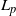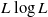1. Introduction and main results
1.1. Description of the model
Branching Brownian motion (BBM) is a spatial population model in which individuals (referred to as particles) move in space according to the law of Brownian motion and reproduce themselves at spatially dependent rates. The particular process that we are going to consider may be described as follows.
It starts with a single particle at time 0 whose spatial position at time
 $t \geq 0$
until the time it dies is given by
$t \geq 0$
until the time it dies is given by
 $X_t$
, where
$X_t$
, where
 ${({X_t})_{t \ge 0}}$
is distributed like a standard Brownian motion. We let T’ and
${({X_t})_{t \ge 0}}$
is distributed like a standard Brownian motion. We let T’ and
 $T_0$
be two random times that are independent conditional on
$T_0$
be two random times that are independent conditional on
 ${({X_t})_{t \ge 0}}$
and satisfy
${({X_t})_{t \ge 0}}$
and satisfy
 $\mathbb{P}(T' > t \!{\mid}\! (X_s)_{s \geq 0}) = \mathrm{e}^{- \beta
t}$
and
$\mathbb{P}(T' > t \!{\mid}\! (X_s)_{s \geq 0}) = \mathrm{e}^{- \beta
t}$
and
 $\mathbb{P}(T_0 > t \!{\mid}\! (X_s)_{s \geq 0}) = \mathrm{e}^{- \beta_0
L_t}$
, where
$\mathbb{P}(T_0 > t \!{\mid}\! (X_s)_{s \geq 0}) = \mathrm{e}^{- \beta_0
L_t}$
, where
 $\beta \geq 0$
and
$\beta \geq 0$
and
 $\beta_0 \geq 0$
are some constants such that
$\beta_0 \geq 0$
are some constants such that
 $\beta$
and
$\beta$
and
 $\beta_0$
are not both 0 and
$\beta_0$
are not both 0 and
 $(L_t)_{t \ge 0}$
is the local time at 0 of
$(L_t)_{t \ge 0}$
is the local time at 0 of
 ${({X_t})_{t \ge 0}}$
. Note that almost surely
${({X_t})_{t \ge 0}}$
. Note that almost surely
 $X_{T_0} =0$
and
$X_{T_0} =0$
and
 $X_{T'} \neq 0$
.
$X_{T'} \neq 0$
.
At time
 $T' \wedge T_0$
, the initial particle dies and is replaced with a random number of new particles. If
$T' \wedge T_0$
, the initial particle dies and is replaced with a random number of new particles. If
 $T_0 < T'$
, then the number of new particles follows some given distribution
$T_0 < T'$
, then the number of new particles follows some given distribution
 ${(q_n)_{n \geq 1}}$
. Otherwise it follows a different distribution
${(q_n)_{n \geq 1}}$
. Otherwise it follows a different distribution
 ${(p_n)_{n \geq 1}}$
. We assume that
${(p_n)_{n \geq 1}}$
. We assume that
 $q_1\neq 1$
and
$q_1\neq 1$
and
 $p_1 \neq 1$
.
$p_1 \neq 1$
.
All the new particles, independently of each other and of the past, then stochastically repeat the behaviour of their parent starting from position
 $X_{T' \wedge T_0}$
. That is, they move like Brownian motions, die after random times giving births to new particles, etc.
$X_{T' \wedge T_0}$
. That is, they move like Brownian motions, die after random times giving births to new particles, etc.
Note that all the particles always produce at least one child upon their death, ruling out the possibility of population extinction.
An equivalent description (up to indistinguishability) would be to say that, after a random time T such that
 $\mathbb{P}(T > t \!{\mid}\! (X_s)_{s \geq 0}) = \mathrm{e}^{- \beta_0 L_t - \beta t}$
, the initial particle dies, and at position x where it died, it is replaced with a random number A(x) of new particles where, for
$\mathbb{P}(T > t \!{\mid}\! (X_s)_{s \geq 0}) = \mathrm{e}^{- \beta_0 L_t - \beta t}$
, the initial particle dies, and at position x where it died, it is replaced with a random number A(x) of new particles where, for
 $n \geq 1$
,
$n \geq 1$
,
 \begin{equation}
\mathbb{P}(A(x) = n) =
\begin{cases}
q_n& \text{if } x = 0, \\
p_n& \text{if } x \neq 0{,}
\end{cases}
\end{equation}
\begin{equation}
\mathbb{P}(A(x) = n) =
\begin{cases}
q_n& \text{if } x = 0, \\
p_n& \text{if } x \neq 0{,}
\end{cases}
\end{equation}
and these new particles then stochastically repeat the behaviour of their parent starting from x. Thus the model can be thought of as the BBM model with spatially inhomogeneous branching rate
 $\beta_0 \delta_0({\cdot}) + \beta$
, where
$\beta_0 \delta_0({\cdot}) + \beta$
, where
 $\delta_0({\cdot})$
is the Dirac delta function, and spatially inhomogeneous offspring distribution given by (1), since informally we may say that
$\delta_0({\cdot})$
is the Dirac delta function, and spatially inhomogeneous offspring distribution given by (1), since informally we may say that
 $L_t = \smash{\int_0^t} \delta_0(X_s) \,\mathrm{d}s$
(this can be made formal via the theory of additive functionals of Brownian motion).
$L_t = \smash{\int_0^t} \delta_0(X_s) \,\mathrm{d}s$
(this can be made formal via the theory of additive functionals of Brownian motion).
Our model combines in a natural way the classical BBM model with constant branching and the BBM model with a single catalytic point. The first one has been studied for many decades and numerous asymptotic results are available (let us mention [Reference Bramson3], [Reference Lalley and Sellke15], [Reference McKean18], and [Reference Roberts19], among many others). The catalytic model has been given less attention and has mostly been studied either in the discrete space (see e.g. [Reference Bulinskaya4] or [Reference Carmona and Hu5]) or in the context of superprocesses (see e.g. [Reference Dawson and Fleischmann6] or [Reference Englander and Turaev8]). For a general review of the topic one may refer to [Reference Klenke, Gorostiza and Ivanoff13].
1.2. Some notation
Using common practice we label the initial particle by
 $\emptyset$
and all the other particles according to the Ullam–Harris convention, so that, for example, particle ‘
$\emptyset$
and all the other particles according to the Ullam–Harris convention, so that, for example, particle ‘
 $\emptyset32$
’ is the second child of the third child of the initial particle.
$\emptyset32$
’ is the second child of the third child of the initial particle.
For two particles u and v we shall write
 $u < v$
if u is an ancestor of v, so for example
$u < v$
if u is an ancestor of v, so for example
 $\emptyset < \emptyset3 < \emptyset32$
. We shall write
$\emptyset < \emptyset3 < \emptyset32$
. We shall write
 $|u|$
for the number of ancestors of the particle u, so for example
$|u|$
for the number of ancestors of the particle u, so for example
 $|\emptyset32| = 2$
.
$|\emptyset32| = 2$
.
We denote the set of all particles in the system at time t by
 $N_t$
, and for every particle
$N_t$
, and for every particle
 $u \in N_t$
, we let
$u \in N_t$
, we let
 $X^u_t$
denote its spatial position at time t and
$X^u_t$
denote its spatial position at time t and
 ${(X^u_s)_{s \in [0,t]}}$
its historical path up to time t with
${(X^u_s)_{s \in [0,t]}}$
its historical path up to time t with
 $L_t^u$
the local time at 0 of
$L_t^u$
the local time at 0 of
 ${(X^u_s)_{s \in [0,t]}}$
. We also define
${(X^u_s)_{s \in [0,t]}}$
. We also define
 ${R_t \,{:\!=}\, \sup \{X^u_t \colon u \in N_t\}}$
${R_t \,{:\!=}\, \sup \{X^u_t \colon u \in N_t\}}$
to be the position of the rightmost particle at time t. We let
 $A_u$
denote the number of offspring produced by particle u when it dies.
$A_u$
denote the number of offspring produced by particle u when it dies.
We let
 $m_0 = \smash{\sum_{n \geq 1}}n q_n$
be the mean of the offspring distribution due to catalytic branching and
$m_0 = \smash{\sum_{n \geq 1}}n q_n$
be the mean of the offspring distribution due to catalytic branching and
 $m = \smash{\sum_{n \geq 1}}n p_n$
the mean of the offspring distribution due to homogeneous branching. For convenience, we also respectively define the effective homogeneous and catalytic branching rates as
$m = \smash{\sum_{n \geq 1}}n p_n$
the mean of the offspring distribution due to homogeneous branching. For convenience, we also respectively define the effective homogeneous and catalytic branching rates as
 $$\hat{\beta} \,{:\!=}\, \beta(m-1)
\quad\text{and}\quad
\hat{\beta}_0 \,{:\!=}\, \beta_0(m_0 - 1).$$
$$\hat{\beta} \,{:\!=}\, \beta(m-1)
\quad\text{and}\quad
\hat{\beta}_0 \,{:\!=}\, \beta_0(m_0 - 1).$$
Finally, we let
 ${(\mathcal{F}_t)_{t \ge 0}}$
be the natural filtration of the branching process, and for the process initiated from position
${(\mathcal{F}_t)_{t \ge 0}}$
be the natural filtration of the branching process, and for the process initiated from position
 $x \in \mathbb{R}$
we let
$x \in \mathbb{R}$
we let
 $\mathbb{P}^x$
be the associated probability measure with the corresponding expectation
$\mathbb{P}^x$
be the associated probability measure with the corresponding expectation
 $\mathbb{E}^x$
. Most of the time it will be assumed that
$\mathbb{E}^x$
. Most of the time it will be assumed that
 $x=0$
, and we shall then write
$x=0$
, and we shall then write
 $\mathbb{P}$
and
$\mathbb{P}$
and
 $\mathbb{E}$
instead of
$\mathbb{E}$
instead of
 $\mathbb{P}^0$
and
$\mathbb{P}^0$
and
 $\mathbb{E}^0$
.
$\mathbb{E}^0$
.
1.3. Motivation
In this subsection we present a few simple calculations which should motivate our main results in the next subsection.
For any
 $x \in \mathbb{R}$
and
$x \in \mathbb{R}$
and
 $t \ge 0$
, let us define
$t \ge 0$
, let us define
 \begin{equation}
N_t^x \,{:\!=}\, \{ u \in N_t \colon X^u_t >x\}
\end{equation}
\begin{equation}
N_t^x \,{:\!=}\, \{ u \in N_t \colon X^u_t >x\}
\end{equation}
to be the set of particles in the system at time t whose spatial position is to the right of x. A simple application of the widely used ‘many-to-one’ formula (see Subsection 2.2) gives an exact expression for
 $\mathbb{E}|N_t^x|$
(to be proved in Subsection 2.2).
$\mathbb{E}|N_t^x|$
(to be proved in Subsection 2.2).
Proposition 1. For any
 $x \geq 0$
and
$x \geq 0$
and
 $t \ge 0$
,
$t \ge 0$
,
 $$
\begin{equation}
\mathbb{E} |N_t^x| = \Phi \bigg( \hat{\beta}_0 \sqrt{t} - {x}/{\sqrt{t}}\bigg) \exp \bigg\{ {1}/{2} \hat{\beta}_0^2t -\hat{\beta}_0x + \hat{\beta}t \bigg\} ,
\end{equation}
$$
$$
\begin{equation}
\mathbb{E} |N_t^x| = \Phi \bigg( \hat{\beta}_0 \sqrt{t} - {x}/{\sqrt{t}}\bigg) \exp \bigg\{ {1}/{2} \hat{\beta}_0^2t -\hat{\beta}_0x + \hat{\beta}t \bigg\} ,
\end{equation}
$$
where
 $${\Phi(x) = \mathbb{P} (\mathcal{N}(0, 1) \leq x) = \int_{- \infty}^x (2 \pi)^{-1/2} \,\mathrm{e}^{- y^2/2} \,\mathrm{d}y}$$
$${\Phi(x) = \mathbb{P} (\mathcal{N}(0, 1) \leq x) = \int_{- \infty}^x (2 \pi)^{-1/2} \,\mathrm{e}^{- y^2/2} \,\mathrm{d}y}$$
is the cumulative distribution function of a standard normal random variable.
In particular, for any
 $\lambda \geq 0$
we obtain
$\lambda \geq 0$
we obtain
 $$\begin{equation}
\mathbb{E}|N_t^{\lambda t}| = \Phi ( (\hat{\beta}_0 - \lambda) \sqrt{t}) \exp \bigg\{ \bigg({1}/{2}\hat{\beta}_0^2 - \hat{\beta}_0\lambda + \hat{\beta}\bigg)t\bigg\}.
\end{equation}$$
$$\begin{equation}
\mathbb{E}|N_t^{\lambda t}| = \Phi ( (\hat{\beta}_0 - \lambda) \sqrt{t}) \exp \bigg\{ \bigg({1}/{2}\hat{\beta}_0^2 - \hat{\beta}_0\lambda + \hat{\beta}\bigg)t\bigg\}.
\end{equation}$$
Using the fact that
 $\Phi(x) \sim (2 \pi)^{-1/2}|x|^{-1} \,\mathrm{e}^{-x^2/2}$
as
$\Phi(x) \sim (2 \pi)^{-1/2}|x|^{-1} \,\mathrm{e}^{-x^2/2}$
as
 $x \to - \infty$
and
$x \to - \infty$
and
 $\Phi(x) \to 1$
as
$\Phi(x) \to 1$
as
 $x \to \infty$
, we can then see that
$x \to \infty$
, we can then see that
 $$\begin{equation}{1}/{t} \log \mathbb{E}|N_t^{\lambda t}| \to \Delta_\lambda \text{ as } t \to \infty ,
\end{equation}$$
$$\begin{equation}{1}/{t} \log \mathbb{E}|N_t^{\lambda t}| \to \Delta_\lambda \text{ as } t \to \infty ,
\end{equation}$$
where
 $$\begin{equation}
\Delta_{\lambda} =
\begin{cases}{1}/{2} \hat{\beta}_0^2 - \hat{\beta}_0 \lambda + \hat{\beta}& \text{if } \lambda \leq \hat{\beta}_0, \\
- {1}/{2} \lambda^2 + \hat{\beta}& \text{if } \lambda \geq \hat{\beta}_0 .
\end{cases}
\end{equation}$$
$$\begin{equation}
\Delta_{\lambda} =
\begin{cases}{1}/{2} \hat{\beta}_0^2 - \hat{\beta}_0 \lambda + \hat{\beta}& \text{if } \lambda \leq \hat{\beta}_0, \\
- {1}/{2} \lambda^2 + \hat{\beta}& \text{if } \lambda \geq \hat{\beta}_0 .
\end{cases}
\end{equation}$$
We can then observe that
 $\Delta_\lambda$
takes positive or negative values according to whether
$\Delta_\lambda$
takes positive or negative values according to whether
 $\lambda < \lambda_{\mathrm{crit}}$
or
$\lambda < \lambda_{\mathrm{crit}}$
or
 $\lambda > \lambda_{\mathrm{crit}}$
, where
$\lambda > \lambda_{\mathrm{crit}}$
, where
 $$\begin{equation}
\lambda_{\mathrm{crit}} =
\begin{cases}
{\hat{\beta}}/{\hat{\beta}_0} + {1}/{2} \hat{\beta}_0& \text{if } \hat{\beta} \leq {1}/{2}\hat{\beta}_0^2, \\
\sqrt{2 \hat{\beta}}& \text{if } \hat{\beta} \geq {1}/{2}\hat{\beta}_0^2 .
\end{cases}
\end{equation}$$
$$\begin{equation}
\lambda_{\mathrm{crit}} =
\begin{cases}
{\hat{\beta}}/{\hat{\beta}_0} + {1}/{2} \hat{\beta}_0& \text{if } \hat{\beta} \leq {1}/{2}\hat{\beta}_0^2, \\
\sqrt{2 \hat{\beta}}& \text{if } \hat{\beta} \geq {1}/{2}\hat{\beta}_0^2 .
\end{cases}
\end{equation}$$
Since the expected number of particles to the right of
 $(\lambda_{\mathrm{crit}} + \varepsilon)t$
decays exponentially with t and the expected number of particles to the right of
$(\lambda_{\mathrm{crit}} + \varepsilon)t$
decays exponentially with t and the expected number of particles to the right of
 ${(\lambda_{\mathrm{crit}} - \varepsilon)t}$
grows exponentially with t, we may interpret
${(\lambda_{\mathrm{crit}} - \varepsilon)t}$
grows exponentially with t, we may interpret
 $\lambda_{\mathrm{crit}}$
for now as the speed of the rightmost particle ‘in expectation’.
$\lambda_{\mathrm{crit}}$
for now as the speed of the rightmost particle ‘in expectation’.
Also, using symmetry or a direct calculation, we may find the expected total population at any time
 $t \ge 0$
:
$t \ge 0$
:
 $$\begin{equation}
\mathbb{E} |N_t| = 2 \mathbb{E} |N_t^0| = 2 \Phi ( \hat{\beta}_0 \sqrt{t}) \exp \bigg\{ \bigg( {1}/{2} \hat{\beta}_0^2 + \hat{\beta}\bigg) t \bigg\} .
\end{equation}$$
$$\begin{equation}
\mathbb{E} |N_t| = 2 \mathbb{E} |N_t^0| = 2 \Phi ( \hat{\beta}_0 \sqrt{t}) \exp \bigg\{ \bigg( {1}/{2} \hat{\beta}_0^2 + \hat{\beta}\bigg) t \bigg\} .
\end{equation}$$
In particular,
 $$\begin{equation}
\mathbb{E} |N_t| \sim \alpha \exp \bigg\{ \bigg( {1}/{2} \hat{\beta}_0^2 + \hat{\beta}\bigg) t \bigg\} \text{ as } t \to \infty ,
\end{equation}$$
$$\begin{equation}
\mathbb{E} |N_t| \sim \alpha \exp \bigg\{ \bigg( {1}/{2} \hat{\beta}_0^2 + \hat{\beta}\bigg) t \bigg\} \text{ as } t \to \infty ,
\end{equation}$$
where
 $\alpha = 1$
if
$\alpha = 1$
if
 $\hat{\beta}_0 = 0$
and
$\hat{\beta}_0 = 0$
and
 $\alpha = 2$
if
$\alpha = 2$
if
 $\hat{\beta}_0 > 0$
.
$\hat{\beta}_0 > 0$
.
1.4. Main results
Our aim is to replace convergences in expectation from the previous subsection with the almost sure convergences.
For all the results in this subsection we shall have to impose an additional condition on the offspring distribution commonly known as the
 $X \log X$
condition (for some discussion see e.g. [Reference Kyprianou14], [Reference Liu, Ren and Song16], or [Reference Lyons17]):
$X \log X$
condition (for some discussion see e.g. [Reference Kyprianou14], [Reference Liu, Ren and Song16], or [Reference Lyons17]):
 $$\begin{equation}
\sum_{n \geq 1} p_n n \log n < \infty \quad\text{and}\quad \sum_{n \geq 1} q_n n \log n < \infty .
\end{equation}$$
$$\begin{equation}
\sum_{n \geq 1} p_n n \log n < \infty \quad\text{and}\quad \sum_{n \geq 1} q_n n \log n < \infty .
\end{equation}$$
This condition is needed to ensure that certain martingales have non-zero limits, as we shall see in Subsection 2.3.
Our first result, which should be compared with (9), is the almost sure approximation of the population size.
Theorem 1. Suppose that condition (10) on the offspring distribution is satisfied. Then
 $$\mathop {\lim }\limits_{t \to \infty } 1/t\log |{N_t}| = 1/2\hat \beta _0^2 + \hat \beta \quad {\mathbb{P - a}}{\rm{.s}}{\rm{.}}$$
$$\mathop {\lim }\limits_{t \to \infty } 1/t\log |{N_t}| = 1/2\hat \beta _0^2 + \hat \beta \quad {\mathbb{P - a}}{\rm{.s}}{\rm{.}}$$
Next, unarguably the most important result of this paper, is the almost sure approximation of
 $|N_t^{\lambda t}|$
, and it should be compared with (5).
$|N_t^{\lambda t}|$
, and it should be compared with (5).
Theorem 2. Suppose that condition (10) is satisfied. Take any
 $\lambda > 0$
and let
$\lambda > 0$
and let
 $\Delta_\lambda$
be as in (6) and
$\Delta_\lambda$
be as in (6) and
 $\lambda_{\mathrm{crit}}$
as in (7).
$\lambda_{\mathrm{crit}}$
as in (7).
If
 $\lambda < \lambda_{\mathrm{crit}}$
, then
$\lambda < \lambda_{\mathrm{crit}}$
, then
 $$\begin{equation}
\lim_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| = \Delta_\lambda\ ({>}\, 0) \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
$$\begin{equation}
\lim_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| = \Delta_\lambda\ ({>}\, 0) \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
If
 $\lambda > \lambda_{\mathrm{crit}}$
, then
$\lambda > \lambda_{\mathrm{crit}}$
, then
 $$\begin{equation}
\lim_{t \to \infty} |N_t^{\lambda t}| = 0 \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
$$\begin{equation}
\lim_{t \to \infty} |N_t^{\lambda t}| = 0 \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
and furthermore
 $$\begin{equation}
\lim_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0) = \Delta_{\lambda}\ ({<}\, 0) .
\end{equation}$$
$$\begin{equation}
\lim_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0) = \Delta_{\lambda}\ ({<}\, 0) .
\end{equation}$$
As a direct corollary of Theorem 2 we establish the almost sure speed of the rightmost particle.
Corollary 1. Suppose that condition (10) is satisfied. Then
 $$\mathop {\lim }\limits_{t \to \infty } {R_t}/t = {\lambda _{{\rm{crit}}}}\quad {\mathbb{P - a}}{\rm{.s}}{\rm{.,}}$$
$$\mathop {\lim }\limits_{t \to \infty } {R_t}/t = {\lambda _{{\rm{crit}}}}\quad {\mathbb{P - a}}{\rm{.s}}{\rm{.,}}$$
where
 $\lambda_{\mathrm{crit}}$
is given in (7).
$\lambda_{\mathrm{crit}}$
is given in (7).
Note that letting either
 $\beta_0 \to 0$
or
$\beta_0 \to 0$
or
 $\beta \to 0$
in Theorem 1, Theorem 2, and Corollary 1, one may recover previously known results for purely homogeneous or purely catalytic branching processes.
$\beta \to 0$
in Theorem 1, Theorem 2, and Corollary 1, one may recover previously known results for purely homogeneous or purely catalytic branching processes.
1.5. Outline of the paper
The rest of this article is organized as follows. In Subsection 2.1 we follow the standard procedure of extending the probability space by constructing the spine process over our branching system. Then in Subsection 2.2 we recall the Many-to-One formula for branching processes and apply it to prove Proposition 1 as well as the upper bounds for Theorem 1 and Theorem 2. In Subsection 2.3 we construct certain change-of-measure martingales and use them to prove the lower bound for Theorem 1 as well as ‘preliminary’ lower bounds for Theorem 2. In Subsections 3.1 and 3.2 we give the heuristic argument and the formal proof for the lower bounds in Theorem 2. Finally, we conclude the paper with the proof of Corollary 1
2. Spine results and applications
2.1 Spine construction
In this section we extend our probability space by introducing the spine process. A more detailed description of this procedure may be found for example in [Reference Hardy and Harris9].
The spine of the branching process, which we shall denote by
 $\xi$
, is the infinite line of descent of particles chosen uniformly at random from all possible lines of descent. It is constructed in the following way. The initial particle
$\xi$
, is the infinite line of descent of particles chosen uniformly at random from all possible lines of descent. It is constructed in the following way. The initial particle
 $\emptyset$
of the branching process begins the spine. When the initial particle dies and is replaced with a random number of new particles, one of them is chosen uniformly at random to continue the spine. This procedure is then repeated recursively: whenever the particle in the spine dies, one of its children is chosen uniformly at random to continue the spine. We may then write the spine as
$\emptyset$
of the branching process begins the spine. When the initial particle dies and is replaced with a random number of new particles, one of them is chosen uniformly at random to continue the spine. This procedure is then repeated recursively: whenever the particle in the spine dies, one of its children is chosen uniformly at random to continue the spine. We may then write the spine as
 $\xi = \{ \xi^{(0)}, \xi^{(1)}, \xi^{(2)}, \ldots\}$
, where
$\xi = \{ \xi^{(0)}, \xi^{(1)}, \xi^{(2)}, \ldots\}$
, where
 $\xi^{(n)}$
is the label of the spine particle in the nth generation and
$\xi^{(n)}$
is the label of the spine particle in the nth generation and
 $\xi^{(0)} = \emptyset$
.
$\xi^{(0)} = \emptyset$
.
We let
 $\tilde{\mathbb{P}}$
denote the probability measure under which the branching process is defined together with the spine. Hence
$\tilde{\mathbb{P}}$
denote the probability measure under which the branching process is defined together with the spine. Hence
 $\mathbb{P} = \tilde{\mathbb{P}}|_{\mathcal{F}_\infty}$
. We let
$\mathbb{P} = \tilde{\mathbb{P}}|_{\mathcal{F}_\infty}$
. We let
 $\tilde{\mathbb{E}}$
be the expectation corresponding to
$\tilde{\mathbb{E}}$
be the expectation corresponding to
 $\tilde{\mathbb{P}}$
.
$\tilde{\mathbb{P}}$
.
Below we introduce some new notation in relation with the spine process.
For any
 $t \ge 0$
we write
$t \ge 0$
we write
 ${{\mathop{\rm node}\nolimits} _t}(\xi )$
for the unique particle
${{\mathop{\rm node}\nolimits} _t}(\xi )$
for the unique particle
 $u \in N_t \cap \xi$
. That is,
$u \in N_t \cap \xi$
. That is,
 ${{\mathop{\rm node}\nolimits} _t}(\xi )$
is the label of the spine particle at time t.
${{\mathop{\rm node}\nolimits} _t}(\xi )$
is the label of the spine particle at time t.
For any
 $t \ge 0$
we write
$t \ge 0$
we write
 $\xi_t$
for
$\xi_t$
for
 $X^u_t$
, where u is the unique particle in
$X^u_t$
, where u is the unique particle in
 $N_t \cap \xi$
. Then
$N_t \cap \xi$
. Then
 $\xi_t$
is the spatial position of the spine particle at time t. It is not hard to check that the process
$\xi_t$
is the spatial position of the spine particle at time t. It is not hard to check that the process
 ${(\xi_t)_{t \ge 0}}$
is a Brownian motion under
${(\xi_t)_{t \ge 0}}$
is a Brownian motion under
 $\tilde{\mathbb{P}}$
. We let
$\tilde{\mathbb{P}}$
. We let
 ${(\tilde{L}_t)_{t \ge 0}}$
denote its local time at the origin.
${(\tilde{L}_t)_{t \ge 0}}$
denote its local time at the origin.
For any
 ${t \ge 0}$
we write
${t \ge 0}$
we write
 $n_t$
for the unique n such that
$n_t$
for the unique n such that
 $\xi^{(n)} \in N_t$
. Then
$\xi^{(n)} \in N_t$
. Then
 ${(n_t)_{t \ge 0}}$
is the counting process of the number of branching events that have occurred along the path of the spine by time t. We denote the sequence of times of these branching events by
${(n_t)_{t \ge 0}}$
is the counting process of the number of branching events that have occurred along the path of the spine by time t. We denote the sequence of times of these branching events by
 $S_n$
and the number of particles produced at each such branching event by
$S_n$
and the number of particles produced at each such branching event by
 $A_n$
,
$A_n$
,
 $n \geq 1$
.
$n \geq 1$
.
Moreover, we would like to distinguish branching events along the spine that occurred due to catalytic branching from those that occurred due to homogeneous branching. In order to do so, we denote the branching times along the spine that took place when the spine was at the origin by
 $S_n^0$
, and the number of particles produced at these times by
$S_n^0$
, and the number of particles produced at these times by
 $A_n^0$
,
$A_n^0$
,
 $n \geq 1$
. Similarly, we denote the branching times along the spine when it was not at the origin by
$n \geq 1$
. Similarly, we denote the branching times along the spine when it was not at the origin by
 $S_n'$
, and the number of particles produced at these times by
$S_n'$
, and the number of particles produced at these times by
 $A_n'$
,
$A_n'$
,
 $n \geq 1$
. We also denote the counting processes for
$n \geq 1$
. We also denote the counting processes for
 ${(S_n^0)_{n \geq 1}}$
and
${(S_n^0)_{n \geq 1}}$
and
 ${(S_n')_{n \geq 0}}$
by
${(S_n')_{n \geq 0}}$
by
 ${(n_t^0)_{t \ge 0}}$
and
${(n_t^0)_{t \ge 0}}$
and
 ${(n_t')_{t \ge 0}}$
, respectively.
${(n_t')_{t \ge 0}}$
, respectively.
Observe that conditional on the path of the spine
 ${(\xi_t)_{t \ge 0}}$
,
${(\xi_t)_{t \ge 0}}$
,
 ${(n_t^0)_{t \ge 0}}$
and
${(n_t^0)_{t \ge 0}}$
and
 ${(n_t')_{t \ge 0}}$
are independent (inhomogeneous in the first case) Poisson processes (or Cox processes) with jump rates
${(n_t')_{t \ge 0}}$
are independent (inhomogeneous in the first case) Poisson processes (or Cox processes) with jump rates
 $\beta_0 \delta_0({\cdot})$
and
$\beta_0 \delta_0({\cdot})$
and
 $\beta$
, respectively, so that
$\beta$
, respectively, so that
 $$\tilde{\mathbb{P}}( n_t^0 = k \!{\mid}\! (\xi_s)_{0 \leq s \leq t}) = {(\beta_0 \tilde{L}_t)^k}/{k!} \,\mathrm{e}^{- \beta_0 \tilde{L}_t}$$
$$\tilde{\mathbb{P}}( n_t^0 = k \!{\mid}\! (\xi_s)_{0 \leq s \leq t}) = {(\beta_0 \tilde{L}_t)^k}/{k!} \,\mathrm{e}^{- \beta_0 \tilde{L}_t}$$
and
 $$\tilde{\mathbb{P}}( n_t' = k \!{\mid}\! (\xi_s)_{0 \leq s \leq t}) = {(\beta t)^k}/{k!} \,\mathrm{e}^{- \beta t}.$$
$$\tilde{\mathbb{P}}( n_t' = k \!{\mid}\! (\xi_s)_{0 \leq s \leq t}) = {(\beta t)^k}/{k!} \,\mathrm{e}^{- \beta t}.$$
Finally, it is convenient to define several filtrations of the now extended probability space in order to take various conditional expectations.
Definition 1. (Filtrations.)
 $\mathcal{F}_t$
was defined in Subsection 1.2. It is the filtration which contains all the information about all the particles’ motion and their genealogy. It does not, however, have any information about the spine.
$\mathcal{F}_t$
was defined in Subsection 1.2. It is the filtration which contains all the information about all the particles’ motion and their genealogy. It does not, however, have any information about the spine.-
$\tilde{\mathcal{F}}_t \,{:\!=}\, \sigma (\mathcal{F}_t,\ {{\mathop{\rm node}\nolimits} _t}(\xi ))$
. Thus
$\tilde{\mathcal{F}}$
has all the information about the branching process and all the information about the spine. This will be the largest filtration.
-
$\mathcal{G}_t \,{:\!=}\, \sigma ( \xi_s \colon 0 \leq s \leq t )$
. This filtration only contains information about the path of the spine but it does not know which particles make up the spine along its path at different times.
-
${\mathcal{G}_t}{\mkern 1mu} : = {\mkern 1mu} \sigma (\mathcal{G}_t,\;{{\mathop{\rm node}\nolimits} _s}(\xi ):0 \le s \le t,\;A_n^0:n \le n_t^0,\;{A_{m}'}:m \le {n_t}')$
. This filtration has information about the path of the spine, its genealogy, and how many particles are born along the path of the spine. However, it has no information about anything happening off the spine.





We note that
 $\mathcal{F}_t \subset \tilde{\mathcal{F}}_t$
and
$\mathcal{F}_t \subset \tilde{\mathcal{F}}_t$
and
 $\mathcal{G}_t \subset \tilde{\mathcal{G}}_t \subset \tilde{\mathcal{F}}_t$
.
$\mathcal{G}_t \subset \tilde{\mathcal{G}}_t \subset \tilde{\mathcal{F}}_t$
.
2.2. Many-to-One Lemma and applications
The proof of the following result with a detailed discussion can be found for example in [Reference Hardy and Harris9] or [Reference Harris and Roberts11].
Lemma 1. (Many-to-One Lemma.) Let Y be a non-negative
 $\tilde{\mathcal{F}}_t$
-measurable random variable. It can be decomposed as
$\tilde{\mathcal{F}}_t$
-measurable random variable. It can be decomposed as
 $$Y = \sum_{u \in N_t} Y(u) \mathbf{1}_{\{ {{\mathop{\rm node}\nolimits} _t}(\xi ) = u \}} ,$$
$$Y = \sum_{u \in N_t} Y(u) \mathbf{1}_{\{ {{\mathop{\rm node}\nolimits} _t}(\xi ) = u \}} ,$$
where for all
 $u \in N_t$
, Y(u) is
$u \in N_t$
, Y(u) is
 $\mathcal{F}_t$
-measurable and then
$\mathcal{F}_t$
-measurable and then
 $$\mathbb{E}^x \bigg( \sum_{u \in N_t} Y(u) \bigg) = \tilde{\mathbb{E}}^x ( Y \,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_t + \hat{\beta} t}) {.}$$
$$\mathbb{E}^x \bigg( \sum_{u \in N_t} Y(u) \bigg) = \tilde{\mathbb{E}}^x ( Y \,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_t + \hat{\beta} t}) {.}$$
In particular, if f is a non-negative functional such that
 $f((\xi_s)_{s \in [0,t]})$
is a
$f((\xi_s)_{s \in [0,t]})$
is a
 $\mathcal{G}_t$
-measurable random variable, then
$\mathcal{G}_t$
-measurable random variable, then
 $$\begin{equation}
\mathbb{E}^x \bigg[ \sum_{u \in N_t} f ( (X^u_s)_{s \in [0,t]}) \bigg] = \tilde{\mathbb{E}}^x [ f((\xi_s)_{s \in [0,t]} ) \,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_t + \hat{\beta} t}] .
\end{equation}$$
$$\begin{equation}
\mathbb{E}^x \bigg[ \sum_{u \in N_t} f ( (X^u_s)_{s \in [0,t]}) \bigg] = \tilde{\mathbb{E}}^x [ f((\xi_s)_{s \in [0,t]} ) \,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_t + \hat{\beta} t}] .
\end{equation}$$
Let us now apply (14) to prove equations (3) and (8) given as the motivation in the first section.
Proof of Proposition 1 and identity (8). Take
 $x \geq 0$
and
$x \geq 0$
and
 $t \ge 0$
. Then
$t \ge 0$
. Then
 $$\begin{align}
\mathbb{E} |N_t^x| = \mathbb{E} \sum_{u \in N_t} \mathbf{1}_{\{X^u_t > x\}}
= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_t > x\}} \,\mathrm{e}^{\hat{\beta}_0 \tilde{L_t} + \hat{\beta} t}] .
\end{align}$$
$$\begin{align}
\mathbb{E} |N_t^x| = \mathbb{E} \sum_{u \in N_t} \mathbf{1}_{\{X^u_t > x\}}
= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_t > x\}} \,\mathrm{e}^{\hat{\beta}_0 \tilde{L_t} + \hat{\beta} t}] .
\end{align}$$
We now make use of the joint density of
 $\xi_t$
and
$\xi_t$
and
 $\tilde{L}_t$
(which for example can be found in [Reference Karatzas and Shreve12]):
$\tilde{L}_t$
(which for example can be found in [Reference Karatzas and Shreve12]):
 $$\tilde{\mathbb{P}} ( \xi_t \in \mathrm{d}y,\ \tilde{L}_t \in \mathrm{d} l) =
{|y| + l}/{\sqrt{2 \pi t^3}} \exp \bigg\{ - {(|y| + l)^2}/{2t}\bigg\} \,\mathrm{d}y \,\mathrm{d}l{,} \quad y \in \mathbb{R},\ l \geq 0{,}$$
$$\tilde{\mathbb{P}} ( \xi_t \in \mathrm{d}y,\ \tilde{L}_t \in \mathrm{d} l) =
{|y| + l}/{\sqrt{2 \pi t^3}} \exp \bigg\{ - {(|y| + l)^2}/{2t}\bigg\} \,\mathrm{d}y \,\mathrm{d}l{,} \quad y \in \mathbb{R},\ l \geq 0{,}$$
to complete the proof of (3). Then
 $$\begin{align*}
\tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_t > x\}} \,\mathrm{e}^{\hat{\beta}_0 \tilde{L_t} + \hat{\beta} t}] &=
\mathrm{e}^{\hat{\beta} t} \int_0^\infty \int_x^{\infty} \,\mathrm{e}^{\hat{\beta}_0 l} {y + l}/{\sqrt{2 \pi t^3}}
\exp \bigg\{ - {(y+l)^2}/{2t}\bigg\} \,\mathrm{d}y \,\mathrm{d}l\\
&= \mathrm{e}^{\hat{\beta} t} \int_0^\infty \,\mathrm{e}^{\hat{\beta}_0 l} {1}/{\sqrt{2 \pi t}}
\exp \bigg\{ - {(x+l)^2}/{2t}\bigg\} \,\mathrm{d}l\\
&= \mathrm{e}^{\hat{\beta} t} \int_0^\infty {1}/{\sqrt{2 \pi t}} \exp \bigg\{ - {1}/{2t} ( l - (\hat{\beta}_0t - x))^2
+ {\hat{\beta}_0^2}/{2}t - \hat{\beta}_0 x \bigg\} \,\mathrm{d}l\\
&= \mathrm{e}^{\hat{\beta}t + {\hat{\beta}_0^2}/{2}t - \hat{\beta}_0 x} \int_{- (\hat{\beta}_0 \sqrt{t} - \frac{x}{\sqrt{t}})}^\infty
{1}/{\sqrt{2 \pi}} \,\mathrm{e}^{- \frac{z^2}{2}} \,\mathrm{d}z\\
&= \Phi \bigg( \hat{\beta}_0 \sqrt{t} - {x}/{\sqrt{t}}\bigg) \exp \bigg\{ {1}/{2} \hat{\beta}_0^2t -\hat{\beta}_0x + \hat{\beta}t \bigg\} .
\end{align*}$$
$$\begin{align*}
\tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_t > x\}} \,\mathrm{e}^{\hat{\beta}_0 \tilde{L_t} + \hat{\beta} t}] &=
\mathrm{e}^{\hat{\beta} t} \int_0^\infty \int_x^{\infty} \,\mathrm{e}^{\hat{\beta}_0 l} {y + l}/{\sqrt{2 \pi t^3}}
\exp \bigg\{ - {(y+l)^2}/{2t}\bigg\} \,\mathrm{d}y \,\mathrm{d}l\\
&= \mathrm{e}^{\hat{\beta} t} \int_0^\infty \,\mathrm{e}^{\hat{\beta}_0 l} {1}/{\sqrt{2 \pi t}}
\exp \bigg\{ - {(x+l)^2}/{2t}\bigg\} \,\mathrm{d}l\\
&= \mathrm{e}^{\hat{\beta} t} \int_0^\infty {1}/{\sqrt{2 \pi t}} \exp \bigg\{ - {1}/{2t} ( l - (\hat{\beta}_0t - x))^2
+ {\hat{\beta}_0^2}/{2}t - \hat{\beta}_0 x \bigg\} \,\mathrm{d}l\\
&= \mathrm{e}^{\hat{\beta}t + {\hat{\beta}_0^2}/{2}t - \hat{\beta}_0 x} \int_{- (\hat{\beta}_0 \sqrt{t} - \frac{x}{\sqrt{t}})}^\infty
{1}/{\sqrt{2 \pi}} \,\mathrm{e}^{- \frac{z^2}{2}} \,\mathrm{d}z\\
&= \Phi \bigg( \hat{\beta}_0 \sqrt{t} - {x}/{\sqrt{t}}\bigg) \exp \bigg\{ {1}/{2} \hat{\beta}_0^2t -\hat{\beta}_0x + \hat{\beta}t \bigg\} .
\end{align*}$$
For the expected total population we could have followed a similar calculation:
 $$\begin{equation}
\mathbb{E}|N_t| = \tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L_t} + \hat{\beta} t} ] = \cdots =
2 \Phi ( \hat{\beta}_0 \sqrt{t}) \exp \bigg\{ \bigg( {1}/{2} \hat{\beta}_0^2 + \hat{\beta}\bigg) t \bigg\}.
\end{equation}$$
$$\begin{equation}
\mathbb{E}|N_t| = \tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L_t} + \hat{\beta} t} ] = \cdots =
2 \Phi ( \hat{\beta}_0 \sqrt{t}) \exp \bigg\{ \bigg( {1}/{2} \hat{\beta}_0^2 + \hat{\beta}\bigg) t \bigg\}.
\end{equation}$$
Let us now prove the upper bound for Theorem 1.
Proposition 2. (Upper bound for Theorem 1.)
 $$\begin{equation}
\limsup_{t \to \infty} {1}/{t} \log |N_t| \leq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
$$\begin{equation}
\limsup_{t \to \infty} {1}/{t} \log |N_t| \leq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
Proof. Fix
 $\varepsilon > 0$
. Then, by the Markov inequality and (8),
$\varepsilon > 0$
. Then, by the Markov inequality and (8),
 $$\begin{align*}
\mathbb{P} \bigg( \dfrac{1}{n} \log |N_n| > \dfrac{1}{2}\hat{\beta}_0^2 +
\hat{\beta} + \varepsilon \bigg) = \mathbb{P} ( |N_n| >
\mathrm{e}^{(\frac{1}{2} \hat{\beta}_0^2 + \hat{\beta} + \varepsilon)n} )
\leq \mathrm{e}^{-(\frac{1}{2} \hat{\beta}_0^2 + \hat{\beta} + \varepsilon)n}
\mathbb{E}|N_n| < 2 \,\mathrm{e}^{- \varepsilon n}.
\end{align*}$$
$$\begin{align*}
\mathbb{P} \bigg( \dfrac{1}{n} \log |N_n| > \dfrac{1}{2}\hat{\beta}_0^2 +
\hat{\beta} + \varepsilon \bigg) = \mathbb{P} ( |N_n| >
\mathrm{e}^{(\frac{1}{2} \hat{\beta}_0^2 + \hat{\beta} + \varepsilon)n} )
\leq \mathrm{e}^{-(\frac{1}{2} \hat{\beta}_0^2 + \hat{\beta} + \varepsilon)n}
\mathbb{E}|N_n| < 2 \,\mathrm{e}^{- \varepsilon n}.
\end{align*}$$
It follows from the Borel–Cantelli lemma that
 $$\mathbb{P} \bigg( \bigg\{ \dfrac{1}{n} \log |N_n| > \dfrac{1}{2}\hat{\beta}_0^2 + \hat{\beta} + \varepsilon \bigg\} \text{ i.o.}\bigg) = 0$$
$$\mathbb{P} \bigg( \bigg\{ \dfrac{1}{n} \log |N_n| > \dfrac{1}{2}\hat{\beta}_0^2 + \hat{\beta} + \varepsilon \bigg\} \text{ i.o.}\bigg) = 0$$
and thus
 $$\limsup_{n \to \infty} \dfrac{1}{n} \log |N_n| \leq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} +
\varepsilon \quad \mathbb{P} \text{-a.s.}$$
$$\limsup_{n \to \infty} \dfrac{1}{n} \log |N_n| \leq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} +
\varepsilon \quad \mathbb{P} \text{-a.s.}$$
By letting
 $\varepsilon \to 0$
we establish (17) with the limit taken over integer times. To get convergence over any real-valued sequence, we note that
$\varepsilon \to 0$
we establish (17) with the limit taken over integer times. To get convergence over any real-valued sequence, we note that
 ${(|N_t|)_{t \ge 0}}$
is a non-decreasing process, and so for any
${(|N_t|)_{t \ge 0}}$
is a non-decreasing process, and so for any
 $t > 0$
$t > 0$
 $$\dfrac{1}{t}\log |N_t| \leq \dfrac{\lceil t \rceil}{t} \dfrac{\log |N_{\lceil t \rceil}|}{\lceil t \rceil}{,}$$
$$\dfrac{1}{t}\log |N_t| \leq \dfrac{\lceil t \rceil}{t} \dfrac{\log |N_{\lceil t \rceil}|}{\lceil t \rceil}{,}$$
and hence
 $$\begin{align*}
\limsup_{t \to \infty} {1}/{t} \log |N_t| \leq \limsup_{t \to \infty}
\dfrac{\log |N_{\lceil t \rceil}|}{\lceil t \rceil}
\leq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} \quad \mathbb{P} \text{-a.s.}\\
\end{align*}$$
$$\begin{align*}
\limsup_{t \to \infty} {1}/{t} \log |N_t| \leq \limsup_{t \to \infty}
\dfrac{\log |N_{\lceil t \rceil}|}{\lceil t \rceil}
\leq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} \quad \mathbb{P} \text{-a.s.}\\
\end{align*}$$
□
For upper bounds of Theorem 2 we need to adjust the previous argument because unlike
 ${(|N_t|)_{t \ge 0}}$
the process
${(|N_t|)_{t \ge 0}}$
the process
 ${(|N_t^{\lambda t}|)_{t \ge 0}}$
is not monotone. We first establish the following result.
${(|N_t^{\lambda t}|)_{t \ge 0}}$
is not monotone. We first establish the following result.
Proposition 3. For
 $\lambda > 0$
and
$\lambda > 0$
and
 $n \in \mathbb{N}\cup\{0\}$
, we define the following set of particles:
$n \in \mathbb{N}\cup\{0\}$
, we define the following set of particles:
 $$\hat{N}^{\lambda n}_n \,{:\!=}\, \Big\{ u \in N_{n+1} \colon \sup_{s \in [n, n+1]}X^u_s \geq \lambda n\Big\}.$$
$$\hat{N}^{\lambda n}_n \,{:\!=}\, \Big\{ u \in N_{n+1} \colon \sup_{s \in [n, n+1]}X^u_s \geq \lambda n\Big\}.$$
Then
 $$\limsup_{n \to \infty} \dfrac{1}{n} \log \mathbb{E} |\hat{N}^{\lambda n}_n| \leq \Delta_\lambda.$$
$$\limsup_{n \to \infty} \dfrac{1}{n} \log \mathbb{E} |\hat{N}^{\lambda n}_n| \leq \Delta_\lambda.$$
Note that for any
 $t \in [n, n+1]$
it is always true that
$t \in [n, n+1]$
it is always true that
 $|N_t^{\lambda t}| \leq |\hat{N}^{\lambda n}_n|$
.
$|N_t^{\lambda t}| \leq |\hat{N}^{\lambda n}_n|$
.
Proof of Proposition 3. By the Many-to-One Lemma we have
 $$\begin{align*}
\mathbb{E} |\hat{N}^{\lambda n}_n| &= \mathbb{E} \sum_{u \in N_{n+1}}
\mathbf{1}_{\{\sup_{s \in [n, n+1]}X^u_s \geq \lambda n\}}\\
&= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\sup_{s \in [n, n+1]}\xi_s \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}]\\
&= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}],
\end{align*}$$
$$\begin{align*}
\mathbb{E} |\hat{N}^{\lambda n}_n| &= \mathbb{E} \sum_{u \in N_{n+1}}
\mathbf{1}_{\{\sup_{s \in [n, n+1]}X^u_s \geq \lambda n\}}\\
&= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\sup_{s \in [n, n+1]}\xi_s \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}]\\
&= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}],
\end{align*}$$
where
 $\bar{\xi}_n \,{:\!=}\, \sup_{s \in [n, n+1]} (\xi_s - \xi_{n + 1})$
and
$\bar{\xi}_n \,{:\!=}\, \sup_{s \in [n, n+1]} (\xi_s - \xi_{n + 1})$
and
 $\bar{\xi}_n \stackrel{d}{=} \sup_{s \in [0,1]} \xi_s \stackrel{d}{=} |\mathcal{N}(0,1)|$
under
$\bar{\xi}_n \stackrel{d}{=} \sup_{s \in [0,1]} \xi_s \stackrel{d}{=} |\mathcal{N}(0,1)|$
under
 $\tilde{\mathbb{P}}$
.
$\tilde{\mathbb{P}}$
.
Then, for any
 $\delta \in (0, \lambda)$
we can split the latter expectation as
$\delta \in (0, \lambda)$
we can split the latter expectation as
 $$\begin{align*}
\tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}] &=
\tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| \leq (\lambda - \delta)n\}}
]\\&\quad\, + \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| > (\lambda - \delta)n\}}] {.}
\end{align*}$$
$$\begin{align*}
\tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}] &=
\tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| \leq (\lambda - \delta)n\}}
]\\&\quad\, + \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| > (\lambda - \delta)n\}}] {.}
\end{align*}$$
We shall refer to the first term in the sum as
 $I_1$
and the second one as
$I_1$
and the second one as
 $I_2$
. First we show that the contribution of
$I_2$
. First we show that the contribution of
 $I_1$
is negligibly small as it has a faster than exponential decay rate:
$I_1$
is negligibly small as it has a faster than exponential decay rate:
 $$\begin{align*}
I_1 &= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| \leq (\lambda - \delta)n\}}]\\
&\leq \tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{\bar{\xi}_n \geq \delta n\}} ]\\
&\leq ( \tilde{\mathbb{E}} [ \mathrm{e}^{2\hat{\beta}_0 \tilde{L}_{n+1} + 2\hat{\beta}(n+1)} ]
)^{{1}/{2}} ( \tilde{\mathbb{P}} ( \bar{\xi}_n \geq \delta n ) )^{{1}/{2} }
\end{align*}$$
$$\begin{align*}
I_1 &= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| \leq (\lambda - \delta)n\}}]\\
&\leq \tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{\bar{\xi}_n \geq \delta n\}} ]\\
&\leq ( \tilde{\mathbb{E}} [ \mathrm{e}^{2\hat{\beta}_0 \tilde{L}_{n+1} + 2\hat{\beta}(n+1)} ]
)^{{1}/{2}} ( \tilde{\mathbb{P}} ( \bar{\xi}_n \geq \delta n ) )^{{1}/{2} }
\end{align*}$$
using the Cauchy–Schwarz inequality in the last line. Then, as we know from (16),
 $$\dfrac{1}{n} \log
( \tilde{\mathbb{E}} [ \mathrm{e}^{2\hat{\beta}_0 \tilde{L}_{n+1} + 2\hat{\beta}(n+1)} ]
)^{{1}/{2} } \to \hat{\beta}_0^2 + \hat{\beta} \quad \text{as } n \to \infty{,}$$
$$\dfrac{1}{n} \log
( \tilde{\mathbb{E}} [ \mathrm{e}^{2\hat{\beta}_0 \tilde{L}_{n+1} + 2\hat{\beta}(n+1)} ]
)^{{1}/{2} } \to \hat{\beta}_0^2 + \hat{\beta} \quad \text{as } n \to \infty{,}$$
while, since
 $\bar{\xi}_n \stackrel{d}{=} |\mathcal{N}(0,1)|$
,
$\bar{\xi}_n \stackrel{d}{=} |\mathcal{N}(0,1)|$
,
 $$\dfrac{1}{n^2} \log ( \tilde{\mathbb{P}} ( \bar{\xi}_n \geq \delta n ) )^{{1}/{2} } \to
\dfrac{-\delta^2}{4} \quad \text{as } n \to \infty.$$
$$\dfrac{1}{n^2} \log ( \tilde{\mathbb{P}} ( \bar{\xi}_n \geq \delta n ) )^{{1}/{2} } \to
\dfrac{-\delta^2}{4} \quad \text{as } n \to \infty.$$
Thus
 $$\begin{equation}
\limsup_{n \to \infty} \dfrac{1}{n^2} \log I_1 \leq - \dfrac{\delta^2}{4}.
\end{equation}$$
$$\begin{equation}
\limsup_{n \to \infty} \dfrac{1}{n^2} \log I_1 \leq - \dfrac{\delta^2}{4}.
\end{equation}$$
On the other hand,
 $$\begin{align*}
I_2 &= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| > (\lambda - \delta)n\}}
]\\
&\leq \tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| > (\lambda - \delta)n\}}]\\
&= 2\tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{\xi_{n+1} > (\lambda - \delta)n\}}]\\
&= 2\mathbb{E}|N_{n+1}^{(\lambda - \delta)n}|{,}
\end{align*}$$
$$\begin{align*}
I_2 &= \tilde{\mathbb{E}} [ \mathbf{1}_{\{\xi_{n+1} + \bar{\xi}_n \geq \lambda n\}}
\,\mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| > (\lambda - \delta)n\}}
]\\
&\leq \tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{|\xi_{n+1}| > (\lambda - \delta)n\}}]\\
&= 2\tilde{\mathbb{E}} [ \mathrm{e}^{\hat{\beta}_0 \tilde{L}_{n+1} + \hat{\beta}(n+1)}
\mathbf{1}_{\{\xi_{n+1} > (\lambda - \delta)n\}}]\\
&= 2\mathbb{E}|N_{n+1}^{(\lambda - \delta)n}|{,}
\end{align*}$$
using symmetry in the third line and identity (15) in the fourth line. Thus, from (3) we can see (just as we did in (4)–(6)) that
 $$\begin{equation}
\limsup_{n \to \infty} \dfrac{1}{n} \log I_2 \leq \Delta_{\lambda - \delta}.
\label{eqn19}
\end{equation}$$
$$\begin{equation}
\limsup_{n \to \infty} \dfrac{1}{n} \log I_2 \leq \Delta_{\lambda - \delta}.
\label{eqn19}
\end{equation}$$
From (18) and (19) we have that, for any
 $\delta \in (0, \lambda)$
,
$\delta \in (0, \lambda)$
,
 $$\limsup_{n \to \infty} \dfrac{1}{n} \log \mathbb{E} |\hat{N}^{\lambda n}_n|
= \limsup_{n \to \infty} \dfrac{1}{n} \log(I_1 + I_2) \leq \Delta_{(\lambda-\delta)}.$$
$$\limsup_{n \to \infty} \dfrac{1}{n} \log \mathbb{E} |\hat{N}^{\lambda n}_n|
= \limsup_{n \to \infty} \dfrac{1}{n} \log(I_1 + I_2) \leq \Delta_{(\lambda-\delta)}.$$
Letting
 $\delta \to 0$
and using continuity and monotonicity of
$\delta \to 0$
and using continuity and monotonicity of
 $\Delta_{\lambda}$
as a function of
$\Delta_{\lambda}$
as a function of
 $\lambda$
, we obtain the sought result.
$\lambda$
, we obtain the sought result.
Proposition 3 can now be applied to prove the upper bounds for Theorem 2.
Proposition 4. (Upper bounds for Theorem 2.)
If
 $\lambda < \lambda_{\mathrm{crit}}$
$\lambda < \lambda_{\mathrm{crit}}$
 $(\Delta_\lambda > 0)$
, then
$(\Delta_\lambda > 0)$
, then
 $$\begin{equation}
\limsup_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \leq \Delta_\lambda
\quad \mathbb{P} \text{-a.s.}
\end{equation}$$
$$\begin{equation}
\limsup_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \leq \Delta_\lambda
\quad \mathbb{P} \text{-a.s.}
\end{equation}$$
If
 $\lambda > \lambda_{\mathrm{crit}}$
$\lambda > \lambda_{\mathrm{crit}}$
 $(\Delta_\lambda < 0)$
, then
$(\Delta_\lambda < 0)$
, then
 $$\begin{equation}
\lim_{t \to \infty} |N_t^{\lambda t}| = 0 \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
$$\begin{equation}
\lim_{t \to \infty} |N_t^{\lambda t}| = 0 \quad \mathbb{P} \text{-a.s.}
\end{equation}$$
and
 $$\begin{equation}
\limsup_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0) \leq \Delta_{\lambda} .
\end{equation}$$
$$\begin{equation}
\limsup_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0) \leq \Delta_{\lambda} .
\end{equation}$$
Proof. For any
 $\lambda >0$
, let
$\lambda >0$
, let
 $\hat{N}_n^{\lambda n}$
be as in the previous proposition and fix
$\hat{N}_n^{\lambda n}$
be as in the previous proposition and fix
 $\varepsilon > 0$
. Then the Markov inequality gives
$\varepsilon > 0$
. Then the Markov inequality gives
 $$\mathbb{P} ( |\hat{N}_n^{\lambda n}| > \mathrm{e}^{(\Delta_\lambda + \varepsilon)n} ) \leq
\mathrm{e}^{-(\Delta_\lambda + \varepsilon)n} \mathbb{E} |\hat{N}_n^{\lambda n}|{,}$$
$$\mathbb{P} ( |\hat{N}_n^{\lambda n}| > \mathrm{e}^{(\Delta_\lambda + \varepsilon)n} ) \leq
\mathrm{e}^{-(\Delta_\lambda + \varepsilon)n} \mathbb{E} |\hat{N}_n^{\lambda n}|{,}$$
and from Proposition 3 the right-hand side decays exponentially fast. Therefore, by the Borel–Cantelli lemma,
 $$\mathbb{P} ( \{|\hat{N}_n^{\lambda n}| > \mathrm{e}^{(\Delta_\lambda + \varepsilon)n} \} \text{ i.o.}) = 0.$$
$$\mathbb{P} ( \{|\hat{N}_n^{\lambda n}| > \mathrm{e}^{(\Delta_\lambda + \varepsilon)n} \} \text{ i.o.}) = 0.$$
This is equivalent to saying that
 $$|\hat{N}_n^{\lambda n}| \leq \mathrm{e}^{(\Delta_\lambda + \varepsilon)n} \quad \text{eventually } \mathbb{P} \text{-a.s.}$$
$$|\hat{N}_n^{\lambda n}| \leq \mathrm{e}^{(\Delta_\lambda + \varepsilon)n} \quad \text{eventually } \mathbb{P} \text{-a.s.}$$
So
 $\mathbb{P}$
-almost surely, for all t sufficiently large,
$\mathbb{P}$
-almost surely, for all t sufficiently large,
 $$|N_t^{\lambda t}| \leq |\hat{N}_{\lfloor t \rfloor}^{\lambda \lfloor t \rfloor}| \leq
\mathrm{e}^{(\Delta_\lambda + \varepsilon)\lfloor t \rfloor}{.}$$
$$|N_t^{\lambda t}| \leq |\hat{N}_{\lfloor t \rfloor}^{\lambda \lfloor t \rfloor}| \leq
\mathrm{e}^{(\Delta_\lambda + \varepsilon)\lfloor t \rfloor}{.}$$
Then, if
 $\lambda > \lambda_{\mathrm{crit}}$
, we can take
$\lambda > \lambda_{\mathrm{crit}}$
, we can take
 $\varepsilon$
sufficiently small that
$\varepsilon$
sufficiently small that
 $\Delta_\lambda + \varepsilon < 0$
and hence
$\Delta_\lambda + \varepsilon < 0$
and hence
 $|N_t^{\lambda t}| < 1$
for t large enough, thus proving (21).
$|N_t^{\lambda t}| < 1$
for t large enough, thus proving (21).
If
 $\lambda < \lambda_{\mathrm{crit}}$
, then we get
$\lambda < \lambda_{\mathrm{crit}}$
, then we get
 $$\limsup_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \leq \Delta_\lambda + \varepsilon
\quad \mathbb{P} \text{-a.s.}{,}$$
$$\limsup_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \leq \Delta_\lambda + \varepsilon
\quad \mathbb{P} \text{-a.s.}{,}$$
and letting
 $\varepsilon \to 0$
yields (20).
$\varepsilon \to 0$
yields (20).
Finally, if
 $\lambda > \lambda_{\mathrm{crit}}$
then (22) follows from the Markov inequality and (5):
$\lambda > \lambda_{\mathrm{crit}}$
then (22) follows from the Markov inequality and (5):
 $$\begin{align*}
\limsup_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0) \leq
\limsup_{t \to \infty} {1}/{t} \log \mathbb{E} |N_t^{\lambda t}| = \Delta_{\lambda}.\\
\end{align*}$$
$$\begin{align*}
\limsup_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0) \leq
\limsup_{t \to \infty} {1}/{t} \log \mathbb{E} |N_t^{\lambda t}| = \Delta_{\lambda}.\\
\end{align*}$$
Remark 1. Note that the
 $X \log X$
condition on the offspring distribution has not been required so far. It will be essential in the next subsection.
$X \log X$
condition on the offspring distribution has not been required so far. It will be essential in the next subsection.
2.3. Additive martingales and applications
Recall that under the probability
 $\tilde{\mathbb{P}}$
the branching process together with the spine may be described as follows.
$\tilde{\mathbb{P}}$
the branching process together with the spine may be described as follows.
The process starts with a single spine particle whose path
${(\xi_t)_{t \ge 0}}$
is distributed like a Brownian motion.At instantaneous rate
$\beta_0 \delta_0({\cdot}) + \beta$
along its path, the spine particle splits into
$A({\cdot})$
particles. If splitting took place at position x, then
$$\begin{equation*}
\tilde{\mathbb{P}}(A(x) = n) =
\begin{cases}
q_n& \text{if } x = 0, \\
p_n& \text{if } x \neq 0.
\end{cases}
\end{equation*}$$
Uniformly at random, one of the new particles is selected to continue the spine and thus to stochastically repeat the behaviour of the initial particle starting from x.
The remaining
$A(x) - 1$
particles initiate independent copies of a branching process with branching rate
$\beta_0 \delta_0({\cdot}) + \beta$
and offspring distribution
$A({\cdot})$
as under
$\mathbb{P}^x$
.








We shall now describe a family of martingale changes of measure that will put a certain bias on the motion of the spine particle as well as the birth rate and the offspring distribution along the path of the spine particle. Again, for a detailed discussion the reader is referred to [Reference Hardy and Harris9].
Let us consider a process of the form
 $$\skew1\tilde{M}_t = \bigg[ \prod_{n=1}^{n_t} A_{n} \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t} \skew1\tilde{M}^{(1)}_t
= \bigg[ \prod_{n=1}^{n_t'} A_{n}' \times \prod_{n=1}^{n_t^0} A_{n}^0 \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t} \skew1\tilde{M}^{(1)}_t,$$
$$\skew1\tilde{M}_t = \bigg[ \prod_{n=1}^{n_t} A_{n} \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t} \skew1\tilde{M}^{(1)}_t
= \bigg[ \prod_{n=1}^{n_t'} A_{n}' \times \prod_{n=1}^{n_t^0} A_{n}^0 \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t} \skew1\tilde{M}^{(1)}_t,$$
where
 ${(A_n)_{n \geq 1}}$
,
${(A_n)_{n \geq 1}}$
,
 ${(A^0_n)_{n \geq 1}}$
,
${(A^0_n)_{n \geq 1}}$
,
 ${(A_n')_{n \geq 1}}$
,
${(A_n')_{n \geq 1}}$
,
 ${(n_t)_{t \ge 0}}$
,
${(n_t)_{t \ge 0}}$
,
 ${(n^0_t)_{t \ge 0}}$
, and
${(n^0_t)_{t \ge 0}}$
, and
 ${(n_t')_{t \ge 0}}$
were defined in Subsection 2.1 and
${(n_t')_{t \ge 0}}$
were defined in Subsection 2.1 and
 ${(\skew1\tilde{M}^{(1)})_{t \ge 0}}$
is a non-negative
${(\skew1\tilde{M}^{(1)})_{t \ge 0}}$
is a non-negative
 ${\tilde{\mathbb{P}}}$
-martingale of mean 1 with respect to the filtration
${\tilde{\mathbb{P}}}$
-martingale of mean 1 with respect to the filtration
 ${(\mathcal{G}_t)_{t \ge 0}}$
. The effect of
${(\mathcal{G}_t)_{t \ge 0}}$
. The effect of
 ${\skew1\tilde{M}^{(1)}}$
, if used as a change of measure martingale, is to put some drift on
${\skew1\tilde{M}^{(1)}}$
, if used as a change of measure martingale, is to put some drift on
 ${(\xi_t)_{t \ge 0}}$
.
${(\xi_t)_{t \ge 0}}$
.
For this particular paper we shall take
 ${\skew1\tilde{M}^{(1)}}$
to be either
${\skew1\tilde{M}^{(1)}}$
to be either
 $${\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda \xi_t - {\lambda^2}/{2}t} , \quad t \ge 0}$$
$${\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda \xi_t - {\lambda^2}/{2}t} , \quad t \ge 0}$$
or
 $$\begin{align*}
\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda |\xi_t| - \lambda \tilde{L}_t - {\lambda^2}/{2}t}
= \mathrm{e}^{\lambda \int_0^t \operatorname{sgn}(\xi_s) \,\mathrm{d}\xi_s - {\lambda^2}/{2}t}
, \quad t \ge 0.
\end{align*}$$
$$\begin{align*}
\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda |\xi_t| - \lambda \tilde{L}_t - {\lambda^2}/{2}t}
= \mathrm{e}^{\lambda \int_0^t \operatorname{sgn}(\xi_s) \,\mathrm{d}\xi_s - {\lambda^2}/{2}t}
, \quad t \ge 0.
\end{align*}$$
The first choice is the classical Girsanov martingale, which has the effect of adding constant drift
 ${\lambda}$
to
${\lambda}$
to
 ${(\xi_t)_{t \ge 0}}$
.
${(\xi_t)_{t \ge 0}}$
.
The second choice has the effect of adding instantaneous drift
 ${\lambda \operatorname{sgn}({\cdot})}$
to
${\lambda \operatorname{sgn}({\cdot})}$
to
 ${(\xi_t)_{t \ge 0}}$
, so that if
${(\xi_t)_{t \ge 0}}$
, so that if
 ${\lambda < 0}$
then this is drift of magnitude
${\lambda < 0}$
then this is drift of magnitude
 ${|\lambda|}$
towards the origin, whereas
${|\lambda|}$
towards the origin, whereas
 ${\lambda> 0}$
then this is drift of magnitude
${\lambda> 0}$
then this is drift of magnitude
 ${\lambda}$
away from the origin. For more details one can refer to [Reference Bocharov and Harris1] or [2, ‘Brownian motion with alternating drift’, pp. 128–129]. Alternatively, the effect of this martingale can be seen as adding constant drift
${\lambda}$
away from the origin. For more details one can refer to [Reference Bocharov and Harris1] or [2, ‘Brownian motion with alternating drift’, pp. 128–129]. Alternatively, the effect of this martingale can be seen as adding constant drift
 ${\lambda}$
to the Brownian motion
${\lambda}$
to the Brownian motion
 ${(|\xi_t| - \tilde{L}_t)_{t \ge 0}}$
.
${(|\xi_t| - \tilde{L}_t)_{t \ge 0}}$
.
Recalling that
 ${{m = \smash{\sum_{n \geq 1}} n p_n}}$
and
${{m = \smash{\sum_{n \geq 1}} n p_n}}$
and
 ${{m_0 = \smash{\sum_{n \geq 1}} n q_n}}$
are the means of the offspring distribution due to homogeneous and catalytic branching, we see that
${{m_0 = \smash{\sum_{n \geq 1}} n q_n}}$
are the means of the offspring distribution due to homogeneous and catalytic branching, we see that
 ${(\skew1\tilde{M})_{t \ge 0}}$
can be decomposed into a product of three martingales:
${(\skew1\tilde{M})_{t \ge 0}}$
can be decomposed into a product of three martingales:
 $${\tilde{M}_t = \skew1\tilde{M}^{(1)}_t \skew1\tilde{M}^{(2)}_t \skew1\tilde{M}^{(3)}_t{,}}$$
$${\tilde{M}_t = \skew1\tilde{M}^{(1)}_t \skew1\tilde{M}^{(2)}_t \skew1\tilde{M}^{(3)}_t{,}}$$
where
 $$\tilde M_t^{(2)} = {m^{{n_{t'}}}}{\mkern 1mu} {{\rm{e}}^{ - \hat \beta t}} \times m_0^{n_t^0}{\mkern 1mu} {{\rm{e}}^{ - {{\hat \beta }_0}{{\tilde L}_t}}}$$
$$\tilde M_t^{(2)} = {m^{{n_{t'}}}}{\mkern 1mu} {{\rm{e}}^{ - \hat \beta t}} \times m_0^{n_t^0}{\mkern 1mu} {{\rm{e}}^{ - {{\hat \beta }_0}{{\tilde L}_t}}}$$
and
 $$\skew1\tilde M_t^{(3)} = \prod\limits_{n = 1}^{{n_{t'}}} {\frac{{{A_{n'}}}}{m}} \times \prod\limits_{n = 1}^{n_t^0} {\frac{{A_n^0}}{{{m_0}}}} .$$
$$\skew1\tilde M_t^{(3)} = \prod\limits_{n = 1}^{{n_{t'}}} {\frac{{{A_{n'}}}}{m}} \times \prod\limits_{n = 1}^{n_t^0} {\frac{{A_n^0}}{{{m_0}}}} .$$
When used as the Radon–Nikodym derivative,
 ${(\tilde{M}^{(2)}_t)_{t \ge 0}}$
has the effect of changing the instantaneous jump rate of
${(\tilde{M}^{(2)}_t)_{t \ge 0}}$
has the effect of changing the instantaneous jump rate of
 ${(n_t')_{t \ge 0}}$
from
${(n_t')_{t \ge 0}}$
from
 ${\beta}$
to
${\beta}$
to
 ${m \beta}$
and the jump rate of
${m \beta}$
and the jump rate of
 ${(n_t^0)_{t \ge 0}}$
from
${(n_t^0)_{t \ge 0}}$
from
 ${\beta_0 \delta_0({\cdot})}$
to
${\beta_0 \delta_0({\cdot})}$
to
 ${m_0 \beta_0 \delta_0({\cdot})}$
. The effect of
${m_0 \beta_0 \delta_0({\cdot})}$
. The effect of
 ${(\skew1\tilde{M}^{(3)}_t)_{t \ge 0}}$
is to change the distribution of random variables
${(\skew1\tilde{M}^{(3)}_t)_{t \ge 0}}$
is to change the distribution of random variables
 ${{(A_n')_{n \geq 1}}}$
from
${{(A_n')_{n \geq 1}}}$
from
 ${{(p_k)_{k \geq 1}}}$
to
${{(p_k)_{k \geq 1}}}$
to
 ${{({k}/{m} p_k)_{k \geq 1}}}$
and the distribution of random variables
${{({k}/{m} p_k)_{k \geq 1}}}$
and the distribution of random variables
 ${{(A_n^0)_{n \geq 1}}}$
from
${{(A_n^0)_{n \geq 1}}}$
from
 ${{(q_k)_{k \geq 1}}}$
to
${{(q_k)_{k \geq 1}}}$
to
 ${({k}/{m_0} q_k)_{k \geq 1}}$
(while keeping them all independent). If we now define a new probability measure
${({k}/{m_0} q_k)_{k \geq 1}}$
(while keeping them all independent). If we now define a new probability measure
 ${\tilde{\mathbb{Q}}}$
as
${\tilde{\mathbb{Q}}}$
as
 $$\begin{equation*}
\dfrac{\mathrm{d} \tilde{\mathbb{Q}}}{\mathrm{d} \tilde{\mathbb{P}}} \bigg\vert_{\tilde{\mathcal{F}}_t} =
\skew1\tilde{M}_t , \quad t \ge 0 {,}
\end{equation*}$$
$$\begin{equation*}
\dfrac{\mathrm{d} \tilde{\mathbb{Q}}}{\mathrm{d} \tilde{\mathbb{P}}} \bigg\vert_{\tilde{\mathcal{F}}_t} =
\skew1\tilde{M}_t , \quad t \ge 0 {,}
\end{equation*}$$
then for any events
 ${E_1 \in \mathcal{G}_t}$
,
${E_1 \in \mathcal{G}_t}$
,
 ${E_2 {\in} \sigma ( (n_s')_{0 \leq s \leq t}, (n_s^0)_{0 \leq s \leq t})}$
and
${E_2 {\in} \sigma ( (n_s')_{0 \leq s \leq t}, (n_s^0)_{0 \leq s \leq t})}$
and
 ${E_3 {\in} \sigma (A_1', \ldots, A_{n_t'}';\ A_1^0, \ldots, A_{n_t^0}^0)}$
we have that
${E_3 {\in} \sigma (A_1', \ldots, A_{n_t'}';\ A_1^0, \ldots, A_{n_t^0}^0)}$
we have that
 $$\begin{align*}
\tilde{\mathbb{Q}}(E_1, E_2, E_3) & = \tilde{\mathbb{E}} ( \mathbf{1}_{E_1} \mathbf{1}_{E_2} \mathbf{1}_{E_3} \skew1\tilde{M}^{(1)}_t \skew1\tilde{M}^{(2)}_t \skew1\tilde{M}^{(3)}_t )\\
& = \tilde{\mathbb{E}} [ \mathbf{1}_{E_1} \skew1\tilde{M}^{(1)}_t \tilde{\mathbb{E}} ( \mathbf{1}_{E_2} \skew1\tilde{M}^{(2)}_t \tilde{\mathbb{E}} ( \mathbf{1}_{E_3} \tilde{M}^{(3)}_t \!{\mid}\! \sigma (\mathcal{G}_t, (n_s')_{0 \leq s \leq t}, (n_s^0)_{0 \leq s \leq t}) ) \!{\mid}\! \mathcal{G}_t ) ]
\end{align*}$$
$$\begin{align*}
\tilde{\mathbb{Q}}(E_1, E_2, E_3) & = \tilde{\mathbb{E}} ( \mathbf{1}_{E_1} \mathbf{1}_{E_2} \mathbf{1}_{E_3} \skew1\tilde{M}^{(1)}_t \skew1\tilde{M}^{(2)}_t \skew1\tilde{M}^{(3)}_t )\\
& = \tilde{\mathbb{E}} [ \mathbf{1}_{E_1} \skew1\tilde{M}^{(1)}_t \tilde{\mathbb{E}} ( \mathbf{1}_{E_2} \skew1\tilde{M}^{(2)}_t \tilde{\mathbb{E}} ( \mathbf{1}_{E_3} \tilde{M}^{(3)}_t \!{\mid}\! \sigma (\mathcal{G}_t, (n_s')_{0 \leq s \leq t}, (n_s^0)_{0 \leq s \leq t}) ) \!{\mid}\! \mathcal{G}_t ) ]
\end{align*}$$
so it can be seen that the effects of
 ${\skew1\tilde{M}^{(1)}}$
,
${\skew1\tilde{M}^{(1)}}$
,
 ${\skew1\tilde{M}^{(2)}}$
, and
${\skew1\tilde{M}^{(2)}}$
, and
 ${\tilde{M}^{(3)}}$
superimpose. Thus, under
${\tilde{M}^{(3)}}$
superimpose. Thus, under
 ${\tilde{\mathbb{Q}}}$
the branching process has the following description.
${\tilde{\mathbb{Q}}}$
the branching process has the following description.
• The process starts with a single spine particle whose path
 ${(\xi_t)_{t \ge 0}}$
is distributed like a Brownian motion with drift imposed by
${(\xi_t)_{t \ge 0}}$
is distributed like a Brownian motion with drift imposed by
 ${\skew1\tilde{M}^{(1)}}$
.
${\skew1\tilde{M}^{(1)}}$
.
• At instantaneous rate
 ${m_0 \beta_0 \delta_0({\cdot}) + m \beta}$
along its path the spine particle splits into
${m_0 \beta_0 \delta_0({\cdot}) + m \beta}$
along its path the spine particle splits into
 ${A({\cdot})}$
particles. If splitting took place at position x, then
${A({\cdot})}$
particles. If splitting took place at position x, then
 $$\begin{equation*}
\tilde{\mathbb{Q}}(A(x) = n) =
\begin{cases}
\dfrac{n}{m_0}q_n& \text{if } x = 0, \\
\dfrac{n}{m}p_n& \text{if } x \neq 0.
\end{cases}
\end{equation*}$$
$$\begin{equation*}
\tilde{\mathbb{Q}}(A(x) = n) =
\begin{cases}
\dfrac{n}{m_0}q_n& \text{if } x = 0, \\
\dfrac{n}{m}p_n& \text{if } x \neq 0.
\end{cases}
\end{equation*}$$
• Uniformly at random, one of the new particles is selected to continue the spine and thus to stochastically repeat the behaviour of the initial particle starting from x.
• The remaining
 ${A(x) - 1}$
particles initiate independent unbiased copies of a branching process with branching rate
${A(x) - 1}$
particles initiate independent unbiased copies of a branching process with branching rate
 ${\beta_0 \delta_0({\cdot}) + \beta}$
and offspring distribution
${\beta_0 \delta_0({\cdot}) + \beta}$
and offspring distribution
 ${A({\cdot})}$
as under
${A({\cdot})}$
as under
 ${\mathbb{P}^x}$
.
${\mathbb{P}^x}$
.
Suppose now that, for all
 ${t \ge 0}$
,
${t \ge 0}$
,
 ${\skew1\tilde{M}^{(1)}_t}$
can be represented as
${\skew1\tilde{M}^{(1)}_t}$
can be represented as
 $$\begin{equation}
\skew1\tilde{M}^{(1)}_t = \sum_{u \in N_t} M^{(1)}_t(u) \mathbf{1}_{\{ {{\mathop{\rm node}\nolimits} _t}(\xi ) = u \}},
\end{equation}$$
$$\begin{equation}
\skew1\tilde{M}^{(1)}_t = \sum_{u \in N_t} M^{(1)}_t(u) \mathbf{1}_{\{ {{\mathop{\rm node}\nolimits} _t}(\xi ) = u \}},
\end{equation}$$
where, for all
 ${u \in N_t}$
,
${u \in N_t}$
,
 ${M^{(1)}_t(u)}$
is
${M^{(1)}_t(u)}$
is
 ${\mathcal{F}_t}$
-measurable. For example, if
${\mathcal{F}_t}$
-measurable. For example, if
 ${\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda |\xi_t| - \lambda \tilde{L}_t - {\lambda^2}/{2}t}}$
, then we get the required representation by taking
${\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda |\xi_t| - \lambda \tilde{L}_t - {\lambda^2}/{2}t}}$
, then we get the required representation by taking
 ${M^{(1)}_t(u) = \mathrm{e}^{\lambda |X^u_t| - \lambda L^u_t - {\lambda^2}/{2}t}}$
. If we define
${M^{(1)}_t(u) = \mathrm{e}^{\lambda |X^u_t| - \lambda L^u_t - {\lambda^2}/{2}t}}$
. If we define
 $$\begin{equation}
M_t \,{:\!=}\, \sum_{u \in N_t} M^{(1)}_t(u) \,\mathrm{e}^{- \hat{\beta}_0 L_t^u - \hat{\beta}t}
, \quad t \ge 0 {,}
\end{equation}$$
$$\begin{equation}
M_t \,{:\!=}\, \sum_{u \in N_t} M^{(1)}_t(u) \,\mathrm{e}^{- \hat{\beta}_0 L_t^u - \hat{\beta}t}
, \quad t \ge 0 {,}
\end{equation}$$
then
 ${(M_t)_{t \ge 0}}$
is a unit-mean
${(M_t)_{t \ge 0}}$
is a unit-mean
 ${\mathbb{P}}$
-martingale such that
${\mathbb{P}}$
-martingale such that
 $$M_t = \tilde{\mathbb{E}} ( \skew1\tilde{M}_t \!{\mid}\!\mathcal{F}_t ).$$
$$M_t = \tilde{\mathbb{E}} ( \skew1\tilde{M}_t \!{\mid}\!\mathcal{F}_t ).$$
To see this, note that
 $$\begin{align*}
\tilde{\mathbb{E}} ( \skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t ) &= \tilde{\mathbb{E}} \bigg( \bigg[ \prod_{n=1}^{n_t} A_n \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t} \skew1\tilde{M}^{(1)}_t \biggm\vert \mathcal{F}_t \bigg)\\&= \tilde{\mathbb{E}} \bigg( \sum_{u \in N_t} \bigg( \bigg[ \prod_{v<u} A_v \bigg] \,\mathrm{e}^{- \hat{\beta}t -
\hat{\beta}_0 L^u_t} \tilde{M}^{(1)}_t(u) \mathbf{1}_{\{{{\mathop{\rm node}\nolimits} _t}(\xi ) = u\}} \bigg) \biggm\vert \mathcal{F}_t \bigg)\\
&= \sum_{u \in N_t} \bigg( \bigg[ \prod_{v<u} A_v \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 L^u_t}
\skew1\tilde{M}^{(1)}_t(u) \tilde{\mathbb{P}} ( u \in \xi\!{\mid}\!\mathcal{F}_t ) \bigg)\\
&= \sum_{u \in N_t} \bigg( \bigg[ \prod_{v<u} A_v \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 L^u_t}
\skew1\tilde{M}^{(1)}_t(u) \prod_{v<u} \dfrac{1}{A_v} \bigg)\\&= M_t.
\end{align*}$$
$$\begin{align*}
\tilde{\mathbb{E}} ( \skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t ) &= \tilde{\mathbb{E}} \bigg( \bigg[ \prod_{n=1}^{n_t} A_n \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t} \skew1\tilde{M}^{(1)}_t \biggm\vert \mathcal{F}_t \bigg)\\&= \tilde{\mathbb{E}} \bigg( \sum_{u \in N_t} \bigg( \bigg[ \prod_{v<u} A_v \bigg] \,\mathrm{e}^{- \hat{\beta}t -
\hat{\beta}_0 L^u_t} \tilde{M}^{(1)}_t(u) \mathbf{1}_{\{{{\mathop{\rm node}\nolimits} _t}(\xi ) = u\}} \bigg) \biggm\vert \mathcal{F}_t \bigg)\\
&= \sum_{u \in N_t} \bigg( \bigg[ \prod_{v<u} A_v \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 L^u_t}
\skew1\tilde{M}^{(1)}_t(u) \tilde{\mathbb{P}} ( u \in \xi\!{\mid}\!\mathcal{F}_t ) \bigg)\\
&= \sum_{u \in N_t} \bigg( \bigg[ \prod_{v<u} A_v \bigg] \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 L^u_t}
\skew1\tilde{M}^{(1)}_t(u) \prod_{v<u} \dfrac{1}{A_v} \bigg)\\&= M_t.
\end{align*}$$
For the martingale property of
 ${(M_t)_{t \ge 0}}$
, we then check that, for any
${(M_t)_{t \ge 0}}$
, we then check that, for any
 ${s \leq t}$
and an event
${s \leq t}$
and an event
 ${A \in \mathcal{F}_s}$
,
${A \in \mathcal{F}_s}$
,
 $$\begin{align*}
\mathbb{E}(M_t \mathbf{1}_A)
& = \tilde{\mathbb{E}}(M_t \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}}(\skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t) \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}} (\skew1\tilde{M}_t \mathbf{1}_A \!{\mid}\! \mathcal{F}_t) ) \\
& = \tilde{\mathbb{E}} (\skew1\tilde{M}_t \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} (\skew1\tilde{M}_s \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}}
(\skew1\tilde{M}_s \mathbf{1}_A \!{\mid}\! \mathcal{F}_s) ) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}}(\skew1\tilde{M}_s \!{\mid}\! \mathcal{F}_s) \mathbf{1}_A)\\
& = \tilde{\mathbb{E}}(M_s \mathbf{1}_A) \\ & = \mathbb{E}(M_s \mathbf{1}_A).
\end{align*}$$
$$\begin{align*}
\mathbb{E}(M_t \mathbf{1}_A)
& = \tilde{\mathbb{E}}(M_t \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}}(\skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t) \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}} (\skew1\tilde{M}_t \mathbf{1}_A \!{\mid}\! \mathcal{F}_t) ) \\
& = \tilde{\mathbb{E}} (\skew1\tilde{M}_t \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} (\skew1\tilde{M}_s \mathbf{1}_A) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}}
(\skew1\tilde{M}_s \mathbf{1}_A \!{\mid}\! \mathcal{F}_s) ) \\
& = \tilde{\mathbb{E}} ( \tilde{\mathbb{E}}(\skew1\tilde{M}_s \!{\mid}\! \mathcal{F}_s) \mathbf{1}_A)\\
& = \tilde{\mathbb{E}}(M_s \mathbf{1}_A) \\ & = \mathbb{E}(M_s \mathbf{1}_A).
\end{align*}$$
If we now define
 ${\mathbb{Q} \,{:\!=}\, \tilde{\mathbb{Q}} \vert_{\mathcal{F}_\infty}}$
, then we would have that
${\mathbb{Q} \,{:\!=}\, \tilde{\mathbb{Q}} \vert_{\mathcal{F}_\infty}}$
, then we would have that
 $${\dfrac{\mathrm{d}\mathbb{Q}}{\mathrm{d}\mathbb{P}} \bigg\vert_{\mathcal{F}_t} = M_t , \quad t \ge 0 {,}}$$
$${\dfrac{\mathrm{d}\mathbb{Q}}{\mathrm{d}\mathbb{P}} \bigg\vert_{\mathcal{F}_t} = M_t , \quad t \ge 0 {,}}$$
since for any event
 ${A \in \mathcal{F}_t}$
,
${A \in \mathcal{F}_t}$
,
 $$\begin{align*}
\mathbb{Q}(A) = \tilde{\mathbb{Q}}(A) = \tilde{\mathbb{E}}(\mathbf{1}_A
\skew1\tilde{M}_t) = \tilde{\mathbb{E}} (\tilde{\mathbb{E}}( \mathbf{1}_A
\skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t) ) = \tilde{\mathbb{E}} ( \mathbf{1}_A
\tilde{\mathbb{E}}(\skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t) ) =
\tilde{\mathbb{E}}(\mathbf{1}_A M_t) = \mathbb{E} (\mathbf{1}_A M_t).
\end{align*}$$
$$\begin{align*}
\mathbb{Q}(A) = \tilde{\mathbb{Q}}(A) = \tilde{\mathbb{E}}(\mathbf{1}_A
\skew1\tilde{M}_t) = \tilde{\mathbb{E}} (\tilde{\mathbb{E}}( \mathbf{1}_A
\skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t) ) = \tilde{\mathbb{E}} ( \mathbf{1}_A
\tilde{\mathbb{E}}(\skew1\tilde{M}_t \!{\mid}\! \mathcal{F}_t) ) =
\tilde{\mathbb{E}}(\mathbf{1}_A M_t) = \mathbb{E} (\mathbf{1}_A M_t).
\end{align*}$$
Since
 ${(M_t)_{t \ge 0}}$
is always a non-negative
${(M_t)_{t \ge 0}}$
is always a non-negative
 ${\mathbb{P}}$
-martingale, it must converge
${\mathbb{P}}$
-martingale, it must converge
 ${\mathbb{P}}$
-almost surely to some non-negative limit
${\mathbb{P}}$
-almost surely to some non-negative limit
 ${M_\infty}$
. Particularly interesting are those martingales whose limit is not almost surely 0.
${M_\infty}$
. Particularly interesting are those martingales whose limit is not almost surely 0.
When investigating whether the limit of M is
 ${\mathbb{P}}$
-almost surely 0 or not, we shall make use of the following result, commonly known as the spine decomposition.
${\mathbb{P}}$
-almost surely 0 or not, we shall make use of the following result, commonly known as the spine decomposition.
Lemma 2. (Spine decomposition.) Let
 ${(M_t)_{t \ge 0}}$
be the
${(M_t)_{t \ge 0}}$
be the
 ${\mathbb{P}}$
-martingale of the form (24) constructed from some given
${\mathbb{P}}$
-martingale of the form (24) constructed from some given
 ${\tilde{\mathbb{P}}}$
-martingale
${\tilde{\mathbb{P}}}$
-martingale
 ${(\skew1\tilde{M}^{(1)}_t)_{t \ge 0}}$
with the property (23). Then
${(\skew1\tilde{M}^{(1)}_t)_{t \ge 0}}$
with the property (23). Then
 $$\tilde{\mathbb{Q}} ( M_t \!{\mid}\! \tilde{\mathcal{G}}_\infty ) = spine(t) + \sum_{n=1}^{n_t} (A_n - 1) spine(S_n),$$
$$\tilde{\mathbb{Q}} ( M_t \!{\mid}\! \tilde{\mathcal{G}}_\infty ) = spine(t) + \sum_{n=1}^{n_t} (A_n - 1) spine(S_n),$$
where
 $${\operatorname{spine}(t) = \skew1\tilde{M}^{(1)}_t \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t}}$$
$${\operatorname{spine}(t) = \skew1\tilde{M}^{(1)}_t \,\mathrm{e}^{- \hat{\beta}t - \hat{\beta}_0 \tilde{L}_t}}$$
and, as defined earlier,
 ${S_n}$
is the time of the nth branching event along the path of the spine process and
${S_n}$
is the time of the nth branching event along the path of the spine process and
 ${A_n}$
is the corresponding number of offspring produced.
${A_n}$
is the corresponding number of offspring produced.
Note that we have used
 ${\tilde{\mathbb{Q}}}$
to denote the expectation corresponding to probability measure
${\tilde{\mathbb{Q}}}$
to denote the expectation corresponding to probability measure
 ${\tilde{\mathbb{Q}}}$
, which is a common practice.
${\tilde{\mathbb{Q}}}$
, which is a common practice.
We have already used the fact that if
 ${A \in \mathcal{F}_t}$
for some
${A \in \mathcal{F}_t}$
for some
 ${t \ge 0}$
, then
${t \ge 0}$
, then
 $${\mathbb{Q}(A) = \int_A M_t \,\mathrm{d}\mathbb{P}.}$$
$${\mathbb{Q}(A) = \int_A M_t \,\mathrm{d}\mathbb{P}.}$$
Let us also recall that if
 ${A \in \mathcal{F}_\infty}$
, then
${A \in \mathcal{F}_\infty}$
, then
 $$\begin{equation}
\mathbb{Q}(A) = \int_A M_\infty \,\mathrm{d}\mathbb{P} + \mathbb{Q} \Big( A \cap \Big\{ \limsup_{t \to \infty} M_t = \infty\Big\} \Big).
\end{equation}$$
$$\begin{equation}
\mathbb{Q}(A) = \int_A M_\infty \,\mathrm{d}\mathbb{P} + \mathbb{Q} \Big( A \cap \Big\{ \limsup_{t \to \infty} M_t = \infty\Big\} \Big).
\end{equation}$$
The latter identity can be found in [7, p. 241].
Proposition 5. Let
 $$\begin{equation}
M_t^\pm \,{:\!=}\, \sum_{u \in N_t} \,\mathrm{e}^{- \hat{\beta}_0 |X^u_t| - \frac{1}{2} \hat{\beta}_0^2 t - \hat{\beta} t} , \quad t \ge 0{,}
\end{equation}$$
$$\begin{equation}
M_t^\pm \,{:\!=}\, \sum_{u \in N_t} \,\mathrm{e}^{- \hat{\beta}_0 |X^u_t| - \frac{1}{2} \hat{\beta}_0^2 t - \hat{\beta} t} , \quad t \ge 0{,}
\end{equation}$$
be the
 ${\mathbb{P}}$
-martingale of the form (24) constructed by taking
${\mathbb{P}}$
-martingale of the form (24) constructed by taking
 $$\begin{align}
\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{- \hat{\beta}_0 |\xi_t| + \hat{\beta}_0 \tilde{L}_t - \frac{1}{2} \hat{\beta}_0^2 t} , \quad t \ge 0.\label{eqn27}\end{align}$$
$$\begin{align}
\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{- \hat{\beta}_0 |\xi_t| + \hat{\beta}_0 \tilde{L}_t - \frac{1}{2} \hat{\beta}_0^2 t} , \quad t \ge 0.\label{eqn27}\end{align}$$
 $$\begin{align}\nonumber
\end{align}$$
$$\begin{align}\nonumber
\end{align}$$
(a) If condition (10) (the
 ${X \log X}$
condition on the offspring distribution) is satisfied, then
${X \log X}$
condition on the offspring distribution) is satisfied, then
 $$M^\pm_\infty > 0 \quad \mathbb{P} \text{-a.s.}$$
$$M^\pm_\infty > 0 \quad \mathbb{P} \text{-a.s.}$$
(b) If condition (10) is not satisfied, then
 $$M^\pm_\infty = 0 \quad \mathbb{P} \text{-a.s.}$$
$$M^\pm_\infty = 0 \quad \mathbb{P} \text{-a.s.}$$
Proof. Let
 ${\mathbb{Q}^\pm}$
and
${\mathbb{Q}^\pm}$
and
 ${\tilde{\mathbb{Q}}^\pm}$
be the probability measures associated with martingales (26) and (27) as previously described in this subsection.
${\tilde{\mathbb{Q}}^\pm}$
be the probability measures associated with martingales (26) and (27) as previously described in this subsection.
A standard argument which relies only on point-recurrence of
 ${(\xi_t)_{t \ge 0}}$
under
${(\xi_t)_{t \ge 0}}$
under
 ${\tilde{\mathbb{P}}}$
(see e.g. [Reference Harris and Harris10]) tells us that
${\tilde{\mathbb{P}}}$
(see e.g. [Reference Harris and Harris10]) tells us that
 ${\mathbb{P}(M^\pm_\infty > 0) \in \{0, 1\}}$
.
${\mathbb{P}(M^\pm_\infty > 0) \in \{0, 1\}}$
.
So if we could show that
 ${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) = 1}$
, then by taking
${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) = 1}$
, then by taking
 ${A = \Omega}$
in (25) we would get
${A = \Omega}$
in (25) we would get
 ${1 = \mathbb{E} M^\pm_\infty + 1}$
and hence
${1 = \mathbb{E} M^\pm_\infty + 1}$
and hence
 ${M^\pm_\infty = 0\ \mathbb{P}}$
-almost surely.
${M^\pm_\infty = 0\ \mathbb{P}}$
-almost surely.
On the other hand, if we could show that
 ${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) = 0}$
then, again by taking
${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) = 0}$
then, again by taking
 ${A = \Omega}$
in (25), we would get
${A = \Omega}$
in (25), we would get
 ${1 = \mathbb{E} M^\pm_\infty + 0}$
and hence
${1 = \mathbb{E} M^\pm_\infty + 0}$
and hence
 ${\mathbb{P}(M^\pm_\infty > 0) > 0}$
, which from the 0–1 law above is the same as
${\mathbb{P}(M^\pm_\infty > 0) > 0}$
, which from the 0–1 law above is the same as
 ${\mathbb{P}(M^\pm_\infty > 0) = 1}$
.
${\mathbb{P}(M^\pm_\infty > 0) = 1}$
.
Thus, to prove the proposition, it is sufficient to show that, if condition (10) is satisfied, then
 ${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) = 0}$
, and if condition (10) is not satisfied, then
${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) = 0}$
, and if condition (10) is not satisfied, then
 ${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) =1}$
.
${\mathbb{Q}^\pm(\limsup_{t \to \infty} M^\pm_t = \infty) =1}$
.
Let us observe that because of the effect of martingale
 ${\skew1\tilde{M}^{(3)}}$
, for any choice of
${\skew1\tilde{M}^{(3)}}$
, for any choice of
 ${c > 0}$
we have
${c > 0}$
we have
 $$\begin{align*}
\dfrac{1}{c} \tilde{\mathbb{E}} (A_1' \log A_1' ) &= \dfrac{1}{c} m \tilde{\mathbb{Q}}^\pm (\log A_1' ) \\
&= m \int_0^\infty \tilde{\mathbb{Q}}^\pm\bigg( \dfrac{1}{c} \log A_1' \geq t\bigg) \,\mathrm{d}t \\
&= m \sum_{n=0}^\infty \int_n^{n+1} \tilde{\mathbb{Q}}^\pm (\log A_1' \geq ct ) \,\mathrm{d}t.
\end{align*}$$
$$\begin{align*}
\dfrac{1}{c} \tilde{\mathbb{E}} (A_1' \log A_1' ) &= \dfrac{1}{c} m \tilde{\mathbb{Q}}^\pm (\log A_1' ) \\
&= m \int_0^\infty \tilde{\mathbb{Q}}^\pm\bigg( \dfrac{1}{c} \log A_1' \geq t\bigg) \,\mathrm{d}t \\
&= m \sum_{n=0}^\infty \int_n^{n+1} \tilde{\mathbb{Q}}^\pm (\log A_1' \geq ct ) \,\mathrm{d}t.
\end{align*}$$
Hence, by monotonicity of
 ${\tilde{\mathbb{Q}}^\pm (\log A_1' \geq ct)}$
, we have
${\tilde{\mathbb{Q}}^\pm (\log A_1' \geq ct)}$
, we have
 $$\begin{equation}
m c \sum_{n=1}^\infty \tilde{\mathbb{Q}}^\pm (\log A_1' \geq c n ) \leq \tilde{\mathbb{E}} (A_1' \log A_1' ) \leq m c \sum_{n=0}^\infty \tilde{\mathbb{Q}}^\pm (\log A_1' \geq c n ).
\end{equation}$$
$$\begin{equation}
m c \sum_{n=1}^\infty \tilde{\mathbb{Q}}^\pm (\log A_1' \geq c n ) \leq \tilde{\mathbb{E}} (A_1' \log A_1' ) \leq m c \sum_{n=0}^\infty \tilde{\mathbb{Q}}^\pm (\log A_1' \geq c n ).
\end{equation}$$
Then, since
 ${(A_n')_{n \geq 1}}$
are i.i.d. random variables, it follows from (28) that
${(A_n')_{n \geq 1}}$
are i.i.d. random variables, it follows from (28) that
 $$\begin{align*}
\sum_{n = 1}^\infty \tilde{\mathbb{Q}}^\pm (A_n' \geq \mathrm{e}^{cn} ) =
\sum_{n = 1}^\infty \tilde{\mathbb{Q}}^\pm (\log A_1' \geq c n) < \infty
\quad \iff \quad \tilde{\mathbb{E}} (A_1' \log A_1' ) = \sum_{n=1}^\infty p_n n \log n < \infty{,}
\end{align*}$$
$$\begin{align*}
\sum_{n = 1}^\infty \tilde{\mathbb{Q}}^\pm (A_n' \geq \mathrm{e}^{cn} ) =
\sum_{n = 1}^\infty \tilde{\mathbb{Q}}^\pm (\log A_1' \geq c n) < \infty
\quad \iff \quad \tilde{\mathbb{E}} (A_1' \log A_1' ) = \sum_{n=1}^\infty p_n n \log n < \infty{,}
\end{align*}$$
and then, by the first and second Borel–Cantelli lemmas,
 $$\begin{equation}
\tilde{\mathbb{Q}}^\pm ( \{ A_n' \geq \mathrm{e}^{cn}\} \text{ i.o.}) =
\begin{cases}
0& \displaystyle \text{if } \sum_{n=1}^\infty p_n n \log n < \infty, \\\\
1& \displaystyle \text{if } \sum_{n=1}^\infty p_n n \log n = \infty.
\end{cases}
\end{equation}$$
$$\begin{equation}
\tilde{\mathbb{Q}}^\pm ( \{ A_n' \geq \mathrm{e}^{cn}\} \text{ i.o.}) =
\begin{cases}
0& \displaystyle \text{if } \sum_{n=1}^\infty p_n n \log n < \infty, \\\\
1& \displaystyle \text{if } \sum_{n=1}^\infty p_n n \log n = \infty.
\end{cases}
\end{equation}$$
An identical argument gives
 $$\begin{equation}
\tilde{\mathbb{Q}}^\pm ( \{ A_n^0 \geq \mathrm{e}^{cn}\} \text{ i.o.}) =
\begin{cases}
0& \displaystyle \text{if } \sum_{n=1}^\infty q_n n \log n < \infty, \\\\
1& \displaystyle \text{if } \sum_{n=1}^\infty q_n n \log n = \infty.
\end{cases}
\end{equation}$$
$$\begin{equation}
\tilde{\mathbb{Q}}^\pm ( \{ A_n^0 \geq \mathrm{e}^{cn}\} \text{ i.o.}) =
\begin{cases}
0& \displaystyle \text{if } \sum_{n=1}^\infty q_n n \log n < \infty, \\\\
1& \displaystyle \text{if } \sum_{n=1}^\infty q_n n \log n = \infty.
\end{cases}
\end{equation}$$
Let us emphasize that dichotomies (29) and (30) hold for any choice of
 ${c>0}$
.
${c>0}$
.
(a) Assume now that condition (10) holds, and recall from Lemma 2.3 that
 $$\begin{align*}
\tilde{\mathbb{Q}}^\pm ( M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty ) &=
\operatorname{spine}(t) + \sum_{n=1}^{n_t'} (A_n' - 1) \operatorname{spine}(S_n') + \sum_{n=1}^{n^0_t} (A_n^0 - 1) \operatorname{spine}(S^0_n)\\&\leq 1 + \sum_{n=1}^\infty (A_n' - 1) \operatorname{spine}(S_n') + \sum_{n=1}^\infty (A_n^0 - 1) \operatorname{spine}(S^0_n),
\end{align*}$$
$$\begin{align*}
\tilde{\mathbb{Q}}^\pm ( M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty ) &=
\operatorname{spine}(t) + \sum_{n=1}^{n_t'} (A_n' - 1) \operatorname{spine}(S_n') + \sum_{n=1}^{n^0_t} (A_n^0 - 1) \operatorname{spine}(S^0_n)\\&\leq 1 + \sum_{n=1}^\infty (A_n' - 1) \operatorname{spine}(S_n') + \sum_{n=1}^\infty (A_n^0 - 1) \operatorname{spine}(S^0_n),
\end{align*}$$
where
 $${\operatorname{spine}(t) = \mathrm{e}^{- \hat{\beta}_0 |\xi_t| - \frac{1}{2} \hat{\beta}_0^2t - \hat{\beta}t}.}$$
$${\operatorname{spine}(t) = \mathrm{e}^{- \hat{\beta}_0 |\xi_t| - \frac{1}{2} \hat{\beta}_0^2t - \hat{\beta}t}.}$$
Recall that martingale (27), when used as the Radon–Nikodym derivative, has the effect of putting constant drift of magnitude
 ${\hat{\beta}_0}$
towards the origin onto
${\hat{\beta}_0}$
towards the origin onto
 ${(\xi_t)_{t \ge 0}}$
or, equivalently, the effect of putting constant drift
${(\xi_t)_{t \ge 0}}$
or, equivalently, the effect of putting constant drift
 ${\hat{\beta}_0}$
onto
${\hat{\beta}_0}$
onto
 ${(\tilde{L}_t - |\xi_t|)_{t \ge 0}}$
. Thus we can see that,
${(\tilde{L}_t - |\xi_t|)_{t \ge 0}}$
. Thus we can see that,
 ${\tilde{\mathbb{Q}}^\pm}$
-almost surely,
${\tilde{\mathbb{Q}}^\pm}$
-almost surely,
 $$\dfrac{\tilde{L}_t - |\xi_t|}{t} \to \hat{\beta}_0,\quad
\dfrac{\xi_t}{t} \to 0,\quad
\dfrac{\tilde{L}_t}{t} \to \hat{\beta}_0$$
$$\dfrac{\tilde{L}_t - |\xi_t|}{t} \to \hat{\beta}_0,\quad
\dfrac{\xi_t}{t} \to 0,\quad
\dfrac{\tilde{L}_t}{t} \to \hat{\beta}_0$$
and consequently
 $$\frac{n_t'}{t} \to m \beta\quad\text{and}\quad
\frac{n^0_t}{\tilde{L}_t} \to m_0 \beta_0\quad \text{as $t \to \infty$.}$$
$$\frac{n_t'}{t} \to m \beta\quad\text{and}\quad
\frac{n^0_t}{\tilde{L}_t} \to m_0 \beta_0\quad \text{as $t \to \infty$.}$$
It follows that
 ${S_n' \sim \frac{1}{(m \beta)}n}$
as
${S_n' \sim \frac{1}{(m \beta)}n}$
as
 ${n \to \infty\ \tilde{\mathbb{Q}}^\pm}$
-a.s., so for any
${n \to \infty\ \tilde{\mathbb{Q}}^\pm}$
-a.s., so for any
 ${\delta > 0}$
${\delta > 0}$
 $$\begin{equation}
\mathrm{e}^{-(K+\delta)n} \leq \operatorname{spine}(S_n') \leq \mathrm{e}^{-(K-\delta)n} \quad \text{eventually } \tilde{\mathbb{Q}}^\pm \text{-a.s.}{,}
\end{equation}$$
$$\begin{equation}
\mathrm{e}^{-(K+\delta)n} \leq \operatorname{spine}(S_n') \leq \mathrm{e}^{-(K-\delta)n} \quad \text{eventually } \tilde{\mathbb{Q}}^\pm \text{-a.s.}{,}
\end{equation}$$
where
 $$K = \bigg(\frac{1}{2} \hat{\beta}_0^2 + \hat{\beta}\bigg)\frac{1}{m \beta} > 0{,}$$
$$K = \bigg(\frac{1}{2} \hat{\beta}_0^2 + \hat{\beta}\bigg)\frac{1}{m \beta} > 0{,}$$
which together with the first line of (29) yields
 $${\sum_{n=1}^\infty (A_n' - 1) \operatorname{spine}(S_n') < \infty \quad \tilde{\mathbb{Q}}^\pm \text{-a.s.}}$$
$${\sum_{n=1}^\infty (A_n' - 1) \operatorname{spine}(S_n') < \infty \quad \tilde{\mathbb{Q}}^\pm \text{-a.s.}}$$
Similarly,
 $$S^0_n \sim \frac{1}{m_0 \beta_0 \hat{\beta}_0}n\quad \text{as $n \to \infty$,}$$
$$S^0_n \sim \frac{1}{m_0 \beta_0 \hat{\beta}_0}n\quad \text{as $n \to \infty$,}$$
which together with the first line of (30) yields
 $${\sum_{n=1}^\infty (A^0_n - 1) \operatorname{spine}(S^0_n) < \infty \quad \tilde{\mathbb{Q}}^\pm \text{-a.s.}}$$
$${\sum_{n=1}^\infty (A^0_n - 1) \operatorname{spine}(S^0_n) < \infty \quad \tilde{\mathbb{Q}}^\pm \text{-a.s.}}$$
We have thus shown that
 $${\limsup_{t \to \infty} \tilde{\mathbb{Q}}^\pm ( M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty ) < \infty.}$$
$${\limsup_{t \to \infty} \tilde{\mathbb{Q}}^\pm ( M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty ) < \infty.}$$
Applying conditional Fatou’s lemma we get
 $$\tilde{\mathbb{Q}}^\pm ( \liminf_{t \to \infty} M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty) \leq
\liminf_{t \to \infty} \tilde{\mathbb{Q}}^\pm (M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty) \leq
\limsup_{t \to \infty} \tilde{\mathbb{Q}}^\pm (M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty) < \infty{,}$$
$$\tilde{\mathbb{Q}}^\pm ( \liminf_{t \to \infty} M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty) \leq
\liminf_{t \to \infty} \tilde{\mathbb{Q}}^\pm (M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty) \leq
\limsup_{t \to \infty} \tilde{\mathbb{Q}}^\pm (M^\pm_t \!{\mid}\! \tilde{\mathcal{G}}_\infty) < \infty{,}$$
which implies that
 ${\liminf_{t \to \infty} M^\pm_t < \infty}$
${\liminf_{t \to \infty} M^\pm_t < \infty}$
 ${\tilde{\mathbb{Q}}^\pm}$
-a.s. and hence also
${\tilde{\mathbb{Q}}^\pm}$
-a.s. and hence also
 ${\mathbb{Q}^\pm}$
-a.s. (as
${\mathbb{Q}^\pm}$
-a.s. (as
 ${\{ \liminf_{t \to \infty} M^\pm_t < \infty \} \in \mathcal{F}_\infty}$
). Then, since
${\{ \liminf_{t \to \infty} M^\pm_t < \infty \} \in \mathcal{F}_\infty}$
). Then, since
 ${{1}/{M^\pm}}$
is a positive supermartingale under
${{1}/{M^\pm}}$
is a positive supermartingale under
 ${\mathbb{Q}^\pm}$
(in fact a true martingale as there is no extinction), it must converge, so
${\mathbb{Q}^\pm}$
(in fact a true martingale as there is no extinction), it must converge, so
 $${\limsup_{t \to \infty} M^\pm_t = \liminf_{t \to \infty} M^\pm_t < \infty \quad \tilde{\mathbb{Q}}^\pm\text{-a.s.}{,}}$$
$${\limsup_{t \to \infty} M^\pm_t = \liminf_{t \to \infty} M^\pm_t < \infty \quad \tilde{\mathbb{Q}}^\pm\text{-a.s.}{,}}$$
which is sufficient to prove part (a) of the proposition.
(b) Assume now that
 ${\sum_{n \geq 1} p_n n \log n = \infty}$
. Then, counting only particles born from the spine, we obtain
${\sum_{n \geq 1} p_n n \log n = \infty}$
. Then, counting only particles born from the spine, we obtain
 $$M^\pm_{S_n'} \geq A_n' \operatorname{spine}(S_n'){,}$$
$$M^\pm_{S_n'} \geq A_n' \operatorname{spine}(S_n'){,}$$
so that the first inequality in (31) and the second line in (29) give us that
 ${\tilde{\mathbb{Q}}^\pm}$
, and hence also
${\tilde{\mathbb{Q}}^\pm}$
, and hence also
 ${\mathbb{Q}^\pm}$
-almost surely
${\mathbb{Q}^\pm}$
-almost surely
 $${\limsup_{n \to \infty} M^\pm_{S_n'} = \infty.}$$
$${\limsup_{n \to \infty} M^\pm_{S_n'} = \infty.}$$
Therefore we also get
 $${\limsup_{t \to \infty} M^\pm_t = \infty \quad \mathbb{Q}^\pm {\text{-a.s.,}}}$$
$${\limsup_{t \to \infty} M^\pm_t = \infty \quad \mathbb{Q}^\pm {\text{-a.s.,}}}$$
which proves the sought result.
If
 ${\sum_{n \geq 1} q_n n \log n = \infty}$
, then we arrive at the same conclusion by replacing
${\sum_{n \geq 1} q_n n \log n = \infty}$
, then we arrive at the same conclusion by replacing
 ${(S_n')_{n \geq 1}}$
with
${(S_n')_{n \geq 1}}$
with
 ${(S^0_n)_{n \geq 1}}$
and
${(S^0_n)_{n \geq 1}}$
and
 ${(A_n')_{n \geq 1}}$
with
${(A_n')_{n \geq 1}}$
with
 ${(A^0_n)_{n \geq 1}}$
in the above argument.
${(A^0_n)_{n \geq 1}}$
in the above argument.
From Proposition 5 we can now easily derive the required lower bound for Theorem 1.
Proposition 6. (Lower bound for Theorem 1.) Suppose that condition (10) on the offspring distribution is satisfied. Then
 $$\mathop {\lim \inf }\limits_{t \to \infty } 1/t\log |{N_t}| \ge 1/2\hat \beta _0^2 + \hat \beta \quad n{\rm{ - a}}{\rm{.s}}{\rm{.}},$$
$$\mathop {\lim \inf }\limits_{t \to \infty } 1/t\log |{N_t}| \ge 1/2\hat \beta _0^2 + \hat \beta \quad n{\rm{ - a}}{\rm{.s}}{\rm{.}},$$
Proof.
 $$|N_t| \,\mathrm{e}^{- \frac{1}{2} \hat{\beta}_0^2 t - \hat{\beta}t} \geq \sum_{u \in N_t}
\,\mathrm{e}^{- \hat{\beta}_0|X^u_t| - \frac{1}{2} \hat{\beta}_0^2t - \hat{\beta}t} = M^\pm_t.$$
$$|N_t| \,\mathrm{e}^{- \frac{1}{2} \hat{\beta}_0^2 t - \hat{\beta}t} \geq \sum_{u \in N_t}
\,\mathrm{e}^{- \hat{\beta}_0|X^u_t| - \frac{1}{2} \hat{\beta}_0^2t - \hat{\beta}t} = M^\pm_t.$$
Then
 $${\frac{\log |N_t|}{t} \geq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} + \frac{\log M^\pm_t}{t}{,}}$$
$${\frac{\log |N_t|}{t} \geq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} + \frac{\log M^\pm_t}{t}{,}}$$
and since, under condition (10),
 ${M^\pm_\infty > 0}$
${M^\pm_\infty > 0}$
 ${\mathbb{P}}$
-almost surely, it follows that
${\mathbb{P}}$
-almost surely, it follows that
 $${\liminf_{t \to \infty} {1}/{t} \log |N_t| \geq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} \quad \mathbb{P} \text{-a.s.}}$$
$${\liminf_{t \to \infty} {1}/{t} \log |N_t| \geq {1}/{2} \hat{\beta}_0^2 + \hat{\beta} \quad \mathbb{P} \text{-a.s.}}$$
In fact we have an even stronger inequality:
 $$\begin{align*}
\liminf_{t \to \infty} \,\mathrm{e}^{- \frac{1}{2} \hat{\beta}_0^2 t - \hat{\beta}t} |N_t| \geq
M^\pm_\infty > 0 \quad \mathbb{P} \text{-a.s.}\\
\end{align*}$$
$$\begin{align*}
\liminf_{t \to \infty} \,\mathrm{e}^{- \frac{1}{2} \hat{\beta}_0^2 t - \hat{\beta}t} |N_t| \geq
M^\pm_\infty > 0 \quad \mathbb{P} \text{-a.s.}\\
\end{align*}$$
Propositions 6 and 2 together prove Theorem 1.
In the rest of this subsection we would like to present some results for a purely homogeneous BBM (
 ${\beta_0 = 0}$
), which we shall make use of in the next section. We begin by stating the following result from [14, Theorem 1].
${\beta_0 = 0}$
), which we shall make use of in the next section. We begin by stating the following result from [14, Theorem 1].
Proposition 7. Consider a BBM with
 ${\beta_0 = 0}$
(only homogeneous branching present). For
${\beta_0 = 0}$
(only homogeneous branching present). For
 ${\lambda \,{\in}\, \mathbb{R}}$
, let
${\lambda \,{\in}\, \mathbb{R}}$
, let
 $$\begin{equation}
M_t^\lambda \,{:\!=}\, \sum_{u \in N_t} \,\mathrm{e}^{ \lambda X^u_t - \frac{1}{2} \lambda^2 t - \hat{\beta} t} , \quad t \ge 0{,}
\end{equation}$$
$$\begin{equation}
M_t^\lambda \,{:\!=}\, \sum_{u \in N_t} \,\mathrm{e}^{ \lambda X^u_t - \frac{1}{2} \lambda^2 t - \hat{\beta} t} , \quad t \ge 0{,}
\end{equation}$$
be the
 ${\mathbb{P}}$
-martingale of the form (24) derived through the procedure described at the beginning of this subsection by taking
${\mathbb{P}}$
-martingale of the form (24) derived through the procedure described at the beginning of this subsection by taking
 $$\begin{equation}
\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda \xi_t - \frac{1}{2} \lambda^2 t} , \quad t \ge 0.
\end{equation}$$
$$\begin{equation}
\skew1\tilde{M}^{(1)}_t = \mathrm{e}^{\lambda \xi_t - \frac{1}{2} \lambda^2 t} , \quad t \ge 0.
\end{equation}$$
(a) If
 ${\sum_{n \geq 1} p_n n \log n < \infty}$
and
${\sum_{n \geq 1} p_n n \log n < \infty}$
and
 $|\lambda | < {\rm{ (}}2\hat \beta {)^{1/2}}$
, then
$|\lambda | < {\rm{ (}}2\hat \beta {)^{1/2}}$
, then
 $$M^\lambda_\infty > 0 \quad \mathbb{P} \text{-a.s.}$$
$$M^\lambda_\infty > 0 \quad \mathbb{P} \text{-a.s.}$$
(b) If
 ${\sum_{n \geq 1} p_n n \log n < \infty}$
and
${\sum_{n \geq 1} p_n n \log n < \infty}$
and
 $|\lambda | < {\rm{ (}}2\hat \beta {)^{1/2}}$
, then
$|\lambda | < {\rm{ (}}2\hat \beta {)^{1/2}}$
, then
 $$M^\lambda_\infty = 0 \quad \mathbb{P} \text{-a.s.}$$
$$M^\lambda_\infty = 0 \quad \mathbb{P} \text{-a.s.}$$
(c) If
 ${\sum_{n \geq 1} p_n n \log n = \infty}$
, then
${\sum_{n \geq 1} p_n n \log n = \infty}$
, then
 $$M^\lambda_\infty = 0 \quad \mathbb{P} \text{-a.s.}$$
$$M^\lambda_\infty = 0 \quad \mathbb{P} \text{-a.s.}$$
The proof is essentially the same as that of Proposition 5. If we define
 ${\mathbb{Q}^\lambda}$
and
${\mathbb{Q}^\lambda}$
and
 ${\tilde{\mathbb{Q}}^\lambda}$
as probability measures associated with martingales (32) and (33), then we would see that under
${\tilde{\mathbb{Q}}^\lambda}$
as probability measures associated with martingales (32) and (33), then we would see that under
 ${\tilde{\mathbb{Q}}^\lambda}$
the
${\tilde{\mathbb{Q}}^\lambda}$
the
 ${\operatorname{spine}(t)}$
term would grow exponentially if
${\operatorname{spine}(t)}$
term would grow exponentially if
 $|\lambda | > {\rm{ (}}2\hat \beta {)^{1/2}}$
and decay exponentially if
$|\lambda | > {\rm{ (}}2\hat \beta {)^{1/2}}$
and decay exponentially if
 $|\lambda | < {\rm{ (}}2\hat \beta {)^{1/2}}$
, which together with dichotomy (29) would lead to the required result.
$|\lambda | < {\rm{ (}}2\hat \beta {)^{1/2}}$
, which together with dichotomy (29) would lead to the required result.
We shall now make use of Proposition 7 to get lower bounds on
 ${|N_t^{\lambda t}|}$
in purely homogeneous branching systems.
${|N_t^{\lambda t}|}$
in purely homogeneous branching systems.
Proposition 8. Consider a BBM with
 ${\beta_0 = 0}$
(only homogeneous branching present). If
${\beta_0 = 0}$
(only homogeneous branching present). If
 $\lambda \in (0,{\rm{ (}}2\hat \beta {)^{1/2}})$
and
$\lambda \in (0,{\rm{ (}}2\hat \beta {)^{1/2}})$
and
 ${\sum_{n \geq 1} p_n n \log n < \infty}$
, then
${\sum_{n \geq 1} p_n n \log n < \infty}$
, then
 $$\mathop {\lim \inf }\limits_{t \to \infty } 1/t\log |N_t^{\lambda t}| \ge \hat \beta - \frac{{{\lambda ^2}}}{2}\quad n{\rm{ - a}}{\rm{.s}}{\rm{.}}$$
$$\mathop {\lim \inf }\limits_{t \to \infty } 1/t\log |N_t^{\lambda t}| \ge \hat \beta - \frac{{{\lambda ^2}}}{2}\quad n{\rm{ - a}}{\rm{.s}}{\rm{.}}$$
Proof. For any choice of
 ${\delta > 0}$
such that
${\delta > 0}$
such that
 $\lambda + \delta < {\rm{ (}}2\hat \beta {)^{1/2}}$
, we have the following lower bound on
$\lambda + \delta < {\rm{ (}}2\hat \beta {)^{1/2}}$
, we have the following lower bound on
 ${|N_t^{\lambda t}|}$
:
${|N_t^{\lambda t}|}$
:
 $$\begin{align}
|N_t^{\lambda t}| &\geq \sum_{u \in N_t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}\nonumber\\&\geq \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - (\lambda + \delta)(\lambda + 2 \delta)t}
\mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}\nonumber\\&= \mathrm{e}^{\hat{\beta}t - \frac{1}{2} (\lambda + \delta)^2t - \delta(\lambda + \delta)t} \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}.
\label{eqn34}\end{align}$$
$$\begin{align}
|N_t^{\lambda t}| &\geq \sum_{u \in N_t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}\nonumber\\&\geq \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - (\lambda + \delta)(\lambda + 2 \delta)t}
\mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}\nonumber\\&= \mathrm{e}^{\hat{\beta}t - \frac{1}{2} (\lambda + \delta)^2t - \delta(\lambda + \delta)t} \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}.
\label{eqn34}\end{align}$$
We now claim that as
 ${t \to \infty}$
${t \to \infty}$
 $$\begin{equation}
\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}} \to M^{\lambda + \delta}_\infty
\quad \mathbb{P} \text{-a.s.,}
\end{equation}$$
$$\begin{equation}
\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}} \to M^{\lambda + \delta}_\infty
\quad \mathbb{P} \text{-a.s.,}
\end{equation}$$
where
 ${M^{\lambda + \delta}}$
is the same martingale as in Proposition 7. Indeed,
${M^{\lambda + \delta}}$
is the same martingale as in Proposition 7. Indeed,
 $$\begin{align}
&\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}} \nonumber\\& \qquad \leq \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}} \,\mathrm{e}^{\delta X^u_t - \delta(\lambda + 2 \delta)t} \nonumber\\
&\qquad = \mathrm{e}^{- \frac{1}{2} \delta^2 t} \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + 2\delta)X^u_t - \frac{1}{2} (\lambda + 2\delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}} \nonumber\\&\qquad \leq \mathrm{e}^{- \frac{1}{2} \delta^2 t} M^{\lambda + 2 \delta}_t \to 0 \quad \mathbb{P} \text{-a.s.}{,}
\label{eqn36}\end{align}$$
$$\begin{align}
&\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}} \nonumber\\& \qquad \leq \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}} \,\mathrm{e}^{\delta X^u_t - \delta(\lambda + 2 \delta)t} \nonumber\\
&\qquad = \mathrm{e}^{- \frac{1}{2} \delta^2 t} \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + 2\delta)X^u_t - \frac{1}{2} (\lambda + 2\delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}} \nonumber\\&\qquad \leq \mathrm{e}^{- \frac{1}{2} \delta^2 t} M^{\lambda + 2 \delta}_t \to 0 \quad \mathbb{P} \text{-a.s.}{,}
\label{eqn36}\end{align}$$
using the fact that
 ${M^{\lambda + 2 \delta}}$
converges
${M^{\lambda + 2 \delta}}$
converges
 ${\mathbb{P}}$
-almost surely to a finite limit. Similarly, we have
${\mathbb{P}}$
-almost surely to a finite limit. Similarly, we have
 $$\begin{align}
&\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \nonumber\\&\qquad \leq \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \,\mathrm{e}^{-\delta X^u_t + \delta \lambda t} \nonumber\\
&\qquad = \mathrm{e}^{- \frac{1}{2} \delta^2 t} \sum_{u \in N_t} \,\mathrm{e}^{\lambda X^u_t - \frac{1}{2} \lambda^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \nonumber\\&\qquad \leq \mathrm{e}^{- \frac{1}{2} \delta^2 t} M^{\lambda }_t \to 0 \quad \mathbb{P} \text{-a.s.}
\label{eqn37}\end{align}$$
$$\begin{align}
&\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \nonumber\\&\qquad \leq \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \,\mathrm{e}^{-\delta X^u_t + \delta \lambda t} \nonumber\\
&\qquad = \mathrm{e}^{- \frac{1}{2} \delta^2 t} \sum_{u \in N_t} \,\mathrm{e}^{\lambda X^u_t - \frac{1}{2} \lambda^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \nonumber\\&\qquad \leq \mathrm{e}^{- \frac{1}{2} \delta^2 t} M^{\lambda }_t \to 0 \quad \mathbb{P} \text{-a.s.}
\label{eqn37}\end{align}$$
Thus from (36) and (37) it follows that
 $$\begin{align*}
&\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}\\&\qquad = M^{\lambda + \delta}_t - \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}}\\&\qquad \quad\, - \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \to M^{\lambda + \delta}_\infty \quad \mathbb{P} \text{-a.s.}{,}
\end{align*}$$
$$\begin{align*}
&\sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ \lambda t \leq X^u_t \leq (\lambda + 2 \delta)t\}}\\&\qquad = M^{\lambda + \delta}_t - \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t > (\lambda + 2 \delta)t\}}\\&\qquad \quad\, - \sum_{u \in N_t} \,\mathrm{e}^{(\lambda + \delta)X^u_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t} \mathbf{1}_{\{ X^u_t < \lambda t\}} \to M^{\lambda + \delta}_\infty \quad \mathbb{P} \text{-a.s.}{,}
\end{align*}$$
proving (35). Moreover, from part (a) of Proposition 7 we know that
 ${M^{\lambda + \delta}_\infty > 0\ \mathbb{P}}$
-almost surely. Hence from (34) and (35) we get
${M^{\lambda + \delta}_\infty > 0\ \mathbb{P}}$
-almost surely. Hence from (34) and (35) we get
 $${\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \hat{\beta} - {1}/{2} (\lambda + \delta)^2 - \delta(\lambda + \delta) \quad \mathbb{P} \text{-a.s.}{,}}$$
$${\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \hat{\beta} - {1}/{2} (\lambda + \delta)^2 - \delta(\lambda + \delta) \quad \mathbb{P} \text{-a.s.}{,}}$$
which proves the proposition after letting
 ${\delta \to 0}$
.
${\delta \to 0}$
.
Proposition 9. Consider a BBM with
 ${\beta_0 = 0}$
(only homogeneous branching present). Let
${\beta_0 = 0}$
(only homogeneous branching present). Let
 $$\begin{equation}
\tilde{N}_t^{\lambda} \,{:\!=}\, \{ u \in N_{t+1} \colon X_s^u > \lambda s\ \text{for all } s \in [t, t+1] \}.
\end{equation}$$
$$\begin{equation}
\tilde{N}_t^{\lambda} \,{:\!=}\, \{ u \in N_{t+1} \colon X_s^u > \lambda s\ \text{for all } s \in [t, t+1] \}.
\end{equation}$$
If
 $\lambda > {(2\hat \beta )^{1/2}}$
and
$\lambda > {(2\hat \beta )^{1/2}}$
and
 ${\sum_{n \geq 1} p_n n \log n < \infty}$
, then
${\sum_{n \geq 1} p_n n \log n < \infty}$
, then
 $$\mathop {\lim \inf }\limits_{t \to \infty } 1/t\log {\mathbb{P}}(|\tilde N_t^\lambda | > 0) \ge \hat \beta - \frac{{{\lambda ^2}}}{2}.$$
$$\mathop {\lim \inf }\limits_{t \to \infty } 1/t\log {\mathbb{P}}(|\tilde N_t^\lambda | > 0) \ge \hat \beta - \frac{{{\lambda ^2}}}{2}.$$
In particular, it is also true that
 $$\begin{equation}
\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0 ) \geq \hat{\beta} - \frac{\lambda^2}{2}.
\end{equation}$$
$$\begin{equation}
\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0 ) \geq \hat{\beta} - \frac{\lambda^2}{2}.
\end{equation}$$
Proof. For any choice of
 ${\delta > 0}$
and
${\delta > 0}$
and
 ${K > 0}$
, consider the following events:
${K > 0}$
, consider the following events:
 $$S^{\lambda, t} \,{:\!=}\, \bigg\{
\begin{aligned}\text{{there exists} } u \in N_{t+1} \colon
& X^u_s > \lambda s\ \text{{for all} } s \in [t, t+1] \\
& X^u_s \leq K + (\lambda + 2 \delta)s\ \text{{for all} } s \in [0, t+1]
\end{aligned}
\bigg\}$$
$$S^{\lambda, t} \,{:\!=}\, \bigg\{
\begin{aligned}\text{{there exists} } u \in N_{t+1} \colon
& X^u_s > \lambda s\ \text{{for all} } s \in [t, t+1] \\
& X^u_s \leq K + (\lambda + 2 \delta)s\ \text{{for all} } s \in [0, t+1]
\end{aligned}
\bigg\}$$
and
 $${\tilde{S}^{\lambda, t} \,{:\!=}\, \{ \xi_s > \lambda s\ \text{{for all} } s \in [t, t+1],\ \xi_s \leq K + (\lambda + 2 \delta)s\ \text{{for all} } s \in [0, t+1] \}.}$$
$${\tilde{S}^{\lambda, t} \,{:\!=}\, \{ \xi_s > \lambda s\ \text{{for all} } s \in [t, t+1],\ \xi_s \leq K + (\lambda + 2 \delta)s\ \text{{for all} } s \in [0, t+1] \}.}$$
One can then see that
 ${S^{\lambda, t} \in \mathcal{F}_{t+1} \subseteq \tilde{\mathcal{F}}_{t+1}}$
,
${S^{\lambda, t} \in \mathcal{F}_{t+1} \subseteq \tilde{\mathcal{F}}_{t+1}}$
,
 ${\tilde{S}^{\lambda, t} \in \mathcal{G}_{t+1} \subseteq \tilde{F}_{t+1}}$
and that
${\tilde{S}^{\lambda, t} \in \mathcal{G}_{t+1} \subseteq \tilde{F}_{t+1}}$
and that
 $${\tilde{S}^{\lambda, t} \subseteq S^{\lambda, t} \subseteq \{|\tilde{N}_t^{\lambda}| > 0 \} \subseteq \{|N_t^{\lambda t}| > 0 \}.}$$
$${\tilde{S}^{\lambda, t} \subseteq S^{\lambda, t} \subseteq \{|\tilde{N}_t^{\lambda}| > 0 \} \subseteq \{|N_t^{\lambda t}| > 0 \}.}$$
We then have the following lower bound on
 ${\mathbb{P}(|\tilde{N}_t^{\lambda}| > 0)}$
:
${\mathbb{P}(|\tilde{N}_t^{\lambda}| > 0)}$
:
 $$ \mathbb{P} ( |\tilde{N}_t^{\lambda}| > 0 )
\geq \mathbb{P} ( S^{\lambda, t} )
= \mathbb{E} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{M^{\lambda + \delta}_{t+1}}{M^{\lambda + \delta}_{t+1}} \bigg)
= \mathbb{Q}^{\lambda + \delta} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg)
= \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg),$$
$$ \mathbb{P} ( |\tilde{N}_t^{\lambda}| > 0 )
\geq \mathbb{P} ( S^{\lambda, t} )
= \mathbb{E} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{M^{\lambda + \delta}_{t+1}}{M^{\lambda + \delta}_{t+1}} \bigg)
= \mathbb{Q}^{\lambda + \delta} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg)
= \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg),$$
where
 ${M^{\lambda + \delta}}$
,
${M^{\lambda + \delta}}$
,
 ${\mathbb{Q}^{\lambda + \delta}}$
, and
${\mathbb{Q}^{\lambda + \delta}}$
, and
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
are the same as in Proposition 7. Then
${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
are the same as in Proposition 7. Then
 $$\tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg)
\geq \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg) \geq \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \vert \tilde{\mathcal{G}}_\infty)}\bigg){,}$$
$$\tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{S^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg)
\geq \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{M^{\lambda + \delta}_{t+1}}\bigg) \geq \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \vert \tilde{\mathcal{G}}_\infty)}\bigg){,}$$
using the conditional Jensen inequality and the pull-through property of conditional expectation in the last inequality. We now recall that
 $$\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \!{\mid}\! \tilde{\mathcal{G}}_\infty)
= \operatorname{spine}(t+1) + \sum_{n=1}^{n_{t+1}} (A_n - 1)\operatorname{spine}(S_n),$$
$$\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \!{\mid}\! \tilde{\mathcal{G}}_\infty)
= \operatorname{spine}(t+1) + \sum_{n=1}^{n_{t+1}} (A_n - 1)\operatorname{spine}(S_n),$$
where
 $${\operatorname{spine}(t) = \mathrm{e}^{(\lambda + \delta)\xi_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t}.}$$
$${\operatorname{spine}(t) = \mathrm{e}^{(\lambda + \delta)\xi_t - \frac{1}{2} (\lambda + \delta)^2t - \hat{\beta}t}.}$$
On the event
 ${\tilde{S}^{\lambda, t}}$
we have that for all
${\tilde{S}^{\lambda, t}}$
we have that for all
 ${s \in [0,t+1]}$
${s \in [0,t+1]}$
 $$\begin{align*}
\operatorname{spine}(s) &\leq \exp \bigg\{(\lambda + \delta) ( K +(\lambda + 2\delta)s ) - \frac{1}{2}(\lambda + \delta)^2s - \hat{\beta}s \bigg\}\\
&= \mathrm{e}^{K(\lambda + \delta)} \exp \bigg\{\bigg( \frac{1}{2}(\lambda + \delta)^2 + \delta(\lambda + \delta) - \hat{\beta}\bigg)s \bigg\}\\&\leq C_\delta \exp \bigg\{\bigg( \frac{1}{2}(\lambda + \delta)^2 + \delta(\lambda + \delta) - \hat{\beta}\bigg)t \bigg\},
\end{align*}$$
$$\begin{align*}
\operatorname{spine}(s) &\leq \exp \bigg\{(\lambda + \delta) ( K +(\lambda + 2\delta)s ) - \frac{1}{2}(\lambda + \delta)^2s - \hat{\beta}s \bigg\}\\
&= \mathrm{e}^{K(\lambda + \delta)} \exp \bigg\{\bigg( \frac{1}{2}(\lambda + \delta)^2 + \delta(\lambda + \delta) - \hat{\beta}\bigg)s \bigg\}\\&\leq C_\delta \exp \bigg\{\bigg( \frac{1}{2}(\lambda + \delta)^2 + \delta(\lambda + \delta) - \hat{\beta}\bigg)t \bigg\},
\end{align*}$$
where
 ${C_\delta}$
is some positive constant. Also from dichotomy (29) we know that
${C_\delta}$
is some positive constant. Also from dichotomy (29) we know that
 ${A_n < \mathrm{e}^{\delta n}}$
eventually
${A_n < \mathrm{e}^{\delta n}}$
eventually
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-almost surely. Thus
${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-almost surely. Thus
 $${\sum_{n=1}^{n_{t+1}} A_n \leq \sum_{n =1}^{n_{t+1}} \,\mathrm{e}^{\delta n} + Y \leq C_\delta' \,\mathrm{e}^{\delta n_{t+1}} + Y,}$$
$${\sum_{n=1}^{n_{t+1}} A_n \leq \sum_{n =1}^{n_{t+1}} \,\mathrm{e}^{\delta n} + Y \leq C_\delta' \,\mathrm{e}^{\delta n_{t+1}} + Y,}$$
where
 ${Y = \sum_{n=1}^\infty A_n \mathbf{1}_{\{A_n > \mathrm{e}^{\delta n}\}}}$
is a
${Y = \sum_{n=1}^\infty A_n \mathbf{1}_{\{A_n > \mathrm{e}^{\delta n}\}}}$
is a
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-almost surely finite random variable independent of
${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-almost surely finite random variable independent of
 ${n_{t+1}}$
and
${n_{t+1}}$
and
 ${(\xi_s)_{0 \leq s \leq t+1}}$
and
${(\xi_s)_{0 \leq s \leq t+1}}$
and
 ${C_\delta'}$
is some positive constant. Thus
${C_\delta'}$
is some positive constant. Thus
 $$\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \!{\mid}\! \tilde{\mathcal{G}}_\infty)
\leq C_\delta \,\mathrm{e}^{( \frac{1}{2} (\lambda + \delta)^2 + \delta(\lambda + \delta) - \hat{\beta})t} ( 1 + Y + C_\delta' \,\mathrm{e}^{\delta n_{t+1}} ).$$
$$\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \!{\mid}\! \tilde{\mathcal{G}}_\infty)
\leq C_\delta \,\mathrm{e}^{( \frac{1}{2} (\lambda + \delta)^2 + \delta(\lambda + \delta) - \hat{\beta})t} ( 1 + Y + C_\delta' \,\mathrm{e}^{\delta n_{t+1}} ).$$
Then, using the fact that
 ${p{1}/{a + b} \geq p{1}/{2ab}}$
whenever a,
${p{1}/{a + b} \geq p{1}/{2ab}}$
whenever a,
 ${b \geq 1}$
, we get
${b \geq 1}$
, we get
 $$\begin{align*}
&\tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \vert \tilde{\mathcal{G}}_\infty)}\bigg)\\
&\qquad \geq \frac{1}{C_\delta}\,\mathrm{e}^{(\hat{\beta} - \frac{1}{2} (\lambda + \delta)^2 - \delta(\lambda + \delta))t}\ \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}}
\frac{1}{1 + Y + C_\delta' \,\mathrm{e}^{\delta n_{t+1}}}\bigg)\\&\qquad \geq \frac{1}{2 C_\delta C_\delta'} \,\mathrm{e}^{(\hat{\beta} - \frac{1}{2} (\lambda + \delta)^2 - \delta(\lambda + \delta))t} \tilde{\mathbb{Q}}^{\lambda + \delta} ( \tilde{S}^{\lambda, t} ) \tilde{\mathbb{Q}}^{\lambda + \delta} ( \mathrm{e}^{- \delta n_{t+1}} ) \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \frac{1}{1 + Y} \bigg).
\end{align*}$$
$$\begin{align*}
&\tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \vert \tilde{\mathcal{G}}_\infty)}\bigg)\\
&\qquad \geq \frac{1}{C_\delta}\,\mathrm{e}^{(\hat{\beta} - \frac{1}{2} (\lambda + \delta)^2 - \delta(\lambda + \delta))t}\ \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}}
\frac{1}{1 + Y + C_\delta' \,\mathrm{e}^{\delta n_{t+1}}}\bigg)\\&\qquad \geq \frac{1}{2 C_\delta C_\delta'} \,\mathrm{e}^{(\hat{\beta} - \frac{1}{2} (\lambda + \delta)^2 - \delta(\lambda + \delta))t} \tilde{\mathbb{Q}}^{\lambda + \delta} ( \tilde{S}^{\lambda, t} ) \tilde{\mathbb{Q}}^{\lambda + \delta} ( \mathrm{e}^{- \delta n_{t+1}} ) \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \frac{1}{1 + Y} \bigg).
\end{align*}$$
We note that
 $${\tilde{\mathbb{Q}}^{\lambda + \delta}\bigg(\frac{1}{1 + Y}\bigg) > 0}$$
$${\tilde{\mathbb{Q}}^{\lambda + \delta}\bigg(\frac{1}{1 + Y}\bigg) > 0}$$
since Y is
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-almost surely finite,
${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-almost surely finite,
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}(\mathrm{e}^{- \delta n_{t+1}}) = \mathrm{e}^{m \beta (t+1) (\mathrm{e}^{- \delta} - 1)}}$
since
${\tilde{\mathbb{Q}}^{\lambda + \delta}(\mathrm{e}^{- \delta n_{t+1}}) = \mathrm{e}^{m \beta (t+1) (\mathrm{e}^{- \delta} - 1)}}$
since
 ${(n_t)_{t \ge 0}}$
is a
${(n_t)_{t \ge 0}}$
is a
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-Poisson process with rate
${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
-Poisson process with rate
 ${m \beta}$
, and
${m \beta}$
, and
 ${\tilde{\mathbb{Q}}^{\lambda + \delta} ( \tilde{S}^{\lambda, t}) \to C_{K, \delta}}$
for some positive constant
${\tilde{\mathbb{Q}}^{\lambda + \delta} ( \tilde{S}^{\lambda, t}) \to C_{K, \delta}}$
for some positive constant
 ${C_{K, \delta}}$
since
${C_{K, \delta}}$
since
 ${(\xi_t)_{t \ge 0}}$
is a Brownian motion with drift
${(\xi_t)_{t \ge 0}}$
is a Brownian motion with drift
 ${\lambda + \delta}$
under
${\lambda + \delta}$
under
 ${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
. Therefore
${\tilde{\mathbb{Q}}^{\lambda + \delta}}$
. Therefore
 $$\begin{align*}
\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} ( |\tilde{N}_t^{\lambda}| > 0 )
&\geq \liminf_{t \to \infty} {1}/{t} \log \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \vert \tilde{\mathcal{G}}_\infty)}\bigg)\\& \geq \hat{\beta} - \frac{1}{2}(\lambda + \delta)^2 - \delta(\lambda + \delta) - m \beta(1 - \mathrm{e}^{-\delta}){,}
\end{align*}$$
$$\begin{align*}
\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} ( |\tilde{N}_t^{\lambda}| > 0 )
&\geq \liminf_{t \to \infty} {1}/{t} \log \tilde{\mathbb{Q}}^{\lambda + \delta} \bigg( \mathbf{1}_{\tilde{S}^{\lambda, t}} \frac{1}{\tilde{\mathbb{Q}}^{\lambda + \delta} ( M^{\lambda + \delta}_{t+1} \vert \tilde{\mathcal{G}}_\infty)}\bigg)\\& \geq \hat{\beta} - \frac{1}{2}(\lambda + \delta)^2 - \delta(\lambda + \delta) - m \beta(1 - \mathrm{e}^{-\delta}){,}
\end{align*}$$
which proves the required result after letting
 ${\delta \to 0}$
.
${\delta \to 0}$
.
Remark 2. Note that Propositions 8 and 9 (equation (39)) already provide sufficient lower bounds for Theorem 2 (equations (11) and (13) respectively) in the case
 ${\lambda \geq \hat{\beta}_0}$
, since a spatially homogeneous branching process can be embedded in a process with homogeneous and catalytic branching both present, by simply not counting any particles born due to catalytic branching.
${\lambda \geq \hat{\beta}_0}$
, since a spatially homogeneous branching process can be embedded in a process with homogeneous and catalytic branching both present, by simply not counting any particles born due to catalytic branching.
3. Remaining proofs
In this subsection we shall complete the proof of Theorem 2 by establishing lower bounds for equations (11) and (13). We shall then finish off the paper with the proof of Corollary 1.
3.1. Heuristic argument
Here we discuss the idea behind the proof in a non-rigorous way in order to help the reader understand the formal argument given in the next subsection.
Our task is to find the optimal way for a particle to reach level
 ${\lambda t}$
at some large time t.
${\lambda t}$
at some large time t.
In the case of spatially homogeneous branching (
 ${\beta_0 = 0}$
), the birth rate along the path of a particle is independent of the path, and so the optimal way would simply be to travel at speed
${\beta_0 = 0}$
), the birth rate along the path of a particle is independent of the path, and so the optimal way would simply be to travel at speed
 ${\lambda}$
all the time (there are of course finer results available, but they are irrelevant to this discussion).
${\lambda}$
all the time (there are of course finer results available, but they are irrelevant to this discussion).
However, in the presence of the catalyst at the origin, travelling at speed
 ${\lambda}$
all the time might be disadvantageous as it will discard any contribution from the catalyst. Thus one might think that a better strategy for a particle would first be to stay near the origin for some positive proportion of time in order to give birth to more particles at an accelerated rate (due to both homogeneous and catalytic branching potential), and then for the remaining time let its children travel at whatever speed is necessary in order to reach the required level.
${\lambda}$
all the time might be disadvantageous as it will discard any contribution from the catalyst. Thus one might think that a better strategy for a particle would first be to stay near the origin for some positive proportion of time in order to give birth to more particles at an accelerated rate (due to both homogeneous and catalytic branching potential), and then for the remaining time let its children travel at whatever speed is necessary in order to reach the required level.
The argument goes as follows. For a large time t we let
 $$\begin{equation*}
q \,{:\!=}\,
\begin{cases}
|N_t^{\lambda t}|& \text{if } \lambda < \lambda_{\mathrm{crit}}, \\
\mathbb{P}(|N_t^{\lambda t}| > 0)& \text{if } \lambda > \lambda_{\mathrm{crit}}{,}
\end{cases}
\end{equation*}$$
$$\begin{equation*}
q \,{:\!=}\,
\begin{cases}
|N_t^{\lambda t}|& \text{if } \lambda < \lambda_{\mathrm{crit}}, \\
\mathbb{P}(|N_t^{\lambda t}| > 0)& \text{if } \lambda > \lambda_{\mathrm{crit}}{,}
\end{cases}
\end{equation*}$$
and we want a lower bound on q.
We fix a number
 ${p \in [0,1]}$
. As we know from Theorem 1, at time pt there are
${p \in [0,1]}$
. As we know from Theorem 1, at time pt there are
 |N_{pt}| \approx \exp \bigg\{ \bigg(\frac{\hat{\beta}_0^2}{2} + \hat{\beta} \bigg)pt \bigg\}$$
|N_{pt}| \approx \exp \bigg\{ \bigg(\frac{\hat{\beta}_0^2}{2} + \hat{\beta} \bigg)pt \bigg\}$$
particles in the system and about half of them lie in the upper half-plane. Next we ignore any catalytic branching that takes place between times pt and t by assuming that every particle
 ${u \in N_{pt}^0}$
starts an independent spatially homogeneous branching process from the position
${u \in N_{pt}^0}$
starts an independent spatially homogeneous branching process from the position
 ${X^u_{pt} > 0}$
.
${X^u_{pt} > 0}$
.
We let
 $$\begin{equation*}
q(u) \,{:\!=}\,
\begin{cases}
|N_T^{\frac{\lambda}{1-p}T}(u)|& \text{if } \frac{\lambda}{1-p} < \sqrt{2 \hat{\beta}},\\\\
\mathbb{P}(|N_T^{\frac{\lambda}{1-p}T}(u)| > 0)& \text{if } \frac{\lambda}{1-p} > \sqrt{2 \hat{\beta}},
\end{cases}
\end{equation*}$$
$$\begin{equation*}
q(u) \,{:\!=}\,
\begin{cases}
|N_T^{\frac{\lambda}{1-p}T}(u)|& \text{if } \frac{\lambda}{1-p} < \sqrt{2 \hat{\beta}},\\\\
\mathbb{P}(|N_T^{\frac{\lambda}{1-p}T}(u)| > 0)& \text{if } \frac{\lambda}{1-p} > \sqrt{2 \hat{\beta}},
\end{cases}
\end{equation*}$$
where
 ${N_T^{(\frac{\lambda}({1-p}))T}(u)}$
is the set of particles which lie to the right of
${N_T^{(\frac{\lambda}({1-p}))T}(u)}$
is the set of particles which lie to the right of
 ${(\frac{\lambda}({1-p}))T}$
at time T of the spatially homogeneous process initiated by u in the time–space frame of this process and where
${(\frac{\lambda}({1-p}))T}$
at time T of the spatially homogeneous process initiated by u in the time–space frame of this process and where
 ${T = (1-p)t}$
. Then, from Propositions 8 and 9, we know that
${T = (1-p)t}$
. Then, from Propositions 8 and 9, we know that
 $$q(u) \ge \exp \{ \hat \beta T - 1/2{(\frac{\lambda }{{1 - p}})^2}T\} = \exp \{ \hat \beta (1 - p)t - \frac{{{\lambda ^2}}}{{2(1 - p)}}t\} .$$
$$q(u) \ge \exp \{ \hat \beta T - 1/2{(\frac{\lambda }{{1 - p}})^2}T\} = \exp \{ \hat \beta (1 - p)t - \frac{{{\lambda ^2}}}{{2(1 - p)}}t\} .$$
Then, since every particle in
 ${N_T^{\frac{\lambda}{1-p}T}(u)}$
for every
${N_T^{\frac{\lambda}{1-p}T}(u)}$
for every
 ${u \in N_{pt}^0}$
also belongs to
${u \in N_{pt}^0}$
also belongs to
 ${N_t^{\lambda t}}$
, we can estimate
${N_t^{\lambda t}}$
, we can estimate
 $$\begin{equation}
q \ge \sum_{u \in N_{pt}^0} q(u)
\approx |N_{pt}^0| \exp \bigg\{ \hat{\beta}(1-p)t - \frac{\lambda^2}{2(1-p)} t \bigg\}
\approx \exp \bigg\{ \bigg( \hat{\beta} + \frac{\hat{\beta}_0^2}{2}p - \frac{\lambda^2}{2(1-p)}
\bigg) t \bigg\}.
\end{equation}$$
$$\begin{equation}
q \ge \sum_{u \in N_{pt}^0} q(u)
\approx |N_{pt}^0| \exp \bigg\{ \hat{\beta}(1-p)t - \frac{\lambda^2}{2(1-p)} t \bigg\}
\approx \exp \bigg\{ \bigg( \hat{\beta} + \frac{\hat{\beta}_0^2}{2}p - \frac{\lambda^2}{2(1-p)}
\bigg) t \bigg\}.
\end{equation}$$
The value of p which maximizes this expression is
 $$\begin{equation}
p^{\ast} \,{:\!=}\,
\begin{cases}
1 - \frac{\lambda}{\hat{\beta}_0}& \text{if } \lambda \leq \hat{\beta}_0,\\
0& \text{if } \lambda \geq \hat{\beta}_0.
\end{cases}
\end{equation}$$
$$\begin{equation}
p^{\ast} \,{:\!=}\,
\begin{cases}
1 - \frac{\lambda}{\hat{\beta}_0}& \text{if } \lambda \leq \hat{\beta}_0,\\
0& \text{if } \lambda \geq \hat{\beta}_0.
\end{cases}
\end{equation}$$
Substituting this value of p into (40), we get
 $$\begin{array}{l}
q \ge \left\{ {\begin{array}{*{20}{l}}
{\exp \{ (\hat \beta + \frac{{\hat \beta _0^2}}{2} - {{\hat \beta }_0}\lambda )t\} }&{{\rm{if }}\lambda \le {{\hat \beta }_0}}&{}\\
{\exp \{ (\hat \beta - \frac{{{\lambda ^2}}}{2})t\} }&{{\rm{if }}\lambda \ge {{\hat \beta }_0}}
\end{array}} \right.\\
\,\,\,\,\, = \exp \{ {\Delta _\lambda }t\} ,
\end{array}$$
$$\begin{array}{l}
q \ge \left\{ {\begin{array}{*{20}{l}}
{\exp \{ (\hat \beta + \frac{{\hat \beta _0^2}}{2} - {{\hat \beta }_0}\lambda )t\} }&{{\rm{if }}\lambda \le {{\hat \beta }_0}}&{}\\
{\exp \{ (\hat \beta - \frac{{{\lambda ^2}}}{2})t\} }&{{\rm{if }}\lambda \ge {{\hat \beta }_0}}
\end{array}} \right.\\
\,\,\,\,\, = \exp \{ {\Delta _\lambda }t\} ,
\end{array}$$
which gives the lower bound on q that we want.
Note that if
 ${\lambda}$
is too large (
${\lambda}$
is too large (
 ${\lambda \geq \hat{\beta}_0}$
) then
${\lambda \geq \hat{\beta}_0}$
) then
 ${p^\ast = 0}$
, and so the best strategy for a particle to reach level
${p^\ast = 0}$
, and so the best strategy for a particle to reach level
 ${\lambda t}$
at time t would indeed be to travel at speed
${\lambda t}$
at time t would indeed be to travel at speed
 ${\lambda}$
, always being driven by a homogeneous branching potential with negligible contribution from catalytic branching. This is consistent with Remark 2 made earlier.
${\lambda}$
, always being driven by a homogeneous branching potential with negligible contribution from catalytic branching. This is consistent with Remark 2 made earlier.
3.2. Lower bounds for (11) and (13)
Before we present the main body of the proof, let us give a couple of preliminary results. The first is a very crude estimate of the number of particles which lie approximately in the upper half-plane at a time t.
Proposition 10. Assume that condition (10) on the offspring distribution is satisfied. Then
 ${\mathbb{P}}$
-almost surely, for any
${\mathbb{P}}$
-almost surely, for any
 ${\delta > 0}$
, there exists a finite time
${\delta > 0}$
, there exists a finite time
 ${T_\delta}$
such that, for all
${T_\delta}$
such that, for all
 ${t \geq T_\delta}$
,
${t \geq T_\delta}$
,
 $$\begin{equation*} \label{eq_positive_pop}
|N^{- \delta t}_t| \geq \exp \bigg\{\bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - c_\delta\bigg)t \bigg\},
\end{equation*}$$
$$\begin{equation*} \label{eq_positive_pop}
|N^{- \delta t}_t| \geq \exp \bigg\{\bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - c_\delta\bigg)t \bigg\},
\end{equation*}$$
where
 ${c_\delta}$
is some positive constant with the property that
${c_\delta}$
is some positive constant with the property that
 ${c_\delta \to 0}$
as
${c_\delta \to 0}$
as
 ${\delta \to 0}$
.
${\delta \to 0}$
.
Proof. Let us observe that
 ${\Delta_\delta \to \hat{\beta} + {\hat{\beta}_0^2}/{2}}$
as
${\Delta_\delta \to \hat{\beta} + {\hat{\beta}_0^2}/{2}}$
as
 ${\delta \to 0}$
. So we may write
${\delta \to 0}$
. So we may write
 ${\Delta_\delta = \hat{\beta} + {\hat{\beta}_0^2}/{2} - c_\delta'}$
for some
${\Delta_\delta = \hat{\beta} + {\hat{\beta}_0^2}/{2} - c_\delta'}$
for some
 ${c_\delta' > 0}$
such that
${c_\delta' > 0}$
such that
 ${c_\delta' \to 0}$
as
${c_\delta' \to 0}$
as
 ${\delta \to 0}$
.
${\delta \to 0}$
.
From Theorem 1 (or Proposition 6 we know that
 ${\mathbb{P}}$
-almost surely, for any
${\mathbb{P}}$
-almost surely, for any
 ${\delta > 0}$
, there exists a finite time
${\delta > 0}$
, there exists a finite time
 ${T_\delta'}$
such that, for all
${T_\delta'}$
such that, for all
 ${t \geq T_\delta'}$
,
${t \geq T_\delta'}$
,
 $$\begin{equation}
|N_t| \geq \exp \bigg\{\bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - \frac{1}{4}c_\delta' \bigg)t \bigg\}.
\end{equation}$$
$$\begin{equation}
|N_t| \geq \exp \bigg\{\bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - \frac{1}{4}c_\delta' \bigg)t \bigg\}.
\end{equation}$$
We also know from Proposition 4 (equation (20)) that
 ${\mathbb{P}}$
-almost surely, for any
${\mathbb{P}}$
-almost surely, for any
 ${\delta > 0}$
, there exists a finite time
${\delta > 0}$
, there exists a finite time
 ${T_\delta''}$
such that, for all
${T_\delta''}$
such that, for all
 ${t \geq T_\delta''}$
,
${t \geq T_\delta''}$
,
 $$|N^{\delta t}_t| \leq \exp \bigg\{\bigg(\Delta_\delta + {1}/{2} c_\delta' \bigg)t \bigg\} =
\exp \bigg\{ \bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - \frac{1}{2}c_\delta' \bigg)t \bigg\}.$$
$$|N^{\delta t}_t| \leq \exp \bigg\{\bigg(\Delta_\delta + {1}/{2} c_\delta' \bigg)t \bigg\} =
\exp \bigg\{ \bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - \frac{1}{2}c_\delta' \bigg)t \bigg\}.$$
Thus, by symmetry it is also true that
 ${\mathbb{P}}$
-almost surely, for any
${\mathbb{P}}$
-almost surely, for any
 ${\delta > 0}$
, there exists a finite time
${\delta > 0}$
, there exists a finite time
 ${T_\delta'''}$
such that, for all
${T_\delta'''}$
such that, for all
 ${t \geq T_\delta'''}$
,
${t \geq T_\delta'''}$
,
 $$\begin{equation}
|N_t| - |N^{- \delta t}_t| \leq \exp \bigg\{ \bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - \frac{1}{2}c_\delta' \bigg)t \bigg\}.
\end{equation}$$
$$\begin{equation}
|N_t| - |N^{- \delta t}_t| \leq \exp \bigg\{ \bigg(\hat{\beta} + \frac{\hat{\beta}_0^2}{2} - \frac{1}{2}c_\delta' \bigg)t \bigg\}.
\end{equation}$$
Subtracting (43) from (42) yields the result.
The next result is basically a version of Chebyshev’s inequality.
Proposition 11. Let N be a random variable supported on
 ${\mathbb{N}}$
and
${\mathbb{N}}$
and
 ${(S_k)_{k \geq 1}}$
a sequence of events independent of each other conditional on N. If, for some
${(S_k)_{k \geq 1}}$
a sequence of events independent of each other conditional on N. If, for some
 ${r \in (0, 1)}$
, it is true that
${r \in (0, 1)}$
, it is true that
 ${\mathbb{P}(S_k\!{\mid}\!N) \geq r}$
${\mathbb{P}(S_k\!{\mid}\!N) \geq r}$
 ${\mathbb{P}}$
-a.s. for all
${\mathbb{P}}$
-a.s. for all
 ${k \geq 1}$
, then
${k \geq 1}$
, then
 $$\mathbb{P} \bigg( \sum_{k = 1}^N \mathbf{1}_{S_k} \leq \frac{r}{2}N \biggm\vert N \bigg)
\leq \frac{4}{rN} \quad \mathbb{P} {\text{-a.s.}}$$
$$\mathbb{P} \bigg( \sum_{k = 1}^N \mathbf{1}_{S_k} \leq \frac{r}{2}N \biggm\vert N \bigg)
\leq \frac{4}{rN} \quad \mathbb{P} {\text{-a.s.}}$$
Sharper inequalities are of course available but are not needed here.
Proof of Proposition 11. Let us assume for simplicity that N is deterministic. Then
 $$\begin{align*}
\mathbb{P} \bigg( \sum_{k = 1}^N \mathbf{1}_{S_k} \leq \frac{r}{2}N \bigg)
&\leq \mathbb{P} \bigg( \sum_{k = 1}^N ( \mathbf{1}_{S_k} - \mathbb{P}(S_k) ) \leq - \frac{1}{2}
\sum_{k = 1}^N \mathbb{P}(S_k) \bigg) \\
&\leq \mathbb{P} \bigg( \biggm\vert \sum_{k = 1}^N ( \mathbf{1}_{S_k} - \mathbb{P}(S_k) ) \bigg\vert \geq {1}/{2} \sum_{k = 1}^N \mathbb{P}(S_k) \bigg)\\
&\leq \frac{\sum_{k=1}^N \ var(\mathbf{1}_{S_k})}{\frac{1}{4} \bigg(\sum_{k = 1}^N \mathbb{P}(S_k) \bigg)^2} \\& \leq \frac{4}{rN}
\end{align*}$$
$$\begin{align*}
\mathbb{P} \bigg( \sum_{k = 1}^N \mathbf{1}_{S_k} \leq \frac{r}{2}N \bigg)
&\leq \mathbb{P} \bigg( \sum_{k = 1}^N ( \mathbf{1}_{S_k} - \mathbb{P}(S_k) ) \leq - \frac{1}{2}
\sum_{k = 1}^N \mathbb{P}(S_k) \bigg) \\
&\leq \mathbb{P} \bigg( \biggm\vert \sum_{k = 1}^N ( \mathbf{1}_{S_k} - \mathbb{P}(S_k) ) \bigg\vert \geq {1}/{2} \sum_{k = 1}^N \mathbb{P}(S_k) \bigg)\\
&\leq \frac{\sum_{k=1}^N \ var(\mathbf{1}_{S_k})}{\frac{1}{4} \bigg(\sum_{k = 1}^N \mathbb{P}(S_k) \bigg)^2} \\& \leq \frac{4}{rN}
\end{align*}$$
using the Markov inequality and the fact that
 ${var\,\,(\mathbf{1}_{S_k}) = \mathbb{P}(S_k) - \mathbb{P}(S_k)^2 \leq \mathbb{P}(S_k)}$
. The same argument will then work for N random if we replace
${var\,\,(\mathbf{1}_{S_k}) = \mathbb{P}(S_k) - \mathbb{P}(S_k)^2 \leq \mathbb{P}(S_k)}$
. The same argument will then work for N random if we replace
 ${\mathbb{P}({\cdot})}$
with
${\mathbb{P}({\cdot})}$
with
 ${\mathbb{P}({\cdot} \!{\mid}\! N)}$
and
${\mathbb{P}({\cdot} \!{\mid}\! N)}$
and
 ${var({\cdot})}$
with
${var({\cdot})}$
with
 ${var({\cdot} \!{\mid}\! N)}$
.
${var({\cdot} \!{\mid}\! N)}$
.
Proposition 12. (Lower bounds for Theorem 2.) Suppose that condition (10) on the offspring distribution is satisfied.
If
 ${\lambda < \lambda_{\mathrm{crit}}}$
${\lambda < \lambda_{\mathrm{crit}}}$
 ${(\Delta_\lambda > 0)}$
, then
${(\Delta_\lambda > 0)}$
, then
 $$\begin{equation}
\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \Delta_\lambda
\quad \mathbb{P} \text{-a.s.}
\end{equation}$$
$$\begin{equation}
\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \Delta_\lambda
\quad \mathbb{P} \text{-a.s.}
\end{equation}$$
If
 ${\lambda > \lambda_{\mathrm{crit}}}$
${\lambda > \lambda_{\mathrm{crit}}}$
 ${(\Delta_\lambda < 0)}$
, then
${(\Delta_\lambda < 0)}$
, then
 $$\begin{equation}
\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0 ) \geq \Delta_\lambda.
\end{equation}$$
$$\begin{equation}
\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0 ) \geq \Delta_\lambda.
\end{equation}$$
Proof. We let
 ${p = p^\ast}$
be the same as in (41). We note that
${p = p^\ast}$
be the same as in (41). We note that
 ${p = 0}$
if and only if
${p = 0}$
if and only if
 ${\lambda \geq \hat{\beta}_0 }$
, while if
${\lambda \geq \hat{\beta}_0 }$
, while if
 ${\lambda \geq \hat{\beta}_0}$
then
${\lambda \geq \hat{\beta}_0}$
then
 ${\Delta_\lambda = \hat{\beta} - \frac{1}{2} \lambda^2}$
, so that (44) and (45) follow from Propositions 8 and 9 (equation (39)) by simply not counting any particles born due to catalytic branching. Thus, for the rest of the proof we shall assume that
${\Delta_\lambda = \hat{\beta} - \frac{1}{2} \lambda^2}$
, so that (44) and (45) follow from Propositions 8 and 9 (equation (39)) by simply not counting any particles born due to catalytic branching. Thus, for the rest of the proof we shall assume that
 ${\lambda < \hat{\beta}_0}$
, so that
${\lambda < \hat{\beta}_0}$
, so that
 ${p = 1 - {\lambda}/{\hat{\beta}_0} > 0}$
and
${p = 1 - {\lambda}/{\hat{\beta}_0} > 0}$
and
 ${\Delta_\lambda = \hat{\beta} + {\hat{\beta}_0^2}/{2} - \hat{\beta}_0 \lambda}$
.
${\Delta_\lambda = \hat{\beta} + {\hat{\beta}_0^2}/{2} - \hat{\beta}_0 \lambda}$
.
We then choose some
 ${\delta > 0}$
and define
${\delta > 0}$
and define
 $$\hat \lambda {\mkern 1mu} : = {\mkern 1mu} \frac{{\lambda + \delta }}{\lambda }{{\hat \beta }_0}.$$
$$\hat \lambda {\mkern 1mu} : = {\mkern 1mu} \frac{{\lambda + \delta }}{\lambda }{{\hat \beta }_0}.$$
We also define
 $$f(\delta) \,{:\!=}\, \bigg( \hat{\beta} - \frac{\hat{\lambda}^2}{2} - \delta \bigg),\quad g(\delta) \,{:\!=}\, \bigg( \hat{\beta} + \frac{\hat{\beta}_0^2}{2} - c_\delta \bigg),$$
$$f(\delta) \,{:\!=}\, \bigg( \hat{\beta} - \frac{\hat{\lambda}^2}{2} - \delta \bigg),\quad g(\delta) \,{:\!=}\, \bigg( \hat{\beta} + \frac{\hat{\beta}_0^2}{2} - c_\delta \bigg),$$
where
 ${c_\delta}$
is the same as in Proposition 10. We let
${c_\delta}$
is the same as in Proposition 10. We let
 ${h(\delta)}$
be such that
${h(\delta)}$
be such that
 $$\begin{align*}
(1-p)f(\delta) + pg(\delta) &= \bigg( \hat{\beta} - \frac{\hat{\lambda}^2}{2} - \delta \bigg)(1-p) +
\bigg( \hat{\beta} + \frac{\hat{\beta}_0^2}{2} - c_\delta \bigg)p\\
&= \hat{\beta} + \frac{\hat{\beta}_0^2}{2} \bigg( 1 - \frac{\lambda}{\hat{\beta}_0} \bigg) - c_\delta p
- {1}/{2} \bigg( \frac{\lambda + \delta}{\lambda} \hat{\beta}_0 \bigg)^2 \frac{\lambda}{\hat{\beta}_0} - \delta(1-p)\\&= \Delta_\lambda - h(\delta).
\end{align*}$$
$$\begin{align*}
(1-p)f(\delta) + pg(\delta) &= \bigg( \hat{\beta} - \frac{\hat{\lambda}^2}{2} - \delta \bigg)(1-p) +
\bigg( \hat{\beta} + \frac{\hat{\beta}_0^2}{2} - c_\delta \bigg)p\\
&= \hat{\beta} + \frac{\hat{\beta}_0^2}{2} \bigg( 1 - \frac{\lambda}{\hat{\beta}_0} \bigg) - c_\delta p
- {1}/{2} \bigg( \frac{\lambda + \delta}{\lambda} \hat{\beta}_0 \bigg)^2 \frac{\lambda}{\hat{\beta}_0} - \delta(1-p)\\&= \Delta_\lambda - h(\delta).
\end{align*}$$
Note that
 ${h(\delta)>0}$
and
${h(\delta)>0}$
and
 ${h(\delta) \to 0}$
as
${h(\delta) \to 0}$
as
 ${\delta \to 0}$
.
${\delta \to 0}$
.
For
 ${t>0}$
we define events
${t>0}$
we define events
 $${\mathcal{A}_t}{\mkern 1mu} : = {\mkern 1mu} \{ |N_{pt}^{ - \delta pt}| \ge {{\rm{e}}^{g(\delta )pt}}\} .$$
$${\mathcal{A}_t}{\mkern 1mu} : = {\mkern 1mu} \{ |N_{pt}^{ - \delta pt}| \ge {{\rm{e}}^{g(\delta )pt}}\} .$$
From Proposition 10 we know that
 ${\mathbb{P}(\mathcal{A}_n) \text{ {eventually}} = 1}$
, so that in particular
${\mathbb{P}(\mathcal{A}_n) \text{ {eventually}} = 1}$
, so that in particular
 ${\mathbb{P}(\mathcal{A}_t) \to 1}$
as
${\mathbb{P}(\mathcal{A}_t) \to 1}$
as
 ${t \to \infty}$
.
${t \to \infty}$
.
Finally, for every particle
 ${u \in N_{pt}}$
, let us consider the subtree initiated by u at time pt. In the time–space frame of this subtree, we define
${u \in N_{pt}}$
, let us consider the subtree initiated by u at time pt. In the time–space frame of this subtree, we define
 ${N_s(u)}$
,
${N_s(u)}$
,
 ${s \geq 0}$
to be the set of particles in the subtree at time s and
${s \geq 0}$
to be the set of particles in the subtree at time s and
 ${Y^v_s}$
the positions of particles
${Y^v_s}$
the positions of particles
 ${v \in N_s(u)}$
at time s. Moreover, by analogy with (2) and (38), we define
${v \in N_s(u)}$
at time s. Moreover, by analogy with (2) and (38), we define
 $$N_s^x(u) \,{:\!=}\, \{ v \in N_s(u) \colon Y_s^v > x\}$$
$$N_s^x(u) \,{:\!=}\, \{ v \in N_s(u) \colon Y_s^v > x\}$$
and
 $$\begin{align*}
\tilde{N}_s^l(u) \,{:\!=}\, \{ v \in N_{s+1}(u) \colon Y_r^v > lr\ \text{for all } r \in [s, s+1]\}.\\
\end{align*}$$
$$\begin{align*}
\tilde{N}_s^l(u) \,{:\!=}\, \{ v \in N_{s+1}(u) \colon Y_r^v > lr\ \text{for all } r \in [s, s+1]\}.\\
\end{align*}$$
Proof of (44), the lower bound for (11). Assume that
 ${\lambda < \lambda_{\mathrm{crit}}}$
so that
${\lambda < \lambda_{\mathrm{crit}}}$
so that
 ${\Delta_\lambda > 0}$
. There are two cases to consider, which require slightly different treatment.
${\Delta_\lambda > 0}$
. There are two cases to consider, which require slightly different treatment.
Case 1:
 ${\hat{\beta} > \frac{1}{2} \hat{\beta}_0^2}$
(equivalently,
${\hat{\beta} > \frac{1}{2} \hat{\beta}_0^2}$
(equivalently,
 ${{\hat \beta }_0} < {\rm{ (}}2\hat \beta {)^{1/2}}$
). We choose
${{\hat \beta }_0} < {\rm{ (}}2\hat \beta {)^{1/2}}$
). We choose
 ${\delta > 0}$
to be sufficiently small that
${\delta > 0}$
to be sufficiently small that
 $\hat \lambda < {\rm{ }}{(2\hat \beta )^{1/2}}$
, and for
$\hat \lambda < {\rm{ }}{(2\hat \beta )^{1/2}}$
, and for
 ${n \geq 1}$
consider events
${n \geq 1}$
consider events
 $${\mathcal{B}_n \,{:\!=}\, \bigg\{ \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \mathcal{B}_n(u)} < \frac{1}{4} |N_{pn}^{- \delta pn}| \bigg\},}$$
$${\mathcal{B}_n \,{:\!=}\, \bigg\{ \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \mathcal{B}_n(u)} < \frac{1}{4} |N_{pn}^{- \delta pn}| \bigg\},}$$
where, for every
 ${u \in N_{pn}^{- \delta pn}}$
,
${u \in N_{pn}^{- \delta pn}}$
,
 $${\mathcal{B}_n(u) = \{ |N_s^{\hat{\lambda}s}(u)| \geq \mathrm{e}^{ f(\delta) s}\ \text{for all } s \in [(1-p)n, (1-p)n + 1] \}.}$$
$${\mathcal{B}_n(u) = \{ |N_s^{\hat{\lambda}s}(u)| \geq \mathrm{e}^{ f(\delta) s}\ \text{for all } s \in [(1-p)n, (1-p)n + 1] \}.}$$
We know that, conditional on
 ${\mathcal{F}_{pn}}$
, events
${\mathcal{F}_{pn}}$
, events
 ${\mathcal{B}_n(u)}$
are independent, since all the subtrees initiated by particles
${\mathcal{B}_n(u)}$
are independent, since all the subtrees initiated by particles
 ${u \in N_{pn}}$
are independent copies of the original branching process starting from positions
${u \in N_{pn}}$
are independent copies of the original branching process starting from positions
 ${X^u_{pn}}$
.
${X^u_{pn}}$
.
Moreover, if we ignore all the catalytic branching taking place in the subtrees initiated by particles
 ${u \in N_{pn}^{- \delta p n}}$
, then we can get from Proposition 8 that there exists some deterministic
${u \in N_{pn}^{- \delta p n}}$
, then we can get from Proposition 8 that there exists some deterministic
 ${n_0}$
such that for all
${n_0}$
such that for all
 ${n \geq n_0}$
,
${n \geq n_0}$
,
 ${\mathbb{P}(\mathcal{B}_n(u) \!{\mid}\! \mathcal{F}_{pn}) \geq {1}/{2} }$
. Hence, by Proposition 11,
${\mathbb{P}(\mathcal{B}_n(u) \!{\mid}\! \mathcal{F}_{pn}) \geq {1}/{2} }$
. Hence, by Proposition 11,
 $$\mathbb{P}(\mathcal{B}_n \!{\mid}\! \mathcal{F}_{pn}) \leq \frac{8}{|N_{pn}^{- \delta pn}|}
\quad\text{$\mathbb{P}$-a.s.}$$
$$\mathbb{P}(\mathcal{B}_n \!{\mid}\! \mathcal{F}_{pn}) \leq \frac{8}{|N_{pn}^{- \delta pn}|}
\quad\text{$\mathbb{P}$-a.s.}$$
for all
 ${n \geq n_0}$
. Then, for all
${n \geq n_0}$
. Then, for all
 ${n \geq n_0}$
, we obtain
${n \geq n_0}$
, we obtain
 $$\begin{align*}
\mathbb{P} (\mathcal{A}_n \cap \mathcal{B}_n ) = \mathbb{E} ( \mathbb{E} ( \mathbf{1}_{\mathcal{A}_n} \mathbf{1}_{\mathcal{B}_n} \!{\mid}\! \mathcal{F}_{pn} ) )
\leq \mathbb{E} \bigg( \mathbf{1}_{\mathcal{A}_n} \frac{8}{|N_{pn}^{- \delta pn}|} \bigg)
\leq 8 \,\mathrm{e}^{- g(\delta) pn}{,}
\end{align*}$$
$$\begin{align*}
\mathbb{P} (\mathcal{A}_n \cap \mathcal{B}_n ) = \mathbb{E} ( \mathbb{E} ( \mathbf{1}_{\mathcal{A}_n} \mathbf{1}_{\mathcal{B}_n} \!{\mid}\! \mathcal{F}_{pn} ) )
\leq \mathbb{E} \bigg( \mathbf{1}_{\mathcal{A}_n} \frac{8}{|N_{pn}^{- \delta pn}|} \bigg)
\leq 8 \,\mathrm{e}^{- g(\delta) pn}{,}
\end{align*}$$
which decays exponentially fast in n (for
 ${\delta}$
sufficiently small). Therefore
${\delta}$
sufficiently small). Therefore
 ${\mathbb{P}(\mathcal{A}_n \cap \mathcal{B}_n \text{ i.o.}) = 0}$
. Then, since
${\mathbb{P}(\mathcal{A}_n \cap \mathcal{B}_n \text{ i.o.}) = 0}$
. Then, since
 ${\mathbb{P}(\mathcal{A}_n \text{ {eventually}}) = 1}$
, it follows that
${\mathbb{P}(\mathcal{A}_n \text{ {eventually}}) = 1}$
, it follows that
 ${\mathbb{P}(\mathcal{A}_n \cap \mathcal{B}_n^c \text{ {eventually}}) = 1}$
. So
${\mathbb{P}(\mathcal{A}_n \cap \mathcal{B}_n^c \text{ {eventually}}) = 1}$
. So
 ${\mathbb{P}}$
-a.s., for all n large enough,
${\mathbb{P}}$
-a.s., for all n large enough,
 $$\sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \mathcal{B}_n(u)} \geq \frac{1}{4} |N_{pn}^{- \delta pn}|
\geq \frac{1}{4} \mathrm{e}^{g(\delta)pn}.$$
$$\sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \mathcal{B}_n(u)} \geq \frac{1}{4} |N_{pn}^{- \delta pn}|
\geq \frac{1}{4} \mathrm{e}^{g(\delta)pn}.$$
Then, noting that, for all
 ${t \in [n, n+1]}$
,
${t \in [n, n+1]}$
,
 $$|N_t^{\lambda t}| \geq \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \mathcal{B}_n(u)} \,\mathrm{e}^{f(\delta)(1-p)n}{,}$$
$$|N_t^{\lambda t}| \geq \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \mathcal{B}_n(u)} \,\mathrm{e}^{f(\delta)(1-p)n}{,}$$
we get that
 ${\mathbb{P}}$
-a.s., for all t sufficiently large,
${\mathbb{P}}$
-a.s., for all t sufficiently large,
 $$|N_t^{\lambda t}| \geq K \,\mathrm{e}^{g(\delta)pt + f(\delta)(1-p)t},$$
$$|N_t^{\lambda t}| \geq K \,\mathrm{e}^{g(\delta)pt + f(\delta)(1-p)t},$$
where K is some positive constant. Hence
 $${\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \Delta_\lambda - h(\delta) \quad \mathbb{P} \text{-a.s.,}}$$
$${\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \Delta_\lambda - h(\delta) \quad \mathbb{P} \text{-a.s.,}}$$
which yields the required value after letting
 ${\delta \to 0}$
.
${\delta \to 0}$
.
Case 2:
 ${\hat{\beta} \leq \frac{1}{2} \hat{\beta}_0^2}$
(equivalently,
${\hat{\beta} \leq \frac{1}{2} \hat{\beta}_0^2}$
(equivalently,
 ${{\hat \beta }_0} \ge {(2\hat \beta )^{1/2}}$
). For
${{\hat \beta }_0} \ge {(2\hat \beta )^{1/2}}$
). For
 ${n \geq 1}$
we consider events
${n \geq 1}$
we consider events
 $${\mathcal{C}_n \,{:\!=}\, \bigg\{ \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{\{ |\tilde{N}_{(1-p)n}^{\hat{\lambda}}(u)| > 0\}} < {1}/{2} \mathrm{e}^{f(\delta)(1-p)n} |N_{pn}^{- \delta pn}|\bigg\}}$$
$${\mathcal{C}_n \,{:\!=}\, \bigg\{ \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{\{ |\tilde{N}_{(1-p)n}^{\hat{\lambda}}(u)| > 0\}} < {1}/{2} \mathrm{e}^{f(\delta)(1-p)n} |N_{pn}^{- \delta pn}|\bigg\}}$$
(note that
 $\hat \lambda > {\rm{ }}{(2\hat \beta )^{1/2}}$
). We know that, conditional on
$\hat \lambda > {\rm{ }}{(2\hat \beta )^{1/2}}$
). We know that, conditional on
 ${\mathcal{F}_{pn}}$
, events
${\mathcal{F}_{pn}}$
, events
 ${\{|\tilde{N}^{\hat{\lambda}}_{(1-p)n}(u)|>0\}}$
are independent. Moreover, if we ignore all the catalytic branching taking place in the subtrees initiated by particles
${\{|\tilde{N}^{\hat{\lambda}}_{(1-p)n}(u)|>0\}}$
are independent. Moreover, if we ignore all the catalytic branching taking place in the subtrees initiated by particles
 ${u \in N_{pn}^{- \delta p n}}$
, then we can get from Proposition 9 that there exists some deterministic
${u \in N_{pn}^{- \delta p n}}$
, then we can get from Proposition 9 that there exists some deterministic
 ${n_0}$
such that, for all
${n_0}$
such that, for all
 ${n \geq n_0}$
,
${n \geq n_0}$
,
 ${\mathbb{P}(|\tilde{N}^{\hat{\lambda}}_{(1-p)n}(u)|>0 \!{\mid}\! \mathcal{F}_{pn}) \geq \mathrm{e}^{f(\delta)(1-p)n}}$
. Hence, by Proposition 11, for all
${\mathbb{P}(|\tilde{N}^{\hat{\lambda}}_{(1-p)n}(u)|>0 \!{\mid}\! \mathcal{F}_{pn}) \geq \mathrm{e}^{f(\delta)(1-p)n}}$
. Hence, by Proposition 11, for all
 ${n \geq n_0}$
,
${n \geq n_0}$
,
 $${\mathbb{P} (\mathcal{C}_n \!{\mid}\! \mathcal{F}_{pn} ) \leq \frac{4}{|N_{pn}^{- \delta pn}|} \,\mathrm{e}^{-f(\delta)(1-p)n} \quad \mathbb{P} \text{-a.s.}}$$
$${\mathbb{P} (\mathcal{C}_n \!{\mid}\! \mathcal{F}_{pn} ) \leq \frac{4}{|N_{pn}^{- \delta pn}|} \,\mathrm{e}^{-f(\delta)(1-p)n} \quad \mathbb{P} \text{-a.s.}}$$
Then, for all
 ${n \geq n_0}$
, we obtain
${n \geq n_0}$
, we obtain
 $$\begin{align*}
\mathbb{P} (\mathcal{A}_n \cap \mathcal{C}_n) &= \mathbb{E} ( \mathbb{E} ( \mathbf{1}_{\mathcal{A}_n} \mathbf{1}_{\mathcal{C}_n} \!{\mid}\! \mathcal{F}_{pn} ) ) \\
& \leq \mathbb{E} \bigg( \mathbf{1}_{\mathcal{A}_n} \frac{4}{|N_{pn}^{- \delta pn}|} \,\mathrm{e}^{-f(\delta)(1-p)n}\bigg)\\
&\leq 4\,\mathrm{e}^{-g(\delta)p -f(\delta)(1-p)}\\
&= 4 \,\mathrm{e}^{ -(\Delta_\lambda - h(\delta) ) n},
\end{align*}$$
$$\begin{align*}
\mathbb{P} (\mathcal{A}_n \cap \mathcal{C}_n) &= \mathbb{E} ( \mathbb{E} ( \mathbf{1}_{\mathcal{A}_n} \mathbf{1}_{\mathcal{C}_n} \!{\mid}\! \mathcal{F}_{pn} ) ) \\
& \leq \mathbb{E} \bigg( \mathbf{1}_{\mathcal{A}_n} \frac{4}{|N_{pn}^{- \delta pn}|} \,\mathrm{e}^{-f(\delta)(1-p)n}\bigg)\\
&\leq 4\,\mathrm{e}^{-g(\delta)p -f(\delta)(1-p)}\\
&= 4 \,\mathrm{e}^{ -(\Delta_\lambda - h(\delta) ) n},
\end{align*}$$
which decays exponentially fast in n (for
 ${\delta}$
chosen sufficiently small). Therefore
${\delta}$
chosen sufficiently small). Therefore
 ${\mathbb{P}(\mathcal{A}_n \cap \mathcal{C}_n \text{ i.o.}) = 0}$
. Then, since
${\mathbb{P}(\mathcal{A}_n \cap \mathcal{C}_n \text{ i.o.}) = 0}$
. Then, since
 ${\mathbb{P}(\mathcal{A}_n \text{ {eventually}}) = 1}$
, it follows that
${\mathbb{P}(\mathcal{A}_n \text{ {eventually}}) = 1}$
, it follows that
 ${\mathbb{P}(\mathcal{A}_n \cap \mathcal{C}_n^c \text{ {eventually}}) = 1}$
. So
${\mathbb{P}(\mathcal{A}_n \cap \mathcal{C}_n^c \text{ {eventually}}) = 1}$
. So
 ${\mathbb{P}}$
-a.s., for all n large enough,
${\mathbb{P}}$
-a.s., for all n large enough,
 $$\sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{\{ |\tilde{N}_{(1-p)n}^{\hat{\lambda}}(u)| > 0\}} \geq {1}/{2} \mathrm{e}^{f(\delta)(1-p)n} |N_{pn}^{- \delta pn}|
\geq 4 \,\mathrm{e}^{(\Delta_\lambda - h(\delta) )n}.$$
$$\sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{\{ |\tilde{N}_{(1-p)n}^{\hat{\lambda}}(u)| > 0\}} \geq {1}/{2} \mathrm{e}^{f(\delta)(1-p)n} |N_{pn}^{- \delta pn}|
\geq 4 \,\mathrm{e}^{(\Delta_\lambda - h(\delta) )n}.$$
Then, noting that, for all
 ${t \in [n, n+1]}$
,
${t \in [n, n+1]}$
,
 $$|N_t^{\lambda t}| \geq \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \{ |\tilde{N}_{(1-p)n}^{\hat{\lambda}}(u)| > 0\}}{,}$$
$$|N_t^{\lambda t}| \geq \sum_{u \in N_{pn}^{- \delta pn}} \mathbf{1}_{ \{ |\tilde{N}_{(1-p)n}^{\hat{\lambda}}(u)| > 0\}}{,}$$
we obtain
 $$\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \Delta_\lambda - h(\delta) \quad \mathbb{P}
\text{-a.s.}{,}$$
$$\liminf_{t \to \infty} {1}/{t} \log |N_t^{\lambda t}| \geq \Delta_\lambda - h(\delta) \quad \mathbb{P}
\text{-a.s.}{,}$$
which yields the required result after letting
 ${\delta \to 0}$
.
${\delta \to 0}$
.
Proof of (45), the lower bound for (13). Assume that
 ${\lambda > \lambda_{\mathrm{crit}}}$
so that
${\lambda > \lambda_{\mathrm{crit}}}$
so that
 ${\Delta_\lambda < 0}$
. Then necessarily
${\Delta_\lambda < 0}$
. Then necessarily
 $\hat \lambda > {{\hat \beta }_0} > \lambda > {\lambda _{{\rm{crit}}}} \ge {\rm{ }}{(2\hat \beta )^{1/2}}$
(the last inequality follows from the fact that, for any given
$\hat \lambda > {{\hat \beta }_0} > \lambda > {\lambda _{{\rm{crit}}}} \ge {\rm{ }}{(2\hat \beta )^{1/2}}$
(the last inequality follows from the fact that, for any given
 ${\hat{\beta}}$
, the minimum value of
${\hat{\beta}}$
, the minimum value of
 ${{\hat{\beta}}/{\hat{\beta}_0} + \frac{1}{2} \hat{\beta}_0}$
over all
${{\hat{\beta}}/{\hat{\beta}_0} + \frac{1}{2} \hat{\beta}_0}$
over all
 ${\hat{\beta}_0 \in (0, \infty)}$
is
${\hat{\beta}_0 \in (0, \infty)}$
is
 ${(2\hat \beta )^{1/2}}$
). We note that
${(2\hat \beta )^{1/2}}$
). We note that
 $$\mathbb{P} ( |N_t^{\lambda t}| > 0) \geq
\mathbb{P} \bigg( \bigcup_{u \in N_{pt}^{- \delta pt}} \{ |N^{\hat{\lambda}t}_{(1-p)t}(u)| > 0 \},\ |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} \bigg).$$
$$\mathbb{P} ( |N_t^{\lambda t}| > 0) \geq
\mathbb{P} \bigg( \bigcup_{u \in N_{pt}^{- \delta pt}} \{ |N^{\hat{\lambda}t}_{(1-p)t}(u)| > 0 \},\ |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} \bigg).$$
We know that conditional on
 ${\mathcal{F}_{pt}}$
events
${\mathcal{F}_{pt}}$
events
 ${\{|N^{\hat{\lambda}t}_{(1-p)t}(u)|>0\}}$
are independent. Moreover, if we ignore all the catalytic branching taking place in the subtrees initiated by particles
${\{|N^{\hat{\lambda}t}_{(1-p)t}(u)|>0\}}$
are independent. Moreover, if we ignore all the catalytic branching taking place in the subtrees initiated by particles
 ${u \in N_{pt}^{- \delta p t}}$
, then we can get from Proposition 9 (equation (39)) that there exists some deterministic
${u \in N_{pt}^{- \delta p t}}$
, then we can get from Proposition 9 (equation (39)) that there exists some deterministic
 ${t_0}$
such that, for all
${t_0}$
such that, for all
 ${t \geq t_0}$
,
${t \geq t_0}$
,
 ${\mathbb{P} (|N^{\hat{\lambda}t}_{(1-p)t}(u)|>0 \!{\mid}\! \mathcal{F}_{pt} ) \geq \mathrm{e}^{f(\delta)(1-p)t}}$
. Hence
${\mathbb{P} (|N^{\hat{\lambda}t}_{(1-p)t}(u)|>0 \!{\mid}\! \mathcal{F}_{pt} ) \geq \mathrm{e}^{f(\delta)(1-p)t}}$
. Hence
 $$\begin{align*}
&\mathbb{P} \Bigg( \bigcup_{u \in N_{pt}^{- \delta pt}} \{ |N^{\hat{\lambda}t}_{(1-p)t}(u)| > 0 \},\ |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} \Bigg)\\
&\qquad = \mathbb{E} \Bigg( \mathbf{1}_{\{|N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt}\}} \Bigg[ 1 - \prod_{u \in N_{pt}^{- \delta pt}} ( 1 - \mathbb{P}(|N^{\hat{\lambda}t}_{(1-p)t}(u)|>0 \!{\mid}\! \mathcal{F}_{pt}))\Bigg]\Bigg)\\
&\qquad \geq \mathbb{P} ( |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} ) [ 1 - ( 1 -
\mathrm{e}^{f(\delta)(1-p)t} )^{\mathrm{e}^{g(\delta )pt}} ]\\
&\qquad \geq \mathbb{P} ( |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} ) \bigg[ \mathrm{e}^{(\Delta_\lambda - h(\delta))t} - {1}/{2} \mathrm{e}^{2(\Delta_\lambda - h(\delta))t} \bigg]
\end{align*}$$
$$\begin{align*}
&\mathbb{P} \Bigg( \bigcup_{u \in N_{pt}^{- \delta pt}} \{ |N^{\hat{\lambda}t}_{(1-p)t}(u)| > 0 \},\ |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} \Bigg)\\
&\qquad = \mathbb{E} \Bigg( \mathbf{1}_{\{|N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt}\}} \Bigg[ 1 - \prod_{u \in N_{pt}^{- \delta pt}} ( 1 - \mathbb{P}(|N^{\hat{\lambda}t}_{(1-p)t}(u)|>0 \!{\mid}\! \mathcal{F}_{pt}))\Bigg]\Bigg)\\
&\qquad \geq \mathbb{P} ( |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} ) [ 1 - ( 1 -
\mathrm{e}^{f(\delta)(1-p)t} )^{\mathrm{e}^{g(\delta )pt}} ]\\
&\qquad \geq \mathbb{P} ( |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} ) \bigg[ \mathrm{e}^{(\Delta_\lambda - h(\delta))t} - {1}/{2} \mathrm{e}^{2(\Delta_\lambda - h(\delta))t} \bigg]
\end{align*}$$
for all t large enough, and where in the last inequality we have used the fact that, for any
 ${a \in (0,1)}$
and
${a \in (0,1)}$
and
 ${b > 0}$
, it is true that
${b > 0}$
, it is true that
 ${(1-a)^b \leq \mathrm{e}^{-ab} \leq 1 - ab + \frac{1}{2} a^2 b^2}$
. Then, noting that as
${(1-a)^b \leq \mathrm{e}^{-ab} \leq 1 - ab + \frac{1}{2} a^2 b^2}$
. Then, noting that as
 ${t \to \infty}$
${t \to \infty}$
 $${\mathbb{P} ( |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} ) \to 1 \quad \mathbb{P} \text{-a.s.}{,}}$$
$${\mathbb{P} ( |N_{pt}^{- \delta pt}| \geq \mathrm{e}^{g(\delta)pt} ) \to 1 \quad \mathbb{P} \text{-a.s.}{,}}$$
we obtain
 $${\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0 ) \geq \Delta_\lambda - h(\delta){,}}$$
$${\liminf_{t \to \infty} {1}/{t} \log \mathbb{P} (|N_t^{\lambda t}| > 0 ) \geq \Delta_\lambda - h(\delta){,}}$$
which yields the required result after letting
 ${\delta \to 0}$
.
${\delta \to 0}$
.
3.3 Proof of Corollary 1.4
Proof of Corollary 1. Assume that condition (10) is satisfied. Then, for any
 ${\lambda < \lambda_{\mathrm{crit}}}$
, as we know from (11),
${\lambda < \lambda_{\mathrm{crit}}}$
, as we know from (11),
 $$\mathbb{P}( R_t > \lambda t \text{ eventually} ) = \mathbb{P} ( |N_t^{\lambda t}| > 0 \text{ eventually} ) = 1.$$
$$\mathbb{P}( R_t > \lambda t \text{ eventually} ) = \mathbb{P} ( |N_t^{\lambda t}| > 0 \text{ eventually} ) = 1.$$
Thus
 ${\liminf_{t \to \infty}\frac{R_t}{t} \geq \lambda}$
${\liminf_{t \to \infty}\frac{R_t}{t} \geq \lambda}$
 ${\mathbb{P}}$
-almost surely. Then, letting
${\mathbb{P}}$
-almost surely. Then, letting
 ${\lambda \to \lambda_{\mathrm{crit}}}$
gives
${\lambda \to \lambda_{\mathrm{crit}}}$
gives
 $${\liminf_{t \to \infty} {R_t}/{t} \geq \lambda_{\mathrm{crit}} \quad \mathbb{P} \text{-a.s.}}$$
$${\liminf_{t \to \infty} {R_t}/{t} \geq \lambda_{\mathrm{crit}} \quad \mathbb{P} \text{-a.s.}}$$
Similarly, if
 ${\lambda > \lambda_{\mathrm{crit}}}$
, then by (12),
${\lambda > \lambda_{\mathrm{crit}}}$
, then by (12),
 $${\mathbb{P} ( R_t \leq \lambda t \text{ eventually} ) = \mathbb{P} ( |N_t^{\lambda t}| = 0 \text{ eventually} ) = 1.}$$
$${\mathbb{P} ( R_t \leq \lambda t \text{ eventually} ) = \mathbb{P} ( |N_t^{\lambda t}| = 0 \text{ eventually} ) = 1.}$$
Thus
 ${\limsup_{t \to \infty} \frac{R_t}{t} \leq \lambda}$
${\limsup_{t \to \infty} \frac{R_t}{t} \leq \lambda}$
 ${\mathbb{P}}$
-a.s. Then, letting
${\mathbb{P}}$
-a.s. Then, letting
 ${\lambda \to \lambda_{\mathrm{crit}}}$
gives
${\lambda \to \lambda_{\mathrm{crit}}}$
gives
 $${\limsup_{t \to \infty} {R_t}/{t} \leq \lambda_{\mathrm{crit}} \quad \mathbb{P} \text{-a.s.}{,}}$$
$${\limsup_{t \to \infty} {R_t}/{t} \leq \lambda_{\mathrm{crit}} \quad \mathbb{P} \text{-a.s.}{,}}$$
which completes the proof.
Acknowledgements
The first author was supported by NSFC grant 11731012. The second author was supported by NSFC grant 11301020.