


Introduction
Although the birth of piglets derived from embryos fertilized in vitro has been successfully achieved (Yoshida et al., Reference Yoshida, Mizoguchi, Ishigaki, Kojima and Nagai1993; Wongsrikeao et al., Reference Wongsrikeao, Otoi, Yamasaki, Agung, Taniguchi, Naoi, Shimizu and Nagai2006a), in vitro production (IVP) of porcine embryos commonly results in highly variable blastocyst development rates. Due to that scenario, many media and additives for in vitro maturation (IVM) have been tested (Wang et al., Reference Wang, Abeydeera, Cantley and Day1997; Yoon et al., Reference Yoon, Shin, Park, Roh, Lim, Lee, Hwang and Lee2000; Akaki et al., Reference Akaki, Yoshioka, Noguchi, Hoshi and Funahashi2009). Additionally, difficulties in selecting high-quality cumulus–oocyte complexes (COCs) (Pawlak et al., Reference Pawlak, Pers-Kamczyc, Renska, Kubickova and Lechniak2011) justified the investigation of distinct selection criteria based upon the oocyte's source and morphology (Yoon et al., Reference Yoon, Shin, Park, Roh, Lim, Lee, Hwang and Lee2000; Schoevers et al., Reference Schoevers, Colenbrander and Roelen2007; Kim et al., Reference Kim, You, Hyun, Lee, Lim and Lee2010).
Grown oocytes with greater developmental competence present reduced activity of the glucose-6-phosphatate dehydrogenase (G6PDH) enzyme. As G6PDH degrades the brilliant cresyl blue dye (BCB), BCB staining has been used for oocyte selection, assuming that highly competent oocytes with greater mitochondrial functionality would be stained blue, due to their inability to metabolize BCB (Roca et al., Reference Roca, Martinez, Vazquez and Lucas1998; Wongsrikeao et al., Reference Wongsrikeao, Otoi, Yamasaki, Agung, Taniguchi, Naoi, Shimizu and Nagai2006a; Egerszegi et al., Reference Egerszegi, Alm, Rátky, Heleil, Brüssow and Torner2010). However, BCB-stained oocytes may have compromised embryo development, due to alterations in mRNA and proteins that act in the fertilization process (Kempisty et al., Reference Kempisty, Jackowska, Piotrowska, Antosik, Wozna, Bukowska, Brüssow and Jaskowski2011) and to disturbances in their mitochondrial function (Santos et al., Reference Santos, Sato, Lucia and Iwata2015). Thus, the efficiency of BCB staining to select high-quality oocytes is still controversial.
Dulbecco's phosphate-buffered saline (DMPBS) is the medium most often used for incubation of BCB-stained oocytes prior to IVM. However, as oocytes commonly are incubated for 1 h, the medium's composition may contribute to impair subsequent embryo development (Pawlak et al., Reference Pawlak, Pers-Kamczyc, Renska, Kubickova and Lechniak2011), especially after a second incubation (Wongsrikeao et al., Reference Wongsrikeao, Otoi, Yamasaki, Agung, Taniguchi, Naoi, Shimizu and Nagai2006a; Kempisty et al., Reference Kempisty, Jackowska, Piotrowska, Antosik, Wozna, Bukowska, Brüssow and Jaskowski2011). Therefore, testing a distinct medium with enriched composition may improve the efficiency of oocyte selection, particularly considering the limitations associated with the use of BCB staining. The objective of this study was to evaluate the efficiency of BCB staining for selection of porcine oocytes incubated in DMPBS and in a modified medium called ReproPel.
Material and methods
Media and chemicals
Except when stated otherwise, all media were prepared with Type 1 ultrapure Milli-Q® water (Merck Millipore, São Paulo, SP, Brazil) and with chemicals from Sigma-Aldrich Chemical Company (St. Louis, MO, USA). DMPBS was used as a control medium, consisting of: 137 mM NaCl; 2.7 mM KCl; 8.1 mM Na2HPO4; 1.47 mM KH2PO4; 0.9 mM CaCl2; 0.49 mM MgCl2; 5.5 mM glucose; 0.33 mM C3H3O3 Na; and 50 μg/ml gentamicin.
The ReproPel medium tested in this study was based on the composition of two known chemically defined media: SOFaaci (Holm et al., Reference Holm, Booth, Schmidt, Greve and Callesen1999) and PZM (Yoshioka et al., Reference Yoshioka, Suzuki and Onishi2008). However, ReproPel was enriched with both fructose (Wongsrikeao et al., Reference Wongsrikeao, Otoi, Taniguchi, Karja, Agung, Nii and Nagai2006b) and glycine (Hong & Lee, Reference Hong and Lee2007) and had low sodium concentration (Funahashi et al., Reference Funahashi, Cantley, Stumpf, Terlouw and Day1994). The composition of ReproPel consisted of: 90 mM NaCl; 10 mM KCl; 5 mM NaHCO3; 0.34 mM C6H5Na3O7.2H2O; 0.2 mM C3H3O3Na; 0.35 mM KH2PO4; 0.4 mM MgSO4; 1.71 mM CaCl2.2H2O; 0.5 mM d-fructose; 2.77 mM myo-inositol; 25 mM HEPES; 10 mM glycine; 0.01 mM EDTA; 5.35 mM sodium lactate; 0.004 mM phenol red; 50 μg/ml gentamicin; 20 ml/l of essential amino acids; and 10 ml/l of non-essential amino acids. Both DMPBS and ReproPel were supplemented with 10% (v/v) BSA.
Harvesting of cumulus–oocyte complexes and follicular fluid
Ovaries from prepubertal gilts weighing 100–110 kg were collected in a slaughterhouse and transported to the laboratory in saline/gentamicin solution (40 mg/ml) at 30–35°C. COCs were aspirated from 3–6 mm follicles using a vacuum pump (14 mmHg, digital vacuum pump WTA®). Only oocytes having homogenous ooplasm and compact cumulus oophorus cells were selected. During morphological selection, COCs were kept in porcine follicular fluid clarified by centrifugation. The follicular fluid was obtained from 3–6 mm follicles aspirated from oocytes of slaughtered pubertal gilts and used as an additive for the IVM medium. The follicular fluid was centrifuged at 1500 g for 30 min, filtered through a 0.20 µM membrane and stored at −20°C.
BCB staining
The COCs previously selected according to their morphology (n = 915) were incubated in a 4-well dish at 39°C with an atmosphere of 5% CO2 for 60 min. Then, oocytes were cultured either in DMPBS or ReproPel (400 µl each), both supplemented with 4 mg/ml BSA. In both media, oocytes were either exposed to 13 µM BCB or unexposed (UN) to BCB (negative controls). After evaluation by stereomicroscopy, oocytes exposed to BCB were classified as stained (BCB+) or unstained (BCB−). Thus, six groups were formed: DMPBS/UN; DMPBS/BCB+; DMPBS/BCB−; ReproPel/UN; ReproPel/BCB+; and ReproPel/BCB−.
In vitro maturation (IVM)
In all subsequent assays, oocytes unexposed to BCB were also incubated for 60 min. The remaining oocytes (BCB+ and BCB−) were submitted to IVM for 44 h, under controlled atmosphere with 5% CO2 with maximum humidity at 39°C. The IVM medium was the NCSU-23 (Wang et al., Reference Wang, Abeydeera, Cantley and Day1997), including: 10% (v/v) porcine follicular fluid; 0.727 mM pyruvate; 5 mM hypotaurine; 0.6 mM cysteine; 10 ng/ml EGF; 1.0 mM glutamine; and 25 µM β-mercaptoethanol. During the first 22 h of culture, the medium was supplemented with: 10 IU hCG (Chorulon®, MSD Animal Health, São Paulo, SP, Brazil); 10 IU eCG (Folligon®, MSD Animal Health, São Paulo, SP, Brazil); and 1 mM dibutyryl cAMP (dbcAMP). After IVM, oocytes were evaluated for nuclear maturation and for migration of cortical granules (CG) and submitted to parthenogenetic activation.
Nuclear maturation
After IVM, mechanically denuded oocytes (n = 384) were placed in contact with 30 µg/ml bis-benzimide (Hoechst 33342), in three replicates. Nuclear maturation was determined through epifluorescence microscopy (Olympus BX 51, America Inc., São Paulo, SP, Brazil). Only oocytes at metaphase II (MII) were considered mature (Naito & Toyoda, Reference Naito and Toyoda1991).
Assessment of cortical granules (CG)
The migration of the CG was evaluated as an indicator of cytoplasmic maturation (Romar et al., Reference Romar, Coya, Gadea and Rath2005). Mechanically denuded oocytes were incubated in 10 µg/ml of RNase for 30 min and 10 µg/ml of propidium iodide for 10 min. In order to assess the exact equatorial plane, oocytes were placed between slide and coverslip in DABCO mounting media (23 mg/ml) surrounded by paraffin, to maintain the its three-dimensional structure. A confocal microscope (FluoView 1000, Olympus®, Tokyo, Japan) was used to evaluate the migration of the CG, with nearly 40 oocytes per group, in three replicates (total n = 123). The percent of the cytoplasmic area occupied by the CG was determined through two concentric halos marked in the equatorial area of the oocytes (Fig. 1), using the Image-J® software. Maturation was considered more efficient in oocytes with greater concentration of CG, occupying a lower percent of the cytoplasmic area (Romar et al., Reference Romar, Coya, Gadea and Rath2005).
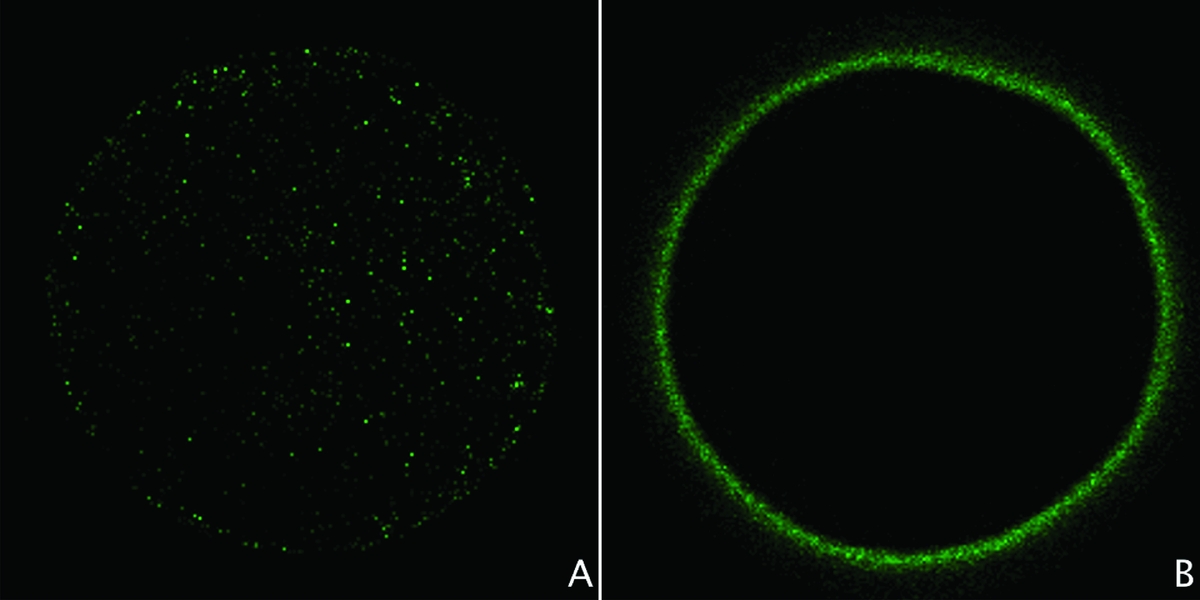
Figure 1 (A, B) Assessment of cortical granules (CG) in porcine oocytes by confocal microscopy (images taken from the equatorial plane).
Determination of genotoxicity
After IVM for 44 h, genotoxicity was evaluated using the comet assay (Tatemoto et al., Reference Tatemoto, Sakurai and Muto2000), to determine the rate of DNA fragmentation of oocytes from DMPBS/BCB+ (n = 175), DMPBS/BCB− (n = 82), ReproPel/BCB+ (n = 257), ReproPel/BCB− (n = 261). Specifically for this assay, oocytes that were not exposed to either tested media and that were kept in porcine follicular fluid during morphological selection (n = 140) were evaluated as a control group. Three replicates were conducted. Initially, COCs were placed in contact with 0.1% hyaluronidase in TCM-199. Then, a 200 µl micropipette was used to remove the cumulus oophorus cells. Oocytes kept in 10 µl PBS were placed in slides containing a 180 µl agarose drop, with low melting point at 50°C (15–20 oocytes per group). After receiving a cover slip, slides were immersed for 24 h in a lysis solution including: 2.5 M NaCl; 100 mM EDTA; 8 mM Tris–HCl; 10% DMSO; and 1% Triton-X. Electrophoresis was done in a dark room in a horizontal cooled vat during 20 min (25 V and 300 Amp). Stained slides were evaluated by optical microscopy at ×100 magnification. The drag formed in the slide determined the level of DNA degradation, which was classified using a score (0–5): 0 meant no fragmentation; and 5 meant maximum fragmentation (Santos et al., Reference Santos, Schiar, Ribeiro, Schwab, Meinerz, Allebrandt, Rocha, Nogueira, Aschner and Barbosa2009).
Embryo development
Denuded oocytes (n = 1,559, with seven replicates) were activated by parthenogenesis (day 0 = D0), as described by Cheng et al. (Reference Cheng, Sun, An, Zhu, Li, Li and Tian2007), with modifications. Oocytes were exposed to 20 µM/ml ionomycin for 5 min in TCM-199 supplemented with 2 mg/ml BSA, and to 2 mM/ml 6-DMAP during 3 h at 39°C. Thereafter, activated oocytes were cultured in four-well dishes, under controlled atmosphere (5% CO2, 5% O2 and 90% N2) at 39°C, under maximum humidity in a bag-system (Vajta et al., Reference Vajta, Holm, Greve and Callesen1997). Subsequent culture (40 oocytes per dish) was done in 400 µl PZM-3 (Yoshioka et al., Reference Yoshioka, Suzuki and Onishi2008), supplemented with 0.2 mM pyruvate, 1 mM glutamine, 5 mM hypotaurine and 3 mg/ml BSA and kept under mineral oil. Cleavage rates were determined at D2. At D4, the medium was supplemented with 10% inactivated fetal bovine serum. Blastocyst development rates were determined at D7.
Statistical analyses
Rates of BCB staining, IVM, cleavage, embryo development and the percent of the area occupied by CG were compared among the tested media through chi-square tests. The DNA fragmentation scores were classified as none (0), mild (1–2), or severe (3–5). The sums of such scores were compared across media through the Kruskal–Wallis analysis of variance for non-parametric data, assuming that lower sums of scores corresponded to greater frequency of oocytes with no DNA fragmentation. All analyses were done with Statistix® (2013).
Results
Among all COCs exposed to BCB, the percent of BCB-stained oocytes was greater (P < 0.05) in ReproPel (86.2%; 288/334) than in DPBS (69.6%; 220/316). Among all evaluated oocytes, 24.0% presented severe DNA fragmentation (Table 1). The frequency of oocytes with undamaged DNA was greater for BCB-stained oocytes in DMPBS than for those in other media (P < 0.05), but similar to that of oocytes incubated in porcine follicular fluid (P > 0.05). Conversely, for oocytes cultured in ReproPel, the frequency of severe DNA fragmentation was greater than for oocytes in other media (P < 0.05).
Table 1 Genotoxicity of swine oocytes stained (+) or unstained (−) with brilliant cresyl blue (BCB) after incubation in distinct media

A.B.C Sum of scores with distinct superscripts differ in the columns by at least P < 0.05.
a.b.c Sum of scores with distinct superscripts differ in the rows by at least P < 0.05.
*None = 0; Mild = 1–2; Severe = 3–5.
Greater MII rates (P < 0.05) were observed for BCB-stained oocytes in both DMPBS and ReproPel and also for unexposed oocytes in ReproPel (Table 2). The percent of cytoplasmic area occupied by CG was lower for unexposed oocytes in ReproPel (P < 0.05) than for unstained oocytes in ReproPel and for unexposed oocytes in DMPBS (Table 2).
Table 2 Nuclear maturation (MII), area of cortical granules (CG) and embryo development for swine oocytes stained (+) or unstained (−) with brilliant cresyl blue (BCB) cultured in distinct media

A,B Rates within each column with distinct superscripts differ by at least P < 0.05.
*Rates were not compared because very few unstained oocytes were available after parthenogenetic activation.
Embryo development rates were compared only for BCB-stained and unexposed oocytes, since very few unstained oocytes were available after parthenogenetic activation. Cleavage rates for stained oocytes were greater (P < 0.05) than for unexposed oocytes in DMPBS (Table 2), but similar to the rates for oocytes in ReproPel (P > 0.05). Blastocyst development rates were greater for oocytes in ReproPel than for those in DMPBS (P < 0.05).
Discussion
The present study was the first one to test a medium with composition more complex than DMPBS as an additional tool for oocyte selection for IVM using BCB staining. This was also the first study to investigate associations between BCB staining and DNA fragmentation by conducting the comet assay without removing the zona pellucida of porcine oocytes exposed to BCB, assuming that denatured DNA fragments migrate outside the cell nucleus during electrophoresis, when stranded DNA is damaged (Berthelot-Ricou et al., Reference Berthelot-Ricou, Perrin, Giorgio, De Meo, Botta and Courbiere2011). As BCB-stained oocytes presented undamaged DNA more frequently when incubated in DMPBS than in ReproPel, it is possible that incubation in ReproPel induced changes in mechanisms of protection against free radicals, which may lead to DNA denaturation, as such mechanisms are dependent upon the G6PDH activity (Takahashi et al., Reference Takahashi, Saka, Takahashi, Kanai, Schultz and Okano1999). Such effect was unexpected, considering that ReproPel was formulated with reduced sodium content and supplemented with pyruvate to increase glutathione's concentration (Funahashi et al., Reference Funahashi, Cantley, Stumpf, Terlouw and Day1994; Reference Funahashi, Koike and Sakai2008). Additionally, the inclusion of fructose in the composition of ReproPel was supposed to help preventing DNA fragmentation (Wongsrikeao et al., Reference Wongsrikeao, Otoi, Taniguchi, Karja, Agung, Nii and Nagai2006b), which apparently did not occur in the present study. Nevertheless, the frequencies of mild DNA fragmentation can be considered fairly high in most treatments, even for oocytes unexposed to BCB conditioned in porcine follicular fluid, which were not incubated for 60 min. Thus, we hypothesize that the high DNA fragmentation observed in this study may not be due to BCB staining or to the additional incubation period, but more likely a result of oxidative stress, which may break the single-stranded DNA, leading to irreparable damages in the double-stranded DNA (Takahashi et al., Reference Takahashi, Saka, Takahashi, Kanai, Schultz and Okano1999). Considering the occurrence of meiosis resumption during IVM and the very intense DNA replication during initial embryo development, DNA breakages may lead to mutations, abnormal gene expression and suppression of subsequent development (Kitagawa et al, Reference Kitagawa, Suzuki, Yoneda and Watanabe2004).
Nuclear maturation was generally lower for unstained oocytes in both tested media, but, in ReproPel, unstained oocytes presented lower MII rate than both stained and unexposed oocytes. Considering that stained oocytes in both media presented similar MII rates, the presence of fructose and pyruvate in ReproPel's composition, which would supposedly aid the capacity of oocytes to progress through meiosis (Funahashi et al., Reference Funahashi, Koike and Sakai2008), apparently did not benefit nuclear maturation. Nonetheless, the area of CG was similar for oocytes in DMPBS, no matter if they were exposed or unexposed to BCB, and generally similar for oocytes cultured in both media (with the exception of unexposed oocytes in ReproPel). Although more BCB-stained oocytes were observed in ReproPel than in DMPBS, the per cent of unstained oocytes was generally low in both media. These results indicate that the efficacy of BCB staining to select viable porcine oocytes is questionable, as efficient oocyte selection may be achieved only using morphological criteria (Pawlak et al., Reference Pawlak, Warzych, Chabowska and Lechniak2014).
Compared with oocytes unexposed to BCB, stained oocytes presented greater cleavage rates in DMPBS, which might suggest that BCB staining could be effective on selecting oocytes. However, the development to blastocyst eliminated such differences between oocytes cultured in DMPBS. Conversely, with incubation in ReproPel, stained and unexposed oocytes achieved similar cleavage rates and greater blastocyst development rates than oocytes in DMPBS. Although that might suggest that ReproPel could be efficient for incubation of porcine oocytes, it could not be recommended due to the increased rate of DNA fragmentation observed in the present study. Future studies could be aimed to refine the composition of ReproPel to minimize oocyte's DNA fragmentation.
In conclusion, after parthenogenetic activation, BCB-stained oocytes incubated in ReproPel presented greater blastocyst development rate compared with BCB-stained oocytes incubated in DMPBS, although nuclear maturation and the area of CG were similar for both such media. However, despite of its enriched composition, ReproPel cannot be recommended for selection of porcine oocytes in vitro due to the high DNA fragmentation. With either media, selection of competent oocytes was not accomplished through BCB staining.
Financial support
This research was funded with a scholarship given to E.C.S. Santos by CAPES.



