Introduction
The longtail butterfly ray, Gymnura poecilura (Shaw, Reference Shaw1804) is distributed widely in the Indo-West Pacific with confirmed reports extending from the south-west and east coast of India, Sri Lanka to Indonesia and China (Muktha et al., Reference Muktha, Akhilesh, Sandhya, Jasmin, Jishnudev and Kizhakudan2016). Reports of G. poecilura from the northern Arabian Sea and regions further west need confirmation as evidence points to the presence of a different species of Gymnura from that area (Muktha et al., Reference Muktha, Akhilesh, Sandhya, Jasmin, Jishnudev and Kizhakudan2016). Given the vulnerability of the species to fisheries, in particular to the trawl fleet, and the general lack of knowledge on its biology, it has been classified as Near Threatened by the IUCN (Bizzarro & White, Reference Bizzarro and White2006).
Elasmobranchs are a vulnerable marine group due to their slow growth rates, long life, low fecundity and late age of maturity (Stevens et al., Reference Stevens, Bonfil, Dulvy and Walker2000). Hence information on their life history is critical for assessing their vulnerabilities. The reproductive biology of G. poecilura has been studied to varying degrees in India and the Bay of Bengal. The first published work on the reproductive biology of this species is possibly that of Woodmason & Alcock (Reference Woodmason and Alcock1891) in which they described the embryos of G. micrura. However, G. micrura as used in the Indian sub-continent has been a misapplication as a synonym of G. poecilura with no connection to the original G. micrura (Bloch & Schneider, Reference Bloch and Schneider1801) from the Atlantic Ocean (James, Reference James1966; Muktha et al., Reference Muktha, Akhilesh, Sandhya, Jasmin, Jishnudev and Kizhakudan2016). Since then there have been reports by Alcock (Reference Alcock1892); Setna & Sarangdhar (Reference Setna and Sarangdhar1950) from Bombay waters; James (Reference James1966) from the Gulf of Mannar and Palk Bay; Devadoss (Reference Devadoss1998) from Chennai; and Raje (Reference Raje2003) and Raje & Zacharia (Reference Raje and Zacharia2009) from Mumbai waters. However, recent biological information is lacking for the species. None of these reports have dealt with diet of G. poecilura in detail.
Given the IUCN classification of the species, lack of recent information on reproduction and lack of detailed information on diet, this study was undertaken to provide up to date information on these aspects of Gymnura poecilura from the western Bay of Bengal which could be helpful for re-assessing its status.
Materials and methods
The study area covers the western Bay of Bengal along the east coast of India, which is fished extensively by trawlers operating from Visakhapatnam Fishing Harbour (17.6962°N 83.3009°E) (Figure 1). Nearly 579 trawlers operate from this fishing harbour, fishing to depths of 100 m (Muktha et al., Reference Muktha, Ghosh, Mini, Divipala, Behera, Loveson, Felix and Raju2018).

Fig. 1. Map showing study location (Visakhapatnam) along east coast of India bordering western Bay of Bengal, with bold arrows indicating the north and south limits of the regular trawling areas along the study area; map of India in inset.
Gymnura poecilura is landed regularly as a by-catch in trawl fisheries from Visakhapatnam Fishing Harbour. Specimens were collected weekly from December 2012 to December 2015. The collected specimens were brought to the laboratory where their disc widths (W D, mm), total weight (W, g) and sex were recorded. The animals were then dissected to check for reproductive status. For females the number, approximate diameter and colour of ovarian follicles were noted. The uterus was also examined for the presence of pups. If pups were present, their W D, W and sex were recorded. In males, since it was difficult to distinguish between a partially calcified clasper and a fully calcified clasper, rigidity of claspers was not used as a criterion for maturity; instead the condition of the testes was used (e.g. Conrath, Reference Conrath, Musick and Ramon2004; Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012).
The sex ratio of females to males was tested using the Pearson's χ2 goodness-of-fit test at 5% level of significance (Zar, Reference Zar1999). Mean size of males and females were tested for significant difference using the Student's t-test at 5% level of significance (Zar, Reference Zar1999). The size–weight relationship was assessed using the standard equation given by Le Cren (Reference Le Cren1951):
 $$W = a \times \lpar {W_{\rm D}} \rpar ^b$$
$$W = a \times \lpar {W_{\rm D}} \rpar ^b$$where a is the intercept (weight at zero width), b is the slope (change in weight per unit change in disc width). Size–weight relationships were fit separately for males and females and compared using Analysis of Covariance (ANCOVA) (Snedecor & Cochran, Reference Snedecor and Cochran1967) at 5% level of significance.
For males, three maturity stages (Figure 2) were assigned: (1) Immature – testis was small, without any lobed structure; (2) Maturing – testis was larger forming 15–20% of body cavity and with small lobes; and (3) Mature – testis was large, occupying >30% of body cavity, with fully formed lobes throughout. The spent stage (based on flaccid testis and the size of the specimen) was also included within the ‘Mature’ category. For females, five maturity stages (Figure 3) were assigned: (1) Immature – ovary was small, with a creamy appearance and no visible ovarian follicles; (2) Maturing – small, yolk-filled ovarian follicles of 1–2 mm diameter in the ovary and with a narrow uterus; (3) Mature – large yolk-filled ovarian follicles of 3–15 mm diameter in the ovary, uterus small, without pups; (4) Gravid – ovary with or without yolk-filled ovarian follicles, embryo(s) in the uterus; and (5) Spent recovering – with yolk-filled ovarian follicles in ovary, uterus large and flaccid but without embryos, indicating a recent release of embryos. The presence of small ovarian follicles in the ‘Spent’ category indicated a recovering stage after an earlier release of pups.

Fig. 2. Left functional testis of Gymnura poecilura (A) Immature – testis immature, without lobes, male of W D 320 mm (B) Maturing – testis maturing with small lobes visible, male of W D 411 mm and (C) Mature – testis with clear lobes, male of W D was 470 mm. All specimens were collected at Visakhapatnam, along east coast of India bordering western Bay of Bengal. Rudimentary non-functional right testis is also visible.

Fig. 3. Reproductive system of female Gymnura poecilura. (A) Immature – ovary is immature, without any visible ovarian follicles, female of W D 328 mm. (B) Maturing – ovary maturing, with small ovarian follicles visible, female of W D 478 mm. (C) Mature – large ovarian follicles visible, female of W D 554 mm. (D) Gravid – embryos found within uterus, large ovarian follicles seen in ovary, female of W D 874 mm; (E) Gravid – Same female with all six embryos displayed, three embryos from each uterus; (F) Spent recovering – Large number of ovarian follicles visible, no embryo in uterus, female of W D 699 mm. All specimens were collected at Visakhapatnam, along east coast of India bordering western Bay of Bengal. Only the left ovary is functional, however a rudimentary right ovary is also present. Both uteri are functional.
Size (W D) at 50% maturity (W D50) was estimated using the procedure outlined by King (Reference King1995). The proportion of mature fish in each 100 mm size class was fit to the following logistic equation:
 $$P = \displaystyle{1 \over {\lpar {1 + {\rm e}^{{-}r\lpar {W_{\rm D}-W_{{\rm D}50}} \rpar}} \rpar}}$$
$$P = \displaystyle{1 \over {\lpar {1 + {\rm e}^{{-}r\lpar {W_{\rm D}-W_{{\rm D}50}} \rpar}} \rpar}}$$where, P is the proportion of mature fish in each length class, r is the slope of the logistic curve, W D is the disc width of specimen (mm) and W D50 is the disc width which corresponds to a proportion of 0.5 in reproductive condition. For females, mature specimens were those in stages ‘Mature’, ‘Gravid’ and ‘Spent’.
Size at birth was estimated based on the size of the largest pup seen within a uterus and the smallest neonatal (free swimming) individual. The parturition season was estimated by studying the monthly size-frequency of neonates and near-term embryos.
The cardiac stomach of each specimen was dissected out and the fullness of stomach was checked to assess the feeding intensity. The degree of fullness of stomach was classified as empty, trace, quarter, half, three-quarters, full and gorged (Pillay, Reference Pillay1952). The stomach was then cut open and contents were weighed (total weight) and the weights and numbers of identified prey taxa recorded. The Index of Relative Importance (IRI) (Pinkas et al., Reference Pinkas, Oliphant and Iverson1971) was calculated from the data as: IRI = (%N + %W) × % F where, %N is the percentage of the particular prey item in total number of prey items, %W is the percentage of the particular prey item in total weight of prey items and %F is the percentage frequency of occurrence of each prey item among total prey items. The IRI values were then converted to percentage IRI values for enabling easier comparisons across prey items (Cortés, Reference Cortés1997). IRI was estimated for male and female fish separately and tested for significant difference using a χ2 test at a 5% level of significance.
The feeding strategy was estimated using the method outlined by Amundsen et al. (Reference Amundsen, Gabler and Staldvik1996). In this method a prey-specific abundance (Pi) is estimated which is defined as the percentage of a prey taxon in relation to all prey items in only those predators in which the actual prey occurs. The prey-specific abundance is estimated as:
 $$P_i = \displaystyle{{\mathop \sum \nolimits S_i} \over {\mathop \sum \nolimits S_t}} \times 100$$
$$P_i = \displaystyle{{\mathop \sum \nolimits S_i} \over {\mathop \sum \nolimits S_t}} \times 100$$where, Si is the stomach content (weight) comprised of prey i and St is the total stomach content in only those predators with prey i in their stomach. The prey-specific abundance was plotted against frequency of occurrence and based on the distribution of prey points; the feeding strategy of the predator was obtained (Amundsen et al., Reference Amundsen, Gabler and Staldvik1996). The vertical axis of the plot indicates feeding strategy with prey points falling in the upper part of the plot indicating specialized feeders and those in the bottom part indicating generalized feeders. The diagonal from lower left to upper right indicates importance of prey items. The diagonal from upper left to lower right indicates the niche width contributions (Amundsen et al., Reference Amundsen, Gabler and Staldvik1996). The dietary niche breadth of G. poecilura was estimated using the Levins Standardized Niche Breadth index (Levins, Reference Levins1968; Hurlbert, Reference Hurlbert1978) which is estimated from the Levins Niche Breadth Index as follows:
 $$B = \displaystyle{1 \over {\mathop \sum p_j^2 }}; \; B_{\rm A} = \displaystyle{{B-1} \over {n-1}}\comma $$
$$B = \displaystyle{1 \over {\mathop \sum p_j^2 }}; \; B_{\rm A} = \displaystyle{{B-1} \over {n-1}}\comma $$where B = Levins Niche Breadth, Pj = proportion of food item j, B A = Levins Standardized Niche Breadth and n = number of food types. B A ranges from 0 to 1, with values close to 0 indicating a specialist feeder and values close to 1 indicating a generalist feeder.
Results
The study covered 630 specimens (377 females and 253 males) of G. poecilura. The overall sex ratio (males:females) was skewed towards females at 1:1.49 (χ2 = 861.5, df = 1, P < 0.05). The sex ratio for the smallest size class was 1:1 (<300 mm; P > 0.05). Males outnumbered females both in the 300–400 mm and 400–500 mm size classes (sex ratio <1; P < 0.05) (Table 1). It was only in the 500–600 mm size class that females outnumbered males (sex ratio >1, P < 0.05). Males were absent in the ≥600 mm W D size class.
Table 1. Size-based sex ratio of Gymnura poecilura collected at Visakhapatnam during December 2012–December 2015

The average W D of females was 576.7 mm (range = 190–920 mm) and for males it was 416.3 mm (range = 230–550 mm). A Student's t-test indicated significant differences between mean W D of males and females (P < 0.05), indicating sexual dimorphism in the size. The maximum W D of males was only 60% of that of the females. The average weight of females was 1721.8 g (SE 73.4) and that of males was only 544 g (SE 22.8).
The W D – weight relationship for males was estimated as:
 $$W = 0.0055 \times \lpar {W_{\rm D}} \rpar ^{3.12}$$
$$W = 0.0055 \times \lpar {W_{\rm D}} \rpar ^{3.12}$$and that for females was estimated as:
 $$W = 0.0063 \times \lpar {W_{\rm D}} \rpar ^{3.08}$$
$$W = 0.0063 \times \lpar {W_{\rm D}} \rpar ^{3.08}$$The slope values of 3.12 (males) and 3.08 (females) showed a significant difference from the hypothesized value of 3 in a Student's t-test. The ANCOVA of the two W D – weight relationships indicated a significant difference between the two relationships, indicating marked differences (P < 0.05) in the growth of males and females.
Reproductive biology
The biological studies were conducted on 626 specimens of G. poecilura (373 females and 253 males). Gymnura poecilura has only a single functional testis on the left side of the animal and a rudimentary testis on the right side (Figure 2), with females having a single functional ovary on the left side of the animal, rudimentary ovary on the right side and two functional uteri (Figure 3). The ovary had yolk-filled follicles ranging in diameter from 0.5–20 mm; ovarian follicles were not visible in immature females. Embryos were found either singly in a uterus or more than one in a uterus or in both uteri. Embryos were placed parallel to the length of the mother's body with the pectoral fins folded under the body. The uterine walls had a dense layer of trophonemata for providing nutrition to the embryos. The colour of the uterine wall and trophonemata changed from light pink (without uterine embryos) to dark red (post-partum).
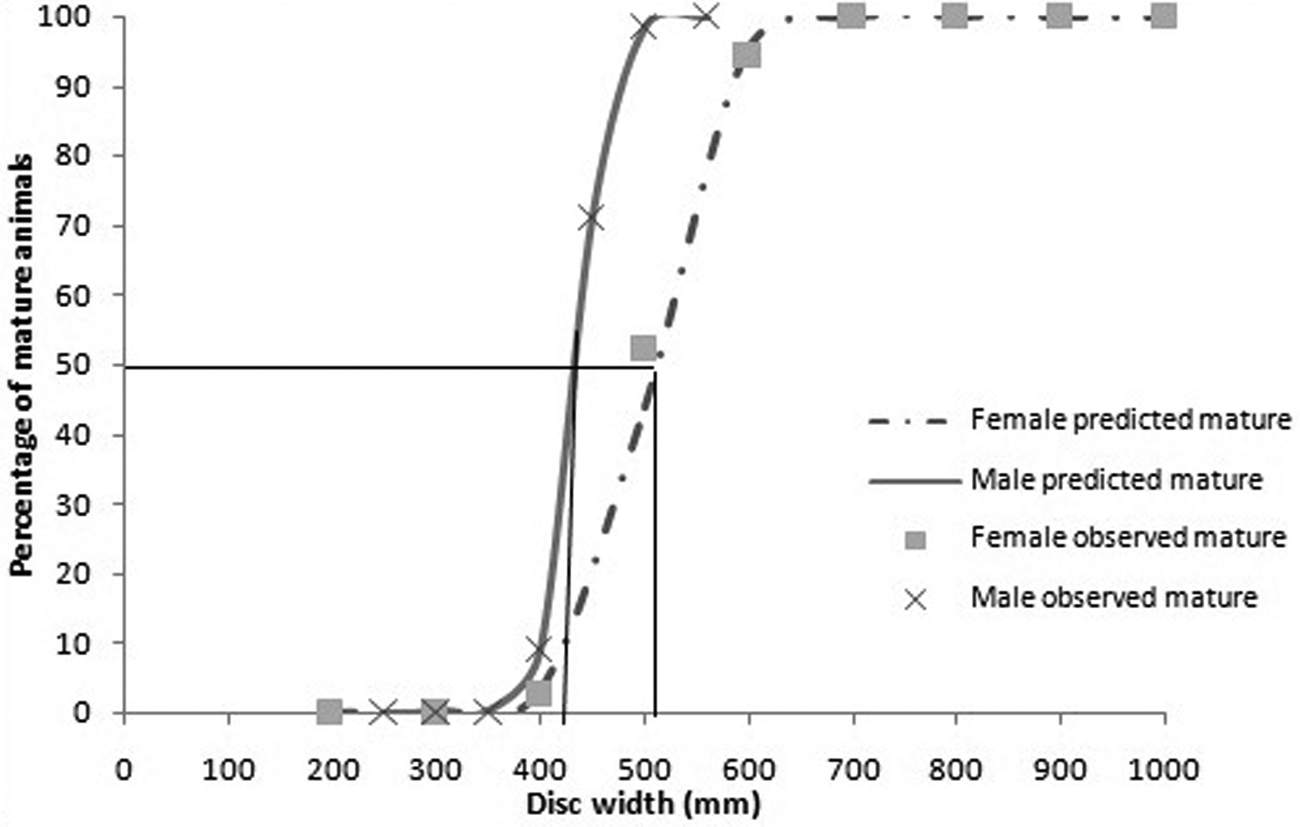
Of the 373 females with information on maturity stages, 78.8% were mature. Of the 253 males with information on maturity stages, 54.9% were mature. The size at 50% maturity for females was estimated to be 506.5 mm W D and that for males was estimated to be 435.9 mm W D (Figure 4). The smallest embryo measured 30 mm W D and 1 g in weight, while the largest measured 220 mm W D and 115 g in weight. The latter was seen in a female of size 655 mm W D and weight 2782 g in February. The smallest free-swimming individual observed was 190 mm W D with a weight of 170 g. Thus the size at birth was estimated to be between 190–220 mm W D.

Fig. 4. Relationship between disc width and maturity of female and male Gymnura poecilura. The size at 50% maturity (W D 50) of female and male Gymnura poecilura was estimated by fitting a logistic curve. Males mature at smaller sizes than females.
Ovarian fecundity was not estimated; however it was observed that the number of eggs decreased with increasing size within the ovary. The number of embryos within a single female ranged from one to eight (for both uteri combined). Only 14.58% of the gravid females had a single embryo, all the others had more than one embryo in the uterus. The mean uterine fecundity was 3.3 (SD 1.8). The number of embryos showed a positive correlation with W D of females (r = 0.67, P < 0.05). The maximum number of eight embryos was observed in a female measuring 920 mm disc width weighing 7.3 kg. In the smallest gravid female (610 mm W D) only a single embryo was observed. Of the 170 embryos seen in the study, 71 were sexed (females = 45 and males = 26) and the sex ratio of the pups was 1:1.73 (M:F) which was significantly different from the 1:1 ratio (χ2 = 5.08, df = 1, P < 0.05).
Embryo size was examined by month (Figure 5) and the highest mean size occurred in January (187.5 mm). However, there were large ranges in embryo size in most months (Figure 5). The largest intra-month variation was seen in April. In April 2015, two gravid females were obtained of 758 and 728 mm W D respectively, with the mean size of their embryos being 204.5 and 70 mm, respectively. Intra-uterine embryos of sizes 190 mm and more were seen in January, February, April, September, November and December. Neonates were occasionally observed at the sampling location during January, March, June, July and August (Table 2), but some small specimens may have been discarded by fishermen. Gravid females and mature males were obtained in samples throughout the year (Figure 6).

Fig. 5. Embryo disc width by month of Gymnura poecilura; the solid black point is the average embryo disc width of the particular month. Highest average value was seen in January and intra-month variations of embryo disc width were high.

Fig. 6. Monthly occurrence of mature males and mature/gravid females of Gymnura poecilura. Mature specimens were found in all months of the study period.
Table 2. Occurrence of neonates of Gymnura poecilura at Visakhapatnam during December 2012–December 2015

Whenever a female was gravid, invariably the ovary had yolk-filled follicles in it. Even when embryos within the uterus were as small as 30 mm W D (~1 g body weight), yolk-filled follicles of 0.5 mm diameter were visible in the ovary. In individuals with larger embryos (150 mm W D, ~40 g body weight), the largest ovarian follicles were ~20 mm. By the time embryos reached 220 mm W D (~120 g body weight), ovarian follicles reached the largest size, ready to be ovulated. The majority of the spent recovering females had well-developed and/or developing ovarian follicles.
Diet
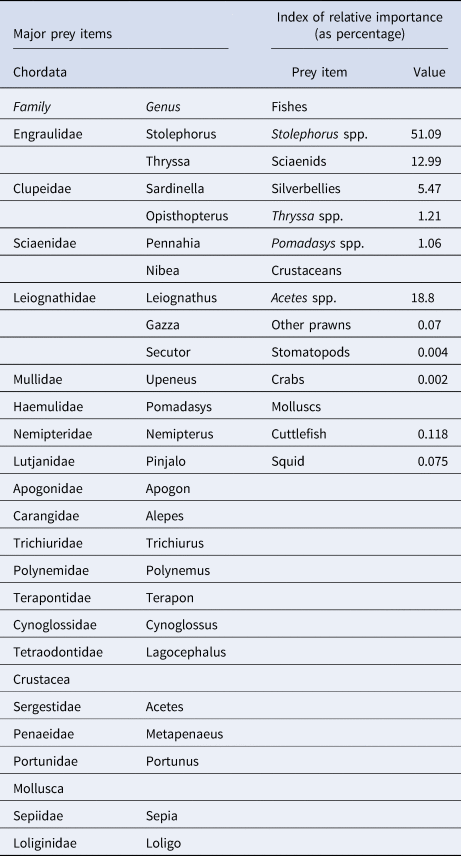
Of the 673 stomachs examined, 53.05% were empty. Teleosts, crustaceans and molluscs were observed in the diet of G. poecilura (Table 3). Teleosts were mainly represented by the families Engraulidae, Clupeidae, Sciaenidae, Leiognathidae and Mullidae. Crustaceans were mainly represented by the family Sergestidae. Molluscs were only represented by cephalopods.
Table 3. Major prey items and Index of Relative Importance of major prey items of Gymnura poecilura

Stolephorus spp. had the highest IRI value, followed by Acetes spp. and croakers (Table 3). These three groups combined formed 83%IRI. Fish accounted for 80.9%IRI, followed by crustaceans (18.9%IRI) and the rest by molluscs. A χ2 test for difference between sexes regarding diet (IRI) indicated no significant difference at the 5% level. Only semi-digested fish showed significant differences between the two sexes (P = 0.023).
The size-based analysis of the diet showed increasing feeding intensity with increase in body size. Empty stomachs dominated in the lower size groups, but the prevalence of non-empty and empty stomachs was equal in the higher size groups (Table 4).
Table 4. Feeding intensity of Gymnura poecilura (all values in percentage) across size groups (W D in mm)

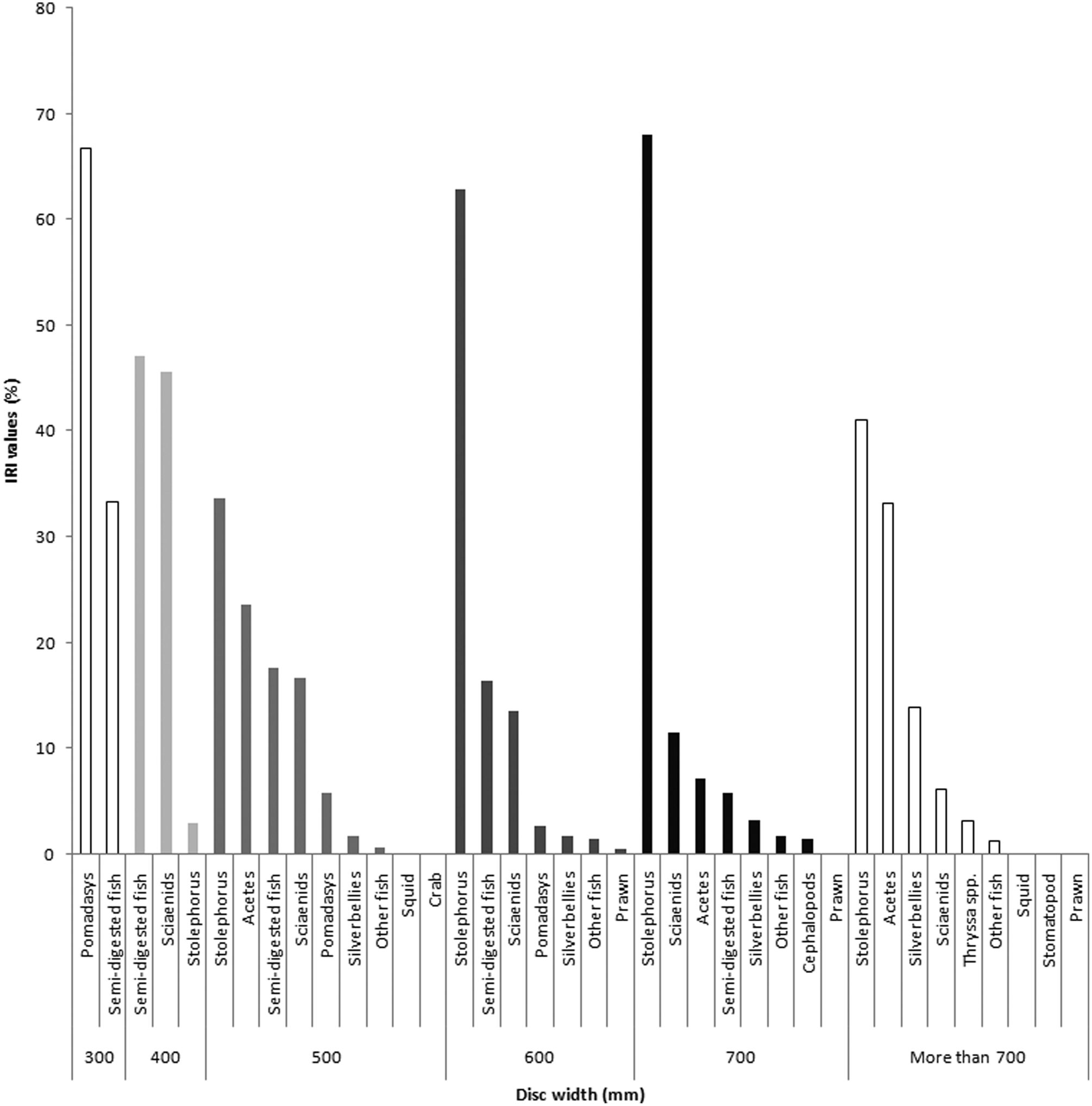
A change in feeding habit was observed in G. poecilura with increasing body size (Figure 7). Gymnura poecilura up to 400 mm W D were exclusively piscivorous feeding mainly on croakers, Pomadasys, Stolephorus, soles and Opisthopterus tardoore. Beyond 400 mm W D rays fed on a variety of food items including fish, crab, prawns, cuttlefish and squid. Despite the different feeding habits, IRI values indicated that fish was the most preferred prey taxon, followed by Acetes (Table 3). Stolephorus spp. and Acetes spp. were the two diet items that were observed in large numbers in a single stomach with their incidence in a single stomach also increasing with size of G. poecilura. The average number of Stolephorus spp. in each stomach ranged from 3 in the smaller (300–400 mm W D) specimens to 6.5 in the largest (>700 mm W D) specimens. The average number of Acetes spp. ranged from 24.4 in medium-sized (400–500 mm W D) individuals to 80.2 in the largest (>700 mm W D) specimens.

Fig. 7. Size-based changes in prey items of Gymnura poecilura; the animals moved from an exclusive piscivorous diet to a broader diet including shellfish as they grew in size.
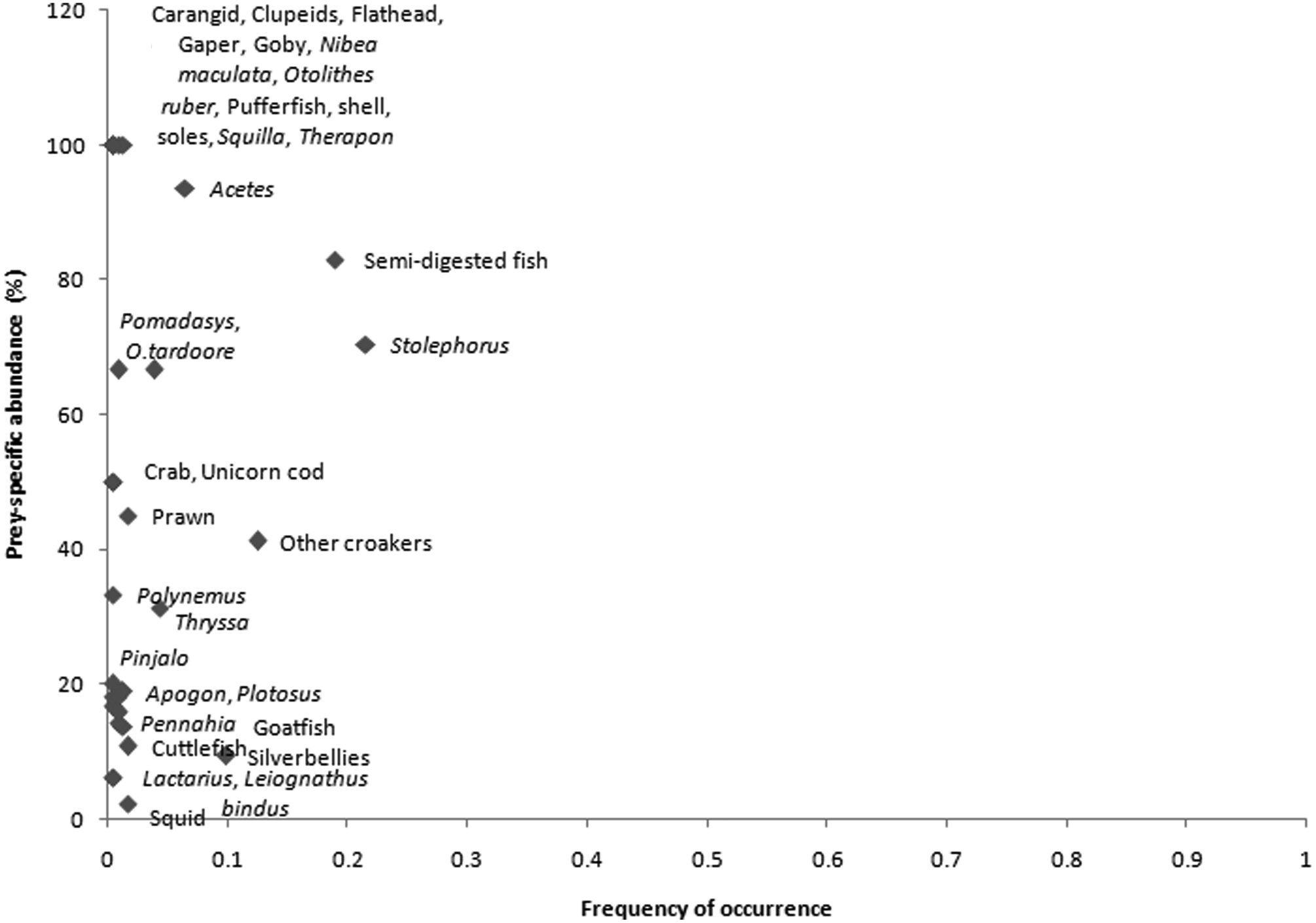
The feeding strategy of G. poecilura is indicated in Figure 8. All the prey points fell below the diagonal from upper left to lower right corner. Furthermore, all the prey points fell on the left hand side of the horizontal axis indicating low frequencies of occurrence. A few prey points, namely Leiognathus bindus, Lactarius lactarius, Alepes spp., Plotosus spp. and Pinjalo spp., fell in the bottom left half of the plot area indicating low prey-specific abundance as well and were classified as occasional prey for G. poecilura. The rest of the prey points fell along the left panel of the plot area ranging from moderate to high prey specific abundance indicating a specialized feeding strategy. The cluster of prey points in the upper left corner of the plot (Carangids, Clupeids, croakers, flathead, gapers, gobies, pufferfish, grunters, soles, stomatopods and Acetes spp.) indicated that there was high between-phenotype contribution to niche width (Amundsen et al., Reference Amundsen, Gabler and Staldvik1996) i.e. different individuals had specialized to feed on different prey items, however each prey item was consumed only by a few individuals. The Levins Standardized Niche Breadth was estimated to be 0.038.

Fig. 8. The feeding strategy plot of Gymnura poecilura; the plot indicated a specialized feeding strategy for the species.
Discussion
This study provided updated information on the reproductive biology of G. poecilura and detailed information on diet of the species from the western Bay of Bengal. The largest specimen sampled (920 mm) is possibly the largest reported size for G. poecilura, with James (Reference James1966) reporting a maximum W D of 915 mm. Reports of larger sizes (>1000 mm W D) for the species from the Arabian Sea (Sommer et al., Reference Sommer, Schneider and Poutiers1996; Raje & Zacharia, Reference Raje and Zacharia2009) must be re-confirmed due to the presence of another closely related species, G. cf. poecilura (Muktha et al., Reference Muktha, Akhilesh, Sandhya, Jasmin, Jishnudev and Kizhakudan2016) from the same region. Other congeners of the species in the Indo-Pacific, namely Gymnura australis 860 mm (Jacobsen & Bennett, Reference Jacobsen and Bennett2009) and Gymnura zonura 1060 mm W D (White et al., Reference White, Last, Stevens, Yearsley, Fahmi and Dharmadi2006) have similar maximum reported sizes.
The W D – weight relationship indicated allometric growth similar to that reported from other batoid species (Basusta et al., Reference Basusta, Basusta, Sulikowski, Driggers, Demirhan and Cicek2012; Naderi et al., Reference Naderi, Zare and Azvar2013). The only report on W D – weight relationship of G. poecilura (G. cf. poecilura) was by Naderi et al. (Reference Naderi, Zare and Azvar2013) from Iran. They too reported allometric growth in the species. Values of b greater than 3 for both male and female G. poecilura, indicated that body width increased more than weight as the ray grew (Froese, Reference Froese2006).
The overall sex ratio reported in this study was 1.49 with females outnumbering males, although the sex ratio varied across size classes. Skewed sex ratios favouring female Gymnura have been reported from India (James, Reference James1966; Raje & Zacharia, Reference Raje and Zacharia2009) and Indonesia (White & Dharmadi, Reference White and Dharmadi2007). Variation in sex ratios across size classes has been reported in G. micrura (Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012) while Capapé et al. (Reference Capapé, Zaouali, Tomasini and Bouchereau1992) reported a male-dominated sex ratio for G. altavela. The sex ratio could be influenced by the fishery (location and gear) or species aggregation (sex-based and spatial) if any, which would need further detailed studies for confirmation. The sex ratio favouring females in our study was primarily due to the absence of male fish at sizes over 550 mm W D.
Gymnura poecilura females grew larger than their male counterparts and matured at sizes larger than males. This pattern of sex-based size difference has been reported from a number of other Gymnurid species (and other elasmobranchs) indicating that this is a common feature in butterfly rays (Capapé et al., Reference Capapé, Zaouali, Tomasini and Bouchereau1992; Raje, Reference Raje2003; Sulikowski et al., Reference Sulikowski, Elzey, Kneebone, Jurek, Huntting Howell and Tsang2007; White & Dharmadi, Reference White and Dharmadi2007; Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009; Smith et al., Reference Smith, Bizzarro, Richards, Nielsen, Márquez- Flarías and Shivji2009; Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012). Sex-based differences in body size are an evolutionary adaptation of live bearers to increase their reproductive potential (Cortés, Reference Cortés2000). Females grow to larger sizes to support larger number of embryos. The positive correlation of female body size with the number of embryos in the uterus provides further proof of this theory. This trend has also been reported in G. australis (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009) and G. micrura (Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012).
The size at maturity for female and male G. poecilura in the western Bay of Bengal was estimated as 506.5 mm W D and 435.9 mm W D, respectively. James (Reference James1966) reported that female G. poecilura in the Gulf of Mannar and Palk Bay produced young from sizes of 410 mm W D. The reported size at maturity of G. poecilura (720 mm males; 780 mm females) and for G. japonica (440 mm males; 520 mm females) by Raje & Zacharia (Reference Raje and Zacharia2009) could possibly relate to G. cf. poecilura and hence are not directly comparable. All other studies on gymnurids have indicated that females mature at larger sizes than males (Capapé et al., Reference Capapé, Zaouali, Tomasini and Bouchereau1992; Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009; Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012; Salazar et al., Reference Salazar, Perez-Jimenez, Mendez-Loeza and Mendoza-Carranza2014; Burgos-Vázquez et al., Reference Burgos-Vázquez, Galván-Magaña, Carrera-Fernández, Ochoa-Báez and Oddone2019).
Gymnura poecilura females matured at 55.06% of maximum size (and males 79.25%). Estimating this percentage from published literature on other gymnurids indicates that females mature from 47.5% in G. australis (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009) to 71.6% in G. altavela (Alkusairy et al., Reference Alkusairy, Ali, Saad, Reynaud and Capape2014), whilst in males this ranged from 65.6% in G. australis (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009) to 86.3% in G. altavela (Alkusairy et al., Reference Alkusairy, Ali, Saad, Reynaud and Capape2014). This is unlike the general trend seen in sharks as indicated by Cortés (Reference Cortés2000) who found that ratio of length at maturity to maximum length was similar for males and females. Thus, at least in gymnurids, it appears that gonadal maturation in males sets in only after a major portion of somatic growth is attained.
The uterine fecundity of G. poecilura ranged from one to eight (subject to the possibility of some embryos or pups being lost on capture) which is similar to that reported elsewhere for this species, and for other gymnurids. James (Reference James1966) reported that G. poecilura in Gulf of Mannar had up to seven pups. Raje & Zacharia (Reference Raje and Zacharia2009) reported a lower number of pups (three per female) for G. japonica (possibly G. cf. poecilura) and G. poecilura (G. cf. poecilura). The maximum number of embryos in other gymnurids is three in G. japonica, four in G. zonura, six in G australis, 11 in G. micrura, nine in G. natalensis and seven in G. altavela (Bigelow & Schroeder, Reference Bigelow, Schroeder, Tee-Van, Breder, Hildebrand, Parr and Schroeder1953; Daiber & Booth, Reference Daiber and Booth1960; Wallace, Reference Wallace1967; White & Dharmadi, Reference White and Dharmadi2007; Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009; Salazar et al., Reference Salazar, Perez-Jimenez, Mendez-Loeza and Mendoza-Carranza2014).
Mature males and females (including gravid females) of G. poecilura were seen throughout the year. The presence of near-term embryos (190–220 mm W D) and neonates in multiple months indicated the lack of a specific pupping season for G. poecilura in the study area. The large intra-monthly variation in embryo size seen in the study indicated that mating was not restricted to a specific time during the year. The continuous development of follicles in the ovary and embryos in the uteri indicated that G. poecilura likely has an almost continuous reproductive cycle throughout the year. This is similar to the spawning season reported by James (Reference James1966), and for G. cf. micrura (possibly misidentified G. cf. poecilura) by Raje (Reference Raje2003). Prolonged breeding seasons are thought to be quite common in rays in tropical oceans in contrast to rays in temperate and subtropical waters (Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012). The presence of uteri without pups and with developing follicles in the ovary indicated a resting period in between spawning events for a single individual. This has also been indicated in G. micrura by Yokota et al. (Reference Yokota, Goitein, Gianeti and Lessa2012).
The size at birth for G. poecilura was estimated to be between 190 and 220 mm W D along western Bay of Bengal. James (Reference James1966) suggested a similar size at birth (237–256 mm W D) for G. poecilura from the Gulf of Mannar and Palk Bay. The size at birth for G. australis was estimated to be 220–260 mm W D (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009). Similar sizes have been reported for G. zonura (140–280 mm W D) and G. japonica (200–210 mm W D) (White & Dharmadi, Reference White and Dharmadi2007). Size at birth for G. micrura was estimated to be 135–175 mm from Brazil (Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2012). Offspring size at birth is positively correlated with maternal body size (Cortés, Reference Cortés2000). Gymnura poecilura too pointed to this trend with similar size at birth for its offspring from other parts of the east coast of India as well as its similar sized congeners in other parts of the world, namely G. australis and G. zonura.
The IRI indicated that the dominant diet content of G. poecilura were Stolephorus and Acetes. Overall, fish accounted for 80.9%IRI, confirming the piscivorous feeding habits. Other than Stolephorus, the majority of the fish in its diet were meso-pelagic and benthic species. The piscivorous feeding nature is similar to that reported for congeners, including G. cf. micrura (G. cf. poecilura) (Raje, Reference Raje2003), G. micrura (Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2013) and G. australis (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009). The %Alimentary Index of fish in G. micrura was 99.5% (Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2013) and teleosts (99.8%IRI) dominated the diet of G. australis (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009). Similarly, fish formed 90% of the diet of G. marmorata (Bizzarro, Reference Bizzarro2005) and was the dominant prey taxa for G. altavela (Yemışken et al., Reference Yemışken, Forero, Megalofonou, Eryilmaz and Navarro2018).
The feeding strategy analysis indicated that individuals in the population were specialized to feed on specific prey items; however some prey items were taken infrequently also. The individuals within the population were specialized in feeding on fish (Carangids, Clupeids, croakers, flathead, gapers, gobies, pufferfish, grunters, soles), stomatopods and Acetes spp. These diet items scored high on prey-specific abundance but the frequency of occurrence was low, indicating that there was high variation between individuals in the studied population (Amundsen et al., Reference Amundsen, Gabler and Staldvik1996). The Levins niche breadth analysis also indicated a highly specialized feeding behaviour. A specialized feeding behaviour for G. micrura has been reported by Yokota et al. (Reference Yokota, Goitein, Gianeti and Lessa2013). In their study from Brazil, they found that G. micrura specialized on feeding on Pomadasys corvinaeformis with some individuals apparently specialized for feeding on a few other items as well. Another congener, G. australis, was described as a fish-feeding specialist by Jacobsen et al. (Reference Jacobsen, Johnson and Bennett2009).
The prevalence of empty stomachs (53.05%) in G. poecilura is similar to that seen in congeners, namely, G. altavela (88%, Daiber & Booth, Reference Daiber and Booth1960), G. marmorata (67%, Daiber & Booth, Reference Daiber and Booth1960), G. cf. micrura (G. cf. poecilura) (55%, Raje, Reference Raje2003), G. australis (66.8%, Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009) and G. micrura (38.5%, Yokota et al., Reference Yokota, Goitein, Gianeti and Lessa2013). The high proportion of empty stomachs indicates an intermittent feeding pattern for the family as a whole (Jacobsen et al., Reference Jacobsen, Johnson and Bennett2009). Gymnura poecilura seems to be no different from its congeners in having an intermittent feeding behaviour. However, the prevalence of empty stomachs reduced as the size of the animal grew, indicating that intermittent feeding behaviour changed as the animal grew. In smaller sizes, the animal tended to eat as opportunity prevailed, becoming more active predators when larger. This is quite logical since it is expected that as the rays grow, their swimming ability and physical capacity to hunt for more types of prey items would also increase. Several reports indicate that Gymnurids actively prey on animals, stunning the prey with their pectoral fins (Schreiber, Reference Schreiber1997; Smale et al., Reference Smale, Sauer and Roberts2001). However, the gorged condition of stomachs containing Acetes spp. indicated a strategy of feeding on aggregating prey, possibly swimming through the shrimp shoal with open mouths to feed on as many numbers as possible or even trapping the prey between their bodies and the seafloor for easy feeding.
Thus, from the diet analysis, it appears that G. poecilura has different feeding behaviours. Smaller individuals (≤400 mm W D) feed mainly on fish, ingesting medium-sized prey in fewer numbers. These individuals feed intermittently and probably opportunistically; it is also possible that these individuals have a smaller capacity to hunt for diverse prey items. Larger individuals on the other hand either feed on large-sized fish in few numbers or feed on small-sized fish/shrimp in large numbers. The latter behaviour, when predating on Stolephorus spp. and Acetes spp., presumably involves the individuals feeding on high densities of prey. However, this needs further confirmation since it could also have fed on these prey items within the trawl net.
This study provided updated information on the reproductive biology and diet of G. poecilura which will be helpful in a more accurate assessment of its status over its distribution range.
Acknowledgements
This study was part of the first author's PhD dissertation. The authors are grateful to Dr A Gopalakrishnan, Director, ICAR-CMFRI, Dr PU Zacharia, Head, Demersal Fisheries Division, ICAR-CMFRI and Dr Shubhadeep Ghosh, Scientist-in-Charge, Visakhapatnam RC of ICAR-CMFRI for encouragement and support during the study. MM acknowledges the help provided by Dr Akhilesh KV, Dr Eldho Varghese, Dr Manas HM, Dr Jasmin F, Dr Madhumita Das, Dr Pralaya Ranjan Behera, Dr Indira Divipala, Mr O Ch Venkateswarulu, Mr MA Jishnudev, Mr P Bhaskar Rao, Mr V Venkateswarulu, Mr P Venkatesh and Mr G Hari during the study. The authors are extremely grateful to two anonymous reviewers and the editor for critically reviewing the manuscript and improving it significantly.
Financial support
This study was fully funded by the Indian Council of Agricultural Research through Central Marine Fisheries Research Institute Project DEM/ELAS/5.