


Introduction
Mesozoic soft, muddy, carbonate marine shelf substrates hosted a variety of infaunal suspension-feeding bivalves similar to the modern Pholadomya candida (Sowerby, Reference Sowerby1823), which burrows in shallow muddy shelves. These large, elongate, thin-shelled bivalves feed by extending the foot through the anterior gape in search of detritus, pedal feeding (Morton, Reference Morton1981). Pholadomyidae survived the end-Permian mass extinction (Stanley, Reference Stanley2015). Beginning in the Triassic and expanding in the Jurassic, this family evolved into several different clades. The more common Cretaceous genera were Pholadomya Sowerby, Reference Sowerby1823; Homomya Agassiz, Reference Agassiz1843; Pachymya J. de C. Sowerby, 1826 (in Sowerby and Sowerby, 1812–Reference Sowerby and Sowerby1846); and Pleuromya Agassiz, Reference Agassiz1843, whose species generally were long ranging within and even across more than one stage.
Two common and relatively diverse Albian genera reported from Comanchean strata are Homomya, order Pholadomyoida Newell (Reference Newell1965), superfamily Pholadomyoidea King (Reference King1844), and Pleuromya, order Pholadida Gray (Reference Gray1854), superfamily Pleuromyoidea Zittel (Reference Zittel1895). The valve morphologies of the two groups are similar in many ways and casts are difficult to distinguish. Both groups have edentulous hinges although some Pholadomyidae King, Reference King1844 species have small knobs at the hinge line. Pholadomyoidae differ from Pleuromyoidae by the ligament structure, which is rarely preserved in external casts. Genera of Pholadomyoidae have an external opisthodetic, parivincular ligament and some have ligamental pseudonymphs. Homomya in the family Pholadomyidae has no lunule, but its escutcheon has marginal ridges that fade anteriorly. The external posterior ligament of Pleuromyoidea is opisthodetic, and the anterior ligament becomes subumbonal supported by weak chondrophores. Pleuromya species have no lunule or escutcheon and the right valve (RV) slightly overlaps the left valve (LV). The posterior shell outline of Homomya tends to be straight and sloping either toward the posteroventral corner or toward the dorsal margin, whereas the posterior margin of Pleuromya is evenly rounded. Consequently, Comanchean species previously assigned to Pleuromya are here reassigned to Homomya.
The order Pholadomyoida and the superfamily Pholadomyoidea contain Pholadomyidae King (Reference King1844), Margaritariidae Vokes (Reference Vokes1964), Ucumariidae Sánchez (Reference Sánchez2003), and Arenigomyidae Carter (Carter et al., Reference Carter2011). The superfamily Pholadomyoidea ranges from Early Ordovician to Holocene. Its common Cretaceous genus Homomya first appeared in Early Triassic and became extinct in middle Eocene but possibly continued into late Oligocene (personal communication, J.G. Carter, University of North Carolina at Chapel Hill, 2016). Carter et al. (Reference Carter2011) placed the family Pleuromyidae in the order Pholadida in the superfamily Pleuromyoidea with the families Ceratomyidae Arkell Reference Arkell1934 and Vacunellidae Astafieva-Urbajtis Reference Astafieva-Urbajtis1973. This classification places Pleuromyidae phylogenetically distant from Pholadomyidae, which is counter to molecular data (Harper et al., Reference Harper, Dreyer and Steiner2006; Bieler et al., Reference Bieler2014). Pleuromyidae ranged from Early Triassic to the end of the Cretaceous (Carter et al., Reference Carter2011) as did Pleuromya (Global Biodiversity Information Facility [GBIF], 2016). Pleuromya diversified during the Jurassic, especially during Middle and Late Jurassic (Duff, Reference Duff1991) and became widespread latitudinally from Antarctica to Russia and Greenland (GBIF, 2016); it was less common during the Early Cretaceous. These families are grouped in the megaorder Poromyata Ridewood, Reference Ridewood1903 by Carter et al. (Reference Carter2011) and in the subclass Anomalodesmata Dall, Reference Dall1889 by Newell (Reference Newell1965), Cox et al. (Reference Cox1969), Morton (Reference Morton1981), Harper et al. (Reference Harper, Dreyer and Steiner2006), and Bieler et al. (Reference Bieler2014).
Albian–lower Cenomanian Comanchean Series species of Pholadomyoidea have been neglected as biostratigraphic markers and paleoecological indicators. Some 22 fossil species have been described (Akers and Akers, Reference Akers and Akers2002). These species have been classed with Pholadomya, Homomya, Pachymya, Myopholas Douvillé (Reference Douvillé1907), Panopea Ménard de Groye (1807), and Pleuromya. Species of these genera are rather large and readily collected. Specimens are generally preserved as external casts that show external features of their uncomplicated shell morphology; internal features are rarely preserved.
These Comanchean infaunal suspension feeders (Stanley, Reference Stanley1970, Reference Stanley2015) are found in lime mud carbonate strata that represent muddy calcareous substrates in shallow shelf environments (Fig. 1). Modern Pholadomya occupies inshore waters and is a passive deep burrower that feeds on sediment detritus by extending its foot through the anterior gape, pedal feeding (Morton, Reference Morton1981, Reference Morton1985).

Figure 1 Middle Albian paleogeographic map of the Comanche Shelf, Texas, and surrounding areas (from Scott, Reference Scott2007). Tan areas represent shallow-water carbonate; light blue is shelf carbonate; dark blue is intrashelf basin or deeper-water carbonate. Location of type localities of Homomyid species: 1. Burnett County; 2. Parker County; 3. Travis County; 4. Grayson County; 5. El Paso County; 6. Comal County; 7. Coahuila, Mexico; 8. Tarrant County.
Ferdinand Roemer discovered and named the first species, Homomya alta, from the Fredericksburg Group during his exploration travels along the Texas Balcones escarpment and up the transecting rivers (Roemer, Reference Roemer1852). Roemer also noted a specimen that he placed in H. Panopea sp. and compared it to the European species Panopea regularis d’Orbigny (Reference d’Orbigny1843, v. 3, pl. 360, figs. 1, 2). Roemer (Reference Roemer1852) named Pholadomya pedernalis from Fredericksburg strata. Subsequent explorers discovered similar specimens in west Texas (Conrad, Reference Conrad1857; Shumard, Reference Shumard1860), in the El Paso area (Böse, Reference Böse1910), and in north Texas (Cragin, Reference Cragin1893, Reference Cragin1894, Reference Cragin1905; Hill, Reference Hill1893), and central Texas (Whitney, Reference Whitney1937). This species group ranges from the upper Aptian–lower Albian Glen Rose Formation of the Trinity Group and into the lower Cenomanian Buda Formation at the top of the Washita Group (Fig. 2). A similar diverse assemblage of shallow-water carbonate-shelf bivalves occupied the Albian Coahuila platform in northwestern Coahuila, Mexico (Perkins, Reference Perkins1961). The Albian Aurora Limestone was deposited on this carbonate platform as fringing buildups and is now exposed in the Sierra de Tlahualilo (Padilla y Sanchez, Reference Padilla y Sanchez1986). The Albian species, Pleuromya knowltoni (Hill, Reference Hill1893), has been proposed as a “guide fossil” of the informal Regional Dense Member (RDM) in the basal part of the Person Formation in central Texas (Small et al., Reference Small, Hanson and Hauwert1996). However, it occurs in both older and younger units; instead, Homomya comalensis Whitney, Reference Whitney1937 is the distinctive species of the RDM.
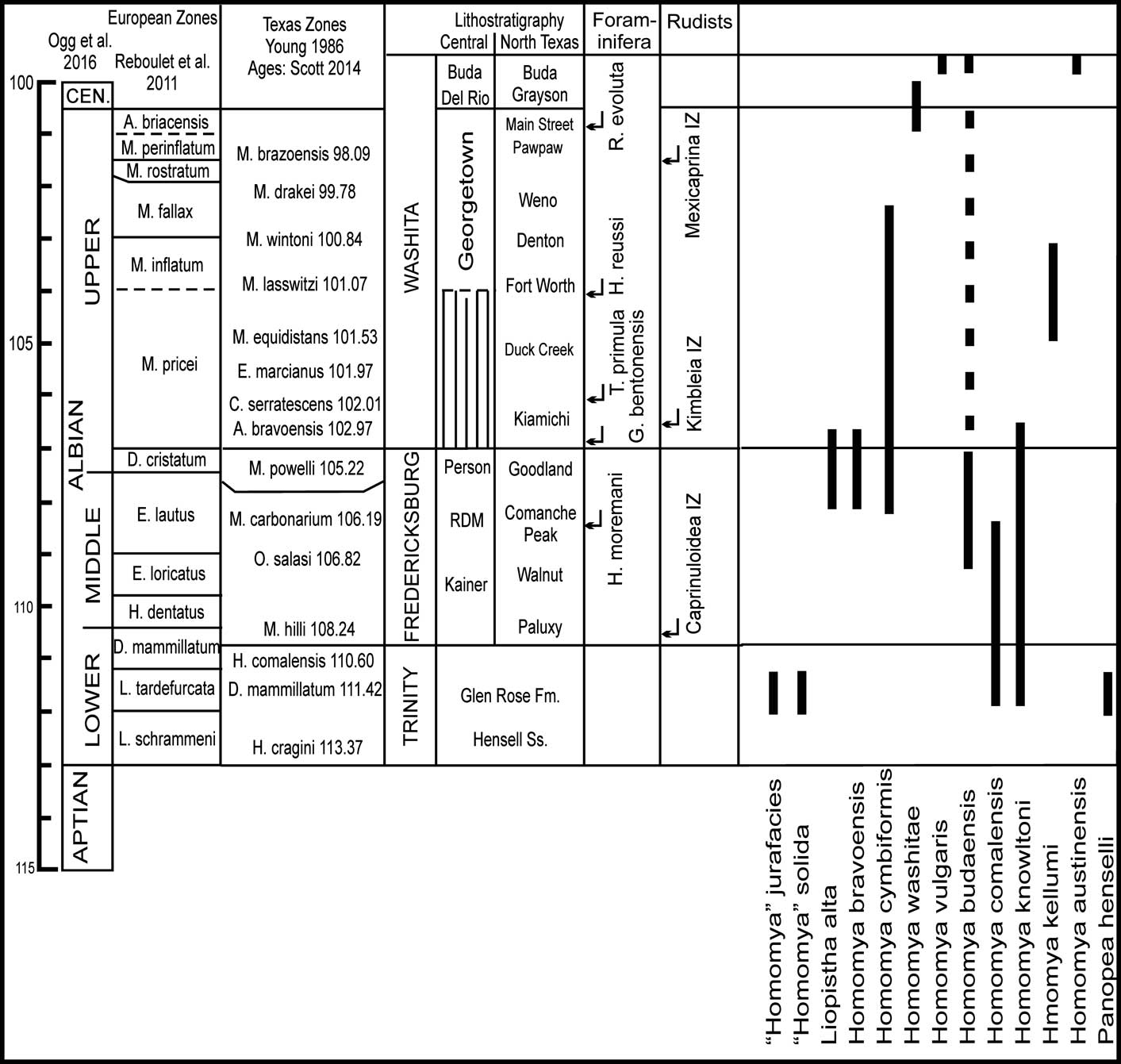
Figure 2 Biostratigraphy of Comanchean Series, Texas (Scott et al., Reference Scott, Benson, Morin, Shaffer and Oboh-Ikuenobe2003), with ranges of Albian Pholadomyidae bivalves. Planktic Foraminifera from Reichelt (Reference Reichelt2005, fig. 51). Rudist zones from Scott and Filkorn (Reference Scott and Filkorn2007).
This report intends to: (1) review the known Comanchean species of the families Poromyidae Dall (Reference Dall1889), Pholadomyidae, and Hiatellidae Gray, Reference Gray1824; (2) present distinctive criteria to identify these taxa consistently; (3) show their stratigraphic utility; and (4) propose paleoecological inferences.
Comanchean Series stratigraphy
The Comanchean Series on the U.S. Gulf Coast and in northern Mexico is a mixed carbonate-siliciclastic interval interbedded with localized evaporate units that spans upper Aptian to lower Cenomanian substages. The Comanchean is composed of three groups: upper Aptian to lower Albian Trinity Group, the middle to basal upper Albian Fredericksburg Group, and the upper Albian to lower Cenomanian Washita Group (Scott et al., Reference Scott, Benson, Morin, Shaffer and Oboh-Ikuenobe2003). Different subdivisions into formations reflect regional lithological changes from North Texas, south to the San Marcos Arch, and to West Texas (Fig. 2). Homomyid species occur in each group in Texas and northern Mexico (Fig. 2) and evolved across this time span. Ranges are relatively long, some up to eight million years and others only one to two million years. The Trinity Group is characterized by the first appearances of Homomya knowltoni (Hill, Reference Hill1895) and Homomya comalensis Whitney Reference Whitney1937; two other species are known only from the Trinity, but their generic assignment is uncertain. Four new Homomya species diversified in the middle-lower upper Albian Fredericksburg Group. One Trinity species, H. comalensis, ranges into the Fredericksburg basal marl member of the Person Formation on the San Marcos Arch, and H. knowltoni persisted into the basal formation of the Washita Group. Homomya washitae Cragin, Reference Cragin1894 spans the Albian/Cenomanian boundary, and three species are found only in the lower Cenomanian Buda Limestone (Fig. 2).
Homomya specimens are diverse and abundant in the Aurora Limestone in the Sierra de Tlahualilo in western Coahuila, Mexico (Perkins, Reference Perkins1961). The upper member of the Aurora is composed of fossiliferous, interbedded thick-bedded and nodular marly limestone up to 110 m (365 ft) thick. Perkins correlated the upper member with the middle-upper Albian Fredericksburg and Washita groups in North Texas. Perkins recognized an older Texigryphaea mucronata Zone and a younger Homomya kellumi Zone. The former species is characteristic of the Fredericksburg, which correlates the lower part of the upper Aurora member with middle-upper Albian. The H. kellumi Zone correlates the upper part of the upper Aurora member with the upper Albian Washita Group. In Texas, this species is in the Fort Worth Formation in the middle part of the Washita, which is characterized by the upper Albian ammonite species Mortoniceras lasswitzi (Young, Reference Young1957) (Fig. 2). Also in the H. kellumi Zone is the caprinid rudist Kimbleia albrittoni (Perkins, Reference Perkins1961), which ranges throughout the upper part of the upper Albian Washita in Texas (Fig. 2) (Scott et al., Reference Scott, Campbell, Hojnacki, Wang and Lai2016). Thus, the H. kellumi Zone in the Aurora correlates with the middle part of the Washita up to the Albian/Cenomanian contact.
Statistical comparison of species
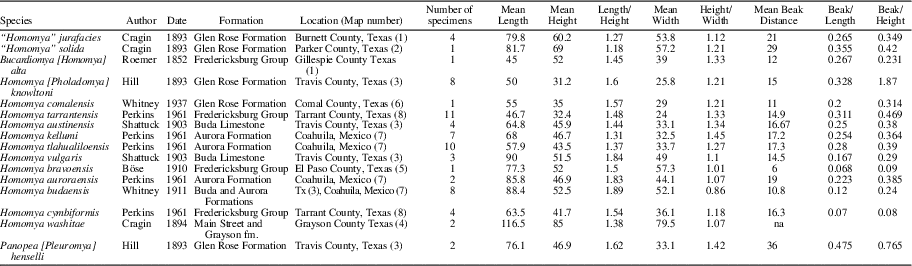
To identify consistently Comanchean species of Homomya, Liopistha (Sergipemya), and Panopea, dimensions of key morphological properties were measured (Fig. 3; Table 1). Because most specimens are preserved as internal or composite molds and most are broken, shell heights are more commonly complete than lengths. Two sets of species are compared, a smaller-sized set of H. knowltoni; H. tarrantensis Perkins, Reference Perkins1961; H. tlahualiloensis Perkins, Reference Perkins1961; H. kellumi Perkins, Reference Perkins1961; and H. comalensis (Fig. 4) and a larger-sized set of H. cymbiformis Perkins, Reference Perkins1961; H. austinensis Shattuck, Reference Shattuck1903; H. vulgaris Shattuck, Reference Shattuck1903; H. budaensis Whitnet, 1911; and H. auroraensis Perkins, Reference Perkins1961 (Fig. 5).
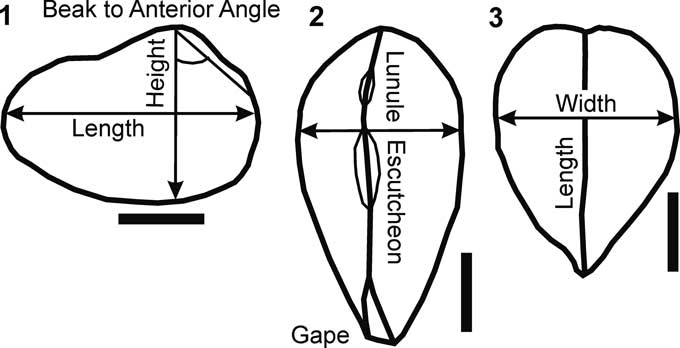
Figure 3 Outlines of a typical Comanchean homomyid showing measured dimensions on Table 1. (1) Right valve lateral view; (2) dorsal view; (3) anterior view. Scale bars=1 cm.

Figure 4 End-member Comanchean homomyid morphotypes. (1) Plot of height/width to valve volume of nine species shows two fields that overlap slightly: streamlined field in which valves are higher than wide and cylindrical field in which valves are as wide as or wider than high. (2, 3) Outlines of morphotype anterior and lateral left valve views: (2) H. knowltoni lectotype NMNH 145652; (3) cylindrical morphotype of H. budaensis hypotype UMMP 32854 (based on Perkins, Reference Perkins1961, pl. 24, figs. 6, 7). Scale bars=1 cm.

Figure 5 Histograms show the difference in shell height of five streamlined homomyid species in stratigraphic succession. STDEV.P is Excel standard deviation function.
Table 1 Species of Homomya and Panopea reported from Albian Strata, Comanche Shelf, Texas and Mexico.
The difference between heights of populations of H. knowltoni and H. tarrantensis are statistically insignificant (two-tailed, paired Student’s t test is 0.3001), and their stratigraphic ranges overlap in the Fredericksburg Group (Fig. 4). Likewise, the difference between heights of populations of H. kellumi and H. tlahualiloensis are statistically insignificant (two-tailed, paired p=.1195, CI=4.637) and both are in the Fort Worth Formation in the middle part of the Washita Group (Fig. 5). Heights of populations of H. cymbiformis, H. austinensis, H. vulgaris, and H. budaensis-auroraensis are also significantly different (Fig. 6). Statistical analyses were performed using equations in Excel spreadsheets and GraphPad software (www.graphpad.com/quickcalcs/ttest2/). Taxonomic implications will be discussed in the Systematic Paleontology section.

Figure 6 Histograms show the difference in shell height of five cylindrical homomyid species in stratigraphic succession. STDEV.P is Excel standard deviation function.
Five species appear to be quite similar, so they are compared on X/Y plots (Fig. 7): length/height (L/H) and height/width (H/W). The difference between the L/H ratios of H. knowltoni and H. tarrantensis is statistically insignificant (two-tailed p=.2004, CI=0.0686), but the H/W ratio is significantly different (two-tailed p=.0230, CI=−0.1040). This statistic indicates that H. knowltoni is significantly wider than H. tarrantensis; however, this difference is not a practical criterion on which to base a species concept. The H/W data points of H. knowltoni overlap those of H. tarrantensis in the Fredericksburg Group (Fig. 7). The difference between H/W ratios of the two younger species in the Washita Group, H. kellumi and H. tlahualiloensis, is also statistically insignificant (two-tailed p=.0097, CI=−0.18229); the lengths of too few specimens were available to form significant populations. These two species are very similar to each other and are distinctly larger than the two Trinity and Fredericksburg group species (Fig. 7). The lower Cenomanian species, H. austinensis, is relatively longer and higher than the other species. This group of Comanchean Homomya species became larger during the early Albian to early Cenomanian time span, approximately 16 million years.
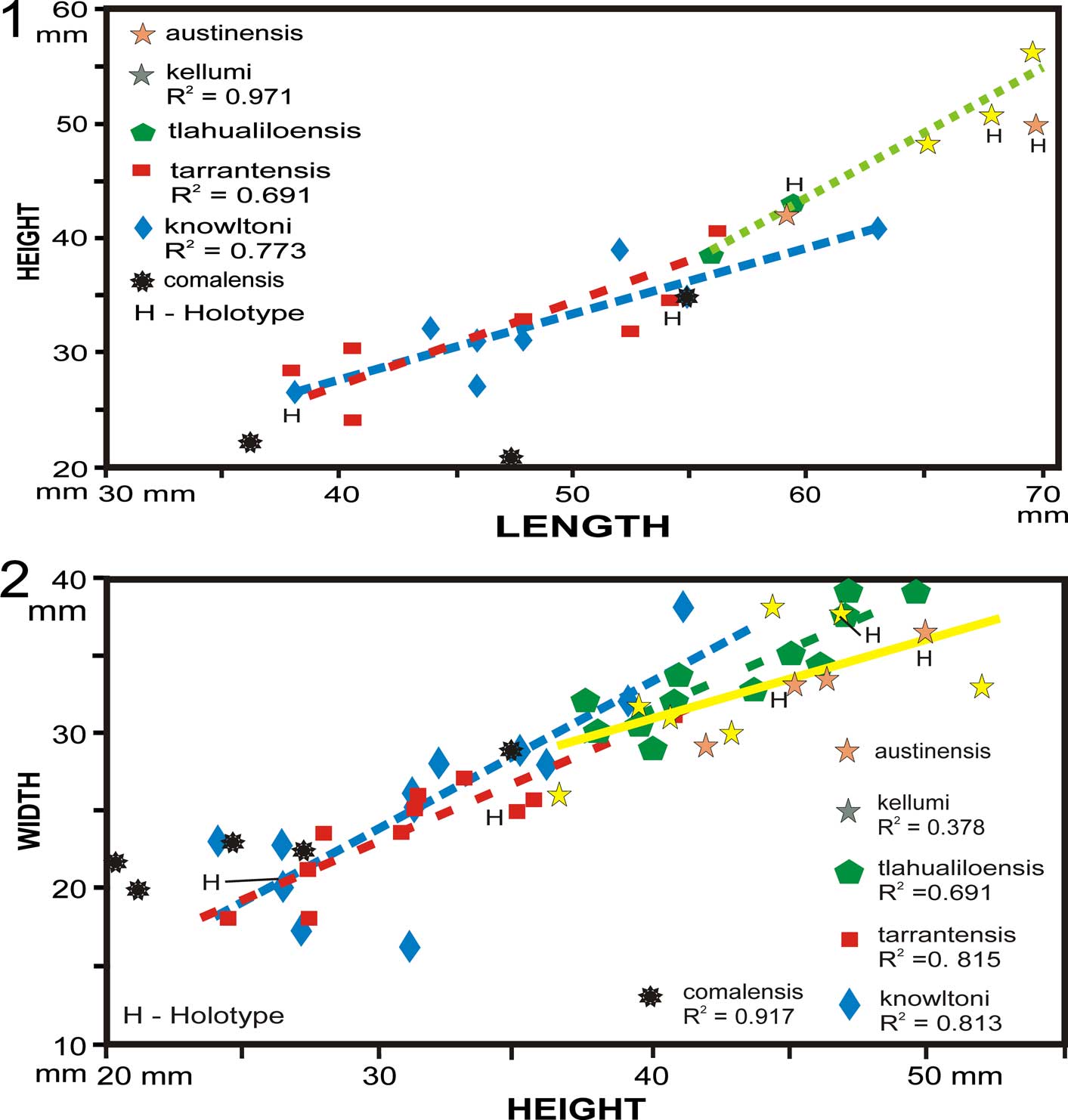
Figure 7 Cross plots of H. knowltoni, H. comalensis, and H. kellumi and their synonyms. (1) Length to height; (2) height to width.
Evolution of homomyid functional morphotypes
Two end-member morphotypes are represented by the “streamlined” Homomya knowltoni, which is an elongate, slightly inflated form with a relatively high umbo, and the cylindrical Homomya budaensis, which is a very elongate, tubular, inflated form with a very low umbo. The geometry of these shell forms is approximated by a cylindrical outline, the volume of which is [π × radius2 × length]; the radius is taken as half the width. The inflation of the valves is approximated by the height-to-thickness (width) ratio. The X/Y plot of these parameters (Fig. 8) separates the “streamlined” morphotype of H. knowltoni and H. cymbiformis from the cylindrical form of H. budaensis and H. auroraensis. The volume of the valves is related to the volume of the interior visceral mass, which includes foot, gills, and stomach. Comanchean Homomya species valves increased in size as the visceral mass increased, suggesting that the amount of sediment processed for organic matter increased. The two shape paradigms, streamlined and cylindrical, may reflect a difference in burrowing activity, which may have responded to substrate stability and fluidity.
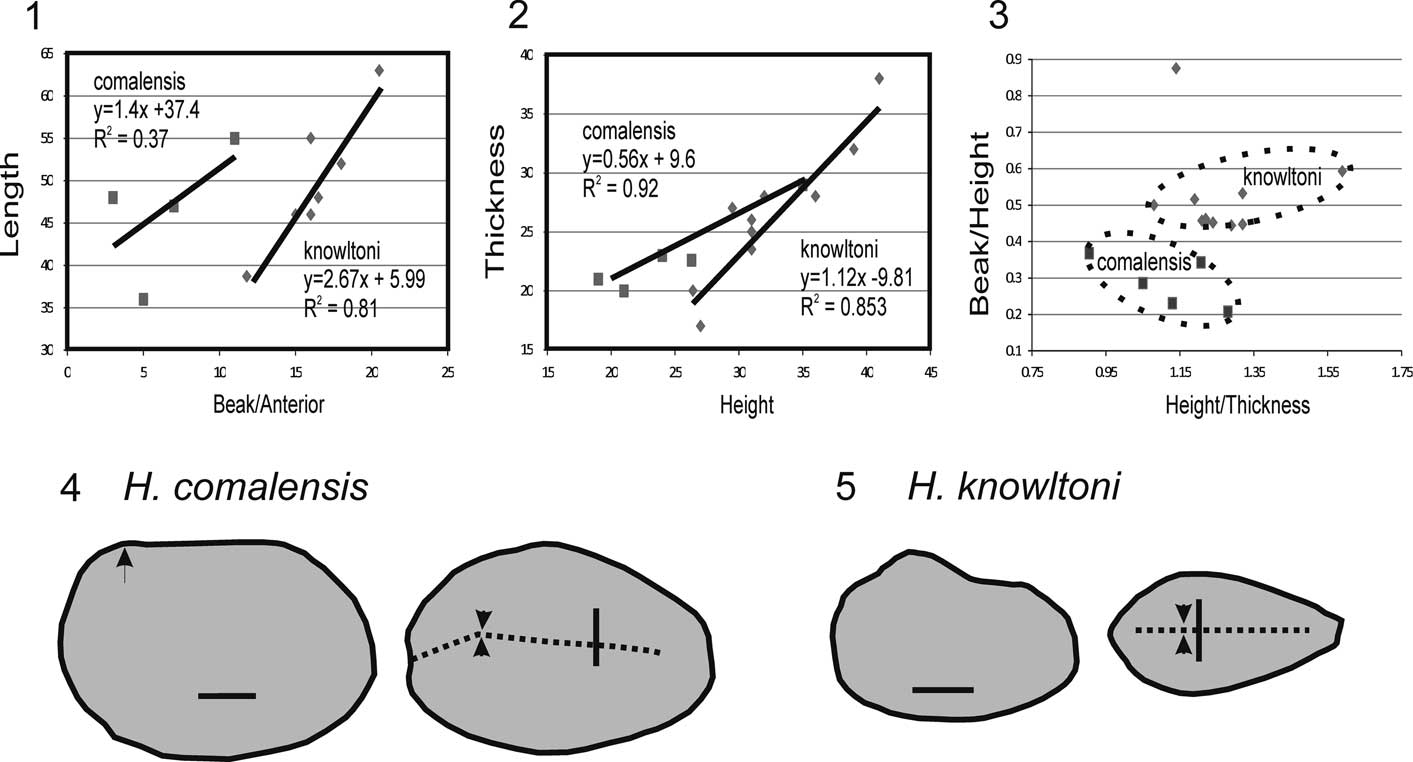
Figure 8 Comparisons of H. comalensis with H. knowltoni. (1) Distance of umbo from anterior margin versus valve length in millimeters. (2) Cross plot of valve height to thickness. (3) Cross plot of ratio of height to thickness versus ratio of umbo distance to height. The area of H. comalensis is separate from that of H. knowltoni. The outlier specimen is from the Comanche Peak Formation NPL44903. (4) Left lateral and dorsal outlines of H. comalensis illustrated by Whitney (Reference Whitney1937, pl. 13, figs. 3, 4). (5) Left lateral and dorsal outlines of H. knowltoni holotype USNM145644. Scale bars=1 cm; dashed line marks the commissure; arrows indicate beaks.
The “streamlined” forms of H. knowltoni, H. kellumi, H. austinensis, and H. cymbiformis first appeared on the Comanchean shelf in early Albian in the middle part of the Glen Rose Formation. These species occupied shallow shelf marls and successive species became larger. The cylindrical forms, H. budaensis, H. auroraensis, H. bravoensis Böse, Reference Böse1910 , H. vulgaris, and H. washitae, first appeared in the late Albian in the Del Norte Formation, southwest Texas. These large species occupied both calcareous shale and muddy carbonate substrates. Future cladistics analysis of all Mesozoic Pholadomyidae is recommended.
Materials and methods
Holotypes and other specimens have been reexamined and measured with calipers and selected specimens have been photographed (Table 1); measurements of all studied specimens are available as supplemental data. Additional new specimens have been collected during numerous field seasons in Texas Comanchean Cretaceous strata since the 1970s and have been measured and used for the analyses. Most species are uncommon to rare at any one outcrop, and some are incompletely preserved so that some species are represented by a small number of specimens. The taxonomy of Carter et al. (Reference Carter2011) is followed.
The suprafamilial classification and thus phylogeny of Bivalvia has advanced significantly since the application of molecular phylogenetics (Harper et al., Reference Harper, Dreyer and Steiner2006; Bieler et al., Reference Bieler, Carter, Coan, Bouchet and Rocroi2010, Reference Bieler2014; Combosch et al., Reference Combosch2017). The resulting classification differs from that based on “all available sources of phylogenetic information, including molecular, anatomical, shell morphological, shell microstructural, bio- and paleobiogeographic as well as stratigraphic, have been integrated into the classification” (Carter et al., Reference Carter2011, p. 1). The two classification positions construct different suprafamilial schemes, but the family-level schemes are similar. Analysis and critique of these two positions are beyond the scope of this paper. Because fossil bivalves in this study are preserved as composite casts, for which only rudimentary morphological features are preserved, the Carter classification is used.
Collections used in this study include the following: NMNH=National Museum of Natural History, Smithsonian Institution, Washington, D.C.; UMMP=University of Michigan Museum of Paleontology; NPL, UT, and WSA=Non-vertebrate Paleontology Laboratory, the Jackson School of Geosciences, the University of Texas at Austin. Former generic names are in brackets; accepted subgenera are in parentheses.
Repositories and institutional abbreviations
Type specimens in the U.S. National Museum of Natural History-Smithsonian, the University of Michigan Museum of Paleontology, and the Non-vertebrate Paleontology Laboratory, the University of Texas, Austin, have been studied.
Systematic paleontology
Class Bivalvia Linnaeus, 1758 in Linnaeus 1758–Reference Linnaeus1759
Clade Eubivalvia Carter in Carter et al., Reference Carter2011
Subclass Autobranchia Grobben, Reference Grobben1894
Infraclass Heteroconchia Hertwig, Reference Hertwig1895
Cohort Cardiomorphi Férussac, Reference Férussac1822 in Férussac Reference Férussac1821–1822
Subcohort Cardioni Férussac, Reference Férussac1822 in Férussac Reference Férussac1821–1822
Infrasubcohort Cardiidia Férussac, Reference Férussac1822 in Férussac Reference Férussac1821–1822
Megaorder Poromyata Ridewood, Reference Ridewood1903
Order Poromyida Ridewood, Reference Ridewood1903
Superfamily Poromyoidea Dall, Reference Dall1886
Remarks
This superfamily is composed of two families, Poromyidae Dall (Reference Dall1886) and Cetoconchidae Ridewood (Reference Ridewood1903), according to Carter et al. (Reference Carter2011).
Family Poromyidae Dall, Reference Dall1886
Remarks
Poromyidae genera are large, rounded to quadrate, most with radial costae, external ligament without a lithodesma, and at least one cardinal tooth in one valve (Keen in Newell, Reference Newell1969). Modern Poromyidae are deep-water infaunal carnivores (Morton, Reference Morton1981).
Genus Liopistha Meek, Reference Meek1864
Type species
Cardium elegantulum Roemer, Reference Roemer1852, p. 48–49, pl. 6, figs. 5a–c.
Diagnosis
Features of the family.
Occurrence
Early to Late Cretaceous.
Subgenus Sergipemya Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015
Liopistha (Sergepimya) alta (Roemer, Reference Roemer1852)
1852 Homomya alta Reference RoemerRoemer, p. 45, pl. 6, fig. 11.
1890 Homomya alta; Reference CraginCragin, p. 76.
1895 Homomya alta; Reference CraginCragin, p. 360.
1928 Homomya alta; Reference AdkinsAdkins, p. 141.
2002 Psilomya alta; Reference Akers and AkersAkers and Akers, p. 388, p. 389, fig. 372.
2015 Liopistha (Sergipemya) alta; Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus AndradeAyoub-Hannaa et al., p. 58.
Holotype
STIPB-Roemer-79, Steinmann-Institut, Paläontologie, Geologisch-paleontologische Institute, University of Bonn.
Diagnosis
Tall triangular shell, anterior margin truncated, posterior margin rounded; ornamented by irregular concentric ridges and grooves; minute, closely spaced nodes top concentric ridges and are vertically aligned in faint radial rows; posterior gape.
Occurrence
Roemer (Reference Roemer1852) collected the specimen of L. alta near Fredericksburg, central Texas, and Adkins (Reference Adkins1928) reported it in the Fredericksburg Group. Cragin (Reference Cragin1890, Reference Cragin1895) reported H. alta in the basal bed of the lower upper Albian Kiowa Formation in Kiowa County, southern Kansas. This 30 cm thick bed is a transgressive lag deposit that yields two ammonite species of the upper Goodland Formation in Texas: Venezoliceras acutocarinatum (Shumard, Reference Shumard1854) and Venezoliceras kiowanum (Twenhofel, Reference Twenhofel1924) (Young, Reference Young1966; Scott, Reference Scott1970).
Description
Shell globose to inflated, heart-shaped, anterior margin truncated, posterior margin subrounded, inflated, ornamented by concentric irregular striae and growth rings; umbo prominent, enlarged, prosogyrate (Roemer, Reference Roemer1852, p. 45, translated from the Latin). Valve length nearly the same as height.
Valve shape triangular, angle between anterior margin and dorsal margin about 65°, posterior margin short, straight, inclined about 145° from dorsal margin; posterior-ventral corner arcuate merging with broadly curved ventral margin that curves broadly into nearly straight anterior margin, wide ovate gape. Umbo broadly curved prosogyrate. Escutcheon narrow, elongate, bordered by low ridges on either valve. Ornamented by low, rounded, concentric growth rugae wider than interspaces, topped by minute, granule-like nodes aligned forming weak indistinct lineations.
Roemer (Reference Roemer1852) listed the length of H. alta as 1″ 9‴, height 1″ 9‴, width 1″ 5‴, which are translated as 1.9 inch and 1.5 inch. Converted to metric units: L=48.26 mm; H=48.26 mm; W=38.10 mm.
Remarks
Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015) designated Homomya alta Roemer (Reference Roemer1852) the type species of their new subgenus, Sergipemya. Their diagnosis of Sergipemya includes “posterior gape lacking; numerous fine commarginal ribs covering the valves and crossed by faint radial tubercles”; however, they state in their description of Liopistha (Sergipemya) alta that “radial ornament is absent” (Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015, p. 58). They also say that a posterior gape is absent. The type specimen of H. alta is ornamented by irregular concentric ridges and grooves (Roemer, Reference Roemer1852); minute, closely spaced nodes top the concentric ridges and are vertically aligned in faint radial rows (Fig. 11.5–11.7), which is consistent with the subgeneric diagnosis. However, the diagnosis should be modified to include the presence of a posterior gape. Roemer’s species is middle to upper Albian, and Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015) extend the range into the Cenomanian.
Roemer (Reference Roemer1852) compared the overall form of H. alta to Pholadomya ligeriensis d’Orbigny (Reference d’Orbigny1843), which Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015, p. 50) reassigned to Pleuromya. The triangular shell morphology of H. alta is very similar to that of Pholadomya (Bucardiomya) Rollier in Cossman (Reference Cossman1912), which, however, has distinct, even, wide radial costae. A species very similar to H. alta is Pholadomya (Bucardiomya) gigantea Jaitly (Reference Jaitly2013) in the Middle Jurassic of western India. This species is about 60 cm tall and has faint indistinct umbonal radial ribs.
A note to clarify the stratigraphic position of Liopistha [Cardium] elegantulum (Roemer, Reference Roemer1852), the type species of Liopistha: Roemer collected his specimen at the waterfall in the Guadalupe River beyond (“unterhalb” was Roemer’s word; Roemer, Reference Roemer1852, p. 49) New Braunfels, Texas. In this area, either the lower Cenomanian Buda Limestone or the middle to upper Albian Edwards Limestone is exposed, so the age of L. elegantulum is not Santonian as suggested by Dhondt and Jagt (Reference Dhondt and Jagt1988) but Albian or lower Cenomanian.
Order Pholadomyida Newell, Reference Newell1965
Superfamily Pholadomyoidea King, Reference King1844
The superfamily Pholadomyoidea is composed of four phylogenetically related families: Pholadomyidae, Arenigomyidae, Margaritariidae, and Ucumariidae (Carter et al., Reference Carter, Campbell and Campbell2006; Carter et al., Reference Carter2011).
Family Pholadomyidae King, Reference King1844
Remarks
The family Pholadomyidae is composed of the subfamilies Pholadomyinae and Chaenomyinae (Carter et al., Reference Carter2011).
Description
Small- to large-sized, equivalved, very inequilateral, oblong, ovate or subtrigonal, moderately to very inflated bivalves; anterior margin evenly curved, ventral margin gently curved to nearly straight, some with shallow sulcus; ventral-posterior margin curved to truncate; dorsal-posterior margin straight, inclined ventrally or dorsally. Posterior gape narrow to wide, anterior gape where present is narrow. Umbos rounded to subangular, anterior of midline; posterior umbonal carina weak or absent; lunule absent or shallow in most species; escutcheon, if present, shallow, bordered by low ridges.
Ligament external, opisthodetic, quasi-parivincular, attached to low, rounded pseudonymph in escutcheon (Carter et al., Reference Carter2012, p. 148); cardinal plate extends from umbo posteriorly, with a resilium of low wave-like swellings and grooves; hinge edentulous, narrow, and straight in most, some with one or more small knob-like projections on hinge. Pallial sinus depth variable among genera; adductor and pedal muscle scars present. Radial ornament is absent or low, radial folds or ribs, some with tubercles, some with fine radial lines or radial rows of pustules, most with distinct to weak concentric growth rugae. Shell composed of aragonitic, thin, nacreous ostracum but rarely preserved (personal communication, J.G. Carter, 2016); Early Triassic to Holocene.
Subfamily Pholadomyinae King, Reference King1844
Remarks
Subfamily comprises 23 genera (Newell, Reference Newell1969), including Homomya and Pachymya.
Genus Homomya Agassiz, Reference Agassiz1843
Type species
Mactra gibbosa J. Sowerby, 1813 in Sowerby and Sowerby, 1812–Reference Sowerby and Sowerby1846, p. 91; Herrmannsen, Reference Herrmannsen1847, p. 541.
Diagnosis
Equivalved, elongate to ovate, moderately protruding umbos situated anterior of valve midline; dorsoposterior margin straight, downwardly sloping, or slightly upturned; edentulous, opisthodetic ligament attached to pseudonymph in escutcheon; deep pallial sinus; ornamented by concentric growth rugae; radial ribs may be present only in juvenile stages.
Occurrence
Early Triassic to middle Eocene.
Homomya knowltoni (Hill, Reference Hill1893)

Figure 9 (1–3) Homomya knowltoni; holotype USNM 145644; (1) right valve; (2) left valve; (3) dorsal view. (4–8) Homomya tarrantense; (4–6) holotype UMMP 33014; (4) right valve; (5) left valve; (6) dorsal view; (7, 8) paratype UMMP 33015; (7) left valve; (8) dorsal view. (9–12) Homomya kellumi holotype UMMP 32855; (9) right valve; (10) left valve; (11) dorsal view; (12) posterior view. (13–16) Homomya tlahualiloensis; (13–15) holotype UMMP 32858; (13) right valve; (14) dorsal view; (15) left valve; (16) right valve paratype UMMP 32859. (17–19) Homomya comalensis lectotype NPL85566; (15) left valve; (18) anterior view; (19) dorsal view. Scale bars=1 cm.
1893 Pholadomya knowltoni Reference HillHill, p. 30, pl. 2, figs. 1, 2.
1901 Pholadomya knowltoni; Reference HillHill, p. 232.
1940 Pholadomya knowltoni; Reference SmithSmith, p. 611.
1982 Pholadomya knowltoni; Reference Offeman, Lewis, Arnette, Akers, Ganshirt, Martin, Akers and LandryOffeman et al., p. 79, fig. 103.
1896 Pleuromya knowltoni; Reference Stanton and VaughanStanton and Vaughan, p. 23.
1910 Pleuromya knowltoni; Reference BöseBöse, p. 17, 136.
1996 Pleuromya knowltoni; Reference Hovorka, Dutton, Ruppel and YehHovorka et al., p. 16.
1996 Pleuromya knowltoni; Reference Small, Hanson and HauwertSmall et al., p. 4, 8, 13.
1928 Pholadomya(?) knowltoni; Reference AdkinsAdkins, p. 138.
1937 Homomya knowltoni; Reference WhitneyWhitney, p. 128, pl. 13, figs. 1, 2.
2002 Homomya knowltoni; Reference Akers and AkersAkers and Akers, p. 367, p. 368, fig. 350, p. 468.
1961 Homomya tarrantensis Reference PerkinsPerkins, p. 84, pl. 26, figs. 3, 4, 6–8.
2002 Homomya tarrantensis; Reference Akers and AkersAkers and Akers, p. 369, fig. 352, p. 468.
Holotype
Homomya tarrantensis Perkins is UMMP 33014; paratypes: UMMP 33015, 33016, 33017, 33018, and 33019.
Lectotype
Homomya knowltoni NMNH 145644 is here designated lectotype from a set of two cotypes; NMNH 145652 is a paralectotype; specimen label: “Glen Rose beds, Trinity, Cretaceous”; from “section along Colorado River between Austin and Paleozoic contact, Burnet County, Texas.”
Diagnosis
A moderately small species 30–60 mm long, equivalved, inequilateral valves, inflated, prosogyrate beaks, distance between beak and anterior margin averages 35% of the length; anterior margin broadly rounded; ventral margin very slightly arched toward truncated posterior margin; dorsal margin nearly straight; no lunule; shallow narrow escutcheon. Concentric growth rings low, rounded.
Occurrence
The type locality of Homomya knowltoni is in exposures along the Colorado River valley in the Austin area. This is in middle parts of the Glen Rose Formation near the mouth of Bull Creek (Adkins, Reference Adkins1928). The species is reported throughout much of the Glen Rose section in north-central Texas above Mesorbitolina texana (Roemer, Reference Roemer1852) with a diverse set of bivalves and gastropods (Whitney, Reference Whitney1937; Rodgers, Reference Rodgers1967). In Hays County, H. knowltoni is in Unit 3 of the Upper Member, Glen Rose (Watson et al., Reference Watson, Broun, Hunt, Wierman and Smith2017). It also is in the Edwards Formation at Austin on East Bull Creek, just east of old Spice Springs Road Crossing, east of West Loop Bridge, Travis County, Texas. H. knowltoni is considered a marker taxon of the Regional Dense member of the Person Formation northwest and southwest of Austin (Small et al., Reference Small, Hanson and Hauwert1996). However, collections in 2015 and 2016 discovered only Homomya comalensis Whitney Reference Whitney1937. Akers and Akers (Reference Akers and Akers2002, p. 367) reported H. knowltoni in the Glen Rose, Edwards, and Kiamichi formations, central Texas and north Texas, and in the Bluff Formation in southwest Texas (Smith, Reference Smith1940). A single new specimen is from the Kiamichi Formation.
The type specimens of Homomya tarrantensis are from the Benbrook Member of the Goodland Formation exposed along Marys Creek in southwest Fort Worth, Tarrant County, Texas. The total range of H. knowltoni is from the lower Albian Upper Glen Rose to basal upper Albian Kiamichi Formation in Texas.
Description
Specimens are external casts showing exterior shape and ornament; most are incomplete. Ornament of unequal concentric growth rugae, rounded, slightly steeper ventrally, continuous to discontinuous anteriorly to posteriorly. Length ranges from about 38 to 63 mm, height is from 24 to 41 mm, and width is from 17 to 38 mm. The distance from anterior margin to beak is from about 14 mm to about 20 mm. Opisthodetic ligament attached to pseudonymph; dentition and musculature unknown.
Remarks
Types of H. knowltoni, H. comalensis, and H. tarrantense display well-defined escutcheons, in which the conjunction of the two valves forms a low pseudonymph (Fig. 4.3, 4.7–4.9). The dorsal margins of the two valves do not overlap as is the case of Pleuromya. Therefore, this set of species is here allocated to Homomya.
Hill (Reference Hill1893, p. 30) compared H. knowltoni to Pholadomya pleuromayaeformis Choffat (Reference Choffat1888) in Albian deposits near Dombey, Angola, West Africa. Examination of Choffat’s drawings of this African species shows that his species differs from the Texas species by having two to four low, rounded radial ribs extending from beak to ventral margin, typical of Pholadomya. The beaks of P. pleuromayaeformis appear to project higher than those of P. knowltoni and the lunule appears to be distinct. Because of the radial ribs, P. pleuromayaeformis is a separate species.
Stanton and Vaughan (Reference Stanton and Vaughan1896), followed by Böse (Reference Böse1910), Hovorka et al. (Reference Hovorka, Dutton, Ruppel and Yeh1996), and Small et al. (Reference Small, Hanson and Hauwert1996), reassigned Pholadomya knowltoni to Pleuromya without explanation. Subsequently, no one has presented a taxonomic explanation of this generic reassignment. Stanton and Vaughan (Reference Stanton and Vaughan1896) and Böse (Reference Böse1910) listed this species in the assemblage from the basal limestone unit at Cerro de Cristo Rey, which is the Finlay Formation and which they correlated with the Fredericksburg Group in north Texas.
Three specimens in the collections of the Non-vertebrate Paleontology Laboratory, University of Texas Austin, are labeled as “Homomya fredericksburgensis n. sp. W.C.I.” (UT 17010, 17011, 17013). The author is identified as W.C.I. and this species name has never been published, so it is a nomen nudum. Each specimen was collected from the upper part of the Walnut Formation near Belton, Texas. These specimens are here assigned to H. knowltoni.
Homomya tarrantensis is here considered a junior synonym of H. knowltoni because it shares the same morphology and its shape and size are within the range of H. knowltoni (Fig. 4). The position of the umbo of H. tarrantensis is about 25% of the length, and that of H. knowltoni is about 35%. H. tarrantensis is common in the middle and upper parts of the Goodland Formation in the Fort Worth, Texas, area (Perkins, Reference Perkins1961).
Pleuromya aff. P. inconstans Castillo and Aguilera (Reference Castillo and Aguilera1895) is quite similar to H. knowltoni, but the posterior-dorsal carina is distinct. This species is in dark gray limestone near Miquihuana, Tamaulipas, Mexico. Imlay (Reference Imlay1937) tentatively correlated this limestone with the Valanginian Stage by associated ammonites, which in Cuba are Valanginian (Myczynski, Reference Myczynski1977).
Pleuromya orbigniana (Rouillier, Reference Rouillier1845) is in the Berriasian-Valanginian Spilsby Sandstone in northeastern England near the North Sea coast (Woods, Reference Woods1909, p. 256–257). This species is much smaller than H. knowltoni and is ornamented by either radial striae or radial rows of nodes. A similar species, Pleuromya peregrina (d’Orbigny, Reference d’Orbigny1845), is found in lowermost Cretaceous near Moscow. Pleuromya hidensis Hayami (Reference Hayami1959) in Aptian strata of Japan and eastern China is smaller than H. knowltoni and taller relative to its length, with a length-to-height ratio of 1.5.
Pholadomya marcoui Cragin (Reference Cragin1905) in the lowermost Cretaceous Malone Formation in southwest Texas is very similar in form to H. knowltoni except for the six faint radial ribs (Akers and Akers, Reference Akers and Akers2002, p. 359–360, fig. 342). If these species are not related they are convergent morphotypes.
Pleuromya ingramensis Pampe (Reference Pampe1975) is an internal cast from the Middle Albian Basal Nodular Member of the Fort Terrett Formation, Fredericksburg Group, north of Kerrville, Texas. It is not a pleuromyid or pholadomyid. Its raised posterodorsal margin along the hinge line and pronounced grooves along the dorsal side are characteristic of arcid taxa such as Cucullaea.
Homomya comalensis Whitney, Reference Whitney1937
1937 Homomya comalensis Reference WhitneyWhitney, p. 130, pl. 13, figs. 3, 4.
2002 Homomya comalensis; Reference Akers and AkersAkers and Akers, p. 367, p. 368, fig. 350, p. 468.
Lectotype
The single specimen illustrated by Whitney would be the holotype, but has not been found in the collections of the Non-vertebrate Laboratory, the University of Texas. Consequently, a lectotype is selected from among two specimens recovered from the Regional Dense Member of the Person Formation in the road cut at Stone Crossing, New Braunfels, Texas (Porter, Reference Porter2017), Texas Memorial Museum Non-vertebrate Paleontology Laboratory NPL85566. Unnumbered paralectoypes are from the Regional Dense Member of the Person Formation at the exposures where highways Loop 1 and Loop 360 intersect in south Austin and from the basal Bee Cave Member of the Walnut Formation on the east road cut in Loop 360 at Bee Cave Road, Austin (Diehl, Reference Diehl2017).
Diagnosis
A moderate-sized species 30–50 mm long equivalved, very inequilateral, ovate, globose valve, longer than high, slightly higher than wide; beaks low, slightly opisthogyrate, very close to rounded anterior margin, distance between beak and anterior margin averages 15% of the length; ventral margin broadly curved; posterior margin narrowly rounded, slightly gaping, and sloping into flat dorsal margin; no lunule, shallow wide escutcheon. Low, rounded concentric growth rings.
Occurrence
Whitney’s specimen of Homomya comalensis is from the middle Glen Rose Formation: “About 275 feet (84 m) above the base … 2.3 miles (3.7 km) west of Fischer Store, Comal County,” Central Texas (Whitney, Reference Whitney1937, p. 131). Newly discovered specimens are in lime mudstone from the middle Albian Regional Dense Member at the base of the Person Formation in Travis and Hays counties and in the Bee Cave Member of the Walnut Formation in Travis County. Total known range is lower to middle Albian.
Description
Specimens are external casts showing exterior shape and ornament; most are incomplete. Beaks wide, very low, barely projecting above the straight dorsal margin. Ornament of weak, rounded, unequal, discontinuous concentric growth rugae, slightly steeper ventrally. Length ranges from 36 to 55 mm, height is from 19 to 35 mm, and width is from 20 to 29 mm. The distance from anterior margin to beak is from about 5 mm to 11 mm. Posterior gape is slight, not flared. Interior dentition, ligament, and musculature are unknown.
Remarks
Whitney (Reference Whitney1937) distinguished Homomya comalensis from H. knowltoni by its greater width, by its rounded gaping posterior margin, and by posterior ridges bordering escutcheon (Akers and Akers, Reference Akers and Akers2002, fig. 230). H. comalensis is as long as smaller specimens of H. knowltoni, but its height is less and it is slightly wider at the same height than H. knowltoni and H. tarrantensis (Figs. 6, 8). The distance of the beak from the anterior margin of H. comalensis is less than of H. knowltoni margin so the beak-to-length and beak-to-height ratios are lower than for H. knowltoni (Fig. 8).
Homomya kellumi (Perkins, Reference Perkins1961)
1910 Homomya aff. H. ligeriensis Reference BöseBöse, p. 137, pl. 29, figs. 1–4.
1928 Homomya aff. H. ligeriensis; Reference AdkinsAdkins, p. 142.
2002 Homomya aff. H. ligeriensis; Reference Akers and AkersAkers and Akers, p. 369, fig. 352.
1961 Homomya kellumi Reference PerkinsPerkins, p. 82, pl. 24, fig. 5; pl. 15, figs. 1–3, 5, 6.
1967 Homomya kellumi; Reference LokkeLokke, p. 352.
2002 Homomya kellumi; Reference Akers and AkersAkers and Akers, p. 468.
1961 Homomya tlahualiloensis Reference PerkinsPerkins, p. 83, pl. 25, figs. 4, 7–10.
Holotype
Homomya kellumi: holotype UMMP 32855; paratypes: UMMP 32856, 32857. Type locality and age: Sierra Tlahualilo, localities 13, 14, 15, 18, 19, 31, and 51, Coahuila, Mexico, upper interval of Aurora Limestone, Lower Cretaceous. Holotype of Homomya tlahualiloensis: UMMP 32858; paratypes UMMP 32859. Type locality and age: Sierra Tlahualilo, localities 14, 18, 26, and 29, Coahuila, Mexico, upper interval of Aurora Limestone, Lower Cretaceous.
Diagnosis
A large-sized, 65–70 mm long, equivalved species, very inequilateral, inflated, prosogyrate beaks 25% to 30% of the length from the anterior margin; anterior margin broadly rounded; ventral margin broadly arched toward narrowly curved posterior margin; dorsal margin nearly straight; gape extends anteriorly along dorsal commissure that merges into a low ridge up to beak.
Occurrence
Type specimens of H. kellumi and H. tlahualiloensis were collected from the upper part of the upper interval of the Aurora Limestone. Two specimens of H. kellumi were also collected from the Fort Worth Limestone near Fort Worth, Tarrant County, Texas (Perkins, Reference Perkins1961, p. 55). Specimens of Homomya aff. H. ligeriensis were collected at Cerro de Muleros (now Cristo de Rey), New Mexico, west of El Paso, Texas, in units 4–6 with the upper Albian ammonite Mortoniceras equidistans (originally Schloenbachia trinodosa) (Cragin, Reference Cragin1893). This stratigraphic interval is now composed of the upper Albian Smeltertown, Muleros and Mesilla Valley formations (Lucas et al., Reference Lucas, Krainer, Spielmann and Durney2010) and correlate with the Washita Group of North Texas.
Remarks
Perkins (Reference Perkins1961) characterized H. kellumi by “its high beaks and pronounced shoulder anterior to the beaks.” He contrasted H. tlahualiloensis with H. kellumi by the “less pointed and less terminal beaks,” its greater width, and by the absence of an anterior shoulder (Perkins, Reference Perkins1961, p. 83). However, these characters intergrade; the distance between the anterior margin and beak of the former is 17.2 ± 3.14 mm compared to 17.3 ± 2.03 mm in the latter (data of Homomya species described here available as Supplementary Data). The mean widths of both species are within the standard deviation: mean width of H. tlahualiloensis is 33.7 ± 3.23 mm; mean width of H. kellumi is 32.5 ± 3.921 mm. These relative proportions, however, are gradational and an insufficient basis upon which to define species among this variable group of homomyids. The “pronounced shoulder anterior to the beaks” of P. kellumi is a subtle and impersistent feature subject to compaction and distortion. In addition, the species occur together at the same sites in the upper part of the Aurora Limestone at Sierra de Tlahualilo, thus have no stratigraphic or geographic significance. Therefore, they are here judged to be synonyms.
Perkins compared H. kellumi to Homomya bravoensis, which has a lower height-to-thickness ratio (Fig. 4). Homomya bravoensis is an oblong species with a length-to-height ratio of 1.38; its anterior margin is nearly at a right angle to the dorsal margin; whereas H. kellumi is a low triangular form with a length-to-height ratio of 1.34 and apical angle of about 115°.
Although Perkins did not compare H. kellumi to the lower Cenomanian Homomya austinensis Shattuck (Reference Shattuck1903, pl. 16, figs. 1–3), they are quite similar in form, proportions, and size (Table 2). The gape even extends anteriorly along the posterior hinge, which merges into a low ridge, the pseudonymph beneath the beaks. However, H. austinensis differs from H. kellumi by its more broadly rounded, wider posterior margin, which reflects a wider siphon.
Perkins also compared H. kellumi to Homomya aff. H. ligeriensis d’Orbigny (Reference d’Orbigny1845), which has finer growth rugae and is less elongate than H. kellumi. Böse (Reference Böse1910) identified a species at Cerro de Muleros (now Cristo de Rey), New Mexico, west of El Paso, Texas, that he compared to Homomya ligeriensis:
The average shell has an ovate form, transversally extended, somewhat convex, posteriorly extended; anterior margin arcuate passing by a curve into the ventral margin; this margin is also somewhat arcuate; posterior margin is not preserved in any of the specimens but nevertheless the outline of lines for a superficial very regular curve; dorsal margin straight; umbos strong large, prominent, prosogyrate; situated very near the anterior of the dorsal margin. The valves are gaping posteriorly. The ornament on the surface consists of very regular concentric costellae separated by relatively deep grooves.
Our species is very similar to Homomya ligerienses of the Cenomanian at Le Mans, the differences are quite small; in the Mexican form the umbos are smaller and lower and the outline is slightly different. We cannot compare our specimens with the French species because none of our examples are complete.
Number of specimens: 4.
Locality: Cerro Muleros close to the monument marking the Mexican-US border.
Horizon: Subdivisions 4, 5 and 6, horizon with Schloenbachia trinodosa, lower Cenomanian. (Böse, Reference Böse1910, p. 137)
Adkins (Reference Adkins1928, p. 142) described Homomya aff. H. ligeriensis as a “form subrectangular, beaks nearly terminal; prominent somewhat irregular growth lines.” The specimen illustrated by Akers and Akers (Reference Akers and Akers2002, fig. 352) is incompletely preserved and is within the size range of H. kellumi. Because the size, shape, and stratigraphic and geographic ranges of Homomya aff. H. ligeriensis are similar to those of H. kellumi, it is placed in synonymy with H. kellumi.
Ayoub-Hannaa et al. (Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015) reassigned the species Pholadomya ligeriensis to Pleuromya. Pleuromya ligeriensis d’Orbigny (Reference d’Orbigny1845) is well known from Cenomanian-Turonian strata in Europe, North Africa, Nigeria, and Brazil (Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015). P. ligeriensis is about the same size as H. kellumi and H. tlahualiloensis but the length and height of P. ligeriensis are nearly the same, whereas the Comanchean species are longer than high. The valve outline of H. ligeriensis differs from the two Albian species in having a straighter anterior margin and a more steeply inclined posterior margin, and the angle between the dorsal and anterior margins is nearly 90° whereas the same angle of the Albian species is greater than 90°. P. ligeriensis differs from H. kellumi by its strongly prosogyrate beaks and by the right angle between dorsal and anterior margins (Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015).
Homomya washitae Cragin, Reference Cragin1894

Figure 10 (1–3) Homomya washitae Cragin, Reference Cragin1894; plaster cast of WSA 10,444, incomplete specimen, Grayson Marl, Denison, Texas; (1) left valve; (2) dorsal view; (3) anterior view. (4) Homomya bravoensis Böse, holotype IGM 384, RV; (2017 image available at: http://www.unipaleo.unam.mx/public/consulta_publica.php). (5, 10) Homomya budaensis Whitney, Reference Whitney1911, hypotype UMMP 32854, collected by Perkins (Reference Perkins1961); (5) left valve; (10) dorsal view. (6, 8, 9) Homomya cymbiformis Perkins, Reference Perkins1961; holotype UMMP 33012; (6) left valve; (8) right valve; (9) dorsal view. (7) Homomya auroraensis Perkins, Reference Perkins1961, holotype UMMP 32853, left valve. Scale bars=1 cm.
1894 Homomya washita (sic) Reference CraginCragin, p. 59.
1928 Homomya washita (sic); Reference AdkinsAdkins, p. 141.
2002 Homomya washita (sic); Reference Akers and AkersAkers and Akers, p. 469.
1961 Homomya washitae; Reference PerkinsPerkins, p. 40.
Holotype
NMNH PAL 32689; topotype: WSA 10444, the label reads “Homomya washitaensis (sic), railroad cut, 0.5 mi SE of Union Depot, Denison.”
Diagnosis
Shell elongate, broadly inflated, beak at anterior margin; escutcheon a narrow shallow depression merging posteriorly with a straight commissure line posterior to beaks; concentric rugae as flat ridges wider than the interspaces; ribs slope slightly ventrally and widen from beak to ventral margin.
Occurrence
Upper Albian Main Street Limestone and lower Cenomanian Grayson Formation “in the abandoned D. B. and N. O. railway cut about half a mile southeast of the Union depot of Denison, Texas; … on summit of Pawpaw creek bluff southeast and east of Dennison” (Cragin, Reference Cragin1894, p. 60). Specimen WSA 10444 (Non-vertebrate Paleontology Laboratory) was collected from a railroad cut in the Grayson Formation 0.5 mi southeast of the Union Depot, Denison, Grayson County, Texas.
Cragin’s Description
“Shell large, curved-oblong, closed or nearly closed anteriorly and closed along the dorsal margin back of the beaks, obliquely truncated and gaping posteriorly, the breadth usually a little greater than the vertical dimension from hinge-margin to ventral margin, the length equal to somewhat more than one and a half times the breadth, the greatest breadth being about half way between the beaks and the mid-region; beaks nearly terminal, low, swollen, obtusely tangent, their bases long in the direction of the length of the shell and rising at a very low angle from their posterior origin to their rather broadly rounded summits; surface marked only with concentric growth-lines and undulations, and sometimes showing distally two or three broad growth-zones or stage” (Cragin, Reference Cragin1894, p. 59). Length 120 mm, height 92 mm, width 80 mm.
Remarks
A plaster cast of the incomplete holotype NMNH 32689 is curated in the Non-vertebrate Paleontology Laboratory at the University of Texas (WSA10444). The posterior portion is missing and the anterior, dorsal, and ventral margins are intact. A lunule is not developed because the umbo is terminal; the escutcheon is directly posterior to the beaks and is a narrow and shallow depression merging posteriorly with a straight commissure line. The concentric rugae are incompletely preserved as flat ridges wider than the interspaces; the ribs slope slightly ventrally and widen from beak to ventral margin.
Cragin (Reference Cragin1894) originally spelled the species name “washita,” which is an incorrect Latinization and therefore not an inadvertent error (ICZN, 2000 Code, Article 32.5, p. 39). The correct Latinized species name is “washitae” as a noun in the genitive case (ICZN, 2000, Code, Article 11.9, p. 13–14).
Homomya austinensis (Shattuck, Reference Shattuck1903)
1903 Homomya austinensis Reference ShattuckShattuck, p. 28, pl. 16, figs. 1–3.
1928 Homomya austinensis; Reference AdkinsAdkins, p. 141.
1961 Homomya austinensis; Reference PerkinsPerkins, p. 42.
2002 Homomya austinensis; Reference Akers and AkersAkers and Akers, p. 468.
Holotype
Homomya austinensis holotype: NMNH PAL 315995; topotypes: NPL 62211, UT 30035, UT32168, UT 32299, UT 32308, UT 32309.
Diagnosis
A moderately large, 50–65 mm long, equivalved, inequilateral, ovate species about one and a half times long as tall, relatively narrow width, weakly inflated, prosogyrate beaks 15% to 20% of the length from the anterior margin; anterior margin broadly rounded; ventral margin merging smoothly into curved posterior margin; dorsal margin nearly straight with a distinct escutcheon.
Occurrence
Lower Cenomanian Buda Limestone, Austin, Texas.
Description
Inflated, equivalved, inequilateral shell; anterior margin rounded, ventral margin broadly rounded, posterior margin narrowly rounded, dorsal margin slightly inclined toward beak, beak about one-fourth the length toward the anterior. Beaks nearly touching, slightly prosogyrate; no lunule, anterior hinge margin short; posterior hinge margin a low ridge at commissure, bordered by shallow groves that are separated from umbo by faint ridges. Slightly gaping. Ornamented by low, rounded, concentric growth rugae, wider than interspaces.
Remarks
The outline and compact thickness of this species are more similar to H. kellumi than to other species in the Buda Limestone. Its escutcheon is shallow, fading out into the posterior gape and with a commissural pseudonymph ridge. Shattuck (Reference Shattuck1903, p. 28) differentiated H. austinensis from H. vulgaris in “that it is much shorter and broader in proportion to its thickness. It is also much more compressed.” H. austinensis is nearly indistinguishable from the French and Brazilian P. ligeriensis (d’Orbigny, Reference d’Orbigny1845, in Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015). The length-to-height ratio of H. austinensis is 1.44 and that of P. ligeriensis is 1.52; H. austinensis is the larger species; the beak positions are virtually the same. P. ligeriensis is a cosmopolitan member of the Mediterranean to eastern South American provinces (Ayoub-Hannaa et al., Reference Ayoub-Hannaa, Bengston, Fürsich and de Jesus Andrade2015), and H. austinensis is a member of the Caribbean Province.
Homomya vulgaris Shattuck, Reference Shattuck1903
1903 Homomya vulgaris Reference ShattuckShattuck, p. 29, pl. 16, figs. 4–5.
1928 Homomya vulgaris; Reference AdkinsAdkins, p. 141.
2002 Homomya vulgaris; Reference Akers and AkersAkers and Akers, p. 371, 372, 469, fig. 354.
Holotype
H. vulgaris holotype: NMNH PAL 315996; topotypes: NPL 62219, WSA 5013, UT 35380B.
Diagnosis
An equivalved, very inflated, elongate shell much longer than tall, with subparallel dorsal and ventral margins; with a shallow, elongate escutcheon.
Occurrence
Uppermost interval of the lower Cenomanian Buda Limestone, upper Washita Group in the Austin, Texas, area; in northern Mexico it is in the upper part of the upper Albian Aurora Limestone.
Description
Large-sized, 70–100 mm long elongate shell, widely inflated; anterior margin broadly rounded, ventral margin broadly curved, posterior margin narrowly rounded, merging with dorsal margin that slopes up to umbo. Umbos broadly incurved, slightly prosogyrate; no lunule; escutcheon wide, shallow, merging smoothly with valve margins. Ornamented by rounded growth rugae wider than interspaces, widening and diverging posteriorly.
Remarks
Perkins (Reference Perkins1961) distinguished H. vulgaris from his new species Homomya auroraensis by several relative differences in shape: (1) the dorsal margin of H. auroraensis is more concave than H. vulgaris; (2) the posterior margin of H. auroraensis is more rounded; (3) beaks of H. auroraensis are more posterior from the anterior margin and less prosogyrate; and (4) the height-to-thickness ratio of H. auroraensis is less. These differences are subject to compaction and distortion. Only the latter two properties can be quantified and the differences are slight based on the small number of specimens. In a subsequent section, H. auroraensis is considered a junior synonym of Homomya budaensis.
H. vulgaris occurs with Homomya budaensis in the Buda Limestone in the Austin, Texas, area. They differ, however, by the valve outlines. H. vulgaris is an elongate shell and its dorsal margin is subparallel with the ventral margin. H. budaensis has a concave dorsal margin and its posterior is expanded and flares dorsally. This difference suggests that soft parts such as siphons or gills were larger in H. budaensis.
Homomya bravoensis Böse, Reference Böse1910
Figure 10.4
1910 Homomya bravoensis Reference BöseBöse, p. 136, pl. 29, figs. 5, 6.
1928 Homomya bravoensis; Reference AdkinsAdkins, p. 141.
1961 Homomya bravoensis; Reference PerkinsPerkins, p. 83.
2002 Homomya bravoensis; Reference AdkinsAkers and Akers, p. 367, p. 368, fig. 350.
Holotype
Homomya bravoensis holotype IGM 384, Colección Nacional de Paleontología (IGM), Universidad Nacional Autónoma de México; upper Albian, Böse’s unit 3 with Ceratostreon [Exogyra] texana, along the Southern Pacific and Bisbee Railroad, near Rio Bravo, Chihuahua, Mexico (source: http://documents.mx/documents/fosiles-tipo-mexicanos.html; image available at: http://www.unipaleo.unam.mx/public/consulta_publica.php).
Diagnosis
Elongate, ovate shell, beaks at anterior margin, posterior margin truncate, inclined, rounded, gaping; ornament of low, rounded concentric rugae.
Occurrence
Type specimens were collected at Cerro de Muleros (now Cerro de Cristo Rey) next to the Southern Pacific and Bisbee railroad near the Rio Bravo, New Mexico, west of El Paso, Texas, in the Del Norte Formation. It is also known in the Edwards Formation west of Round Rock, Williamson, and in Bell and Parker counties, Texas, in Walnut to Goodland formations, Fredericksburg Group (Adkins, Reference Adkins1928). It is reported in the Benevides Formation in the southern part of the Quitman Mountains (Albritton and Smith, Reference Albritton and Smith1965). It also was collected from the University Mesa Marl near Fort Stockton, I-10, Pecos County, Texas (NPL 74414). In Mexico, it is in the lower part of the Upper Aurora Limestone. It ranges from middle Albian to lower upper Albian.
Description
According to Böse:
Shell moderately oval, transversally extended, very convex, posteriorly extended; anterior margin short, obliquely curved ventrally, posterior margin longer, regularly curved; dorsal margin nearly straight, long, subparallel with ventral margin which is slightly arcuate; umbo strong, elevated, globose, curved towards anterior and below; positioned nearly at the extreme anterior of the valve. The valves are open at the posterior margin. The ornament consists of undulations and concentric grooves. Dimensions: length 55 mm, height 40 mm, width of one valve 18.2 mm.
Our species rather resembles Homomya vulgaris Shattuck in its general form, but is distinguished by the position of the umbos, which in our species are positioned very near the anterior margin and by the less extended form. The species is rather rare and is encountered only in the lower part of the Cretaceous deposits at Cerro Muleros; Stanton and Vaughan cited the same horizon of Pleuromya knowltoni Hill; we have not found this species at Cerro Muleros. (Freely translated from Böse, Reference Böse1910, p. 136.)
According to Adkins:
Form roughly elongate rectangular, rounded, oblique, quite inflated, beaks subterminal, proportionately less tall and more inflated than H aff. ligeriensis; reduced concentric lines. (Adkins, Reference Adkins1928, p. 142)
Remarks
Böse noted that the overall form of H. bravoensis is similar to that of Homomya vulgaris Shattuck, but distinguished it by the position of the umbo, which is flush with the anterior margin. The escutcheon is elongate, ovate, but covered in lime mud (personal communication, J. Alvarado Ortega, 2017, Nacional de Paleontología del Instituto de Geología de la UNAM). H. bravoensis is from strata of the same age as L. (Sergipemya) alta but differs by its low umbo and oblong form; L. alta is characterized by a tall umbo very close to the anterior margin. The length and height of L. alta are subequal but the types of H. bravoensis are longer than high. H. bravoensis is similar to H. budaensis but tends to be shorter relative to its height, and the distance of the beak to the anterior margin of H. bravoensis is less than 10% and that of H. budaensis is nearly 30%.
Homomya budaensis Whitney, Reference Whitney1911
1911 Homomya budaensis Reference WhitneyWhitney, p. 15, pl. 3, figs. 1, 2, pl. 4, figs. 1, 2.
1928 Homomya budaensis; Reference AdkinsAdkins, p. 141.
1961 Homomya budaensis; Reference PerkinsPerkins, p. 81, pl. 24, figs. 4, 6, 7.
2002 Homomya budaensis; Reference Akers and AkersAkers and Akers, p. 369, 468, fig. 352.
1961 Homomya auroraensis Reference PerkinsPerkins, p. 81, pl. 24, figs. 1–3.
2002 Homomya auroraensis; Reference Akers and AkersAkers and Akers, p. 468.
Holotype
Homomya auroraensis holotype UMMP 32853. Reference specimens in the Non-vertebrate Paleontology Laboratory: UT 32300, UT 30019, UT 32298, UT 32294.
Lectotype
Homomya budaensis lectotype UT 32300; topotypes: UT 30019, UT 32298, UT 32294.
Diagnosis
Medium to large, 75–100 mm long, equivalved, greatly inflated shell; prosogyrate beaks nearly even with anterior margin, anterior margin truncated, ventral margin concave, posterior margin expanded, dorsal margin concave, dorsal-posterior corner inclined upward, higher than anterior margin.
Occurrence
Whitney’s specimens are from the upper 2 m of the Buda Limestone at Shoal Creek, Austin, Travis County, Texas. Perkins’s specimens of H. budaensis are from seven sites in the upper Aurora Limestone in the P. kellumi zone, which correlates with the Washita Group of north Texas (Scott et al., Reference Scott, Benson, Morin, Shaffer and Oboh-Ikuenobe2003). The holotype of H. auroraensis co-occurs with H. budaensis at Perkins’s site 18 in the H. kellumi zone of the upper Albian Aurora Limestone, Ojo de Agua, Sierra de Tlahualilo, Coahuila, Mexico. The total known range of H. budaensis is middle Albian to lower Cenomanian.
Description
“Shell large, globose, equivalved, inequilateral, greatly produced and broadened posteriorly; anterior margin short, curved; ventral margin deeply curved; posterior margin broadly rounded; dorsal margin curved, subparallel with ventral; umbos globose, approximate, elevated, curved inward and slightly forward, situated anteriorly; shell closed anteriorly, gaping posteriorly. Surface marked by lines of growth, which vary considerably in size” (Whitney, Reference Whitney1911, p. 15–16). The valves gape posteriorly; beaks are low, large, subterminal in position; surface ornament of irregular, coarse, concentric growth lines (Perkins, Reference Perkins1961, p. 81–82).
H. budaensis: length=~110 mm; height=60 mm; width=61 mm (Buda Formation; Whitney, Reference Whitney1911, p. 16); mean length=88.37 mm; mean height=48.0 mm; mean width=52.1 mm; mean beak distance to anterior margin 10.8 mm (Aurora Limestone). H. auroraensis: length=85.8 mm; height=46.9 mm; width=44 mm (Table 1).
Remarks
Whitney distinguished H. budaensis from H. vulgaris by its overall shape, the more curved dorsal and ventral margins, the more expanded, flaring posterior margin, the greater width, and the variable growth rings. The dorsal-posterior corner flares up and the posterior margin is straight and tilted upward. Perkins (Reference Perkins1961, p. 81) distinguished H. auroraensis from H. budaensis by its “more concave dorsal border, a greater height in relation to the thickness of the shell and in not having the broad posterior, vertical expansion.” Perkins also noted that the growth rings of H. auroraensis seemed less coarse. The height-to-width ratio of the holotype of H. auroraensis is 1.2 and that of eight specimens of H. budaensis is 0.923 ± 0.066 (Fig. 6). Furthermore, the shapes and dimensions are very similar except that the beak-to-anterior-margin distance is greater in H. auroraensis, 19 mm compared to 10.8 mm in budaensis. Because these two species are morphologically similar and because H. auroraensis occurs together with H. budaensis in the H. kellumi zone, Aurora Limestone, the species are synonymized.
Perkins (Reference Perkins1961) identified four specimens as Homomya budaensis from the Texigryphaea mucronata Zone in the lower part of the upper member of the Aurora Limestone, Coahuila, Mexico: UMMP 32972, 32974, 32976, and 32977. Two relatively complete specimens are indistinguishable from the holotype. This extends the range of H. budaensis from the upper part of the Washita Group–equivalent Aurora Limestone into the lower part of the Fredericksburg Group–equivalent.
Homomya cymbiformis Perkins, Reference Perkins1961
1961 Homomya cymbiformis Reference PerkinsPerkins, p. 83, pl. 26, figs. 1, 2, 5.
2002 Homomya cymbiformis; Reference Akers and AkersAkers and Akers, p. 367, 468, fig. 350.
Holotype
Homomya cymbiformis holotype UMMP 33012, an external cast; paratype: UMMP 33013, cast of incomplete specimen.
Diagnosis
“A Homomya distinguished by the strong central concavity of the dorsal border and its cymbiform profile” (Perkins, Reference Perkins1961, p. 83).
Occurrence
The type locality of H. cymbiformis is the lower part of the Marys Creek Member of the Goodland Formation in Fort Worth, Tarrant County, Texas, Fredericksburg Group, middle Albian. An additional specimen in the Non-vertebrate Paleontological Laboratory, University of Texas, is from the Weno Formation, Washita Group on Old Mansfield Road, Tarrant County, Texas (UT 35247), which extends the species range into the middle part of the upper Albian Washita.
Description
A medium-sized, 60–70 mm long, equivalved, elongate, compressed shell characterized by a concave dorsal margin and dorsally arched, expanded posterior margin resulting in the so-called cymbiform margin. Prosogyrate umbos close to the anterior margin, distance about 25% of length. Ornament of simple concentric, co-marginal, rounded growth rugae.
Remarks
H. cymbiformis is similar in outline to H. budaensis and their ranges overlap from uppermost middle Albian to middle upper Albian. The valves of H. cymbiformis are more equidimensional than H. budaensis; the length-to-height ratio is 1.48 versus 1.69. H. cymbiformis is higher than wide, 1.25 versus 1.08, and its volume is less than half of the volume of H. budaensis: 73,419 mm3 versus 164,760 mm3.
Species doubtfully attributed to Homomya
Three Comanchean species were placed in the genus Homomya by their authors. However, their valve morphologies differ substantially from species of Homomya, and their internal features are not known because the type specimens are preserved as external molds. Therefore, these species are provisionally retained in “Homomya.”
“Homomya” jurafacies Cragin, Reference Cragin1893

Figure 11 (1, 2) Homomya jurafacies Cragin, Reference Cragin1893; lectoparatype UT 17236, left valve, right valve. (3, 4) Homomya solida, holotype UT 21724; (3) right valve; (4) dorsal view. (5–8) Liopistha (Sergipemya) alta (Roemer, Reference Roemer1852), holotype STIPB-Roemer-79, Bonn Museum; (5) drawing of right valve (Roemer, Reference Roemer1852, pl. 6, fig. 11); (6) right valve, note minute nodes on concentric rugae (photo by Georg Heumann, Steinmann-Institut, Paläontologie); (7) left valve, minute granule-like nodes on concentric rugae create a granular texture; (8) posterior view showing gape and elongate oval escutcheon on dorsal margin. (9, 10) Panopea henselli; (9) left valve; (10) dorsal views. (11) Homomya comalensis in situ, Regional Dense member, basal Person Formation, Stone Crossing outcrop, New Braunfels, Texas; dotted lines approximate bedding. (1–10) Scale bar=1 cm; (11) scale bar=5 cm.
1893 Homomya jurafacies Reference CraginCragin, p. 191, pl. 39, figs. 1, 2.
1928 Homomya jurafacies; Reference AdkinsAdkins, p. 140.
1937 Liopistha (Psilomya) jurafacies; Reference WhitneyWhitney, p. 143, pl. 28, figs. 1, 2.
1952 Liopistha (Psilomya) jurafacies; Reference WhitneyWhitney, p. 68.
2002 Psilomya jurafacies; Reference Akers and AkersAkers and Akers, p. 388, 389, figs. 371, 372.
Lectotype
Two syntypes collected by J. Taff are cataloged as UT 17236, Non-vertebrate Paleontology Laboratory, which are from Hickory Creek, Burnet County, Texas. The smaller specimen is better preserved and is here designated the lectotype, and the larger specimen becomes the paralectotype. Topotypes are UT 8307, UT 8309, UT 17236.
Occurrence
Lower Albian Glen Rose Formation in Burnet and Bosque counties, central Texas.
Description
A large, equivalved, ovate-trapezoidal shell with a tall, angular, prosogyrate umbo, average length-to-height ratio 1.27 ± 0.106; umbo close to anterior margin about 27% of the length; anterior margin evenly rounded, ventral margin broadly curved, posterior margin nearly straight, inclined about 150° to the dorsal margin; lunule short, narrow; escutcheon longer, shallow; pallial line a faint shallow groove at both anterior-ventral and posterior-dorsal margins. Dentition not evident on type specimens.
Remarks
“Homomya” jurafacies is very similar to “Homomya” solida. The type specimens of “H.” jurafacies were collected from the Glen Rose Formation in ‘the Alternating beds’ from central Texas in Burnet and Bosque counties and the types of “H.” solida are from the Glen Rose formation in Parker, Hood, Erath, and Travis counties, north Texas. The small number of available specimens is not a statistically significant population; however, their measurements are quite similar: the length of four specimens of “H.” jurafacies ranges 73–88 mm, height 53–69 mm, articulated thickness 51–57.2 mm, and the ratio of umbo distance from anterior margin to length is 0.18–0.32; a single specimen of “H.” solida is 81.7 mm long, height 69 mm, thickness 57.2 mm, with the ratio of umbo distance to length 0.35. The posterior margin of “H.” jurafacies is straighter and more truncate than that of “H.” solida.
Whitney (Reference Whitney1937, Reference Whitney1952) reclassified this species as well as “H.” solida in Psilomya White; however, Psilomya is a member of Poromyidae Dall, which is characterized by radial striae, a single cardinal tooth, an external ligament, and no posterior gape. Neither “H.” jurafacies nor “H.” solida bears these features. Although Whitney (Reference Whitney1937) described two cardinal teeth in both valves of “H.” jurafacies, these are not visible on the internal mold she illustrated (Whitney, Reference Whitney1937, pl. 28, fig. 2). Molds of teeth are not visible in specimens UT 8307, UT 8309, and UT 17236. Thus, both species are here provisionally retained in the genus “Homomya.”
‘Homomya’ solida Cragin, Reference Cragin1893
1893 Homomya solida Reference CraginCragin, p. 191, pl. 39, figs. 3, 4.
1928 Homomya solida; Reference AdkinsAdkins, p. 141.
1937 Liopistha (Psilomya) solida; Reference WhitneyWhitney, p. 141, pl. 25, figs. 1, 2; 1952, p. 69.
1952 Liopistha (Psilomya) fletcheri Reference WhitneyWhitney, p. 68, pl. 16, figs. 3, 4.
2002 Psilomya solida; Reference Akers and AkersAkers and Akers, p. 390, 471, figs. 373, 374.
Holotype
“Homomya” solida holotype BEG-UT 21724; this single specimen becomes the holotype by original designation. Three additional specimens in the Non-vertebrate Paleontology Laboratory collections are: UT 38950, “not illustrated, Granbury, Brazos bluffs at railroad bridge, Texas, Stone collector”; UT 34126, Parker County; and UT 17296. The type specimen of Liopistha fletcheri is unknown.
Occurrence
Lower Albian Glen Rose Formation in north and central Texas. The holotype specimen label states: “syntype, Hinor, Parker Co., Tx, Glen Rose, Taff collector.” Whitney collected L. fletcheri from the Salenia texana zone, Lower Member, Glen Rose Formation at Julian Creek, Bandera, Texas.
Remarks
The posterior margin of this large, ovate shell is rounded in contrast to the straight margin inclined toward dorsal margin of ‘Homomya’ jurafacies. Whitney (Reference Whitney1952) distinguished Liopistha fletcheri from “H.” solida by the overall shape: “a more prolonged posterior, a concave posterior cardinal margin and a more rounded ventral margin” (p. 69). These subtle differences are evident by comparing photographed specimens but have not been substantiated by statistical analysis. L. fletcheri is larger but the length-to-height and height-to-width ratios are close to those of “H.” solida and do not support the definition of a separate species. The generic assignment of “H.” solida is indeterminate because dentition of this species is unknown and radial ornament is not evident.
Megaorder Solenata Dall, Reference Dall1889
Order Hiatellida Carter in Carter et al., Reference Carter2011
Superfamily Hiatelloidea Gray, Reference Gray1824
Family Hiatellidae Gray, Reference Gray1824
Subfamily Panopeinae Bronn, Reference Bronn1862
Genus Panopea Ménard de la Groye, Reference Ménard de la Groye1807
Type species
Mya glycimeris Born, Reference Born1778 (ICZN, 1986).
Description
Medium to large, equivalved, quadrate to trapezoidal, elongate, gaping valves; ligamental nymphs tall; one or two small cardinal teeth; wide pallial sinus. The generic name, Panopea, has been conserved (ICZN, 1986).
Panopea henselli (Hill, Reference Hill1893)
1893 Pleuromya (?) henselli Reference HillHill, p. 31, pl. 4, figs. 1, 2.
1967 Pleuromya (?) henselli; Reference RodgersRodgers in Lokke, p. 127.
1928 Panope (sic) henselli; Reference AdkinsAdkins, p. 170.
1924 Panopea henselli; Reference GilletGillet, p. 233.
2002 Panopea henselli; Reference Akers and AkersAkers and Akers, p. 473.
1901 Pholadomya henselli; Reference HillHill, p. 161.
1937 Panopea hilli; Reference WhitneyWhitney p. 172, pl. 15, figs. 2, 3.
Lectotype
Panopea henselli two cotypes NMNH 145646; here the smaller is designated the lectotype.
Diagnosis
An elongate shell, subcentral beaks, anterior and posterior margins evenly rounded, slight posterior gape; ornament of low, rounded commarginal growth rugae.
Occurrence
Lower Albian, Somervell County, Texas, about 12 m below top of Glen Rose.
Description
Equivalved, elliptical, elongate valve, umbo subcentral, not prominent; anterior margin rounded, posterior margin subrounded, gaping slightly; angle around beak between dorsal anterior and posterior margins 135°–140°. Ornamented by simple rounded commarginal growth rugae. The average length of the two specimens is 76.1 mm; height is 46.95 mm; thickness is 33.1 mm.
Remarks
Hill (Reference Hill1893) questioned the generic assignment of this species but created this species because it is characteristic of the Glen Rose Formation. Hill noted the similarities between P. henselli and Thracia myaeformis White (Reference White1880). Similar species in Texas Comanchean strata are Panopea texana Shumard Reference Shumard1854 at ‘camp 4 Cross Timbers,’ and Panopea newberryi Shumard Reference Shumard1860 from the Edwards Limestone in north Texas. Both species are illustrated by hand drawings; both are relatively higher than P. henselli. Roemer (Reference Roemer1852, p. 45) noted an indeterminate Panopaea (sic) sp. together with Mesorbitolina texana (Roemer) near Pedernales that he compared to Panopea regularis d’Orbigny (Reference d’Orbigny1843). Panopea sellardsi Whitney Reference Whitney1952 from the middle part of the Glen Rose Formation in central Texas is much smaller than P. henselli, narrower posteriorly, and relatively more inflated.
Panopea is a common genus in pre-Albian strata in Switzerland and in Argentina (Weaver, Reference Weaver1931). Panopea dupiana d’Orbigny Reference d’Orbigny1843 is about twice as long as high and the umbo is closer to the anterior margin than in P. henselli. Panopea neocomiensis (Leymerie, Reference Leymerie1842) is ornamented by very fine radial striae. Modern Panopea bivalves have aragonite shells that dissolve quickly following death, leaving external molds, which are filled with fine crystalline carbonate. Because of this taphonomy, the resulting casts show mainly external characters.
Conclusions
Species of Pholadomyidea are common and relatively diverse in Albian–lower Cenomanian Comanchean strata in Texas and northern Mexico. Some 22 species have been classed with Pholadomya, Homomya, Pachymya, Myopholas, Panopea, and Pleuromya. Two genera most commonly reported are Homomya, order Pholadomyoida, superfamily Pholadomyoidea and Pleuromya, order Pholadida, superfamily Pleuromyoidea. Since 1852, more than a dozen species have been identified as either Homomya or Pleuromya. Because valve morphologies of the two genera are similar in many ways, casts are difficult to separate. Statistical analysis of height and width objectively differentiates species. Eight species are retained in Homomya, four are synonymized with these; two species are provisionally retained in “Homomya” although they differ significantly.
Homomyid species in Texas and Mexico range from lower Albian to lower Cenomanian. They are found in each of three Gulf Coast Comanchean Series groups: upper Aptian to lower Albian Trinity Group, middle to basal upper Albian Fredericksburg Group, and upper Albian to lower Cenomanian Washita Group. They evolved across this interval, and their ranges vary from one to two million years to up to eight million years. Two sets of species form two clades, the smaller-sized H. knowltoni, H. tarrantensis, H. tlahualiloensis, H. kellumi (Fig. 4) and the larger-sized set of H. cymbiformis, H. austinensis, H. vulgaris, H. budaensis, and H. auroraensis. Two end-member morphotypes are represented by the ‘streamlined’ Homomya knowltoni, which is an elongate, slightly inflated form with a relatively high umbo, and the cylindrical Homomya budaensis, which is a very elongate, tubular, inflated form with a very low umbo. These infaunal suspension feeders occupied offshore calcareous mud and carbonate shelf substrates.
Acknowledgments
G. Heumann, Sammlungsverwaltung Paläontologie & Goldfuß-Museum, Steinmann-Institut, Paläontologie, Bonn, kindly provided new photographs of Roemer’s type specimen of Homomya alta. M.S. Florence, Museum Specialist, Smithsonian Institution, arranged loan of Hill’s types of Pholadomya knowltoni and Pleuromya (?) henselli. A. Molineux and A. Thompson, Non-vertebrate Paleontological Laboratory, University of Texas, hosted our visit to the museum and facilitated loan of M.I. Whitney’s specimens and access to the complete Cretaceous collections. D. Miller, University of Michigan Museum of Paleontology, hosted our visit to the museum and arranged loan of Perkins’s types. New photographs of the holotype of Homomya bravoensis were prepared by J. Alvarado Ortega, Chief of Collections, and his staff at Nacional de Paleontología del Instituto de Geología de la UNAM. J.G. Carter graciously shared unpublished descriptions of the generic and suprageneric categories included here. The University of Tulsa Geosciences graduate students B. Diehl, A. Porter, and X. Tan helped collect new specimens from the Person Formation. Precision Stratigraphy Associates provided financial support for this project, and the University of Tulsa Geosciences Department provided laboratory facilities and financial support.
Accessibility of supplemental data
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.40s3m.












