ground growth to below-ground resource acquisition (Figure 1). Fisher et al. (Reference FISHER, MALHI, TORRES, METCALFE, VAN DE WEG, MEIR, SILVA-ESPEJO and HUASCO2013) concluded that N fertilization of TMCF promoted higher stem growth not by increasing C assimilation but through effects on below-ground allocation. Some studies have demonstrated that root : shoot ratios increase with altitude on tropical mountains (Kitayama & Aiba Reference KITAYAMA and AIBA2002, Leuschner et al. Reference LEUSCHNER, MOSER, BERTSCH, RODERSTEIN and HERTEL2007), and Girardin et al. (Reference GIRARDIN, ARAGÃO, MALHI, HUARACA HUASCO, METCALFE, DURAND, MAMANI, SILVA-ESPEJO and WHITTAKER2013) provided evidence for a step increase in the cloud immersion zone. Moreover, some evidence suggests that below-ground production is especially high in TMCF (Girardin et al. Reference GIRARDIN, MALHI, ARAGÃO, MAMANI, HUASCO, DURAND, FEELEY, RAPP, SILVA-ESPEJO, SILMAN, SALINAS and WHITTAKER2010, Moser et al. Reference MOSER, LEUSCHNER, HERTEL, GRAEFE, SOETHE and IOST2011), although measurement of below-ground production is notoriously difficult and uncertain. Taken together, these observations suggest that the low stature and low ANPP of TMCF results in part from relatively high below-ground C allocation associated with chronic N (and sometimes P) limitation (Figure 1).
Excess soil water and reducing conditions
Although not all TMCFs exhibit frequent and persistent soil saturation (Roman et al. Reference ROMAN, SCATENA, BRUIJNZEEL, Bruijnzeel, Scatena and Hamilton2010), this is clearly a common feature in many TMCFs (Schawe et al. Reference SCHAWE, GEROLD, BACH, GRADSTEIN, Bruijnzeel, Scatena and Hamilton2010). Chronically high water content of soils leads to the depletion of oxygen. Indeed, field studies indicate that anaerobic conditions increase along some tropical montane elevation gradients (Silver et al. Reference SILVER, LUGO and KELLER1999). Low soil oxygen has been suggested as a factor limiting the LAI of some TMCF (Santiago et al. Reference SANTIAGO, JONES, GOLDSTEIN, Bruijnzeel, Scatena and Hamilton2010). However, it seems likely that the tolerance of low soil oxygen varies considerably among TMCF species so that the contribution of reducing conditions will vary with the phylogenetic history of the extant floras. Rapp et al. (Reference RAPP, SILMAN, CLARK, GIRARDIN, GALIANO and TITO2012) attributed the low ANPP of TMCF in part to such species effects. Notably, some tall, highly productive forests are found in low oxygen floodplain soils and swamps (Grubb Reference GRUBB1977) including TMCF (e.g. at Monteverde, Costa Rica; R. Lawton, pers. comm.).
Under anaerobic conditions organic matter decomposition is slow and incomplete, contributing to the high accumulation of soil organic matter in TMCF. Schuur (Reference SCHUUR2001) demonstrated that lower tissue quality of leaf litter in wetter sites also can contribute to reduced decay rates, high SOM accumulation and delayed nutrient recycling. Such a mechanism appears not to be universally important in TMCF, however, as Silver et al. (Reference SILVER, THOMPSON, HERMAN, FIRESTONE, Bruijnzeel, Scatena and Hamilton2010) indicated, net N mineralization rates are not consistently different between TMCF and low-elevation tropical forest in Puerto Rico. Moreover, TMCF in their study exhibited higher gross N mineralization rates than low-elevation soils and no effect of anaerobic conditions was observed. Clearly, further study of the interactions between soil N dynamics and soil saturation in TMCF is warranted.
Other mechanisms associated with excess soil water could contribute to nutrient limitation in TMCF. For example, anaerobic conditions can limit root growth and physiological activity in many species. Indeed, aerial and canopy roots are a common feature of wet TMCF (Gill Reference GILL1969, Nadkarni Reference NADKARNI1981, Santiago et al. Reference SANTIAGO, GOLDSTEIN, MEINZER, FOWNES and MUELLER-DOMBOIS2000). Low-oxygen conditions also may favour losses of N by denitrification, and high losses of soil nutrients may accompany the greater hydrological outputs in wetter tropical soils. Although losses of mineral N forms should be constrained on N-limited sites (but see Brookshire et al. Reference BROOKSHIRE, HEDIN, NEWBOLD, SIGMAN and JACKSON2012), leaching of organic N may be favoured by the accumulation of recalcitrant organic matter under cool or anaerobic conditions (Perakis & Hedin Reference PERAKIS and HEDIN2002). Also, by restricting the depth distribution of tree roots, anaerobic conditions may either restrict access to available nutrients or promote their losses. It is also possible that shallow rooting in some TMCF reflects ease of access to soil water and superficial nutrient supply; however, as noted previously, shallow rooting also may reduce the wind-firmness of trees and increase the frequency of disturbance with possible long-term effects on nutrient losses.
Topographic effects
Variation of topography in tropical montane landscapes clearly plays a role in regulating TMCF structure, and provides further indications about the contributing mechanisms. Most TMCFs occur under highly variable topography including steeper slopes (Asner et al. Reference ASNER, ANDERSON, MARTIN, KNAPP, TUPAYACHI, SINCA and MALHI2014) and more landslides (Larsen & Torres-Sanchez Reference LARSEN and TORRES-SÁNCHEZ1998, Shreve Reference SHREVE1914) than lower montane landscapes. Tanner (Reference TANNER1977) demonstrated that systematic variation in forest structure of TMCF was related to landscape position and subsequent studies have confirmed this relationship (Wardle et al. Reference WARDLE, BELLINGHAM, KARDOL, GIESLER and TANNER2015). For example, several studies report that canopy height in TMCF is lower on steep slopes (Roman et al. Reference ROMAN, SCATENA, BRUIJNZEEL, Bruijnzeel, Scatena and Hamilton2010), an observation that would also question the universal role of poor soil drainage in shaping TMCF ecosystems. This pattern might be explained in part by disturbance history associated with landslides or more generally by soil stability and tree rooting (Soethe et al. Reference SOETHE, LEHMANN and ENGELS2006). Werner & Homeier (Reference WERNER and HOMEIER2015) proposed a nutrient feedback mechanism to explain striking contrasts in forest structure and composition between slope positions in tropical montane landscapes: downslope nutrient transport could result in spatial contrasts in soil nutrient availability, reinforced by differences in organic recycling owing to litter quality responses to soil fertility. However, in some situations TMCF stunting is most strongly expressed in saddles where soils are enriched and moisture may accumulate (Wilcke et al. Reference WILCKE, BOY, GOLLER, FLEISCHBEIN, VALAREZO, ZECH, Bruijnzeel, Scatena and Hamilton2010). Girardin et al. (Reference GIRARDIN, SILVA ESPEJOB, DOUGHTY, HUASCO, METCALFE, DURAND-BACA, MARTHEWS, ARAGAO, FARF~N-RIOS, GARCIA-CABRERA, HALLADAY, FISHER, GALIANO-CABRERA, HUARACA-QUISPE, ALZAMORA-TAYPE, EGUILUZ-MORA, SALINAS-REVILLA, SILMAN, MEIR and MALHI2014b) associated the strong expression of the TMCF structural features with local topography that favoured cloud convergence and formation. At Monteverde, Costa Rica, maximum TMCF stunting also has been associated with topographic effects on wind exposure and cloud formation (Lawton Reference LAWTON1982, Lawton et al. Reference LAWTON, NAIR, RAY, REGMI, POUNDS, WELCH, Bruijnzeel, Scatena and Hamilton2010). Conversely, Schwarzkopf et al. (Reference SCHWARZKOPF, RIHA, FAHEY and DEGLORIA2011) noted that extreme differences in TMCF structure were unrelated to topographic variables in an Andean landscape.
Forest structure in TMCF
Stunting of trees and low canopy height are commonly observed in TMCF (Grubb Reference GRUBB1977). Because trees in closed-canopy forest compete intensely for the light resource, height growth is important to individual tree success. Thus, unless there are other advantages of being stunted, trees would be expected to attain the maximum height that resource limitations permit; however, greater relative competition for soil resources than light in TMCF might contribute to lower investment in height growth. In any case, all the environmental factors that constrain plant production (low insolation, cold temperatures, nutrient limitation) undoubtedly contribute to tree stunting in TMCF (Figure 1). Less clear is the explanation for increases in the ratio of tree diameter to height with increasing elevation on tropical mountains (Grubb Reference GRUBB1977, Lieberman et al. Reference LIEBERMAN, LIEBERMAN, PERALTA and HARTSHORN1996). The consistency of this relationship is indicated by a summary for 11 altitudinal transects in different regions of the world (Figure 2).

Figure 2. Tree diameter to canopy height ratio (cm m−1) for forests along 11 altitudinal transects on tropical mountains. Mean diameter for trees >10 cm dbh except for Ecuador (>5 cm dbh). References as follows: Ecuador (Leuschner et al. Reference LEUSCHNER, MOSER, BERTSCH, RODERSTEIN and HERTEL2007); Ecuador-2 (Homeier et al. Reference HOMEIER, HERTEL, CAMENZIND, CUMBICUS, MARAUN, MARTINSON, POMA, RILIG, SANDMANN, SCHEU, VELDKAMP, WILCKE, WULLAERT and LEUSCHNER2012); Borneo (Kitayama Reference KITAYAMA1992); Borneo-1 (Aiba & Kitayama Reference AIBA and KITAYAMA1999 non-ultrabasic soils); Borneo-2 (Aiba & Kitayama Reference AIBA and KITAYAMA1999 ultrabasic soils); Malaysia (Proctor et al. Reference PROCTOR, LEE, LANGLEY, MUNRO and NELSON1988); Costa Rica (Lieberman et al. Reference LIEBERMAN, LIEBERMAN, PERALTA and HARTSHORN1996); Hawaii (Raich et al. Reference RAICH, RUSSELL and VITOUSEK1997); Dominican Republic (DR, Sherman et al. Reference SHERMAN, MARTIN and FAHEY2005); Peru (Andes Biodiversity and Ecosystem Research Group [ABERG] 2014). Long-term plot inventory data. Unpublished data at http://www.andesconservation.org.
The factors influencing D : H relationships in trees have been analysed and debated, and include mechanical constraints (King et al. Reference KING, DAVIES, TAN and NOOR2009, McMahon Reference McMAHON1973); drought and hydraulic constraints (Niklas & Spatz Reference NIKLAS and SPATZ2004, Sperry et al. Reference SPERRY, MEINZER and McCULLOH2008); influences of neighbouring trees on both access to light and exposure to wind (King Reference KING1996); and phylogenetic constraints. Because canopy height is low in TMCF the advantage of monopodial growth to limit light competition is reduced and multi-stemmed architecture can have selective value (Givnish Reference GIVNISH, Medina, Mooney and Vazquez-Yanes1984). Such architecture is common in many TMCFs (Figure 1; Bellingham & Sparrow Reference BELLINGHAM and SPARROW2009, Culmsee et al. Reference CULMSEE, LEUSCHNER, MOSER and PITOPANG2010).
In addition, trees would be expected to maintain a safety factor against mechanical buckling to reduce risks associated with storm winds (de Gouvenain & Silander Reference DE GOUVENAIN and SILANDER2003) although for multi-stemmed individuals a sprouting trunk can compensate for failure of stems. King et al. (Reference KING, DAVIES, TAN and NOOR2009) suggested trees in locations protected from wind in lowland tropical forest approached the theoretical buckling height, whereas most TMCF forests exhibit very low values (Table 2). Feldpausch et al. (Reference FELDPAUSCH, BANIN, PHILLIPS, BAKER, LEWIS, QUESADA, AFFUM-BAFFOE, ARETS, BERRY, BIRD, BRONDIZIO, DE CAMARGO, CHAVE, DJAGBLETEY, DOMINGUES, DRESCHER, FEARNSIDE, FRANÇA, FYLLAS, LOPEZ-GONZALEZ, HLADIK, HIGUCHI, HUNTER, IIDA, SALIM, KASSIM, KELLER, KEMP, KING, LOVETT, MARIMON, MARIMON-JUNIOR, LENZA, MARSHALL, METCALFE, MITCHARD, MORAN, NELSON, NILUS, NOGUEIRA, PALACE, PATIÑO, PEH, RAVENTOS, REITSMA, SAIZ, SCHRODT, SONKÉ, TAEDOUMG, TAN, WHITE, WÖLL and LLOYD2011) summarized the factors related to D:H variation across a large dataset for the world's tropical forests. Among the climatic influences was an effect of temperature, but this effect was confined only to the influence of altitude, reflecting the relationships in Figure 2. Recently, Asner et al. (Reference ASNER, ANDERSON, MARTIN, KNAPP, TUPAYACHI, SINCA and MALHI2014) observed that TMCF trees maintained crown shapes in which a greater proportion of the foliage was held low in the canopy than for trees at lower altitudes which might contribute to mechanical stability.
Table 2. Canopy height, estimated water-saturated biomass of canopy epiphytes and ratio of actual canopy height (Hact) to buckling height (Hcrit) for six tropical montane forests. Epiphyte biomass from Kӧhler et al. (Reference KÖHLER, TOBÓN, FRUMAU and BRUIJNZEEL2007), except for Peru (from Horwath Reference HORWATH2011). Hcrit = C ×
 $(\frac{E}{\gamma})$
1/3 × D2/3, where C = constant (0.79 for columnar tapering); E is elastic modulus; ⋎ is stem wood density; and D is stem diameter;
$(\frac{E}{\gamma})$
= 117.6 m for angiosperms (Niklas Reference NIKLAS1994; and pers. comm.).
$(\frac{E}{\gamma})$
1/3 × D2/3, where C = constant (0.79 for columnar tapering); E is elastic modulus; ⋎ is stem wood density; and D is stem diameter;
$(\frac{E}{\gamma})$
= 117.6 m for angiosperms (Niklas Reference NIKLAS1994; and pers. comm.).

Perhaps the single most likely cause of the increasing D : H ratio with altitude is increasing windspeeds (Woodward Reference WOODWARD1993). As noted above Lawton (Reference LAWTON1982) attributed high D : H ratio in a cloud-forest tree species to wind exposure, and King et al. (Reference KING, DAVIES, TAN and NOOR2009) observed a higher mechanical safety factor for lowland rain-forest trees on exposed ridges than protected coves. The possible role of soil properties influencing tree stability has also been suggested (Soethe et al. Reference SOETHE, LEHMANN and ENGELS2006) as shallow rooting in thin or waterlogged soils could necessitate wide root crowns for trees to remain wind-firm.
Another possible factor influencing the low canopy height and high D : H ratio of TMCF is the extra canopy loading associated with high epiphyte and canopy soil biomass in the cloud immersion zone (Köhler et al. Reference KÖHLER, TOBÓN, FRUMAU and BRUIJNZEEL2007). Although the dry biomass of canopy epiphytes is typically only a small fraction of the tree canopy mass, epiphytic bryophytes can hold several times their dry mass in water when they are immersed in clouds (Köhler et al. Reference KÖHLER, TOBÓN, FRUMAU and BRUIJNZEEL2007), and canopy soil adds still more to the canopy load (Bohlman et al. Reference BOHLMAN, MATELSON and NADKARNI1995). The water-saturated mass of canopy epiphytes in several cloud forests ranges from 13 to 45 Mg ha−1, values which are substantial relative to leaf and branch biomass. For example, the biomass of wet epiphytes and canopy soil in the Monteverde cloud forest in Costa Rica (33.1 Mg ha−1; Nadkarni Reference NADKARNI1984) is about half of leaf and branch biomass (60 Mg ha−1; Nadkarni et al. Reference NADKARNI, SCHAEFER, MATELSON and SOLANO2004), and comparable proportions are likely in other cases (Horwath Reference HORWATH2011). The theoretical critical height (Hcritical) at which buckling would occur can be calculated on the basis of tree architecture and wood properties (Niklas Reference NIKLAS1994), and notably, the ratio of actual canopy height to buckling height (Hactual : Hcritical; the static mechanical safety factor) is particularly low for some epiphyte-laden forests (0.25–0.31; Table 2); much higher values are noted for lower montane forest (e.g. 0.7 at 750 m asl in Costa Rica; Lieberman et al. Reference LIEBERMAN, LIEBERMAN, PERALTA and HARTSHORN1996). However, some stunted cloud forests have much smaller epiphyte loads; the Hactual : Hcritical ratio of the stunted mor forest in Jamaica is 0.30, yet the epiphyte mass is only 2.1 Mg ha−1. Thus, although the possible role of mechanical constraints associated with epiphyte loading in stunted TMCF deserves further study, the evidence at present is not conclusive.
The role of phylogenetic factors affecting canopy height and forest structure also deserves attention. In some notable cases particular species – e.g. from Pinaceae, Fagaceae, Eucalyptus spp. – attain much greater heights within the cloud zone than the species that form the main canopy. In the Dominican Republic, the native Pinus occidentalis grows along the entire elevation gradient, attains heights much greater than the TMCF trees and is also somewhat stunted in the main cloud immersion zone (Martin et al. Reference MARTIN, SHERMAN and FAHEY2007). Plastic species such as this could lend themselves to experimental investigations of the relative importance of the potential causal factors. For example, temperature and light can be varied independently in field experiments with potted plants, though it will be more difficult to manipulate direct contact of leaves with fog or canopy epiphyte loading.
Feedbacks and non-linearity in TMCF
Early concepts and observations in tropical montane forest ecology suggested the existence of spatial discontinuities in the composition and structure of vegetation along the altitudinal gradient (Holdridge Reference HOLDRIDGE1967). Such a pattern might arise either as a result of discontinuity in one or more key macro-environmental drivers (e.g. geological substrates, topography or climatic variables) or because of strong feedbacks between vegetation (or other biota) and micro-environmental factors. Although some detailed studies have discounted the existence of discontinuous variation in forest composition across the altitudinal gradient in tropical mountains (Lieberman et al. Reference LIEBERMAN, LIEBERMAN, PERALTA and HARTSHORN1996), some cases of sharp discontinuities involving the vegetation feedback switch have been documented. For example, Martin et al. (Reference MARTIN, SHERMAN and FAHEY2007) demonstrated that a sharp ecotone between species-rich TMCF and monodominant pine forest is maintained by vegetation effects on natural fire disturbance, together with the influence of frequent cloud immersion especially during the dry season (Martin & Fahey Reference MARTIN and FAHEY2014).
Distinctive TMCF structure, particularly in its most extreme form, also may depend upon feedback mechanisms in which vegetation traits and soil properties reinforce the limitation on the productivity and stature of the forest (Werner & Homeier Reference WERNER and HOMEIER2015). Recent syntheses conclude that nutrient limitation of TMCF productivity results not from lower amounts of limiting soil nutrients but by the inability of trees to access these nutrients (Benner et al. Reference BENNER, VITOUSEK, OSTERTAG, Bruijnzeel, Scatena and Hamilton2010, Roman et al. Reference ROMAN, SCATENA, BRUIJNZEEL, Bruijnzeel, Scatena and Hamilton2010). Some evidence supports a positive feedback mechanism whereby litter of lower chemical quality is produced in TMCF, suppressing efficient recycling of nutrients by microbial decomposition (Schuur et al. Reference SCHUUR, CHADWICK and MATSON2001, Werner & Homeier Reference WERNER and HOMEIER2015). The role of soil saturation on such a feedback also seems plausible: inhibition of root function by anaerobic soil could reduce water uptake (Weaver et al. Reference WEAVER, BYER and BRUCK1973) reinforcing soil saturation and contributing to low litter quality (Schuur Reference SCHUUR2001).
An additional vegetation-soil feedback that contributes to TMCF structure and function is the apparently high proportional allocation of C to root systems (Girardin et al. Reference GIRARDIN, MALHI, ARAGÃO, MAMANI, HUASCO, DURAND, FEELEY, RAPP, SILVA-ESPEJO, SILMAN, SALINAS and WHITTAKER2010, Kitayama & Aiba Reference KITAYAMA and AIBA2002, Moser et al. Reference MOSER, LEUSCHNER, HERTEL, GRAEFE, SOETHE and IOST2011), leaving less C to support above-ground growth and LAI. Again, a feedback involving soil saturation seems plausible. For example, the higher below-ground allocation could limit LAI, and thereby promote soil saturation by reducing AET. Also, observations of Leuschner et al. (Reference LEUSCHNER, MOSER, BERTSCH, RODERSTEIN and HERTEL2007) emphasize the much higher coarse-root biomass in high-elevation TMCF with frequently saturated soils; high allocation to coarse roots could contribute to overcoming structural stability problems in saturated soils (Coutts Reference COUTTS1983, Soethe et al. Reference SOETHE, LEHMANN and ENGELS2006).
In conclusion, recent research has contributed to an improved understanding of tropical montane cloud forest structure and function and its dependence on unique climatic conditions in the altitudinal zone where fog is persistent (Figure 1). However, additional research is needed to clarify several features of TMCF ecosystem dynamics: (1) The role of phylogeny vs. phenotype in shaping the characteristic traits of TMCF vegetation; (2) Photosynthetic physiology of TMCF foliage in fog-shrouded environments; (3) Below-ground carbon allocation and fine-root production and turnover in TMCF; (4) Factors leading to N vs. P limitation of TMCF productivity, including the role of mycorrhizal associations; (5) The role of saturated soils and low soil oxygen in TMCF ecosystems; (6) Causes of contrasting patterns of variation in TMCF structure in relation to topographic position among different locations; (7) Drivers of high diameter : height ratio of TMCF; (8) The role of high epiphyte abundance in regulating TMCF ecosystem dynamics.
Recognizing the great variation of vegetation structure and dynamics among TMCF locations, comparative research approaches across TMCF sites should help to provide new insights into the causal factors driving the behaviour of these remarkable forests.
ACKOWLEDGEMENTS
The authors appreciate the insightful comments and suggestions provided by P. Bellingham, R. Lawton, P. Martin and an anonymous reviewer.




