



1. Introduction
The potential of ichnology to decipher macro-evolutionary trends in animal behaviour has a long history. Compilation of ichnological data was a major part of the work of A. Seilacher and P. Crimes. Seilacher (Reference Seilacher1974, Reference Seilacher, Crimes and Harper1977, Reference Seilacher, Kitchell and Nitecki1986) focused on the variety of graphoglyptid trace fossils from the deep sea (i.e. patterned trace fossils forming nets, regular meanders and spirals), aiming to understand their environmental adaptation through time (e.g. size changes, functional optimization). Seilacher (Reference Seilacher1956) was the first to recognize the potential of trace fossils to delineate the Precambrian–Cambrian boundary; Crimes (Reference Crimes1987, Reference Crimes1992a, Reference Crimes and Donovan1994) extended this idea by reviewing worldwide literature, and developed an ichnostratigraphic scheme that helped to define the Cambrian GSSP (Narbonne et al. Reference Narbonne, Myrow, Landing and Anderson1987; Brasier et al. Reference Brasier, Cowie and Taylor1994). In Crimes’ comprehensive work, horizontal spiral trace fossils (as defined in this contribution) were consistently absent from the Ediacaran and the Cambrian systems (Crimes, Reference Crimes1987, Reference Crimes1992a, b, Reference Crimes and Donovan1994), only appearing in the Ordovician System in the deep-marine realm (Crimes et al. Reference Crimes, Marcos and Perez-Estaun1974, Reference Crimes, Garcia Hidalgo and Poire1992). Crimes suspected that most deep-marine graphoglyptids originated in shallow-marine environments during the Cambrian Period (Crimes, Reference Crimes1987; Crimes & Anderson, Reference Crimes and Anderson1985; Crimes & Fedonkin, Reference Crimes and Fedonkin1994), but the absence of regular planispiral trace fossils in the Cambrian System was then problematic (Crimes et al. Reference Crimes, Garcia Hidalgo and Poire1992).
The Ediacaran–Cambrian transition (c. 539 Ma) was a time of striking changes in Earth ecosystems. Ediacaran seafloors were dominated by microbially stabilized surfaces on which epifaunal and very shallow infaunal grazers thrived (Seilacher & Pflüger, Reference Seilacher, Pflüger, Krumbein, Paterson and Stal1994; Gehling, Reference Gehling1999). Macroscopic animals of the earliest Cambrian Period started to disrupt the sediment at depth, affecting the substrate ventilation (e.g. Mángano & Buatois, Reference Mángano and Buatois2014; Gougeon et al. Reference Gougeon, Mángano, Buatois, Narbonne and Laing2018a), trophic webs (e.g. Bottjer et al. Reference Bottjer, Hagadorn and Dornbos2000; Meysman et al. Reference Meysman, Middelburg and Heip2006) and geochemical cycles (e.g. Logan et al. Reference Logan, Hayes, Hieshima and Summons1995; Canfield & Farquhar, Reference Canfield and Farquhar2009; Boyle et al. Reference Boyle, Dahl, Bjerrum and Canfield2018). In northwestern France, the Brioverian series of central Brittany is a thick siliciclastic succession that was deposited during the Cadomian cycle (c. 750–540 Ma). Despite the report of fossils since the 19th century, the age of its uppermost beds (Ediacaran or early Cambrian) is a long-standing conundrum. Recently, new investigations unravelled a unique assemblage of trace and body fossils in the vicinity of Rennes (Néraudeau et al. Reference Néraudeau, Gougeon, Dabard and Poujol2016, Reference Néraudeau, Dabard, El Albani, Gougeon, Mazurier, Pierson-Wickmann, Poujol, Saint Martin and Saint Martin2019; Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b, Reference Gougeon, Néraudeau, Poujol and Loi2019). Trace fossils are dominantly simple, horizontal and associated with microbially stabilized surfaces; of these, planispirals represent a surprising discovery.
The aim of this study is threefold: (1) to describe a new assemblage of planispiral trace fossils from the Brioverian series of northwestern France; (2) to place this assemblage within a macro-evolutionary framework and to interpret its significance; and (3) to provide new radiometric dating in order to discuss the age of the uppermost Brioverian deposits.
2. General background
2.a. Geological setting and previous work
The Brioverian series (c. 660–540 Ma; Le Corre et al. Reference Le Corre, Auvray, Ballèvre and Robardet1991; Guerrot et al. Reference Guerrot, Peucat and Dupret1989, Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992) is the informally named thick sedimentary succession deposited during the Cadomian cycle (c. 750–540 Ma) in northwestern France (Fig. 1a, b; Chantraine et al. Reference Chantraine, Egal, Thiéblemont, Le Goff, Guerrot, Ballèvre and Guennoc2001; Ballèvre et al. Reference Ballèvre, Bosse, Dabard, Ducassou, Fourcade, Paquette, Peucat and Pitra2013). In the Rennes area (Fig. 1b), only the uppermost Brioverian beds are exposed, with a thickness evaluated at c. 1300 m (Trautmann et al. Reference Trautmann, Paris and Carn1999). The Brioverian series of Brittany lies unconformably on an Icartian basement (c. 2200–1800 Ma) and is unconformably overlain either by the Ordovician Red Bed Series (‘Séries Rouges Initiales’) or by the Ordovician Armorican Sandstone (‘Grès Armoricain’; Cogné, Reference Cogné1959; D’Lemos et al. Reference D’Lemos, Strachan, Topley, D’Lemos, Strachan and Topley1990; Le Corre et al. Reference Le Corre, Auvray, Ballèvre and Robardet1991). The terrigenous siliciclastic sediments of the Brioverian series resulted from the erosion of the Cadomian belt in northern Brittany and accumulated in a marginal, intra-plate basin in central Brittany (Denis, Reference Denis1988; Dissler et al. Reference Dissler, Doré, Dupret, Gresselin and Le Gall1988; Rabu et al. Reference Rabu, Chantraine, Chauvel, Denis, Balé, Bardy, D’Lemos, Strachan and Topley1990; Dabard et al. Reference Dabard, Loi and Peucat1996). Locally, carbonaceous cherts (‘phtanites’), limestones and igneous intrusions have also been reported (Denis & Dabard, Reference Denis and Dabard1988; Dabard, Reference Dabard1990, Reference Dabard2000; Chantraine et al. Reference Chantraine, Egal, Thiéblemont, Le Goff, Guerrot, Ballèvre and Guennoc2001). On a regional scale, the correlation of the Brioverian sedimentary deposits is hindered by the discontinuous cropping out, facies changes, the absence of biostratigraphic markers and the metamorphic overprint from the Devonian–Carboniferous Variscan orogeny (Denis & Dabard, Reference Denis and Dabard1988; D’Lemos et al. Reference D’Lemos, Strachan, Topley, D’Lemos, Strachan and Topley1990; Le Corre et al. Reference Le Corre, Auvray, Ballèvre and Robardet1991; Ballèvre et al. Reference Ballèvre, Bosse, Dabard, Ducassou, Fourcade, Paquette, Peucat and Pitra2013). While traditionally interpreted as deeper-marine turbiditic deposits (Dangeard et al. Reference Dangeard, Doré and Juignet1961; Denis, Reference Denis1988; Trautmann et al. Reference Trautmann, Paris and Carn1999), the Brioverian sedimentary beds also show evidence of shallow-marine storm-influenced (Dabard & Loi, Reference Dabard and Loi1998; Dabard & Simon, Reference Dabard and Simon2011) and marginal-marine tidally influenced conditions (Graindor, Reference Graindor1957; Dabard, Reference Dabard1990, Reference Dabard2000; Néraudeau et al. Reference Néraudeau, Dabard, El Albani, Gougeon, Mazurier, Pierson-Wickmann, Poujol, Saint Martin and Saint Martin2019).

Fig. 1. Geological map of northwestern France, and new U–Pb dating. (a) Location of the Brioverian deposits in northwestern France. (b) Close-up showing the Brioverian deposits, and the three localities with planispiral trace fossils. NASZ – North Armorican Shear Zone; SASZ – South Armorican Shear Zone. (c) Kernel density estimation diagrams for La Lammerais and Le Lorinou samples.
Fossils recovered from the Brioverian series are algal or bacterial organic-walled microfossils in cherts and limestones (Cayeux, Reference Cayeux1894; Deflandre, Reference Deflandre1955; Chauvel & Schopf, Reference Chauvel and Schopf1978; Chauvel & Mansuy, Reference Chauvel and Mansuy1981; Mansuy & Vidal, Reference Mansuy and Vidal1983), macroscopic body fossils of unknown origin (Néraudeau et al. Reference Néraudeau, Dabard, El Albani, Gougeon, Mazurier, Pierson-Wickmann, Poujol, Saint Martin and Saint Martin2019) and trace fossils. Ichnofossils were first discovered in the late 19th century (Lebesconte, Reference Lebesconte1886), but did not draw the attention of the scientific community for a long time. Recently, new investigations in the vicinity of Rennes (Fig. 1b) have unravelled an assemblage dominated by simple horizontal grazing trails (Circulichnis, Gordia, Helminthoidichnites, Helminthopsis), passively filled horizontal burrows (Palaeophycus) and horizontal spiral trace fossils (Spirodesmos; Néraudeau et al. Reference Néraudeau, Gougeon, Dabard and Poujol2016; Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b, Reference Gougeon, Néraudeau, Poujol and Loi2019). In addition, microbially textured surfaces (MISS of Noffke et al. Reference Noffke, Gerdes, Klenke and Krumbein2001) are common both in fossiliferous and azoic intervals (Lebesconte, Reference Lebesconte1886; Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b).
The age of the uppermost Brioverian sedimentary beds in Brittany, Normandy and Mayenne has been highly debated (Fig. 1b). In Brittany, the overlying Red Bed Series gave an age of 472 ± 5 Ma (Rb–Sr dating from volcanic rocks; Auvray et al. Reference Auvray, Mace, Vidal and Van der Voo1980), 465 ± 1 Ma (U–Pb dating from volcanic rocks; Bonjour et al. Reference Bonjour, Peucat, Chauvel, Paris and Cornichet1988; Bonjour & Odin, Reference Bonjour and Odin1989) and 486 ± 28 Ma (Pb–Pb dating from volcanic rocks; Guerrot et al. Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992), placing these beds within the Ordovician Period (contra McMahon et al. Reference McMahon, Davies and Went2017; Went, Reference Went2017). In the westernmost part of Brittany (Crozon area; Fig. 1b), Guerrot et al. (Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992) obtained an age of 543 ± 18 Ma (Pb–Pb dating) for a tuff intercalated within Brioverian beds, whereas a maximum deposition age of 546 ± 2 Ma (U–Pb dating) has been obtained by Ballouard et al. (Reference Ballouard, Poujol and Zeh2018) from detrital zircon grains extracted from a sandstone (see also Dabard et al. Reference Dabard, Loi, Pavanetto, Meloni, Hauser, Matteini and Funedda2021). In the vicinity of Rennes (Fig. 1b), detrital zircon grains gave a maximum deposition age of c. 550 Ma (U–Pb dating from sandstone and siltstone; Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b); however, five zircon grains dated at 532.1 ± 3.9 Ma were problematic to interpret (Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b). In Normandy, Brioverian sediments were deposited in a different palaeogeographic domain than in central Brittany, as they are separated by the North Armorican Shear Zone (Fig. 1b; Chantraine et al. Reference Chantraine, Chauvel, Dupret, Gatinot, Icart, Le Corre, Rabu, Sauvan and Villey1982; Guerrot et al. Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992). In this domain, granitoid intrusions within Brioverian sediments have been dated at 540 ± 10 Ma (U–Pb dating on monazite; Pasteels & Doré, Reference Pasteels, Doré and Odin1982). In Mayenne, where the Brioverian series is in continuity with its equivalent of central Brittany (Fig. 1b), radiometric dating on zircon grains yielded an age of 540 ± 17 Ma (tuff and detrital horizons; Guerrot et al. Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992).
2.b. Outcrops under study and depositional environments
The outcrop at La Lammerais village nearby St-Gonlay (Fig. 1b) yields slates with trace fossils, stacked in a pile c. 2 m high and 50 m long (outcrops for this contribution are located on private properties, which do not allow details on their exact locations). These slates were extracted from a pit that was exploited by locals to build houses and pathways decades ago; unfortunately, the pit is now covered with vegetation and therefore impossible to sample in situ. Slates are made of siltstone and rare very-fine- to fine-grained sandstone. One loose sandstone sample was collected for U–Pb dating, coming from a nearby agricultural field. At La Lammerais outcrop, sedimentary structures are parallel-lamination organized in siltstone–sandstone bundles (i.e. rhythmite-like; Néraudeau et al. Reference Néraudeau, Dabard, El Albani, Gougeon, Mazurier, Pierson-Wickmann, Poujol, Saint Martin and Saint Martin2019, fig. 3), current-ripples (Néraudeau et al. Reference Néraudeau, Dabard, El Albani, Gougeon, Mazurier, Pierson-Wickmann, Poujol, Saint Martin and Saint Martin2019, fig. 2), tool-marks/spindle-shaped flute-marks, possible load-casts, and pustular and wrinkled microbially textured surfaces (Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b, figs 4, 7).
Nearby St-Gonlay, another outcrop has been investigated at Le Lorinou locality, situated 1.4 km to the east of La Lammerais. This outcrop is very poor in trace fossils (no spiral trace fossils were found there), but beds are preserved in situ and a sandstone sample was collected for U–Pb zircon dating.
The outcrop of Le Bois-du-Buisson is located at the entrance of a small forest in Montfort-sur-Meu (Fig. 1b). It consists of a small quarry of c. 3 m high and 10 m long, with vegetation extensively covering the sedimentary beds. However, a few siltstone beds are accessible and reveal fresh surfaces with trace fossils. Sedimentary structures are parallel-lamination organized in siltstone–sandstone bundles (i.e. rhythmite-like, similar to what is found at La Lammerais), and pustular and wrinkled microbially stabilized surfaces.
In addition, Montfort-sur-Meu is the host of Les Grippeaux quarry, where P. Lebesconte recovered fossils for the first time in the late 19th century (Lebesconte, Reference Lebesconte1886; Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b). Nowadays, the quarry is secured by a fence preventing any access. Many samples were collected in the late 19th and early 20th century by P. Lebesconte, F. Kerforne and other geologists; they are housed at the Geological Institute of the University of Rennes 1 and at the Museum of Natural History of Nantes, and are available for study. Sedimentary structures are pustular microbially stabilized surfaces.
The outcrop in Crozon (Fig. 1b) is located on the coastal cliff at La Plage-du-Goulien. This outcrop has not been visited by the authors, and the only trace fossil discovered was reported by E. Hanson in 2014 (pers. comm.). The sedimentology of the Brioverian series from the Bay of Douarnenez and the Cove of Dinan (both in the vicinity of Crozon) has been studied in two doctoral theses (J.R. Darboux, unpubl. Ph.D. thesis, University of Brest, 1973; Denis, Reference Denis1988). The succession displays parallel-laminated/bedded sandstone and siltstone with flute-casts, load-casts, tool-marks, rip-up clasts, carbonate concretions, normal and reverse grading, convolute bedding, flame structures, and current, wave and climbing ripples. Both authors interpreted the succession as deposited by turbidites, located either below the limit of the storm wave-base action, or deeper in an abyssal plain. However, Denis (Reference Denis1988) noted the presence of oscillatory flow structures, lenticular bedding (Facies 3 of Denis, Reference Denis1988) and mud-drapes, which are more typical of shallower environments.
In Montfort-sur-Meu and St-Gonlay, the dominance of siltstone intercalated with laminated very-fine-grained sandstone, and the record of rhythmite-like bundles and current ripples, suggest a marginal-marine, tidally influenced depositional environment (cf. Nio & Yang, Reference Nio, Yang, Smith, Reinson, Zaitlin and Rahmani1991; Tessier et al. Reference Tessier, Archer, Lanier, Feldman, Flemming and Bartholomä1995; Dalrymple, Reference Dalrymple, James and Dalrymple2010). This conclusion is strengthened by observations in Chanteloup and Nouvoitou (both in the vicinity of Rennes), where a sandstone facies displays mudstone drapes within fine-grained sandstone samples (i.e. flaser lamination; see online Supplementary Fig. S1); these areas could represent the seaward, sandier part of the intertidal system. Sedimentary structures made by oscillatory flows (e.g. wave ripples, hummocky cross-stratification) have not been observed in the area so far. These conclusions are preliminary and await further support, notably from Brioverian outcrops revealing bedding architecture, and from more sampling of sedimentary structures.
3. Materials and methods
3.a. Terminology of planispiral trace fossils
This contribution focuses only on spirals formed on a horizontal plane (i.e. planispirals). Three-dimensional, vertically (e.g. Gyrolithes, Lapispira) or horizontally (e.g. Avetoichnus, Helicodromites, Helicolithus) oriented spirals are not comparable with the Brioverian material. In order to describe spiral morphologies, the following terms will be used: a regular spiral maintains a constant distance between whorls (Fig. 2b, c); an irregular spiral has a variable distance between whorls (Fig. 2a, d); a one-way spiral is a simple spiral with no central turnaround (Fig. 2a, b, d; Seilacher, Reference Seilacher, Crimes and Harper1977; Crimes & McCall, Reference Crimes and McCall1995); a two-way spiral is a double spiral with a central turnaround (Fig. 2c; Seilacher, Reference Seilacher, Crimes and Harper1977; Crimes & McCall, Reference Crimes and McCall1995); and a bounded spiral is an irregular, one-way spiral that decreases the distance between whorls outward (Fig. 2d).

Fig. 2. Terminology of planispirals: (a) an irregular one-way spiral; (b) a regular one-way spiral; (c) a regular two-way spiral; and (d) a bounded spiral. See text for further explanations.
3.b. U–Pb dating method
A classic mineral separation procedure has been applied to concentrate zircon grains for U–Pb dating. Rocks were crushed and only the powder fraction with a diameter < 250 µm was kept. Heavy minerals were successively concentrated by Wilfley table, heavy liquids and an isodynamic Frantz separator. Zircon grains were then handpicked under a binocular microscope to produce the most representative sampling, with the aim of avoiding any intentional bias (see Malusà et al. Reference Malusà, Carter, Limoncelli, Villa and Garzanti2013). Selected grains were then embedded in epoxy mounts, grounded and polished. Zircon grains were imaged by cathodoluminescence (CL) using a Reliotron CL system equipped with a digital colour camera available at the GeOHeLiS analytical platform (University of Rennes 1).
U–Pb geochronology was conducted by in situ laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) at the GeOHeLiS analytical platform using an ESI NWR193UC Excimer laser coupled to a quadripole Agilent 7700× ICP-MS. Instrumental conditions are reported in online Supplementary Table S1, while the analytical protocol can be found in Manzotti et al. (Reference Manzotti, Poujol and Ballèvre2015). Kernel density diagrams for analyses that are 100 ± 10% concordant were generated using IsoplotR (Vermeesch, Reference Vermeesch2018). When dealing with detrital zircon geochronology, a minimum of three different ages obtained on three different zircon grains overlapping in age at 2σ has been demonstrated to produce a statistically robust maximum deposition age (Dickinson & Gehrels, Reference Dickinson and Gehrels2009). The second important criteria in order to determine this maximum deposition age is the degree of concordance of the individual analysis used to calculate this age. Most authors consider all analyses that are 90% concordant or more, while others only consider analyses that are at least 95% concordant. In this study, because of the complexity of one of the datasets (La Lammerais), only analyses that were at least 95% concordant were considered to calculate the maximum deposition age, in order to avoid using apparent ages that could be younger than the true age due to a non-negligible Pb loss.
3.c. Museum repository
From La Lammerais and Le Bois-du-Buisson, samples were collected and reposited at the Geological Institute of the University of Rennes 1 (collections Gougeon and Néraudeau). Historical specimens from Les Grippeaux are reposited at the Museum of Natural History of Nantes (collections Barrois and Lebesconte) and the Geological Institute of the University of Rennes 1 (collections Kerforne, Rolland and Rouault). The trace fossil from La Plage-du-Goulien has not been collected and was only photographed in the field by E. Hanson.
4. Results
4.a. Planispiral trace fossils from the Brioverian series
The Brioverian series of central Brittany contains a rich assemblage of simple horizontal trace fossils, with Helminthoidichnites and Helminthopsis being the most common forms. Originally, Lebesconte (Reference Lebesconte1886, pl. 34, fig. 7) figured a planispiral trace fossil from Montfort-sur-Meu without further discussion. Since then, spiral trace fossils have not been reported in the Brioverian series. Here, we describe two types of planispiral trace fossils: (1) irregular, one-way spiral trace fossils; and (2) regular, one-way spiral trace fossils.
Four irregular, one-way spiral trace fossils were recovered from La Lammerais, Le Bois-du-Buisson and Les Grippeaux (Fig. 3a). Specimens are 0.3–1 mm wide, have 1¼–1¾ whorls, and are preserved in positive and negative reliefs (preservation as epirelief or hyporelief is unknown because slates with trace fossils are not preserved in situ). One specimen (Fig. 3a) has a different infill than the host rock and a lining; this is potentially a burrow. Rarely, they are associated with Helminthoidichnites, small-scale branching trace fossils (cf. Pilichnus), and pits of uncertain affinity. They are commonly found on pustular microbially textured surfaces.

Fig. 3. Spirodesmos isp. and Spirodesmos archimedeus from the uppermost Brioverian beds of Brittany. (a) Spirodesmos isp. (irregular one-way spiral trace fossil) from Le Bois-du-Buisson (IGR 2852). (b, c) Spirodesmos archimedeus (regular one-way spiral trace fossils) from (b) Les Grippeaux (IGR 112628) and (c) La Plage-du-Goulien. Scale bars are 1 cm across.
Two regular, one-way spiral trace fossils were recovered from La Plage-du-Goulien and Les Grippeaux (Fig. 3b, c). Specimens are 1–3 mm wide, have 2¼–2½ whorls, and are preserved in positive and negative reliefs. The distance between whorls remains constant until the last whorl, where the course detaches from the spiral system and progressively disappears. They are associated with Helminthopsis and pits of uncertain affinity. The surfaces they are found on are not textured.
Spirodesmos, Spirophycus and Spirorhaphe are the most common planispirals from the trace fossil record: however, their morphological boundaries are unclear. Spirodesmos is a regular to irregular, one-way spiral trace fossil (Geinitz, Reference Geinitz, Geinitz and Liebe1867; Andrée, Reference Andrée1920; Huckriede, Reference Huckriede1952; Xia et al. Reference Xia, Lu, Xiong, He and Hu1987). For Seilacher (Reference Seilacher, Crimes and Harper1977), Spirodesmos has a wide spacing between whorls; although this is clearly so in the type ichnospecies S. interruptus Andrée, Reference Andrée1920, S. archimedeus Huckriede, Reference Huckriede1952 has a narrower spacing between whorls. This issue becomes critical with Spirodesmos kaihuaensis Xia, He & Hu, Reference Xia, Lu, Xiong, He and Hu1987 and S. spiralis (Geinitz, Reference Geinitz, Geinitz and Liebe1867), both having irregular courses with variable distances between whorls. Spirophycus is a regular to irregular, one-way spiral trace fossil that commonly grades into meanders (Heer, Reference Heer1876; Häntzschel, Reference Häntzschel1975). Seilacher (Reference Seilacher, Crimes and Harper1977) argued that Spirophycus has wide strings with a tubercular surface and backfilled laminae (see also Książkiewicz, Reference Książkiewicz1977; but see Uchman, Reference Uchman1998). The spiral portion of Spirophycus (e.g. Heer, Reference Heer1876, pl. 66, fig. b; Sacco, Reference Sacco1888, pl. 2, fig. 14), with regular whorls distinctly spaced from each other, can however be very similar to Spirodesmos archimedeus. Spirorhaphe is a regular to irregular spiral trace fossil with either a one-way (S. azteca, S. graeca) or two-way (S. involuta) course (Seilacher, Reference Seilacher, Crimes and Harper1977; Crimes & McCall, Reference Crimes and McCall1995). The inclusion of one-way spirals in Spirorhaphe is overlapping with Spirodesmos and Spirophycus morphologies, which is problematic.
Despite these taxonomical issues (see also Crimes & Crossley, Reference Crimes and Crossley1991; Uchman, Reference Uchman1998; Minter & Braddy, Reference Minter and Braddy2009), planispirals from the Brioverian series are comparable to Spirodesmos. Brioverian irregular, one-way spiral trace fossils are reminiscent of Spirodesmos spiralis (cf. Geinitz, Reference Geinitz, Geinitz and Liebe1867; Stepanek & Geyer, Reference Stepanek and Geyer1989). However, because of its irregular course and the poor extent of its whorls, Spirodesmos spiralis needs to be re-evaluated taxonomically; in this study, Brioverian forms are referred to Spirodesmos isp. Conversely, Brioverian regular, one-way spiral trace fossils belong to Spirodesmos archimedeus (cf. Huckriede, Reference Huckriede1952; Zapletal & Pek, Reference Zapletal and Pek1971; Horn, Reference Horn1989). Spirodesmos ranges from the Ediacaran–Cambrian (this study) to the Holocene periods (e.g. Kitchell et al. Reference Kitchell, Kitchell, Johnson and Hunkins1978; Smith et al. Reference Smith, Holland and Ruhl2005).
4.b. U–Pb dating
For the sandstone sample from La Lammerais, 118 zircon grains were analysed, among which 107 analyses have a concordance of 100 ± 10%. Their U (49–1195 ppm) and Pb (6–493 ppm) contents, as well as their Th/U ratios (0.02–1), are highly variable (see online Supplementary Table S2). A first group of 12 analyses yields apparent ages between 2.8 and 1.06 Ga. The remaining analyses form two major peaks at 600 and 550 Ma, with minor peaks around 850 and 650 Ma (Fig. 1c). The 10 youngest analyses that are more than 95% concordant yield a weighted average 206Pb/238U date of 540 ± 5 Ma (mean square weighted deviation (MSWD), 1.2) that we consider as the maximum deposition age for this sandstone.
For the sandstone sample from Le Lorinou, 89 grains were analysed, out of which 68 are 100 ± 10% concordant (online Supplementary Table S2). They are characterized by variable U and Pb contents (21–726 ppm and 2–242 ppm, respectively), with Th/U ratios between 0.05 and 1.4. A first group of 22 zircon grains yields Neoarchean (2.9 Ga) to Palaeoproterozoic (1.8 Ga) ages, followed by a gap until the end of the Mesoproterozoic. The remaining grains present apparent ages around 1000, 900, 800 and 680 Ma, with a major peak around 600 Ma (Fig. 1c). The youngest three grains provide a weighted 206Pb/238U date of 551 ± 7 Ma (MSWD, 0.009) that we consider as the maximum deposition age for this sandstone.
5. Discussion
5.a. Radiometric age of the uppermost Brioverian beds with trace fossils
At Le Lorinou (St-Gonlay), zircon grains from a sandstone bed associated with trace fossils yielded a maximum deposition age of 551 ± 7 Ma, whereas zircon grains from a loose sandstone sample at La Lammerais (St-Gonlay) gave a maximum deposition age of 540 ± 5 Ma (this study). In Crozon, the youngest U–Pb dating on zircon grains from Brioverian tuff gave an age of 543 ± 18 Ma (Guerrot et al. Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992), whereas another U–Pb dating on zircon grains from a sandstone gave a maximum deposition age of 546 ± 2 Ma (Ballouard et al. Reference Ballouard, Poujol and Zeh2018). In Montfort-sur-Meu, no radiometric dating has been done so far.
The radiometric age for the base of the Cambrian is given by U–Pb dating on zircon grains from tuff in southern Oman, dated at 541.0 ± 0.13 Ma (Bowring et al. Reference Bowring, Grotzinger, Condon, Ramezani, Newall and Allen2007). However, recent U–Pb dating on zircon grains from tuff in southern Namibia constrained the age of the basal Cambrian within a 538.6–538.8 Ma interval (Linnemann et al. Reference Linnemann, Ovtcharova, Schaltegger, Gärtner, Hautmann, Geyer, Vickers-Rich, Rich, Plessen, Hofmann, Zieger, Krause, Kriesfeld and Smith2019). If we consider a radiometric age of c. 539 Ma for the base of the Cambrian System, the Brioverian beds of Crozon could either be Ediacaran (c. 635–539 Ma) in age or younger, while the Brioverian of St-Gonlay could be Ediacaran but is suspected to be Fortunian (c. 539–529 Ma) in age or younger. Indeed, the new results of this study agree with earlier dating elsewhere (maximum deposition age of c. 550 Ma in Néant-sur-Yvel, Brittany, with five zircons grains dated at 532.1 ± 3.9 Ma; 540 ± 10 Ma in Normandy; and 540 ± 17 Ma in Mayenne; Pasteels & Doré, Reference Pasteels, Doré and Odin1982; Guerrot et al. Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992; Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b) and suggest an early Cambrian age for the uppermost Brioverian beds, from a radiometric standpoint (see also Guerrot et al. Reference Guerrot, Peucat and Dupret1989, Reference Guerrot, Calvez, Bonjour, Chantaine, Chauvel, Dupret and Rabu1992; Dabard et al. Reference Dabard, Loi, Pavanetto, Meloni, Hauser, Matteini and Funedda2021). In terms of trace fossil biostratigraphy, the matground ecology of the Brioverian series is more typical of late Ediacaran assemblages (Gougeon et al. Reference Gougeon, Néraudeau, Dabard, Pierson-Wickmann, Polette, Poujol and Saint-Martin2018b). Because of the differences provided by these two proxies (radiometric dating and ichnostratigraphy), definitive conclusions on the age of the uppermost Brioverian beds with trace fossils are not possible at this point.
5.b. Critical review on Ediacaran and Cambrian planispiral trace fossils
Several trace fossils inaccurately described or suggested to be planispirals, have been reported from the Ediacaran and the Cambrian systems. Fedonkin (Reference Fedonkin, Sokolov and Ivanovskiy1985, Reference Fedonkin, Sokolov and Iwanoski1990) erected Planispiralichnus Fedonkin, Reference Fedonkin, Sokolov and Ivanovskiy1985 and Protospiralichnus Fedonkin, Reference Fedonkin, Sokolov and Ivanovskiy1985 from the Fortunian Kessyuse Formation of northern Russia. Planispiralichnus is made of dense, overlapping loops (Fedonkin, Reference Fedonkin, Sokolov and Iwanoski1990; Marusin & Kuper, Reference Marusin and Kuper2020), whereas Protospiralichnus starts as a bounded spiral until it scribbles abundantly (Fedonkin, Reference Fedonkin, Sokolov and Iwanoski1990); because of their scribbling patterns, neither of them represent spirals (Buatois et al. Reference Buatois, Wisshak, Wilson and Mángano2017). Jenkins (Reference Jenkins1995, pl. 2, fig. E) reported cf. Protospiralichnus from the Ediacaran Rawnsley Quartzite of southern Australia; the development of a full circle and/or a loop affiliates this trace fossil to Circulichnis or Gordia instead (see also Buatois & Mángano, Reference Buatois, Mángano, Mángano and Buatois2016, fig. 2.8e). Multilaqueichnus Yang & Yin (in Yang et al. Reference Yang, Yin and He1982) from the Cambrian Stage 3 Jiulaodong Formation of central China has overlapping loops (Yang et al. Reference Yang, Yin and He1982, pl. 2, fig. 1); these trace fossils distinctly scribble and are not spirals either (contra Mángano & Buatois, Reference Mángano, Buatois, Mángano and Buatois2016, Reference Mángano and Buatois2020). Finally, a trace fossil from the Fortunian part of the Chapel Island Formation of eastern Canada was considered a spiral by Crimes & Fedonkin (Reference Crimes and Fedonkin1994, fig. 2i). This trace fossil has been observed in the field by one of the authors (R.G.) and represents the scribbling burrow of a large infaunal deposit-feeder.
In addition, non-ichnological structures from the Ediacaran and the Cambrian have been mistaken for planispiral trace fossils. An important debate arose with the report of Precambrian spiral fossils from the Lower Vindhyan Limestone of northeastern India (Beer, Reference Beer1919) and from the Belt Series of northwestern USA (Walcott, Reference Walcott1899). Both authors suggested a trace fossil origin, an opinion followed by Seilacher (Reference Seilacher1956). However, Cloud (Reference Cloud and Drake1968) suspected an algal origin, and re-evaluation of both materials confirmed that view (Walter et al. Reference Walter, Oehler and Oehler1976; Runnegar, Reference Runnegar1991). Arenicolites spiralis Billings, Reference Billings1872 and Helminthoidichnites sangshuanensis Du (in Du et al. Reference Du, Tian and Li1986; Yan & Liu, Reference Yan and Liu1998) are certainly of similar algal affinity (Hofmann, Reference Hofmann1971; Walter et al. Reference Walter, Du and Horodyski1990; Shaowu, Reference Shaowu1998). Furthermore, Aceñolaza (Reference Aceñolaza2005) reported circular structures from the Cambrian Mesón Group of northwestern Argentina, and erected the new ichnospecies Spirodesmos milanai; Minter et al. (Reference Minter, Buatois, Lucas, Braddy and Smith2006) argued these structures were formed by shrinkage cracks in matgrounds instead (cf. Pflüger, Reference Pflüger1999; Buatois et al. Reference Buatois, Netto, Mángano and Carmona2013; Sedorko et al. Reference Sedorko, Netto and Horodyski2019).
Finally, rare horizontal spiral trace fossils have been reported from the Ediacaran and the Cambrian systems. Planispiralichnus rarus Menasova, Reference Menasova2003 is a one-way spiral trace fossil discovered in the Fortunian Khmelnitsky Formation of western Ukraine. The holotype is made of three whorls with angular intervals along the course; the first two whorls are continuous, while the last one is made of unconnected segments (Ivantsov et al. Reference Ivantsov, Gritsenko, Paliy, Velikanov, Konstantinenko, Menasova, Fedonkin, Zakrevskaya and Serezhnikova2015, pl. 7, fig. 4a, b). This specimen possesses the key features of a spiral trace fossil and should not be affiliated to Planispiralichnus as described by Fedonkin (Reference Fedonkin, Sokolov and Iwanoski1990). Jensen & Palacios (Reference Jensen and Palacios2016, fig. 4b) reported one-way spiral trace fossils from the Ediacaran–Fortunian Cíjara Formation of central Spain. The photographed specimen is a continuous to discontinuous trail, with 2½ whorls and an irregular course. Carbone & Narbonne (Reference Carbone and Narbonne2014, fig. 4.5) also figured an irregular one-way spiral with 1¾ whorls and a continuous course from the Fortunian part of the Ingta Formation of northwestern Canada. Finally, Runnegar (Reference Runnegar and Schopf1992, fig. 3.9) figured an irregular two-way spiral transitional with a meandering trace fossil from the Ediacaran Rawnsley Quartzite of southern Australia. Jensen (Reference Jensen2003) considered this trace fossil to represent Helminthorhaphe grading into Spirorhaphe.
5.c. Macro-evolutionary profile and onshore–offshore migration
Our detailed literature review (Section 5.b above and online Supplementary Material) unravelled the environmental and temporal distribution of Spirodesmos (Fig. 4). The Ediacaran System and the Fortunian Stage are then marked by the appearance of a Spirodesmos pool, composed of irregular and regular forms colonizing marginal-marine to shelf environments. The Brioverian assemblage represents a key component of that pool, holding the oldest regular one-way planispirals (Spirodesmos archimedeus) both in marginal-marine and open-shelf settings (in Montfort-sur-Meu and Crozon, respectively). During the Ediacaran and the Cambrian periods, deposit-feeding was the dominant feeding strategy (MacNaughton & Narbonne, Reference MacNaughton and Narbonne1999; Carbone & Narbonne, Reference Carbone and Narbonne2014), and early Spirodesmos were arguably made by epifaunal detritus-feeders and shallow-infaunal deposit-feeders. Indeed, ‘surplus stretches’ as observed in Spirorhaphe and inferring open-burrow systems (Seilacher, Reference Seilacher and Hediger1967a, b, Reference Seilacher, Crimes and Harper1977) have not been observed in Ediacaran–Fortunian material. Moreover, Ediacaran–Fortunian Spirodesmos are often preserved on microbially stabilized surfaces, which could represent the nutritive resource of their tracemaker (Carbone & Narbonne, Reference Carbone and Narbonne2014). Possible producers are enteropneusts and nematodes, both suspected to first appear during the Cambrian Period or before (Knoll & Carroll, Reference Knoll and Carroll1999; Budd & Jensen, Reference Budd and Jensen2000; Maletz, Reference Maletz2014; Cunningham et al. Reference Cunningham, Liu, Bengtson and Donoghue2017). Enteropneusts produce regular horizontal spirals on the modern deep-sea floor on areas of greater nutritional values, using tactile sensory systems in their head (Lemche et al. Reference Lemche, Hansen, Madsen, Tendal and Wolff1976; Smith et al. Reference Smith, Holland and Ruhl2005; Jones et al. Reference Jones, Alt, Priede, Reid, Wigham, Billett, Gebruk, Rogacheva and Gooday2013). Nematodes spiral by contracting all the muscles of one side of their body (Wharton, Reference Wharton, Gaugler and Bilgrami2004). However, spiralling in nematodes has been suggested for other purposes than feeding (e.g. responses to increasing temperature, osmotic stress, desiccation and for reproduction; Huettel, Reference Huettel, Gaugler and Bilgrami2004; Wharton, Reference Wharton, Gaugler and Bilgrami2004).
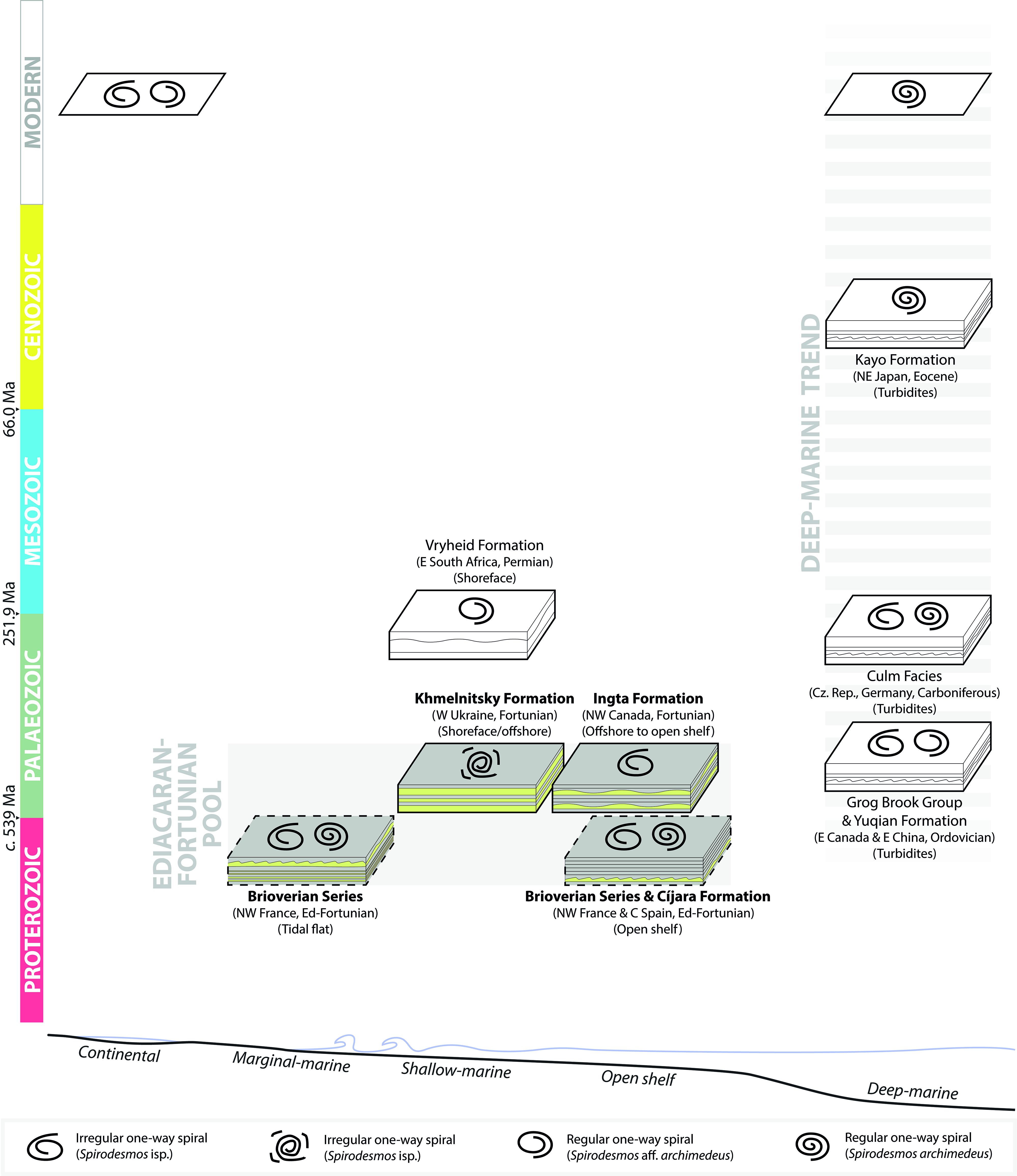
Fig. 4. Macro-evolutionary profile of Spirodesmos comparable to the Brioverian material, with emphasis on the Ediacaran and the Cambrian systems (coloured diagrams). Positions suggested for the Brioverian material are emphasized by diagrams with dashed lines. Brioverian Spirodesmos belonged to an Ediacaran–Fortunian, marginal-marine to shelf pool. A Spirodesmos onshore–offshore migration is suggested, during Ediacaran–Fortunian to Ordovician time. See text for further explanations.
Although deep-marine deposits with trace fossils have been reported both from the Ediacaran (e.g. Narbonne & Hofmann, Reference Narbonne and Hofmann1987; Liu et al. Reference Liu, McIlroy and Brasier2010) and the Cambrian (e.g. Hofmann et al. Reference Hofmann, Cecile and Lane1994; Seilacher et al. Reference Seilacher, Buatois and Mángano2005) systems, planispirals are consistently absent. However, planispirals are common through the rest of the Phanerozoic Eonothem in the deep-sea, and Spirodesmos forms a conspicuous Ordovician–Recent deep-marine trend (Fig. 4; online Supplementary Material). Deep-marine seafloors are characterized by an absence of light, high hydrostatic pressure, oxygen and temperature fluctuations, and low nutrient content (Sanders & Hessler, Reference Sanders and Hessler1969; Gage & Tyler, Reference Gage and Tyler1991, pp. 9–29; Rex & Etter, Reference Rex and Etter2010, pp. 1–49). These stressful conditions play an important role on animal fitness and their physiology (e.g. Childress & Thuesen, Reference Childress, Thuesen, Rowe and Pariente1992; Yancey et al. Reference Yancey, Rhea, Kemp and Bailey2004; van der Grient & Rogers, Reference Van der Grient, Rogers and Curry2015). However, with the increased competition for space and food on early Cambrian shelves (Orr, Reference Orr2001), planispiral tracemakers may have adapted their metabolisms to the deep-sea. An onshore–offshore migration in Spirodesmos is then suggested during Ediacaran–Fortunian to Ordovician time, similarly to the migration observed in graphoglyptids (Crimes & Anderson, Reference Crimes and Anderson1985; Crimes et al. Reference Crimes, Garcia Hidalgo and Poire1992; Crimes & Fedonkin, Reference Crimes and Fedonkin1994; Orr, Reference Orr2001; Uchman, Reference Uchman2003). The existence of an onshore–offshore migration is also reinforced by the presence of an important gap (c. 230 Ma) between the Ediacaran–Fortunian Spirodesmos pool and the next shallow-marine Spirodesmos report from the Permian Vryheid Formation of eastern South Africa (Mason et al. Reference Mason, Stanistreet and Tavener-Smith1983; Fig. 4).
6. Conclusion
The age of the uppermost Brioverian deposits of central Brittany, northwestern France, is a long-standing question. Here, two U–Pb detrital zircon grain datings on sandstone samples collected in St-Gonlay gave maximum depositional ages of 551 ± 7 Ma and 540 ± 5 Ma. Although an Ediacaran age for the uppermost Brioverian beds cannot be discarded, a Fortunian age is suggested in this study, following previous dating in Brittany, Normandy and Mayenne. However, the intervals of error of the radiometric dating and trace fossil biostratigraphy do not allow definitive conclusions.
A unique assemblage of irregular and regular, one-way planispiral trace fossils of Spirodesmos affinity has been recovered from the uppermost Brioverian beds in Crozon, Montfort-sur-Meu and St-Gonlay. Planispiral trace fossils are unusual in the Ediacaran and the Cambrian systems, and an in-depth literature review revealed that the Brioverian assemblage belonged to an Ediacaran–Fortunian, marginal-marine to shelf Spirodesmos pool. Ediacaran–Fortunian Spirodesmos were arguably made by detritus- or deposit-feeders, possibly related to enteropneusts or nematodes. However, by the Ordovician Period, Spirodesmos became conspicuous mostly in the deep-marine realm, underscoring an onshore–offshore migration similar to what has been reported in graphoglyptids.
Acknowledgements
This manuscript is a follow-up to a poster presented at the International Meeting on the Ediacaran System and the Ediacaran–Cambrian Transition (IMECT) that took place in Guadalupe, Spain in October 2019. Discussions and feedback from L. Buatois, P. Crimes, G. Mángano, V. Marusin and M. Paz are kindly acknowledged. We also thank M. Ferguson for help with language proofing; D. Gendry and S. Regnault for granting access to the museum of the Geological Institute of the University of Rennes 1 and the Museum of Natural History of Nantes, respectively; N. Hallot for the rock crushing; E. Hanson for sharing her photographs of the trace fossil from Crozon; X. Le Coz for the rock sawing and preparation of thin sections; Y. Lepagnot for mineral separations; F. Polette and L. Guillois for their assistance with U–Pb dating; and the owners of the outcrops for granting access to their private properties. The manuscript benefited from insightful reviews by A. Uchman and an anonymous reviewer, as well as editor S. Jensen. The work of A.L. has been partially supported by the ‘Fondazione Banco di Sardegna’ and by the ‘Regione Autonoma della Sardegna’ (grant numbers F74I19000960007, J81G17000110002). This publication is a contribution to the VIBRIO project of the INSU InterrVie program, and benefitted from financial support from the Observatoire des Sciences de l’Univers de Rennes (OSUR). We are grateful to have had the chance to work with our colleague M.-P. Dabard, who passed away too soon.
Declaration of interest
None.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0016756820001430






