



Introduction
The Hirnantian (latest Ordovician) was a special time in Earth's history, marked by the second largest extinction of the Phanerozoic (Sepkoski, Reference Sepkoski and Walliser1996) and the onset of a major glaciation associated with the extinctions (e.g., Brenchley et al., Reference Brenchley, Marshall, Carden, Robertson, Long, Meidla, Hints and Anderson1994; Sheehan, Reference Sheehan2001). The Hirnantian glaciation, centered on the northern margin of Gondwana, led to dramatic sea level changes and, consequently, to changes in oceanic circulation, water chemistry, oxygen concentrations, and nutrients availability, causing severe disturbances of global ecosystems (Sheehan, Reference Sheehan2001). Traditionally, two major extinction pulses are recognized during the Hirnantian (Brenchley et al., Reference Brenchley, Carden and Marshall1995; Harper et al., Reference Harper, Hammarlund and Rasmussen2014). However, some authors have recognized a prolonged three-phased interval of biodiversity loss and suggested that the extinctions could have started earlier (e.g., Rasmussen et al., Reference Rasmussen, Kröger, Nielsen and Colmenar2019). Others (e.g., Wang et al., Reference Wang, Zhan and Percival2019) defended a single-pulse extinction, followed by different recovery phases through the Hirnantian and early Silurian.
Regardless of its causes and pulses, brachiopod-dominated communities originated and thrived worldwide in shallow-marine environments at intermediate to low latitudes after the first extinction pulse. These communities characterize the Hirnantian deposits globally, known as the Hirnantia Fauna (Temple, Reference Temple1965). The appearance, lifespan, composition, and geographical distribution of these survivor communities are essential to understand what triggered the extinction and how it happened. Recently, Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a) analyzed all the reported occurrences of brachiopod Hirnantian fossil assemblages and recognized two successive evolutionary faunas related to the first and second phases of the extinction: the Hirnantia Fauna sensu stricto, a widespread and diachronous community related to the glacial acme in the early-middle Hirnantian; and the newly named Edgewood-Cathay Fauna, adapted to the warmer waters of the late Hirnantian and early Rhuddanian (Silurian) post-glacial stage, occupying the low-latitude biotopes left vacant by the disappearance of the Hirnantia Fauna.
In Belgium, the occurrence of Hirnantian sequences was first postulated by Billiaert (Reference Billiaert2000) and Herbosch et al. (Reference Herbosch, Verniers, Debacker, Billiaert, de Schepper and Belmans2002) based on the presence of coarse terrigenous beds in the Génicot Formation of the Central Condroz Inlier, which was interpreted as the sedimentological expression of the Hirnantian glaciation sea level drop (Vanmeirhaeghe and Verniers, Reference Vanmeirhaeghe and Verniers2004). Shortly after, Vanmeirhaeghe (Reference Vanmeirhaeghe2006a) reassigned the Génicot Formation to the Aeronian (Silurian). However, based on its stratigraphical position and sedimentological characteristics, he recognized another probable Hirnantian succession in the Central Condroz Inlier: the uppermost part of the Fosses Formation, which he defined as the Tihange Member (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a). Mortier (Reference Mortier2014), in his PhD thesis, confirmed the Hirnantian age for the upper part of the Tihange Member based on brachiopods.
In this paper, we describe, illustrate, and discuss the first macrofossiliferous Hirnantia Fauna assemblage from Belgium, from the Tihange Member at Tihange (Huy). A systematic paleontology section is provided for all the identified groups. The age and depositional settings of the upper member of the Fosses Formation are herein discussed based on the biostratigraphy and the paleobiogeographical affinities of this new assemblage, framed within the most recent perspectives on the latest Ordovician Hirnantian brachiopod faunas (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a).
Geographical and geological setting
Location of the study area
The studied material is from central-eastern Belgium, geologically located in the eastern part of the Condroz Inlier, in two sections of the Tihange Member, in the village of Tihange (Huy), between Namur and Liège (Fig. 1.1, 1.2).
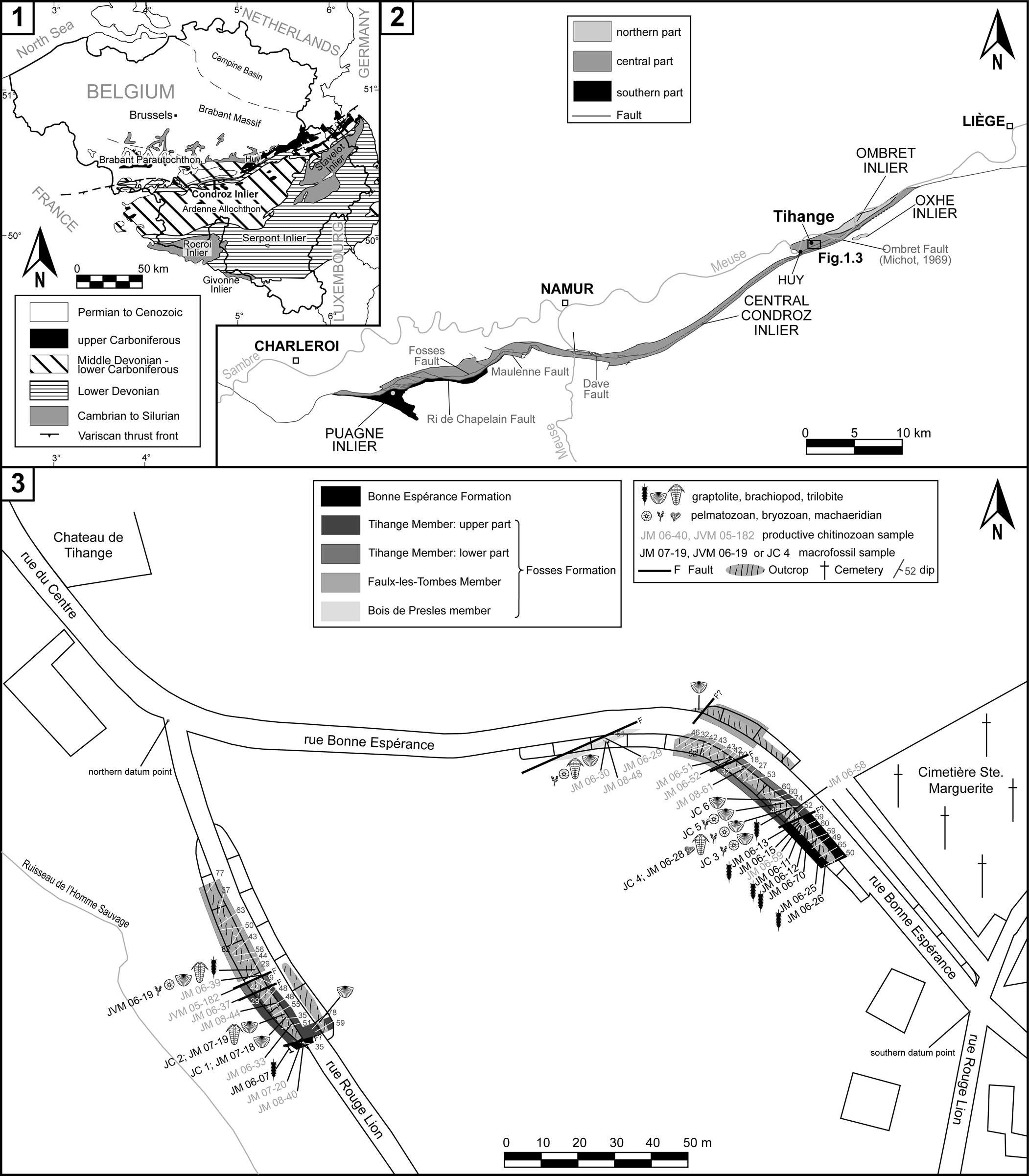
Figure 1. Geographical and geological setting of the Condroz Inlier and the studied sections. (1) Simplified geological map of Belgium (adapted from Fielitz and Mansy, Reference Fielitz, Mansy, Sintubin, Vandycke and Camelbeeck1999; Mortier, Reference Mortier2014); (2) location of the Tihange area and the main outcrop localities of the Condroz Inlier (map compiled from Michot, Reference Michot1932a, Reference Michotb, Reference Michot1934, Reference Michot1944, Reference Michot1969; Vandevelde, Reference Vandevelde1976; Maes et al., Reference Maes, Rickards, Rombouts and Vandevelde1978; and modified after Mortier, Reference Mortier2014); (3) overview of the sections of rue Bonne Espérance and rue Rouge Lion, with exact positions of the fossiliferous levels.
The studied sections are exposed on the ‘Rue Bonne Espérance’ (50°31′35.6″N, 5°15′42.7″E) and the ‘Rue Rouge Lion’ (50°31′33.2″N, 5°15′37.1″E; Figs. 2, 3). Two data points were created to locate the samples (Mortier, Reference Mortier2014): a northern one at the northern junction of the Rue Bonne Espérance and the Rue Rouge Lion, and a southern one at the southern junction of these same streets (Fig. 1.3). Along the Rue Bonne Espérance, the outcrops start at 99.8 m east of the northern datum point, being almost continuous from 142–213 m. In this section, the best exposures are located along the southwestern side of the street, the succession being poorly represented and weathered on the opposite side. Along the Rue Rouge Lion section, the exposures are mainly on its southwestern side, from 45.6–96.7 m south of the northern datum point. These sections were studied previously by Malaise (Reference Malaise1907), Michot (Reference Michot1932a, Reference Michot1934, Reference Michot and Fourmarier1954), Martin (Reference Martin1969), Vanmeirhaeghe (Reference Vanmeirhaeghe2006a), and Mortier (Reference Mortier2014).

Figure 2. Lithostratigraphy of the two studied sections at Tihange, with exact positions of the fossiliferous levels. Abbreviations: A., Akidograptus; bioz., biozones; C., Cystograptus; complan., complanatus; D., Dicellograptus; extraord., extraordinarius; Fm, Formation; M., Metabolograptus; Mb, Member; N., Normalograptus; pars., part; Obs., Observational; P., Pleurograptus; Par., Parakidograptus.

Figure 3. Examples of the interbedded accumulations layers: (1) and (2) from JC3 level; (3) and (4) from JC4 level. (1) IRSNB a13492, which shows several complete Eostropheodonta hirnantensis (M'Coy, Reference M'Coy1851) valves, numerous fragments of brachiopods, and several disarticulated pelmatozoan columnal plates; the white arrow indicates a Pentagonocyclicus (col.) sp. columnal; (2) IRSNB a13504, which shows numerous brachiopod and bryozoan fragments, as well as disarticulated pelmatozoan columnal plates; the white arrow indicates a Conspectocrinus (col.) celticus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973, columnal; (3) IRSNB a13509, which shows numerous brachiopod and bryozoan fragments, as well as disarticulated pelmatozoan columnal plates; the white arrow on the left indicates a trilobite sclerite; the white arrow on the right indicates a machaeridian plate; (4) IRSNB a13508, which shows numerous brachiopod and bryozoan fragments, as well as disarticulated pelmatozoan columnal plates; the white arrow indicates a Cyclocharax (col.) paucicrenellatus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973, columnal. All scale bars = 2 mm.
The Central Condroz Inlier and Ordovician stratigraphy of the Tihange Sector
The Condroz Inlier, also known as the ‘Bande de Sambre-et-Meuse,’ ‘Bande condrusienne,’ ‘Ride du Condroz,’ or ‘Bande de Dave’ (e.g., d'Omalius d'Halloy, Reference d'Omalius d'Halloy1842; Malaise, Reference Malaise1910; Maillieux, Reference Maillieux1939; Verniers and Van Grootel, Reference Verniers, Van Grootel, Andre, Herbosch, Vanguestaine and Verniers1991), is a long (~65 km) and narrow (0.5–4 km) WSW-ESE to SW-NE strip of Ordovician and Silurian of predominantly siliciclastic sedimentary rocks exposed roughly between Charleroi and Liège (Fig. 1.2). They contact with the Middle Devonian to lower Carboniferous succession of the Brabant Parautochthon to the north and the Lower Devonian of the Ardenne Allochthon to the south. Together with the Brabant Massif and the four Ardennes Inliers, this is one of the six areas in Belgium where the lower Paleozoic crops out. At least three tectonic zones can be distinguished within the Condroz Inlier, all of them having a distinct evolutionary history prior to conjoining together in this narrow band by Variscan tectonics (Michot, Reference Michot1980; Verniers et al., Reference Verniers, Herbosch, Vanguestaine, Geukens, Delcambre, Pingot, Belanger, Hennebert, Debacker, Sintubin and De Vos2001).
The study area (Fig. 1.2) belongs to the central and main part of the Condroz Inlier termed the ‘Central Condroz Inlier,’ comprising a post-Cambrian metasedimentary sequence, ranging from the Lower Ordovician (only represented in the Wépion borehole) to the upper Silurian, represented by siliciclastic rocks deposited in a deeper shelf setting (Verniers et al., Reference Verniers, Van Grootel, Louwye and Diependaele2002). Continuous sections are rare owing to the difficulty of properly correlating individual sedimentary successions and lithostratigraphic units, in addition to estimating their thicknesses across the inlier. Therefore, herein we will focus only on the litho- and biostratigraphical data from the study sections at Tihange village, avoiding uncertain stratigraphical correlations with other Condroz Inlier sectors.
The studied stratigraphic succession at Tihange (Fig. 2) was assigned to the Fosses Formation (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a). Its contact with the underlying lithostratigraphical unit (lithology and stratification are similar to those of the Rue de Corrière Member of the Vitrival-Bruyère Formation) is faulted (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a). The lower unit of the Fosses Formation, the Bois de Presles Member, is represented only in the Rue Bonne Espérance Section, being composed of brown-gray decalcified mudstones, bearing limestone nodules with marcasite and few macrofossil remains (Michot, Reference Michot and Fourmarier1954; Vanmeirhaeghe, Reference Vanmeirhaeghe2006a; Mortier, Reference Mortier2014). Michot (Reference Michot1932a, Reference Michot1934, Reference Michot and Fourmarier1954) cited the brachiopod Orthis calligramma Dalman, Reference Dalman1828, from this member in Rue Bonne Espérance Section, reidentified later by Sheehan (Reference Sheehan1987) as Portranella akymatata Sheehan, Reference Sheehan1987, which is restricted to middle to upper Katian rocks. Chitinozoans collected by Mortier (Reference Mortier2014) also suggest correlation with the middle to upper Katian.
Upwards, the lithology changes to gray-green, micaceous siltstones containing dark gray fusiform to elliptic, bioturbation traces (the ‘schistes mouchetés’ of Lassine, Reference Lassine1913) and small rusty cubes (possibly weathered pyrite) at the top of the Faulx-les-Tombes Member. The rare macrofossils found in this member (brachiopods, trilobites, and graptolites), together with a long-ranging chitinozoan assemblage, provide poor age constraints for this unit at this location (Mortier, Reference Mortier2014). However, in its type locality (Faulx-les-Tombes, Namur), this member is assigned to the Rawtheyan (=upper Katian, Ka4 stage slice; Vanmeirhaeghe, Reference Vanmeirhaeghe2006a, Reference Vanmeirhaegheb). A possible fault marks the boundary with the overlying Tihange Member. This member may be lithostratigraphically subdivided into a lower and an upper unit. The lower unit shows dark gray micaceous mudstones and rusty patches similar to the underlying Faulx-les-Tombes Member, suggesting a gradual transition between both members. Most of the chitinozoan specimens could not be attributed to species level. The specimens that are identifiable are only long ranging or reworked (Mortier, Reference Mortier2014).
The upper part of the Tihange Member is represented by light gray micaceous mudstones that coarsen upwards to cross-bedded fine-grained sandstones, fining to mudstones towards the top. The macrofossil assemblages are from this latter section of the Tihange Member, discussed in detail below. Mortier (Reference Mortier2014) recovered a poorly preserved specimen of the chitinozoan Ancyrochitina ellisbayensis? Soufiane and Achab, Reference Soufiane and Achab2000, (bed JM 06-58 in Fig. 2), suggesting a late Hirnantian age (Hi2) for the upper part of the Tihange Member. This unit is overlain by the Bonne Espérance Formation (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a, Reference Vanmeirhaeghe2007), a lower Rhuddanian (Silurian) succession of laminated dark graptolitic mudstones (Michot, Reference Michot1932a, Reference Michot1934, Reference Michot and Fourmarier1954; Verniers et al., Reference Verniers, Nestor, Paris, Dufka, Sutherland and Van Grootel1995; Mortier, Reference Mortier2014).
The Tihange Member
Vanmeirhaeghe (Reference Vanmeirhaeghe2006a, Reference Vanmeirhaeghe2007) erected the Tihange Member, which later was promoted to formation rank by Mortier (Reference Mortier2014). The unit, however, <20 m thick (Figs. 1, 2) and is not mappable at a regional scale (sensu Murphy and Salvador, Reference Murphy and Salvador1992). We thus follow Vanmeirhaeghe (Reference Vanmeirhaeghe2006a, Reference Vanmeirhaeghe2007) in recognizing the unit as a member. The Tihange Member corresponds to the “c” level of Michot (Reference Michot and Fourmarier1954, p. 65), a 15 m succession of sandy shale and thin sandstone beds that coarsen upwards in the Rue Bonne Espérance Section. So far, this unit is known only from the Tihange study area.
Malaise (Reference Malaise1907) was probably the first to document these levels, when he reported for the first time from Belgium the brachiopod species “Strophomena siluriana” from the ‘schistes quartzeux’ immediately underlying the Silurian graptolitic mudstone. Malaise's identification is questionable, and probably corresponds to Eostropheodonta hirnantensis (M'Coy, Reference M'Coy1851), which is one of the most common brachiopod species in the Tihange Member fossil assemblage of the Rue Bonne Espérance section. Unfortunately, Malaise (Reference Malaise1907) neither figured the specimens nor detailed their stratigraphic position, simply listing the species together with other fossil taxa, without specifying if they all occurred in the same bed or across several meters below the ‘Llandovery.’ This may have led the importance of this report to go unnoticed by subsequent authors, who probably considered Malaise's identification of “Strophomena siluriana” to correspond to other taxa occurring in the diverse brachiopod assemblage known from the lower members of the Fosses Formation (Sheehan, Reference Sheehan1987). Furthermore, the Tihange specimens identified as “Strophomena siluriana” were not reported from Malaise's Ordovician brachiopod collection when revised by Sheehan (Reference Sheehan1987) and Lespérance and Sheehan (Reference Lespérance and Sheehan1987). It is possible that Malaise's (Reference Malaise1907) material is lost.
The lower part of the Tihange Member crops out in the Rue Bonne Espérance section, 160–174 m eastwards from the northern datum point, with a thickness of 8.7 m. The boundary with the upper part of the Tihange Member is apparently conformable. In the Rue Rouge Lion section, from 74.4–77.8 m southwards of the northern datum point, we measured a thickness of 2.2 m; the boundary with the upper part of the Tihange Member is also a conformable stratigraphical contact, although modified by faults.
Mortier (Reference Mortier2014) distinguished five subdivisions within the upper part of the Tihange Member, which we follow here (Fig. 2). The succession starts with light gray, micaceous mudstones with the same grain size as the lower part of the Tihange Member (subdivision 1). Upwards, it becomes progressively coarser (subdivision 2), from very coarse-grained mudstones, to laminated siltstones and very fine-grained sandstones (subdivision 3), with beds up to 5 cm thick in the coarsest part that show crossbedding, indicating normal stratigraphical polarity in both sections. When grain size is coarser, lamination is observed. Towards the top, the grain size becomes finer again (subdivision 4), with mudstones having a similar grain size as those occurring at the base, and rare 3 cm beds of coarser mudstone towards the top (subdivision 5). The rocks are dark gray (not as dark as in the lower part of the Tihange Member) with rusty patches, similar to those in the lower part of the Tihange Member. In the Rue Bonne Espérance section, the thickness of the upper part of the Tihange Member, exposed from 174–184 m eastwards of the northern datum point, is 9.4 m, possibly with a faulted contact with the overlying lithostratigraphical unit. In the Rue Rouge Lion section, from 77.8–95.2 m southwards of the northern datum point, the thickness is 19.3 m, again with a possible faulted contact marking its upper limit. The fossil assemblages are from subdivision 3 of the upper part of the Tihange Member (Figs. 1.3, 2).
Materials and methods
The material originates from the eastern part of the Condroz Inlier, from two nearby sections of the Tihange Member that are exposed in the village of Tihange (Huy), between Namur and Liège (Fig. 1.2). Collecting took place during several field campaigns conducted by JM in 2007–2009 and recent campaigns in the spring and summer of 2020 by JC and SP.
Six fossiliferous beds from the upper part of the Tihange Member were sampled (Figs. 1.3, 2), two from the Rue Rouge Lion section (beds JC1 and JC2, corresponding, respectively, to JM 07–18 and JM 07–19 of Mortier, Reference Mortier2014) and four from the Rue Bonne Espérance section (beds JC3, JC4, JC5, and JC6; JC4 corresponding to JM 06–28 of Mortier, Reference Mortier2014; see detailed description below). These beds yielded a total of 168 brachiopods, 22 trilobites, and four graptolite specimens, as well as numerous crinoid columnals, disarticulated machaeridian plates, and bryozoans. Groups occurring in each sampled bed are shown in Figure 2.
Rue Rouge Lion section (Figs. 1.3, 2)
The assemblages from this section yielded abundant brachiopod and a few trilobite remains. The material is fragmentary, occurring in very fine-grained sandstones (subdivision 3) of the upper part of the Tihange Member, and the preservation, especially of the brachiopods, is quite poor. Two fossiliferous levels have yielded identifiable elements of the Hirnantia Fauna: JC 1 (=JM 07–18) level produced brachiopods, exclusively represented by Eostropheodonta hirnantensis (~49 specimens); JC 2 (=JM 07–19) level yielded a similar association to that of JC 1, but the brachiopod remains are less abundant with just few fragmentary valves of Eostropheodonta hirnantensis (~6 identifiable specimens), and 22 trilobite fragments were collected, seven of them identifiable as Mucronaspis sp.
Rue Bonne Espérance section (Figs. 1.3, 2)
The assemblages from this section are more diverse, and the preservation is generally better than in the Rue Rouge Lion Section. The material also is from subdivision 3 of the upper part of the Tihange Member, but in this case the fossils occur in very coarse-grained mudstones that alternate with the fine-grained sandstones of this subdivision. Six fossiliferous levels have yielded macrofossil remains: JC 3 level occurs in a bed of coarse mudstones, 3 cm thick, in which well-preserved brachiopods and graptolites occur, and a thin interbedded layer (<1 cm) consisting of accumulations of fragments (mostly isolated pelmatozoan columnals, ramose bryozoans and brachiopod fragments), with occasionally some complete brachiopod valves (Fig. 3.1, 3.2). This is the richest fossiliferous level, yielding ~83 brachiopod specimens (Eostropheodonta hirnantensis 89%, Plectothyrella crassicosta 7%, Hirnantia sp. 3%, Trucizetina? sp. 1%), 11 identifiable pelmatozoan columnals (Pentagonocyclicus [col.] sp. 64%, Cyclocharax [col.] paucicrenulatus 18%, Conspectocrinus [col.] celticus 9%, Xenocrinus sp. 9%), four graptolites (Normalograptus normalis 50%, Cystograptus ancestralis 25%,?Metabolograptus sp. 25%), and indeterminate ramose bryozoans. JC 4 (=JM 06–28) level occurs in a bed of very fine-grained sandstones, ~3 cm thick, with a thin interbedded layer (<1 cm) of accumulations of fragments, mostly isolated pelmatozoan columnals, ramose bryozoans, brachiopod fragments, occasionally with some complete brachiopod valves, disarticulated machaeridian plates, and rare trilobite sclerites (Fig. 3.3, 3.4). This level has yielded ~23 identifiable brachiopods (Eostropheodonta hirnantensis 95%, Plectothyrella crassicosta 5%), eight identifiable disarticulated pelmatozoan columnals (Pentagonocyclicus [col.] sp. 75%, Cyclocharax [col.] paucicrenulatus 25%), one machaeridian plate, a few indeterminate trilobite sclerites, and ramose bryozoans. JC 5 level consists of a bed of very fine-grained sandstones, ~2 cm thick, with a thin interbedded layer (<1 cm) of accumulations of fragments, mostly isolated pelmatozoan columnals, ramose bryozoans, and brachiopod fragments, with some occasional complete brachiopod valves and occasional trilobite sclerites. This level has yielded only a few identifiable remains, of which about four are identifiable brachiopods (Eostropheodonta hirnantensis 75%, Plectothyrella crassicosta 25%), three identifiable disarticulated pelmatozoan columnals (Pentagonocyclicus [col.] sp. 66%, Cyclocharax [col.] paucicrenulatus 33%), a few indeterminate trilobite sclerites, and ramose bryozoans. JC 6 level consists of a bed of very fine-grained sandstones, ~3 cm thick, yielding isolated brachiopod shells, only three of which are identifiable (Eostropheodonta hirnantensis 66%, Plectothyrella crassicosta 33%).
Repository and institutional abbreviation
The study material is housed at the Royal Belgian Institute of Natural Sciences (RBINS), Brussels, Belgium, with numbers IRSNB a13491 to a13523 (figured specimens) and Invert-34332 (non-figured specimens).
Systematic paleontology
The use of open nomenclature follows Bengtson (Reference Bengtson1988). Systematic assignment of the clade Machaeridia follows the proposal of Parry et al. (Reference Parry, Edgecombe, Sykes and Vinther2019).
Phylum Brachiopoda Duméril, Reference Duméril1806
Subphylum Rhynchonelliformea Williams et al., Reference Williams, Carlson, Brunton, Holmer and Popov1996
Class Strophomenata Williams et al., Reference Williams, Carlson, Brunton, Holmer and Popov1996
Order Strophomenida Öpik, Reference Öpik1934
Superfamily Strophomenoidea King, Reference King1846
Family Leptostrophiidae Caster, Reference Caster1939
Genus Eostropheodonta Bancroft, Reference Bancroft and Lamont1949
Type species
Orthis hirnantensis M'Coy, Reference M'Coy1851 from the Hirnant Formation (Hirnantian), Upper Ordovician, Aber Hirnant, near Bala, Wales, UK.
Eostropheodonta hirnantensis (M'Coy, Reference M'Coy1851)
Figure 4
- Reference M'Coy1851
Orthis hirnantensis M'Coy, p. 395.
- Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b
Eostropheodonta hirnantensis (M'Coy, Reference M'Coy1851); Rong et al., p. 13, figs.16C, G, H. [and the synonymy therein]

Figure 4. Eostropheodonta hirnantensis (M'Coy, Reference M'Coy1851) from the upper part of the Tihange Member, Fosses Formation, Tihange (Huy), all from JC3 level except (9) from JC4 level. (1–3) IRSNB a13491: (1) latex cast of exterior, (2) internal mold, and (3) latex cast of interior of a ventral valve; (4–7) IRSNB a13492: (4) latex cast of exterior, (5) internal mold, (6) detail of pseudopunctae, and (7) latex cast of interior of a dorsal valve; (8) IRSNB a13493: latex cast of exterior of a ventral valve showing mucronate extremities; (9) IRSNB a13507: latex cast of exterior of a dorsal valve showing mucronate extremities showing mucronate cardinal extremities; (10, 11, 15) IRSNB a13494: (10) internal mold, (11) detail of the ichnofossil Vermiforichnus Cameron, Reference Cameron1969, on the internal mold surface, and (15) latex cast of interior of a ventral valve; (12–14) IRSNB a13495: (12) internal mold, (13) latex cast of interior, and (14) detail of the crenulated dental plates of a ventral valve; (16–19) IRSNB a13496: (16) internal mold, (17) latex cast of interior, and (18) detail of the cardinalia showing the mesocardinal ridge of a dorsal valve; (19) IRSNB a13497: latex cast of interior of a dorsal valve in latero-oblique view showing the crenulated socket ridges. All scale bars = 2 mm
Holotype
Holotype was not selected by Davidson (Reference Davidson1871). Lectotype selected by Temple (Reference Temple1965, p. 410), internal mold of ventral valve (A 28831) from the Hirnant Formation (Hirnantian) of North Wales, UK (M'Coy, Reference M'Coy1852, pl. 1H, fig. 11c), figured again in Temple (Reference Temple1965, pl. XIX, fig. 1).
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Rouge Lion section, JC 2 [=JM 07–19] and Rue Bonne Espérance section, JC 3, JC 4 [=JM 06–28], JC 5, JC 6), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Other occurrences of Eostropheodonta hirnantensis
Zalesie Formation, Hirnantian of the Holy Cross Mountains, Poland (Baltica); Langøyene and Husbergøya formations, Hirnantian of Norway (Baltica); Kyrkås Formation, Jämtland and Tommarp Formation, Hirnantian of Västergötland, Sweden (Baltica), Kuldiga Formation, Hirnantian of Estonia, Lithuania, and Latvia (Baltica); Dorozhninsky horizon, Hirnantian of Russia (Gorny Altai); White Head Formation, Hirnantian of Québec (Laurentia); Tirekhtyakh Formation, Hirnantian of Russia (Kolyma Terrane); Hwe Mawng and Pangshsa-Pye formations, Hirnantian of Burma (Sibumasu); Keisley Limestone, Hirnantian of Westmorland, England (Lakesman Terrane); Kildare Limestone, Hirnantian of Ireland (Leinster-Lakesman Terrane); Tirnaskea Formation, Hirnantian of Northern Ireland (Midland Valley Terrane); Glyn, Plas Uchaf, and Cyrn-y-brain formations and Hirnant Limestone, Hirnantian of N. Wales (E. Avalonia); Haverford, Cwm Clyd, and Wenallt formations, Hirnantian of Mid Wales (E. Avalonia); Gögeren Formation, Hirnantian of NW Turkey (Pontides); Don Braulio Formation, Hirnantian of Argentina (Precordillera); Kosov Formation, Hirnantian of Czech Republic (Bohemia); Marmairane Formation, Hirnantian of France (Mouthoumet); “volcaniclastic unit,” Hirnantian of N. Spain (Iberia).
Materials
About 150 specimens, all preserved as internal and external molds (figured specimen numbers: IRSNB a13491–a13497 and a13507). Some of the material is fragmented, always disarticulated, but normally well preserved, showing details such as the papillate interior well (Fig. 4.6).
Remarks
The strong, crenulated dental plates, socket ridges crenulated posteriorly, and the elongated, erect cardinal process lobes indicate inclusion of this material within the genus Eostropheodonta. There are several species ascribed to this genus, ranging from the Katian (Upper Ordovician) to the Rhuddanian (Silurian). Among all the species assigned to Eostropheodonta, the Belgian shells most strongly resemble E. hirnantensis (M'Coy, Reference M'Coy1851) from the Hirnant Formation of Wales, E. squamosa Havlíček, Reference Havlíček1971 from the Kosov Formation of Bohemia, E. intermedia Havlíček, Reference Havlíček1971 from the Upper Ktaoua Formation of the Moroccan Anti-Atlas, E. discumbata Cocks and Brunton in Cocks et al., Reference Cocks, Brunton, Rowell and Rust1970, from the Cedarberg Formation of South Africa, and E. martnai (Rõõmusoks, Reference Rõõmusoks1993) from the Adila Formation of Estonia, by having mucronate to acute cardinal extremities. The Belgian material shows a ridge/process supporting the chilidium, located between the cardinal process lobes. This character is barely distinguishable, but present, in E. hirnantensis (Cocks et al., Reference Cocks, Brunton, Rowell and Rust1970) and well developed in the other species mentioned above, as well as in some specimens of E. parvicostellata Rong, Reference Rong1984, from the Kuanyinchiao Formation of Yichang, China and in Sinomena typica Zeng, Chen, and Zhang in Zeng et al., Reference Zeng, Chen, Wang, Zhang and Han2016, now considered a junior synonym of E. hirnantensis (see Rong et al., Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b). This structure has been considered as a third lobe of the cardinal process (Zeng et al., Reference Zeng, Chen, Wang, Zhang and Han2016), but more recently has been interpreted as the attachment structure for the adductor muscles in the dorsal valve (Rong et al., Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b). This ridge might alternatively, or additionally, correspond to a mesocardinal ridge (see Pope, Reference Pope1976), which is present in other strophomenoids, usually supporting the chilidium when preserved (Colmenar, Reference Colmenar2016), or even might be hidden under the chilidium (Pope, Reference Pope1976). The Belgian material also resembles E. squamosa by having a similar rib density of ~12–13 costellae per 5 mm measured at 10 mm anteromedially from the umbo, but differs from the Bohemian material in lacking the strongly marked concentric lamellae typical of the Bohemian species. Eostropheodonta intermedia differs from the material described here in having a much finer radial ornament of ~23 costellae at the same valve length and by lacking the mesocardinal ridge. Eostropheodonta discumbata differs from the Belgian material in having more elongated cardinal process lobes extending anteriorly as long as socket ridges (while in the material described here, they are slightly shorter) and by having a thicker radial ornament counting ~7–8 ribs per 5 mm measured at 10 mm anteromedially from umbo. Eostropheodonta martnai also shows a mesocardinal ridge, but differs from the Belgian specimens by having a more convex ventral umbonal region and slightly concave dorsal valves. Eostropheodonta hirnantensis shares all characters such as shell outline, dimensions, rib density, occasional presence of concentric lamellae, size and divergence of dental plates, cardinal process, and notothyrial platform, with the Belgian material, but for the poor development of the mesocardinal ridge. Despite this single difference from the type material of the Welsh species, we assigned the Belgian valves to E. hirnantensis.
Class Rhynchonellata Williams et al., Reference Williams, Carlson, Brunton, Holmer and Popov1996
Order Orthida Schuchert and Cooper, Reference Schuchert and Cooper1932
Suborder Dalmanellidina Moore, Reference Moore, Moore, Lalicker and Fischer1952
Superfamily Dalmanelloidea Schuchert, Reference Schuchert and v Zittel1913
Family Dalmanellidae Schuchert, Reference Schuchert and v Zittel1913
Subfamily Dalmanellinae Schuchert, Reference Schuchert and v Zittel1913
Genus Trucizetina Havlíček, Reference Havlíček1974
Type species
Trucizetina subrotundata Havlíček, Reference Havlíček1974, from the Kosov Formation (Hirnantian), Upper Ordovician of Bohemia.
Trucizetina? sp.
Figure 5.10–5.12

Figure 5. Brachiopods from the upper part of the Tihange Member, Fosses Formation, Tihange (Huy). All from JC3 level. Plectothyrella crassicosta (Dalman, Reference Dalman1828) (1–9), (1–3) IRSNB a13498: (1) latex cast of exterior, (2) internal mold, and (3) latex cast of interior of a dorsal valve; (4–6, 8) IRSNB a13499: latex cast of a specimen with conjoined valves in (4) ventral, (5) posterior, and (8) lateral views, (6) internal mold of the precedent in ventral view; (7) IRSNB a13500: internal mold of a dorsal valve; (9) IRSNB a13501: internal mold of a ventral valve. Trucizetina? sp. (10–12) IRSNB a13502: (10) latex cast of exterior, (11) internal mold, and (12) latex cast of interior of a dorsal valve. Hirnantia sp. (13, 14) IRSNB a13503: (13) internal mold and (14) latex cast of interior of a juvenile dorsal valve. All scale bars = 2 mm.
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Materials
Single internal and external molds of a dorsal valve available (figured specimen number IRSNB a13502).
Remarks
The generic assignment of this dalmanellid shell is difficult, based on the only specimen. The dorsal valve shares some characteristics with the genus Trucizetina (e.g., the fascicostellate ornament; the slender, plate-like cardinal process that is shorter than the brachiophore bases, posteriorly protruding through the notothyrium; and the subcircular dorsal muscle field), but the incipient shallow dorsal sulcus contrasts with the characteristic deep, angular dorsal median sulcus of this genus. However, the last character varies within a population, as can be seen in the type material of this genus from the Hirnantian Kosov Formation of Bohemia, with some specimens showing a shallow sulcus (e.g., Havlíček, Reference Havlíček1977, pl. 31, fig. 8), as well as in some others assigned to this genus occurring in the upper part of the Trapiche Formation, probably late Katian in age, of the Precordillera (e.g., Benedetto, Reference Benedetto1999, figs. 8L, 8T, 8U, 8W, Reference Benedetto2002, pl. 1, fig. 28), and from the Hirnantian Hwe Mawng Purple Shale Member of the Naungkangyi Group Myanmar (e.g., Rong et al., Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b, fig. 19J), which in appearance even lack the sulcus. Until more material is available, the specimen is assigned tentatively to Trucizetina.
Suborder Dalmanellidina Moore, Reference Moore, Moore, Lalicker and Fischer1952
Superfamily Enteletoidea Waagen, Reference Waagen1884
Family Draboviidae Havlíček, Reference Havlíček1950
Genus Hirnantia Lamont, Reference Lamont1935
Type species
Orthis sagittifera M'Coy, Reference M'Coy1851 from the Hirnant Formation (Hirnantian), Upper Ordovician of North Wales.
Hirnantia sp.
Figure 5.13, 5.14
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance Section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Materials
Two incomplete internal molds of dorsal valves available from locality JC3 (figured specimen number IRSNB a13503).
Remarks
The only dorsal valve available displays a long, thin ridge-like cardinal process, located on a wide notothyrial platform, continuing anteriorly as a wide median ridge bisecting the adductor muscle field, which most probably belongs to Hirnantia. The small size of the valve, ~10 mm wide, and the ridge-like cardinal process without a lobate and crenulated myophore, a character that is usually developed in adult specimens, might indicate that it is likely a juvenile specimen. In the absence of the external characters and the poor preservation of the internal mold, specific determination is not possible.
Order Rhynchonellida Kuhn, Reference Kuhn1949
Superfamily Rhynchotrematoidea Schuchert, Reference Schuchert and v Zittel1913
Family Trigonirhynchiidae Schmidt, Reference Schmidt1965
Subfamily Rostricellulinae Rozman, Reference Rozman1969
Genus Plectothyrella Temple, Reference Temple1965
Type species
Plectothyrella platystrophoides; OD; =Plectothyrella crassicosta (Dalman, Reference Dalman1828) from the Hirnant Formation (Hirnantian) of North Wales.
Plectothyrella crassicosta (Dalman, Reference Dalman1828)
Figure 5.1–5.9
- Reference Dalman1828
Atrypa? crassicostis Dalman, p. 131.
- Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b
Plectothyrella crassicostis (Dalman, Reference Dalman1828); Rong et al., p. 21, fig. 23A–F. [and the synonymy therein]
Holotype
Internal mold and portion of external mold of conjoined valves (A 52001) from the Hirnant Formation (Hirnantian) of North Wales, UK (Temple, Reference Temple1965, PI. XX, fig. 3).
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance section, JC3, JC6), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Other occurrences of Plectothyrella crassicosta
Tirekhtyakh Formation, Hirnantian (Kolyma Terrane); Kuanyinchiao Formation, Hirnantian of China (South China); Wanyaoshu Formation, Hirnantian of China (N. Sibumasu); Xainza Formation, Hirnantian of Tibet (S. Tibet); White Head Formation, Hirnantian of Québec (Laurentia); Tommarp Formation, Hirnantian of Västergötland, Sweden (Baltica); Kuldiga Formation, Hirnantian of Estonia and Latvia (Baltica); Zalesie Formation, Hirnantian of the Holy Cross Mountains, Poland (Baltica); Kildare Limestone, Hirnantian of Ireland (Leinster-Lakesman Terrane); siltstones at Pond Pitch and Haskell Rock, Hirnantian of Maine (Miramichi Terrane); Glyn Formation and Hirnant Limestone, Hirnantian of N. Wales (E. Avalonia); Graig-wen Sandstone, Hirnantian of C. Wales (E. Avalonia); Wenallt Formation, Hirnantian of Mid Wales (E. Avalonia); Rio San Marco Formation, Hirnantian of Italy (Sardinia).
Materials
Five internal and external molds (three dorsal valves, two ventral valves) and a specimen with conjoined valves (figured specimen numbers IRSNB a13498–a13501).
Nomenclatural notes
The original species-group name by Dalman (Reference Dalman1828), crassicostis, was an attempt to create a combination of the adjective crassa (thick) + costa (rib). However, the adjective of costa would be costatus/costata. As it is, “crassicostis” is a dative plural form, a declination of the nominative crassicosta. According to the ICZN article 32.5.2.7, it must be corrected to the nominative singular (i.e., to crassicosta). For this reason, Bergström's (Reference Bergström1968) emendation to crassicosta, whatever his reasons may have been, is correct. Because crassicosta is a nominative singular noun, it does not need to agree with the gender of the genus (although it does phonetically agree with Plectothyrella), so the name crassicosta should be conserved regardless of future generic assignments.
Remarks
All the internal and external characters indicate inclusion of this material within the genus Plectothyrella Temple, Reference Temple1965. Among the species described, the Belgian specimens probably belong to the species Plectothyrella crassicosta (Dalman, Reference Dalman1828) based on its ribs. Villas et al. (Reference Villas, Lorenzo and Gutiérrez-Marco1999) considered the rib density as a criterion for discriminating between Plectothyrella crassicosta subspecies. Rong et al. (Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b) considered the Villas et al. (Reference Villas, Lorenzo and Gutiérrez-Marco1999) method inaccurate because the number of ribs on Plectothyrella crassicosta is highly variable, even within a population, and dependent on the ontogenetic stage of the specimen (i.e., the ribs in both sulcus and fold bifurcate and increase in number during ontogeny). Rib density and the zig-zag pattern formed in the commissure of the Rhynchonellata have been interpreted as a way of increasing the length of the commissure, and thus increasing the volume fluid flow, while preventing entrance to the mantle cavity b grains of sediment exceeding the shell gaps (Benton and Harper, Reference Benton and Harper2009). If this premise is correct, the size of this gap, and thus the thickening and rib density, may be an adaptation for a specific type of substrate and grain size, which explains the variability of this character in Plectothyrella species and the presence of numerous ecomorphotypes. Until this hypothesis is properly investigated, it is suggested here to cease using subspecies of Plectothyrella crassicosta.
Phylum Arthropoda Siebold and Stannius, Reference Siebold and Stannius1845
Class Trilobita Walch, Reference Walch1771
Order Phacopida Salter, Reference Salter1864
Suborder Phacopina Richter, Richter, and Struve, Reference Richter, Richter, Struve and Moore1959
Superfamily Dalmanitoidea Vodges, Reference Vodges1890
Family Dalmanitidae Vodges, Reference Vodges1890
Subfamily Mucronaspidinae Holloway, Reference Holloway1981
Genus Mucronaspis Destombes, Reference Destombes1963
Type species
Dalmanitina (Mucronaspis) termieri Destombes, Reference Destombes1963, from the Upper Ktaoua Formation, upper Katian (Kralodvorian of the Ibero-Bohemian regional scale) of Morocco.
Remarks
Zhou et al. (Reference Zhou, Zhou and Yuan2011) recognized two different subgenera within Mucronaspis: M. (Mucronaspis) and M. (Songxites). The type-species, Mucronaspis termieri, two other species from the Upper Ordovician of Morocco (M. zagoraensis Destombes, Reference Destombes1972, and M. greti [Destombes, Reference Destombes1963]) were included in Mucronaspis (Mucronaspis), and the M. mucronata group, together with Chinese occurrences of the genus, assigned to M. (Songxites). Zhou et al. (Reference Zhou, Zhou and Yuan2011) emphasized differences in the cephalic lateral border furrow of both, being weak and meeting the lateral margin posteriorly in the Moroccan types and distinct and confluent with the posterior border furrow at the genal angle in Mucronaspis (Songxites). The dependence of the lateral border furrow on preservation, and differences in its confluence with the posterior border furrow at species level within Dalmanitidae (Pereira, Reference Pereira2017, pl. 45–49), suggest this character alone does not justify the subgeneric classification of Zhou et al. (Reference Zhou, Zhou and Yuan2011). Other differences listed by Zhou et al. (Reference Zhou, Zhou and Yuan2011) for subgeneric classification (e.g., hypostome denticles, pygidial border width) also are not reliable in our opinion. The delicate hypostome denticles are entirely comparable in both sets of species (e.g., Destombes, Reference Destombes1963, pl. 3, fig. 11; Zhou et al., Reference Zhou, Zhou and Yuan2011, fig. 4J), and the pygidial border width is relevant only at the species level in many dalmanitids (e.g., Henry, Reference Henry1980). For this reason, we prefer to avoid subgeneric assignments within the Mucronaspis mucronata species-group.
Mucronaspis sp.
Figures 6.9–6.11, 6.14–6.16, 6.20, 6.21

Figure 6. Pelmatozoans, trilobites, machaeridians, bryozoans, and graptolites from the upper part of the Tihange Member, Fosses Formation, Tihange (Huy). Trilobite remains all from JC2 level, all graptolite remains from JC3 level. (1) IRSNB a13504: Conspectocrinus (col.) celticus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973, internal mold of a disarticulated columnal plate from JC3 level. (2) IRSNB a13505: Xenocrinus sp., latex cast of a disarticulated columnal plate from JC3 level. (3, 4, 7, 8) IRSNB a13509: Indeterminate pelmatozoan columnals, latex casts of three different taxa; (3) from JC5 level; (4) IRSNB a13510 from JC3 level; (7) IRSNB a13511; (8) IRSNB a13512 from JC4 level. (5) IRSNB a13506: Pentagonocyclicus (col.) sp., latex cast of a disarticulated columnal plate from JC3 level. (6) IRSNB a13508: Cyclocharax (col.) paucicrenellatus Le Menn, Reference Chauvel and Le Menn1973, latex cast of a disarticulated columnal plate from JC4 level. Mucronaspis sp. (9–11; 14–16; 20, 21). (9) IRSNB a13513: internal mold of a cranidium from JC2 level in dorsal view; (10) IRSNB a13514: internal mold of a fragmented fixigena in dorsal view; (11) IRSNB a13515: external mold of a fragmented fixigena and genal spine in dorsal view; (14) IRSNB a13516: latex cast of external mold of an ocular surface in lateral view; (15, 16) IRSNB a13517: (15) internal mold and (16) latex cast of the external mold of a meraspid pygidium in dorsal view; (20) IRSNB a13518: internal mold of a hypostome in ventral view; (21) IRSNB a13519: internal mold of a fragmented pygidial pleura in dorsal view. (12) IRSNB a13509: Machaeridian plate indet., external mold. (13) IRSNB a13520: Cystograptus ancestralis Štorch, Reference Štorch1985, complete, partly damaged specimen. (17) IRSNB a13521: ?Metabolograptus sp., distal part of the rhabdosome. (18, 19) Normalograptus normalis (Lapworth, Reference Lapworth1877). (18) IRSNB a13522: specimen preserved in profile view; (19) IRSNB a13523: incomplete specimen in sub-scalariform view. All scale bars = 2 mm.
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Rouge Lion section, JC2 [=JM 07–19] and Rue Bonne Espérance section, JC4 [JM 06–28]), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Materials
The available materials are fairly fragmentary, consisting of isolated sclerites. One cranidium (internal mold: IRSNB a13513); two fixigenae (internal mold: IRSNB a13514 and external mold: IRSNB a13515); one eye surface (external mold: IRSNB a13516); one hypostome (internal mold: IRSNB a13518); one pygidial fragment (IRSNB a13519); one meraspid pygidium (internal and external molds: IRSNB a13517).
Remarks
The available material is crushed and very incomplete, thus best left under open nomenclature. Nevertheless, it allows some comparisons with named species. The genus assignment is possible due to the typical glabellar lobation, eye lobe position, fixigenal border furrows, long genal spine aligned with the lateral border, hypostome morphology and sculpture, and one meraspid pygidium, which entirely matches juveniles of Mucronaspis (e.g., Temple, Reference Temple1952b, pl. 10, figs. 4, 5). Although the palpebral lobe is not entirely preserved, it is possible to verify it does not extend back farther than L2, which differentiates our material from Mucronaspis matutina (Dean, Reference Dean1962) from the Dufton Shales, upper Katian of Cumbria (England). Similarly, the eye lobe does not seem to be so short and the posterior pleural bands of the pygidium are not inflated, as in the Chinese wuningensis species-group (sensu Zhou et al., Reference Zhou, Zhou and Yuan2011). S1 is not bifurcate, as in M. guizhouensis (Yin in Yin and Li, Reference Yin and Li1978) from the upper Katian of Guizhou Province. The Belgian specimens also differ from a group of Mucronaspis species documented by Hints et al. (Reference Hints, Pärnaste and Gailite2012), called morphs D and E, from the Hirnantian of the East Baltic (Hints et al., Reference Hints, Pärnaste and Gailite2012, fig. 4D–F, M, N), which bear straighter axial furrows, not abaxially deviated by L3. Configuration of the lateral and posterior border furrows (Fig. 6.10) differentiates the Belgian material from the type species, M. termieri, as well as from M. zagoraensis (Destombes, Reference Destombes1972) and M. greti (Destombes, Reference Destombes1972), both from the Katian of Morocco. According to Temple's (Reference Temple1952a) ontogenetic study of Mucronaspis, the number of pygidial segments is constant early in development, therefore the Belgian material, which bears at least seven pleural ribs, also is differentiated from M. termieri and M. zagoraensis (six ribs). Nevertheless, and counter to the smooth surface described by Temple (Reference Temple1952a) in Mucronaspis olini (Temple, Reference Temple1952b) or Budil (Reference Budil1996) in Mucronaspis grandis (Barrande, Reference Barrande1852) meraspis, the external and internal surfaces of the Belgian meraspid pygidium exhibits small tubercles aligned along the anterior band of pleurae (Fig. 6.15, 6.16), previously undescribed for Mucronaspis juveniles (probably due to preservation). The only available hypostome (Fig. 6.20) does not permit observation of denticles in the posterior border, but it is morphologically identical with documented hypostomes for the genus (e.g., Kielan, Reference Kielan1960; Destombes, Reference Destombes1972; Ingham, Reference Ingham1977; Zhou et al., Reference Zhou, Zhou and Yuan2011). Its sculpture, together with the characteristic coarse tubercles in the median body, also includes a fine tuberculation covering its surface, like that described for M. zagoraensis (Destombes, Reference Destombes1972, p. 55). Other described species of Mucronaspis (see Hammann and Leone, Reference Hammann and Leone2007) are too poorly known to allow comparison.
Despite its poor state of preservation, the studied material allows a distinction from several Mucronaspis species and agrees with the features of the cosmopolitan Mucronaspis mucronata (Brongniart, Reference Brongniart, Brongniart and Desmarest1822), originally described from the Jonstorp Formation (upper Katian) in Sweden and distributed almost globally in the Hirnantian (e.g., Kielan, Reference Kielan1960; Owen, Reference Owen1981; Lespérance, Reference Lespérance, Cocks and Rickards1988; Yolkin et al., Reference Yolkin, Obut and Sennikov1988; Zhou et al., Reference Zhou, Zhou and Yuan2011).
Mucronaspis sp. is the first record of this genus in Belgium.
Phylum Echinodermata Bruguière, Reference Bruguière1791
Subphylum Pelmatozoa Leuckart, Reference Leuckart1848
Class Crinoidea Miller, Reference Miller1821
Subclass Camerata Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Order Monobathrida Moore and Laudon, Reference Moore and Laudon1943
Family Xenocrinidae Miller, Reference Miller1890
Genus Xenocrinus Miller, Reference Miller1881
Type species
Xenocrinus pencillus Miller, Reference Miller1881 from Richmondian strata (upper Katian, Upper Ordovician) of Ohio, USA.
Xenocrinus sp.
Figure 6.2
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Material
One single disarticulated columnal plate (number IRSNB a13505).
Remarks
Xenocrinus columnals are easily recognized by their characteristic square outline. This genus is known from at least three Hirnantian localities in Wales (Donovan and Veltkamp, Reference Donovan and Veltkamp1993). Ausich and Cournoyer (Reference Ausich and Cournoyer2019) confirmed the presence of Xenocrinus, also across the Ordovician-Silurian boundary, based on occurrences of isolated columnals and complete specimens of the species X. rubus Ausich and Copper, Reference Ausich and Copper2010 in Rhuddanian rocks of the Becsie Formation, Anticosti. The occurrence of columnals of Xenocrinus in the Tihange Formation associated with Rhuddanian graptolites suggests this genus also occurs in the Silurian of Europe (see Age of the Tihange Member fossil associations section for further information).
Pelmatozoa incertae sedis
Remarks
The pelmatozoan columnals described herein are crinoids, blastozoans, or a mixture of both, and therefore will be considered here as incertae sedis until better material becomes available. Morphogeneric names of pelmatozoans carry the suffix “(col.)” after the genus, as recommended by Stukalina (Reference Stukalina1968) and Donovan (Reference Donovan1986).
Morphogenus Conspectocrinus Stukalina, Reference Stukalina1969
Type species
Conspectocrinus conspectus Stukalina, Reference Stukalina1969 from the Upper Ordovician of Kazakhstan, Russia
Conspectocrinus (col.) celticus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973
Figure 6.1
- Reference Chauvel and Le Menn1973
Conspectocrinus celticus, Le Menn in Chauvel and Le Menn, p. 48, pl. 1, figs. 7–9.
- Reference Jacinto2015
Conspectocrinus (col.) celticus; Jacinto et al., p, 76, fig. 1N. [and the synonymy therein]
- Reference Colmenar, Pereira, Pires, da Silva, Sá and Young2017
Conspectocrinus (col.) celticus; Colmenar et al., p, 453, fig. 5U.
Holotype
Specimen LPB-1999 (Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973, pl. I, fig. 7) from the Upper Ordovician rocks of Coat-Carrec, Argol (Finistère, France).
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance Section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Other occurrences of Conspectocrinus (col.) celticus.—Obikalon Beds, lower Sandbian–lower Katian of Uzbekistan (Tien Shan); Zeravshan Range, Sandbian–Katian of Tadzhikistan; Sholeshook Limestone Formation, upper Katian of S. Wales (E. Avalonia); Portixeddu and Tuviois formations Katian of Italy (Sardinia); Rosan Formation, Katian of France (Armorican Massif); Montjoi Formation, upper Katian of France (Mouthoumet Massif); Cystoid Limestone Formation, upper Katian of the Iberian Chains (Iberia); uppermost La Mora Slates, Katian of the Catalonian Coastal Ranges (Iberia); La Devesa Formation, upper Katian of the Cantabrian Zone (Iberia); La Aquiana Limestone and Urbana Limestone, upper Katian of the Spanish Central Iberian Zone (Iberia); Porto de Santa Anna and Ferradosa formations, upper Katian of the Portuguese Central Iberian Zone (Iberia).
Material
One single disarticulated columnal plate (number IRSNB a13504).
Remarks
This discoidal columnal, with pentagonal lumen and depressed areola formed by five petaloid lobes, is characteristic of the species Conspectocrinus (col.) celticus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973. This species has been reported so far in upper Katian rocks; the occurrence in Belgium may represent the youngest record of this taxon, co-occurring with elements of the Hirnantia Fauna (see discussion on the age of the Tihange Member).
Morphogenus Cyclocharax Moore and Jeffords, Reference Moore and Jeffords1968
Type species
Cyclocharax fasciatus Moore and Jeffords, Reference Moore and Jeffords1968, from the Silurian of Indiana, USA.
Cyclocharax (col.) paucicrenellatus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973
Figures 6.6
- Reference Chauvel and Le Menn1973
Cyclocharax paucicrenellatus, Le Menn in Chauvel and Le Menn, p. 44, pl. 2, fig. 8.
- Reference Jacinto2015
Cyclocharax (col.) paucicrenellatus; Jacinto et al., p. 76, fig. 1M. [and the synonymy therein]
Holotype
Specimen LPB-1923 (Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973, pl. II, fig. 8) from the Upper Ordovician rocks of Coat-Carrec, Argol (Finistère, France).
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance Section, JC3, JC4 [JM 06–28], JC5), Rhuddanian, Condroz Inlier, Belgium.
Other occurrences of Cyclocharax (col.) paucicrenellatus
Portixeddu and Tuviois formations, Katian of Italy (Sardinia); Rosan Formation, Katian of France (Armorican Massif); Montjoi Formation, upper Katian of France (Mouthoumet Massif); Cystoid Limestone Formation, upper Katian of the Iberian Chains (Iberia); La Aquiana Limestone and La Devesa Formation, upper Katian of the Cantabrian Zone (Iberia); Urbana Limestone, upper Katian of the Spanish Central Iberian Zone (Iberia); Porto de Santa Anna and Ferradosa formations, upper Katian of the Portuguese Central Iberian Zone (Iberia).
Materials
Five disarticulated columnal plates available (figured specimen number IRSNB a13508).
Remarks
The circular outline, five pointed star lumen, and fine radial crenulae suggest that this columnal plate belongs to Cyclocharax (col.) paucicrenellatus Le Menn in Chauvel and Le Menn, Reference Chauvel and Le Menn1973. This species has been reported so far in upper Katian rocks. The occurrence in Belgium may represent the youngest occurrence of this taxon, co-occurring with Hirnantia Fauna elements (see discussion on the age of the Tihange Member).
Morphogenus Pentagonocyclicus Yeltysheva and Schewtschenko, Reference Yeltysheva and Schewtschenko1960
Type species
Pentagonocyclicus haldaranensis Yeltysheva and Schewtschenko, Reference Yeltysheva and Schewtschenko1960 from the lower Carboniferous of Russia.
Pentagonocyclicus (col.) sp.
Figure 6.5
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance section, JC3, JC4 [JM 06–28], JC5), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Materials
About 15 disarticulated columnal plates (figured specimen number IRSNB a13506).
Remarks
The circular outline, pentagonal lumen, rounded pentagonal areola about the same width than the crenularium, and the fine and numerous radial crenulae suggest inclusion of this material within morphogenus Pentagonocyclicus Yeltysheva and Schewtschenko, Reference Yeltysheva and Schewtschenko1960. The Belgian specimens strongly resemble those occurring in the upper Katian la Devesa Formation of the Spanish Cantabrian Zone (Gutiérrez-Marco et al., Reference Gutiérrez-Marco, Aramburu, Arbizu, Méndez-Bedia, Rábano and Villas1996), the upper Katian Porto de Santa Anna and Ferradosa formations of the Portuguese Central Iberian Zone (Jacinto, Reference Jacinto2015, pl. 5, fig. g), and the basal part of the Hirnantian Ribeira do Braçal Formation of the Portuguese Central Iberian Zone (Colmenar et al., Reference Colmenar, Pereira, Young, da Silva and Sá2019, fig. 6.14).
Phylum Annelida Lamarck, Reference Lamarck1809
Order Phyllodocida Dales, Reference Dales1962
Suborder Aphroditiformia Levinsen, Reference Levinsen1883
Machaeridia Withers, Reference Withers1926
Plate indet.
Figures 3.3, 6.12
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance Section, JC4 [JM 06-28]), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Material
One internal mold of an incomplete plate (number IRSNB a13509).
Remarks
One single incomplete, tiny plate (2 mm long), preserved as an internal mold, does not allow a further identification. The rugae are distinct (~24), continuous, widely spaced in the apical region of the plate, nearly twice as closely spaced as those on the anterior portion (18 rugae per mm), meeting the lateral margin at a slightly obtuse angle. One single gentle inflection is preserved, and no longitudinal folds are present on the plate. The incompleteness of the studied specimen does not allow for a definite differentiation among the three classically accepted families for the group: Lepidocoleidae, Plumulitidae, and Turrilepadidae (Adrain, Reference Adrain1992). The high density of dorso-ventrally aligned rugae and, specially, the more widely spaced rugae towards the apex are the most conspicuous characters, comparable with those present in some lepidocoleids (Lepidocoleus grayae Withers, Reference Withers1922, from the Ashgill of Girvan or L. sarlei Clarke, Reference Clarke1896, from the Wenlock of New York, bearing high density of rugae and aberrant spacing towards the apex, respectively). Nevertheless, a turrilepadid or a plumulitid association is also possible.
Phylum Hemichordata Bateson, Reference Bateson1885
Class Pterobranchia Lankester, Reference Lankester1877
Order Graptolithina Bronn, Reference Bronn1849
Suborder Axonophora Frech, Reference Frech1897
Family Normalograptidae Štorch and Serpagli, Reference Štorch and Serpagli1993
Genus Normalograptus Legrand, Reference Legrand1987
Type species
Climacograptus scalaris var. normalis Lapworth, Reference Lapworth1877; by original designation; from the Llandovery of County Down, Ireland.
Normalograptus normalis (Lapworth, Reference Lapworth1877)
Figures 6.18–6.19, 7.2–7.3
- Reference Lapworth1877
Climacograptus scalaris var. b normalis, Lapworth, p. 138, pl. 6, fig. 31.
- Reference Loydell2007
Normalograptus normalis; Loydell, p. 38, pl. 1, figs. 4, 7, text-figs. 12E, F, 16C, H, N, 19. [and the synonymy therein]

Figure 7. Drawings of the graptolites from the JC3 level of the upper part of the Tihange Member, Fosses Formation, Tihange (Huy). (1) IRSNB a13521: ?Metabolograptus sp., distal part of the rhabdosome with prominent nema, median septum, and little geniculated thecae. (2, 3) Normalograptus normalis (Lapworth, Reference Lapworth1877); (2) IRSNB a13522: incomplete specimen preserved in profile view, (3) IRSNB a13523: proximal part of sub-scalariform flattened specimen with particularly long virgella. (4) IRSNB a13520: Cystograptus ancestralis Štorch, Reference Štorch1985, partly damaged specimen with characteristic semicircular proximal end. Scale bar = 1 mm.
Holotype
Specimen (BU 1136) from the acuminatus Biozone of the Birkhill Shales at Dob's Linn, Moffat, southern Scotland, UK (Lapworth, Reference Lapworth1877, pl. 6, fig. 31; refigured by Elles and Wood, Reference Elles and Wood1906, pl. 26, fig. 2a).
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Materials
Two incomplete rhabdosomes available from locality JC3 (numbers IRSNB a13522, a13523).
Remarks
The two specimens, one preserved in profile and one flattened in sub-scalariform view, can be assigned to Normalograptus normalis with reasonable certainty. Both proximal parts represent a slightly, but markedly tapering rhabdosome with long and stout virgella. The dorso-ventral width (DVW) increases from 1.0 mm across the apertures of the first thecal pair, through 1.7 mm at th10, to the maximum observed DVW of 1.8 mm at th12. In addition, the two thecae repeat distance (2TRD 2 = 1.4–1.5 mm; 2TRD 10 = 1.8 mm) matches values reported by Loydell (Reference Loydell2007) in his revision of the type material. Normalograptus normalis is a cosmopolitan species with a long stratigraphic range (Goldman et al., Reference Goldman, Mitchell, Melchin, Fan, Wu and Sheets2011), comprising the entire Hirnantian and Rhuddanian. Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005) noted that N. normalis disappeared from the fossil record during early Hirnantian and reappeared as a Lazarus taxon in the late Hirnantian and early Rhuddanian. Some proliferation of the species can be observed in the ascensus-acuminatus Biozone, although misidentification with Normalograptus ajjeri (Legrand, Reference Legrand1977), another species of almost equal stratigraphic range, is common (Loydell, Reference Loydell2007), especially in northwestern Gondwana and peri-Gondwanan terranes. Normalograptus ajjeri can be differentiated from N. normalis by having a less-tapering rhabdosome and lesser maximum width (1.5 mm). Normalograptus premedius (Waern, Reference Waern1948), which is characteristic of Hirnantian/Rhuddanian boundary beds in Sweden (Koren’ et al., Reference Koren', Ahlberg, Nielsen, Ortega and Aceñolaza2003), can be readily differentiated from the Belgian specimens by its more tapering rhabdosome with a narrower proximal end and widely spaced thecae. Closely similar Normalograptus transgrediens (Waern, Reference Waern1948), which is also reported from the Ordovician/Silurian boundary interval in Sweden (Koren’ et al., Reference Koren', Ahlberg, Nielsen, Ortega and Aceñolaza2003), exhibits a slightly narrower rhabdosome and generally shorter virgella. Some other diagnostic features, such as point of insertion of median septum, are not preserved in our specimens.
Family Neodiplograptidae Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Genus Cystograptus Hundt, Reference Hundt1942
Type species
Diplograpsus vesiculosus Nicholson, Reference Nicholson1868; subsequently designated by Jones and Rickards (Reference Jones and Rickards1967), from the Llandovery of Dumfriesshire, Scotland.
Cystograptus ancestralis Štorch, Reference Štorch1985
Figures 6.13, 7.4
- Reference Štorch1985
Cystograptus ancestralis, Štorch, p. 97, pl. 4, figs. 1, 3–5; text-fig 3A–D.
- Reference Štorch and Serpagli1993
Cystograptus ancestralis; Štorch and Serpagli, p. 16, pl. 2, fig. 3, pl. 3, figs. 2–4, 7, 8, text-fig. 5C, J–M.
- Reference Piçarra, Štorch, Gutiérrez-Marco and Oliveira1995
Cystograptus ancestralis; Piçarra et al., fig. 3.7.
- Reference Maletz1999
“Neodiplograptus” sp., Maletz, fig. 2.10.
- ?Reference Maletz1999
Cystograptus vesiculosus; Maletz, p. 350, figs. 2.13, 4.11.
- Reference Koren', Ahlberg, Nielsen, Ortega and Aceñolaza2003
Cystograptus ancestralis; Koren’ et al., fig. 3.34.
- Reference Štorch and Feist2008
Cystograptus ancestralis; Štorch and Feist, p. 948, figs. 5.12, 14, 21; 9.10, 10.8.
- Reference Piçarra, Robardet, Oliveira, Paris and Lardeux2009
Cystograptus ancestralis; Piçarra et al., fig. 4C.
- Reference Loydell, Walasek, Schovsbo and Nielsen2017
Cystograptus ancestralis; Loydell et al., fig. 12F.
- Reference Štorch, Roqué Bernal and Gutiérrez-Marco2019
Cystograptus ancestralis; Štorch et al., figs 8n, 12j.
Holotype
Specimen (PŠ 73/1) from the lower acuminatus Biozone of the Želkovice Formation at Prague-Řepy, Czech Republic (Štorch, Reference Štorch1985, pl. 4, fig. 1, text-fig. 3A).
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance Section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Material
One complete but partly damaged rhabdosome available from locality JC3 (number IRSNB a13520).
Remarks
A single specimen of Cystograptus ancestralis shows a semicircular proximal end and a markedly incised, concave sicular aperture characteristic of this species. The present assignment is further supported by slightly convex and moderately inclined ventral walls and broad, distally facing apertures of the subsequent thecae. Thecal geniculation is almost absent or hidden under apertural margin of preceding theca. Basic morphometry of the specimen (DVW th1 = 1.85 mm, DVW th3 = 1.95 mm, DVW th7–8 = 2.45 mm, 2TRD 2 = 1.75 mm; 2TRD 7 = 2.0 mm) matches values measured in the Czech type material (Štorch, Reference Štorch1985). This characteristic and widespread species, confined in the upper ascensus and lower acuminatus biozones (i.e., middle part of combined ascensus-acuminatus Biozone), has been recorded by Štorch (Reference Štorch1985) in central Bohemia; Štorch and Serpagli (Reference Štorch and Serpagli1993) in Sardinia; Piçarra et al. (Reference Piçarra, Štorch, Gutiérrez-Marco and Oliveira1995) in south Portugal; Koren’ et al. (Reference Koren', Ahlberg, Nielsen, Ortega and Aceñolaza2003) in Scania, Sweden; Štorch and Feist (Reference Štorch and Feist2008) in Montagne Noire, France; Piçarra et al. (Reference Piçarra, Robardet, Oliveira, Paris and Lardeux2009) in Brittany, France; Loydell et al. (Reference Loydell, Walasek, Schovsbo and Nielsen2017) in Bornholm, Denmark; Štorch et al. (Reference Štorch, Roqué Bernal and Gutiérrez-Marco2019) in the Pyrenees, Spain; and in some other regions of Avalonia, Baltica, and northwestern Gondwana. A specimen with a semicircular proximal end and rather short sicula, reported from the middle part of the acuminatus Biozone of Deerlijk 404 well (Maletz, Reference Maletz1999) can be assigned to this species with some reservation; similarly Cystograptus vesiculosus (Nicholson, Reference Nicholson1868) and Diplograptus modestus, figured from British acuminatus Biozone by Rickards (Reference Rickards, Cocks and Rickards1988, fig. 1m, q), and Cystograptus vesiculosus reported from the acuminatus Biozone of Seville Province, Spain, by Jaeger and Robardet (Reference Jaeger and Robardet1979). Cystograptus sp., however, recorded in the acuminatus Biozone of northwestern Spain by Gutiérrez-Marco and Robardet (Reference Gutiérrez-Marco and Robardet1991) clearly belongs to Cystograptus ancestralis.
Genus ?Metabolograptus Obut and Sennikov, Reference Obut, Sennikov, Betekhtina and Zhuravleva1985
Type species
Diplograptus modestus sibiricus Obut, Reference Obut1955; by original designation; from the lower Llandovery of Siberia.
?Metabolograptus sp.
Figures 6.17, 7.1
Occurrence
Upper part of the Tihange Member (subdivision 3; Fig. 2) of Fosses Formation in Tihange, Huy (Rue Bonne Espérance section, JC3), lower Rhuddanian A. ascensus-P. acuminatus Biozone, Condroz Inlier, Belgium.
Material
One incomplete rhabdosome available from locality JC3 (number IRSNB a13521).
Remarks
The available specimen represents the distal part of a septate biserial graptolite with long and robust nema, 2TRD = 2.0–2.1 mm and DVW = 2.2–2.4 mm, characters that match either Metabolograptus ex gr. M. persculptus, a primitive Rickardsograptus, or one of the robust early Rhuddanian species assigned to Korenograptus. Flattened specimens of M. persculptus usually possess less-robust nema and more-geniculated thecae with apertures that occupy less of the rhabdosome width (Štorch and Loydell, Reference Štorch and Loydell1996). Two thecae repeat distance (2TRD) and the thecal overlap match those of M. persculptus, but the rhabdosome width (DVW) is greater. More specific determination of the specimen is not possible in the absence of a proximal part with diagnostic characters.
Affinities of the Belgian Hirnantia Fauna assemblage
Based on a comprehensive dataset, Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a) recognized two successive Hirnantian evolutionary brachiopod faunas,: the Hirnantia Fauna and the succeeding Edgewood-Cathay Fauna. The former, widespread and diachronous, was related to the glacial acme in the early-mid Hirnantian, and the latter thrived during post-glacial, warmer, shallow-water carbonate and siliciclastic environments of low latitudes (e.g., Laurentia, Baltica, Kolyma, Central Asia, Siberia, and South China). Through statistical methods, Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a) determined the core members of both faunas. The Belgian assemblage comprises three of the Hirnantia Fauna key taxa (Eostropheodonta, Plectothyrella, and Hirnantia), indicating a reliable assignment to this instead of the later Edgewood-Cathay Fauna. The only other brachiopod occurring in the studied assemblage is tentatively assigned to Trucizetina, which is an uncommon yet distinctive taxon of the Hirnantia Fauna, known from the Miramichi Terrane, Gorny Altai, Perunica, Sibumasu, South China, and the Precordillera (upper Katian–Hirnantian? Trapiche Formation). Furthermore, presence of the trilobite Mucronaspis, the core taxon of the so-called Mucronaspis Fauna (Lespérance, Reference Lespérance1974), which concurs with the Hirnantia Fauna, supports this assignment. Brachiopods are predominant in abundance and diversity in the studied assemblage (~84% of the entire reported association), which is largely dominated by Eostropheodonta hirnantensis (73%), followed by Plectothyrella crassicosta (5%). The remaining occurring groups (crinoids, bryozoans, machaeridians, and graptolites), present only at some of the fossiliferous levels, are in line with what has been reported for Hirnantia Fauna communities worldwide (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a, fig. 22).
Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a) analyzed faunal changes of the Hirnantia Fauna according to water depth, substrate, latitude, and geographical position. The Belgian association occurs in fine-grained sandstones, a somewhat coarser substrate than usual for the Hirnantia Fauna, especially for Plectothyrella and Trucizetina (see Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a, fig. 12). Nevertheless, Plectothyrella occasionally also occurs in coarser substrates, such as siltstone (e.g., Bergström, Reference Bergström1968; Benedetto, Reference Benedetto1986) and sandstone (e.g., Brenchley and Cullen, Reference Brenchley, Cullen and Bruton1984; Leone et al., Reference Leone, Hammann, Laske, Serpagli and Villas1991; Villas et al., Reference Villas, Lorenzo and Gutiérrez-Marco1999; Brenchley et al., Reference Brenchley, Marshall, Harper, Buttler and Underwood2006). Furthermore, the presence and preservation of graptolites in laminated siltstone intercalations may indicate the existence of slightly calmer (protected or deeper) environments where fauna could have inhabited. In terms of water depth distribution, the studied brachiopod assemblage suggests assignation to Benthic Assemblage (BA) 3 (Boucot, Reference Boucot1975) due to the predominance of Eostropheodonta (see Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a, fig. 11). The occurrence of several fairly fragmented specimens of Plectothyrella, typically common in the slightly shallower environments of BA2, might suggest assignment of the Belgian assemblage to the upper BA3, where those specimens might have been transported and accumulated from the shoreward-adjacent BA. Two different Hirnantia Fauna provinces have been classically differentiated based on latitudinal distribution of some key endemic brachiopod genera: the Kosov (subtropical and temperate latitudes) and Bani (polar latitudes) provinces (Rong and Harper, Reference Rong and Harper1988). During the early Paleozoic, Belgium was positioned in southeastern Avalonia (e.g., Owens and Servais, Reference Owens and Servais2007; Harper et al., Reference Harper, Rasmussen, Liljeroth, Blodgett, Candela, Jin, Percival, Rong, Villas, Zhan, Harper and Servais2013), a boundary position between low and middle latitudes. The low brachiopod diversity obtained from the studied assemblage is represented by genera co-occurring in both latitudinal settings. The boundaries of these provincial divisions are, in our opinion, less well defined; their limits having faded as more high-latitude Hirnantia Fauna associations are described (e.g., Bernárdez et al., Reference Bernárdez, Colmenar, Gutiérrez-Marco, Rábano, Zamora, Pankhurst, Castiñeiras and Sánchez Martínez2015; Álvaro et al., Reference Álvaro, Colmenar, Monceret, Pouclet and Vizcaïno2016; Colmenar et al., Reference Colmenar, Villas and Rasmussen2018; Popov et al., Reference Popov, Legrand, Bouterfa and Ghobadi Pour2019). Focusing on the identified core taxa of the Hirnantia-Mucronaspis fauna, the Belgian assemblage is comparable in composition and diversity with others from eastern Avalonia, especially some from North and Central Wales (e.g., Temple, Reference Temple1965, Cocks and Price, Reference Cocks and Price1975, Brenchley and Cullen, Reference Brenchley, Cullen and Bruton1984) and the Ganderian Leinster-Lakesman Terrane (Kildare, Ireland; Wright, Reference Wright1968), where Eostropheodonta hirnantensis dominates a low-diversity assemblage with fewer representatives of Plectothyrella crassicosta, Hirnantia, Mucronaspis, and, for the Aber Hirnant assemblage (N. Wales), fewer pelmatozoan columnals and bryozoans (Cocks and Price, Reference Cocks and Price1975). It is also interesting to note that the Hirnantia Fauna assemblages of the above-mentioned regions, especially that of the Kildare Limestone, overlie Katian brachiopod- and trilobite-bearing beds with several elements in common with the lower members of the Fosses Formation (Lespérance and Sheehan, Reference Lespérance and Sheehan1987; Sheehan, Reference Sheehan1987). The Belgian Hirnantia Fauna also bears some resemblance to occurrences from Baltica (e.g., Västergötland, Sweden; Bergström, Reference Bergström1968; Chen and Rong, Reference Chen and Rong2019) and even marginal Laurentia (Québec, Canada; Lespérance and Sheehan, Reference Lespérance and Sheehan1976). However, it is less diverse than those, which possess the same basic elements, but with the addition of other taxa. In fact, the low diversity of the Belgian Hirnantia Fauna and the broad geographical distribution of the identified species, make a comparison of its composition less robust. On the other hand, differences in the brachiopod community from Belgium, such as the absence of Dalmanella and Leptaena, which are some of the most characteristic taxa in the Avalonia domain, may be more related to other causes (e.g., age [see below], substrate, or geographical position) rather than only to latitude (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a).
Age of the Tihange Member fossil associations
Prior to this work, an Hirnantian age was proposed for the Tihange Member based on: (1) the stratigraphical position overlying the upper Rawtheyan (uppermost Katian) Faulx-les-Tombes Member and underlying the Silurian Bonne Espérance Formation (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a); (2) the presence of a brachiopod Hirnantia-Fauna assemblage, plus a poorly preserved specimen of the chitinozoan Ancyrochitina ellisbayensis? (Mortier, Reference Mortier2014); and (3) the lithological succession reflecting a relative sea level drop consistent with the pronounced glacio-eustatic sea-level fall, globally recorded in Hirnantian sediments (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a; Mortier, Reference Mortier2014). However, a Rhuddanian age cannot be excluded in any of these cases (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a). Biostratigraphical data from the lowermost overlying Bonne Espérance Formation indicate a range from the upper(?) part of the P. acuminatus graptolite Biozone to the A. atavus graptolite Biozone (lower to middle Rhuddanian); the basal Silurian ascensus graptolite Biozone has not been detected in this unit. On the other hand, Ancyrochitina ellisbayensis, an identification that was tentative and the preservation state insufficient (Mortier, Reference Mortier2014), is not restricted to the Hirnantian. Although this species biozone, defined on Anticosti Island, characterizes the uppermost Hirnantian (Soufiane and Achab, Reference Soufiane and Achab2000), this chitinozoan was reported by Verniers and Vandenbroucke (Reference Verniers and Vandenbroucke2006) from 12 cm below to 5 cm above the global boundary stratotype section and point (GSSP) for the base of the Silurian System, located in Dob's Linn, southern Scotland (Williams, Reference Williams, Cocks and Rickards1988). Finally, there are a few reported occurrences of taxa typical of the Hirnantia Fauna crossing the Ordovician-Silurian boundary and others lacking detailed biostratigraphic data (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a).
The JC 3 level of the Rue Bonne Espérance section, located ~1.5 m below the sample (JM 06–58) bearing Ancyrochitina ellisbayensis?, has yielded Cystograptus ancestralis, a characteristic graptolite of the Akidograptus ascensus-Parakidograptus acuminatus Zone, suggesting an early Rhuddanian (Rh1 stage slice) age for the uppermost part of the Tihange Member. The GSSP for the base of the Silurian (Williams, Reference Williams, Cocks and Rickards1988; Melchin, Reference Melchin, Ortega and Aceñolaza2003; Fan et al., Reference Fan, Melchin, Williams, Sheehan and Servais2005) is placed at the base of the A. ascensus Biozone, characterized by the first appearance of A. ascensus and P. praematurus. Hence, because graptolites have formed the basis for reliable correlation of Hirnantian rocks worldwide and for determining the age of the Hirnantia Fauna (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a), their presence is critical for discussion on the age of the upper part of the Tihange Member. In light of these new biostratigraphic data, we discuss the significance of this Hirnantia Fauna assemblage from Belgium and the implications of a Rhuddanian age for this unit.
An early Rhuddanian age for the upper part of the Tihange Member somewhat calls into question the presumed continuous sedimentation in the Tihange area (eastern Condroz Inlier) during the Hirnantian (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a; Mortier, Reference Mortier2014). For this to be confirmed, the lower part of the Tihange Member would have been a condensed sequence comprising most of the Hirnantian. The grain size of the lower part of the Tihange Formation is similar to that of the Faulx-les-Tombes Member, so it was probably deposited before eustatic sea-level fall when we assume that the shelf was not uplifted. An age of late Katian is preferred, although an age of late Katian to early Hirnantian cannot be excluded. On the other hand, the tectonic complexity of the outcropping Tihange sections makes evaluation of stratigraphical continuity difficult, so we are unable to draw any further conclusions.
The co-occurrence in the JC3 level of core brachiopods of the Hirnantia Fauna (Eostropheodonta hirnantensis and Plectothyrella crassicosta) and disarticulated pelmatozoan columnals (Pentagonocyclicus [col.] sp., Cyclocharax [col.] paucicrenulatus, Conspectocrinus [col.] celticus), all known so far from upper Katian rocks, is problematic and confuse the dating of these levels. Underlying this level, three fossiliferous beds in the upper part of the Tihange Member (JC 4–6) yield a similar assemblage with Eostropheodonta hirnantensis, Plectothyrella crassicosta, Pentagonocyclicus (col.) sp., and Cyclocharax (col.) paucicrenulatus, but no graptolites were recovered. On the other hand, the lowermost Bonne Espérance Formation, immediately overlying the Tihange Member, has a diverse graptolite assemblage characteristic of the P. acuminatus to A. atavus biozones, where, among those previously reported Tihange Formation graptolites, only N. normalis is still present (Mortier, Reference Mortier2014).
Besides the tentative identification of Ancyrochitina ellisbayensis, the uppermost Tihange Member (subdivision 4, level JM 06–33) in the Rue Rouge Lion section also provided poorly preserved specimens of Desmochitina erinacea Eisenack, Reference Eisenack1931, and Desmochitina juglandiformis Laufeld, Reference Laufeld1967 (Mortier, Reference Mortier2014), both characteristic of upper Sandbian–lower Katian deposits (Webby et al., Reference Webby, Cooper, Bergström, Paris, Webby, Droser, Paris and Percival2004; Vandenbroucke, Reference Vandenbroucke2005). Nevertheless, their presence is probably caused by reworking, as suggested by Mortier (Reference Mortier2014), as well as by Vanmeirhaeghe (Reference Vanmeirhaeghe2006a, sample JVM 05-182, table 10) for a poorly preserved and questionable D. juglandiformis specimen at the base of the Tihange Member in the same section. Furthermore, chitinozoans have not been reported from the Hirnantian GSSP, located in Wangjiawan, Yichang (Vandenbroucke et al., Reference Vandenbroucke, Hennissen, Zalasiewicz and Verniers2008), limiting the confidence in its applicability for worldwide correlations.
Assuming that the age of the upper part of the Tihange Member and, consequently, that of the studied macrofossiliferous assemblages is lower Rhuddanian, based on a graptolite assemblage indicative of the Akidograptus ascensus-Parakidograptus acuminatus combined biozone, we propose alternative hypotheses to explain this (apparent or not) incongruent occurrence of normally temporally separated faunal-representative assemblages.
Hypothesis 1
Relict Hirnantia Fauna: all fossils are contemporaneous and Rhuddanian in age. Globally, many Hirnantia Fauna occurrences are chronostratigraphically well constrained from continuous graptolite-bearing successions across the Ordovician and Silurian transition. Both regional and worldwide correlations show that most Hirnantia Fauna assemblages occur in lower to middle Hirnantian units, correlating mostly with the M. extraordinarius Biozone, straddling the interface with or being overlain by the M. persculptus Biozone (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a, fig. 4). Fewer, but still representative, were correlated with the M. persculptus Biozone, both with its lower (e.g., Leone and Benedetto, Reference Leone and Benedetto2019) and its uppermost (e.g., Štorch et al., Reference Štorch, Roqué Bernal and Gutiérrez-Marco2019) part. Based on this worldwide chronostratigraphic distribution, Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a) proposed diachronism of the Hirnantia Fauna, with its predominance in the lower to middle Hirnantian interpreted as an ecological preference for cool-water environments established during the major Gondwanan glaciation, in contrast with the succeeding post-glacial warm-water environment and consequent sea-level rise in the late Hirnantian and afterwards. Nevertheless, some Hirnantia Fauna occurrences are not calibrated with graptolites—their age assignment to lower or middle Hirnantian based on lithological correlation (e.g., Daijiagou and Honghuayuan, Tongzi, China; Chen et al., Reference Chen, Rong, Mitchell, Harper, Fan, Zhan, Zhang, Li and Wang2000) or assumed through the presence of a typical assemblage of this fauna (e.g., Ribeira do Braçal Formation, Portugal; Colmenar et al., Reference Colmenar, Pereira, Young, da Silva and Sá2019). On the other hand, there are upper Hirnantian assemblages bearing core taxa of the Hirnantia Fauna (e.g., uppermost Kosov Formation, Czech Republic, Havlíček, Reference Havlíček1989; Don Braulio Formation, Argentina, Leone and Benedetto, Reference Leone and Benedetto2019), and high-latitude peri-Gondwana Hirnantia Fauna assemblages occurring in post-glacial sediments (e.g., Amouktir Member of the Upper Second Bani Formation, Anti-Atlas of Morocco; Colmenar and Álvaro, Reference Colmenar and Álvaro2015), which may result from the diachronism suggested by Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a). There are also other occurrences that do not entirely fit in composition or expected chronostratigraphical distribution of the Hirnantia or Edgewood-Cathay faunas (e.g., Wenallt Formation, Central Wales, Williams and Wright, Reference Williams and Wright1981; Cedarberg Formation, South Africa, Rong and Harper, Reference Rong and Harper1988).
Following Hirnantian glaciation, a global warming occurred, triggering ice cap melting, sea level rise, expansion of the oxygen minimum zone (OMZ) towards shallower waters, and subsequent global anoxia, which provoked the second phase of the Hirnantian extinction (LaPorte et al., Reference LaPorte, Holmden, Patterson, Loxton, Melchin, Mitchell, Finney and Sheets2009; Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a). These caused extinction of most of the Hirnantia Fauna, and its replacement by the Edgewood-Cathay faunas, in shallow-water settings (BA2-3) that likely were unaffected by the anoxia (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a). But a few Hirnantia Fauna assemblages were located in refugia in deep, poorly oxygenated environments (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a). A clear example is the Hirnantia Fauna assemblage reported from the lowermost Silurian, P. acuminatus graptolite biozone of the Skelgill Formation, Yewdale Beck, in the Lake District, England (Harper and Williams, Reference Harper and Williams2002), representing the youngest documented occurrence of this fauna so far. The assemblage is composed of small but mature specimens, interpreted as a relict fauna seeking refuge in less-oxygenated, deep-water environments of the Cumbria basin (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a). This is the only currently accepted Silurian record of the Hirnantia Fauna, occurring interbedded with a graptolite assemblage typical of the A. ascensus-P. acuminatus biozone (Hutt, Reference Hutt1974, Reference Hutt1975).
The described assemblage herein from Belgium possibly represents an additional relict Hirnantia Fauna from the Rhuddanian, sharing characteristics with the Yewdale Beck occurrence that support and strengthen this hypothesis. Both regions were paleogeographically close during the Late Ordovician–early Silurian, with Belgium (Condroz Inlier) located in East Avalonia and Yewdale Beck in the Ganderian Leinster-Lakesman Terrane (Fig. 8), thus increasing the chances that, if there were a surviving Hirnantia Fauna, it would have developed and inhabited contiguous areas. The geographical proximity between both regions during the Late Ordovician is also highlighted by the close similarity of the Katian fossil assemblages from the Fosses Formation and those from coeval sequences within Cautley and Dent districts (Sheehan, Reference Sheehan1987, and Ingham, Reference Ingham1966, respectively). Both alleged relict assemblages are interbedded with graptolites of the A. ascensus-P. acuminatus Biozone, and in both sequences (upper part of the Tihange Member and lowest Skelgill Formation) there are different brachiopod-rich levels intercalating laminated shales/siltstones. The remaining shelly assemblage also bears some elements in common, namely pelmatozoan columnals and machaeridian plates (Harper and Williams, Reference Harper and Williams2002, p. 77), but these have never been illustrated or published for further comparison. Despite the similarities, the Belgian assemblage is composed of larger individuals and has a slightly lower diversity than that of the Yewdale Beck, with which it only shares one species (Plectothyrella crassicosta) and, questionably, Hirnantia. The genus Eostropheodonta, the most abundant taxon of the Belgian assemblage, was not reported from the Yewdale Beck relict assemblage. Nevertheless, these composition differences are not significant because, in both assemblages, specimens are disarticulated, broken, and accumulated in certain levels that are inferred to have been transported with granulometric hydrodynamic selection, and because faunal changes are highly dependent on water depth, temperature, and substrate (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a). A good example is that of the uppermost Kuanyinchiao Formation (Tongzi, Guizhou, China), in which a change in faunal composition, from a Dalmanella, Plectothyrella, and Hindella (DP) Community to an Eostropheodonta-dominated (EDP) Community, was interpreted as being related to substrate and temperature changes due to differences in environmental settings. Lastly, although brachiopod specimens are often disarticulated and fragmented, there are no obvious signs of reworking in the fossils of the Tihange Member, nor clasts of different lithologies suggesting a more complex taphonomic history. On the other hand, the presence of disarticulated pelmatozoan columnals, known so far from the upper Katian, may be better related to poor documentation of the occurrences of this group, which is commonly reported but not described in Hirnantian–Rhuddanian fossil assemblages, stressing the importance of including, whenever possible, the associations in their entirety and not only the most-studied groups (brachiopods and trilobites), to avoid biasing chronostratigraphical distribution.

Figure 8. Paleogeographic map of late Hirnantian–early Rhuddanian times showing positions of the Condroz Inlier and other terranes across the Iapetus Ocean. Paleoreconstruction based on Liljeroth et al. (Reference Liljeroth, Harper, Carlisle and Nielsen2017) and Torsvik and Cocks (Reference Torsvik and Cocks2016). Note that a modified version of the Gall's projection is used. Abbreviations: BL, Brompton Line; BVL, Baie Verte Line; C Nfl., Central Newfoundland Terrane; Dashw, Dashwoods Terrane; E. Avalonia, East Avalonia; E Nfl., East Newfoundland; Expl, Exploits Zone; Fm, Formation; Kilb, Kilbucho; Leins, Leinster-Lakesman Terrane; Monian C, Monian Composite Terrane; MV, Midland Valley Terrane; Nova Sc, Nova Scotia; Österg. Östergötland; PANTHA. PANTHALASSIC; RIL, Red Indian Line; S. Mayo, South Mayo Terrane; Shrop, Shropshire; SU, Southern Uplands Terrane; TH, Table Head (west Newfoundland, foreland basin); S Wales, South Wales; W. Avalonia, West Avalonia.
Skelgill Formation deposition was characterized by low-density turbiditic flows (e.g., Rickards and Woodcock, Reference Rickards and Woodcock2005), whereas a slightly different, likely shallower depositional context may be represented in the upper part of the Tihange Formation, in which the fossiliferous beds would correspond to storm-dominated intervals of accumulation (tempestites) periodically disturbing a relatively low-energy environment in which laminated mudstones and siltstones were deposited. The presence of Eostropheodonta, normally characteristic of an upper BA3 (Rong and Harper, Reference Rong and Harper1988), as well as the normal (large) size of the specimens in the Belgian assemblages, contrasts with the small (but mature) individuals of the lower Skelgill Formation, interpreted as inhabiting deep-water, poorly oxygenated environments (Rong et al., Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a) of the lower BA3 (Harper and Williams, Reference Harper and Williams2002). Brett et al. (Reference Brett, Boucot and Jones1993), using different sedimentological evidence published in the literature, estimated mean depths of ~30–40 m for BA3 (ranging from ~10–20 m to 60 m), even documenting cases with clear influence of the fair weather waves. These mean depths likely correspond to the offshore transition zone (OTZ) from the shoreface to the offshore, defined as the zone comprised from directly below fair weather wave base (FWWB) to storm weather base (SWB) (Nichols, Reference Nichols2009; Peters and Loss, Reference Peters and Loss2012; Ortiz and Ashton, Reference Ortiz and Ashton2016).
The causes or determinant factors for survival of these Hirnantia Fauna assemblages are not obvious. A possible explanation could be the geographical position of eastern Avalonia (~30°S) during Late Ordovician–early Silurian times, with an active tectonic context, which may have favored establishment of refugia, likely controlled by particular oceanic circulation patterns that may have geographically isolated these Avalonian sectors, delaying post-glacial effects of rising temperature and sea level. Geographic isolation previously has been noted as the most likely cause for the high diversity and unusual provinciality in shallow-water regimes of South China during the Late Ordovician–early Silurian (Wang et al., Reference Wang, Boucot, Rong and Yang1984; Zhan and Cocks, Reference Zhan and Cocks1998; Rong et al., Reference Rong, Boucot, Harper, Zhan and Neuman2006; Rasmussen and Harper, Reference Rasmussen and Harper2011). On the other hand, the Belgian assemblage may be a product of the prolonged process of the later stage of glaciation in this particular geographical sector, which would be consistent with the low diversity and absence of other biological groups commonly present in warmer waters.
Hypothesis 2
Relict Hirnantia Fauna with reworked pelmatozoan columnals: Rhuddanian assemblage mixed with older reworked fossils (Fig. 9). Vanmeirhaeghe (Reference Vanmeirhaeghe2006a) compared the sequences of the Puagne Inlier (western Condroz Inlier) with those of the central Condroz Inlier (e.g., in Faulx-les-Tombes) and farther to the east in the Tihange area (eastern part, central Condroz Inlier). Middle Katian successions in the eastern sectors are represented by shallower facies, inferring a platform deepening towards the west, where coeval successions are represented by deeper facies (Fig. 9.1). During the late Katian, an inversion followed (Fig. 9.2), which Vanmeirhaeghe (Reference Vanmeirhaeghe2006a) explained by tectonic uplift in the western Condroz Inlier with two possible causes: the Ardennian Deformation Phase or emplacement of a large pluton nearby. In view of the important sea-level drop occurring worldwide during the early Hirnantian (e.g., Brenchley, Reference Brenchley, Cocks and Rickards1988; Nielsen, Reference Nielsen, Webby, Paris, Droser and Percival2004), this model would explain the putative continuous sedimentation in eastern sectors (Tihange), while in the western sectors (Puagne Inlier) emersion would inhibit sedimentation from the Hirnantian (Fig. 9.3) up to the late mid Aeronian (Fig. 9.4), creating the stratigraphical hiatus. This emersion certainly led to erosion of the youngest units in the western sectors, possibly the Faulx-les-Tombes Member (the unit immediately underlying the sedimentary hiatus in the Puagne Inlier) and/or a younger unit (uppermost Katian–lowermost Hirnantian?), that has since been completely eroded. In fact, the Faulx-les-Tombes Member has a limited thickness in the Puagne Inlier (Michot, Reference Michot1928) compared to the central and western sectors of the Condroz Inlier, supporting an erosional event prior to deposition of the overlying Génicot Formation (Aeronian). Furthermore, evidence of erosion in the Puagne Inlier is provided by reworked microfossils from the Fosses Formation within the basal part of the Génicot Formation (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a).

Figure 9. Proposed model for the Condroz Inlier basin evolution during (1) mid to late Katian (Ka2–Ka3); late Katian (Ka4) (1, 2); early to late Hirnantian (Hi1–Hi2) (3); and early Rhuddanian (Rh1) (4) times, illustrating the preferred hypothesis for the age of the studied assemblages (modified after Vanmeirhaeghe, Reference Vanmeirhaeghe2006a). Uplift may be related to the Ardennian Deformation Phase or to thermal uplift. Abbreviations: FWWB, Fair Weather Wave Base; Mb, Member; SWB, Storm Wave Base; u., upper.
Considering the easterly slope of the Condroz Inlier and the putative continuous sedimentation in the Tihange area during the Hirnantian–Rhuddanian (Mortier, Reference Mortier2014), it is expected that western-eroded Katian material provided a sediment source for Hirnantian and Rhuddanian deposition in the Tihange sector, as suggested by the presence of reworked Katian chitinozoans in the Tihange Member (Vanmeirhaeghe, Reference Vanmeirhaeghe2006a; Mortier, Reference Mortier2014). Thus, the studied assemblages of the Tihange Member also may include reworked macrofossils, which, considering the fine granulometry of the units, could have been transported and accumulated during high-energy events. The presence of disarticulated pelmatozoan columnals in the Tihange Member, which are known so far from the upper Katian, despite the previously mentioned biasing of poorly documented groups in the Hirnantia Fauna occurrences, allow us to hypothesize that they may represent reworked material from the western sectors (Fig. 9.3, 9.4), where the Fosses Formation is characterized by fossiliferous calcareous shales, with trilobites (Lespérance and Sheehan, Reference Lespérance and Sheehan1987), brachiopods (Sheehan, Reference Sheehan1987), and abundant crinoid fragments in some levels (Michot, Reference Michot1934). The pelmatozoans columnals from the studied macrofossil assemblage in the Tihange Member occur mostly as disarticulated debris in particular levels, with rare, partially articulated stems and fewer well-preserved specimens. Each columnal is a single calcite crystal, favoring them to resist and behave like a clast when eroded, thus being hard to distinguish between accumulated/resedimented and reworked entities (sensu Fernández-López, Reference Fernández-López1991). The occurrence of reworked pelmatozoan columnals in younger units is quite common (e.g., Zamora et al., Reference Zamora, Álvaro and Vizcaïno2009; Ernst et al., Reference Ernst, Königshof, Taylor and Bohatý2011; Donovan et al., Reference Donovan, Jagt and Deckers2016), including pluricolumnal specimens robust enough to resist reworking processes (e.g., Donovan et al., Reference Donovan, Deckers, Jagt and de Winter2020). In terms of crinoid erosion indexes (e.g., CEI in Debout and Denayer, Reference Debout and Denayer2018), the Tihange pelmatozoan assemblage includes intact, unbroken columnals, to abraded and non-recognizable elements (Figs. 3, 6.1–6.8), all preserved as molds, which makes it impossible to evaluate reworking signatures, such as coatings, or evidence of pre-fossilization (Brett and Baird, Reference Brett, Baird and Lash1990; Clausen and Smith, Reference Clausen and Smith2008). Reworked pelmatozoan columnal debris is common within TST sequences (Carlucci et al., Reference Carlucci, Westrop, Brett and Burkhalter2014), roofed by black shales (Brett and Baird, Reference Brett, Baird and Lash1990), which is expected in upper Hirnantian–lower Rhuddanian successions due to global sea-level rise (e.g., basal part of the upper? Hirnantian Ribeira do Braçal Formation of Portugal; Colmenar et al., Reference Colmenar, Villas and Rasmussen2018, figs. 6–19). The remaining fossil assemblage of the Tihange Member (e.g., brachiopods, trilobites, machaeridians, and graptolites) show some fragmentation but are also represented by complete sclerites (Fig. 6.9, 6.15, 6.16) and specimens with conjoined valves (Fig. 5.4–5.6, 5.8), lacking clear evidence of reworking. Furthermore, somatic remains of brachiopods, trilobites, machaeridians, and graptolites are not as susceptible to reworking as single-crystal pelmatozoan columnals. Thus, the fossil assemblage from the Tihange Member may be composed of accumulated Rhuddanian fossils, including graptolites and representatives of the Hirnantia Fauna, together with reworked Katian pelmatozoan columnals that were disarticulated and concentrated in debris levels (Fig. 2). Such surfaces could be associated with major tempestites, during which the sediment was reworked, or alternatively with condensed sections formed during times of rapid sea-level rise (e.g., Brett and Baird, Reference Brett, Baird and Lash1990). The implications of a relict Rhuddanian Hirnantia Fauna assemblage in Belgium, discussed for Hypothesis 1, also apply in Hypothesis 2.
Hypothesis 3
Reworked Hirnantia Fauna: graptolites are the only Rhuddanian fossils, mixed with a reworked Hirnantian assemblage (Fig. 9.4). This hypothesis is raised here due to the problematic presence of a typical Hirnantia Fauna assemblage in the Rhuddanian, as previously discussed. Considering the stratigraphical hiatus in the western sectors (Puagne Inlier), spanning the Hirnantian and Rhuddanian stages, and following the facies model of Vanmeirhaeghe (Reference Vanmeirhaeghe2006a) for the Katian–Rhuddanian of the Condroz Inlier (Fig. 9), emersion may have led to erosion of a Hirnantian unit that had since been completely eroded before the deposition of the Génicot Formation (Aeronian). If this hypothetical unit were fossiliferous, incorporation of eroded sediments in the upper part of the Tihange Member could have resulted in the presence of reworked Hirnantian fossils (the studied Hirnantia Fauna assemblage) in this Rhuddanian unit. However, we would like to emphasize that this hypothesis is unlikely because there is no clear evidence of reworking, other than fragmentation, in the fossils of the index taxa that typify the Hirnantia Fauna of the Tihange Member. This hypothesis is also raised because the only other known Rhuddanian Hirnantia Fauna record, from Yewdale Beck, in the Lake District (Harper and Williams, Reference Harper and Williams2002), was challenged in the same way by Cocks (Reference Cocks2019, p. 19), who suggested that “…whilst the beds themselves are of undoubted Llandovery age, the shells themselves may have undergone redeposition from unconsolidated older muds at the edge of the continental shelf…”.
In the Tihange assemblage, Eostropheodonta hirnantensis and Plectothyrella crassicosta, index species of the typical Hirnantia Fauna, are represented by internal and external molds of large and complete isolated valves. Among testable taphonomic criteria, some of these fossils show no fracture surfaces in fragile areas (e.g., posterolateral mucronate cardinal extremities, cardinalia, etc.), ferruginous or phosphatic coatings, or traces of bioerosion or encrusting organisms, except for perforations found on some E. hirnantensis valves (see Fig. 4.1–4.3, 4.9–4.13, 4.15) likely attributed to the ichnogenus Vermiforichnus Cameron, Reference Cameron1969, probably produced by a symbiotic or parasitic polychaete (e.g., Pickerill, Reference Pickerill1976). There are also no differences in lithological composition between inner mold and the matrix of one specimen with conjoined valves of P. crassicosta (Fig. 5.6). This would imply that reworking would necessarily take place before complete lithification of older units and dissolution of the original shells and exoskeletons. Although under favorable conditions some modern brachiopod shells that are more resistant than fossil mineralized shells may be preserved in fair-weather deposits generated under high energy conditions (Simões et al., Reference Simões, Rodrigues, de Moraes Leme and Pires-Domingues2007), they usually show extremely rounded margins and a pronounced bias among the ventral and dorsal valve ratio (Holland, Reference Holland1988), which is not observed in the studied material. Brett and Bordeaux (Reference Brett, Bordeaux, MacKinnon, Lee and Campbell1991) showed that the fragile nature of brachiopod shells, especially regarding the brachial valves, resulted in a high degree of attrition, being rapidly broken down to unrecognizable fragments. Thus, it is very unlikely that brachiopod shells could survive intact following reworking. Finally, with this hypothesis one would expect the occurrence of Hirnantian biozones in the reworked chitinozoan assemblages of the Tihange Member; however, Mortier (Reference Mortier2014) documented mainly Desmochitina juglandiformis and Desmochitina erinacea (besides the doubtful specimen attributed to A. ellisbayensis, which, due to poor preservation, does not allow support or rejection of any of the hypotheses), which characterize the Katian associations of the Faulx-les-Tombes and Bois de Presles members. Evidence of reworked brachiopod shells is generally rare, and the documented occurrences are usually of shells incorporated into hardgrounds or preserved as reworked fragmented fossil debris (e.g., Gil-Peña et al., Reference Gil-Peña, Barnolas, Sanz-López, García-Sansegundo and Palau2001; Pérez-Pueyo et al., Reference Pérez-Pueyo, Bádenas and Villas2018).
Based in all the available data, Hypothesis 1 is the most robust interpretation for these biostratigraphically incongruous assemblages. Both the Tihange and the Yewdale Beck assemblages are more likely to represent relict Hirnantia Fauna occurrences than taphonomical reworked records. Although rare, occurrences of relict faunas following mass extinction, a phenomenon termed “dead clade walking” by Jablonski (Reference Jablonski2002), have been described from several time intervals (e.g., Baarli and Harper, Reference Baarli and Harper1986; Landing et al., Reference Landing, Westrop, Kroeger and English2011), as well as today (e.g., Spikkeland et al., Reference Spikkeland, Kinsten, Kjellberg, Nilssen and Väinölä2016). Refugia occur during the most catastrophic extinction events (e.g., Schulte et al., Reference Schulte, Alegret, Arenillas, Arz, Barton, Bown, Bralower, Christeson, Claeys, Cockell and Collins2010), and a combination of the incompleteness of the fossils record and artificial factors (sampling bias) may contribute to these records.
Conclusions
The Belgian association reported herein represents an unexpectedly late record for a typical Hirnantia fauna, co-occurring with a graptolite assemblage indicative of the Akidograptus ascensus-Parakidograptus acuminatus Biozone, from the lower Rhuddanian (Silurian). Although reworking processes could justify inconsistent temporal occurrences of geochronologically well-constrained communities, the index species of the Belgian Hirnantia Fauna are represented by complete specimens with no evident signs of reworking from lower horizons. In this sense, the Belgian association represents an additional relict Hirnantia Fauna in the Silurian, sharing characteristics with the only other known Silurian occurrence, from lower Rhuddanian rocks at Yewdale Beck (Lake District, England). Both regions were located paleogeographically close during the Late Ordovician–early Silurian. These anomalous occurrences are possibly related to a delay of post-glacial effects in this geographical sector, which may have favored the establishment of refugia. The lowermost Silurian occurrence of Hirnantia Fauna supports the diachronous establishment of this community reported by Rong et al. (Reference Rong, Harper, Bing, Rongyu, Xiaole and Di2020a), in this case particularly induced by temporal and spatial differences at the termination of the major glaciation.
Acknowledgments
We thank Associate Editor C. Sproat as well as B. Gudveig Baarli and Y. Candela, whose comments helped to improve the quality of this manuscript. We also thank C. Esteves and A. Cravinho for their assistance during fieldwork, C. Marques da Silva for various discussions, and S. Morrisson for providing literature. We also acknowledge M. Liljeroth for kindly sharing her detailed paleogeographic maps for the terranes across the Iapetus that helped preparation of the paleogeographic reconstruction presented here.
This study was supported by Portuguese funds from Fundação para a Ciência e a Tecnologia, I.P. (Portugal) in the frame of UID/Multi00073/2019, UIDB/00073/2020, and UIDP/00073/2020 projects of the I & D unit Geosciences Center (CGEO). SP was supported by a SYNTHESYS (Synthesis of Systematic Resources) grant (BE-TAF-2620). JC and JCGM were supported by CGL2017-87631-P project from the Spanish MICINN. JM was supported during his PhD by the BOF of Ghent University and the FWO-Flandres. PŠ acknowledges in-house support no. RVO 67985831 received from the Institute of Geology of the Czech Academy of Sciences. This is a contribution to the IGCP project 735 “Rocks and the Rise of Ordovician Life: Filling knowledge gaps in the Early Palaeozoic Biodiversification.”









