




















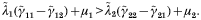
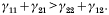










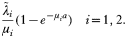


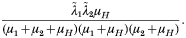



















































Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Jackson, J.A.
Pleass, R.J.
Cable, J.
Bradley, J.E.
and
Tinsley, R.C.
2006.
Heterogenous interspecific interactions in a host–parasite system.
International Journal for Parasitology,
Vol. 36,
Issue. 13,
p.
1341.
PION, S. D. S.
CLARKE, P.
FILIPE, J. A. N.
KAMGNO, J.
GARDON, J.
BASÁÑEZ, M.-G.
and
BOUSSINESQ, M.
2006.
Co-infection withOnchocerca volvulusandLoa loamicrofilariae in central Cameroon: are these two species interacting?.
Parasitology,
Vol. 132,
Issue. 6,
p.
843.
Cattadori, Isabella M
Albert, Réka
and
Boag, Brian
2007.
Variation in host susceptibility and infectiousness generated by co-infection: the myxoma–Trichostrongylus retortaeformiscase in wild rabbits.
Journal of The Royal Society Interface,
Vol. 4,
Issue. 16,
p.
831.
Stear, Michael J
Fitton, Lesley
Innocent, Giles T
Murphy, Lisa
Rennie, Kerry
and
Matthews, Louise
2007.
The dynamic influence of genetic variation on the susceptibility of sheep to gastrointestinal nematode infection.
Journal of The Royal Society Interface,
Vol. 4,
Issue. 16,
p.
767.
Basáñez, María-Gloria
Razali, Karina
Renz, Alfons
and
Kelly, David
2007.
Density-dependent host choice by disease vectors: epidemiological implications of the ideal free distribution.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 101,
Issue. 3,
p.
256.
BRADLEY, J. E.
and
JACKSON, J. A.
2008.
Measuring immune system variation to help understand host-pathogen community dynamics.
Parasitology,
Vol. 135,
Issue. 7,
p.
807.
FENTON, ANDY
2008.
Worms and germs: the population dynamic consequences of microparasite-macroparasite co-infection.
Parasitology,
Vol. 135,
Issue. 13,
p.
1545.
BEHNKE, J. M.
2008.
Structure in parasite component communities in wild rodents: predictability, stability, associations and interactions .... or pure randomness?.
Parasitology,
Vol. 135,
Issue. 7,
p.
751.
BEHNKE, J. M.
EIRA, C.
ROGAN, M.
GILBERT, F. S.
TORRES, J.
MIQUEL, J.
and
LEWIS, J. W.
2009.
Helminth species richness in wild wood mice,Apodemus sylvaticus, is enhanced by the presence of the intestinal nematodeHeligmosomoides polygyrus.
Parasitology,
Vol. 136,
Issue. 7,
p.
793.
Karvonen, Anssi
Seppälä, Otto
and
Tellervo Valtonen, E.
2009.
Host immunization shapes interspecific associations in trematode parasites.
Journal of Animal Ecology,
Vol. 78,
Issue. 5,
p.
945.
Pugliese, Andrea
2010.
Modelling Parasite Transmission and Control.
Vol. 673,
Issue. ,
p.
32.
Fenton, Andy
Viney, Mark E.
and
Lello, Jo
2010.
Detecting interspecific macroparasite interactions from ecological data: patterns and process.
Ecology Letters,
Vol. 13,
Issue. 5,
p.
606.
Cornell, Stephen J.
2010.
Modelling Parasite Transmission and Control.
Vol. 673,
Issue. ,
p.
66.
Basáñez, María-Gloria
McCarthy, James S.
French, Michael D.
Yang, Guo-Jing
Walker, Martin
Gambhir, Manoj
Prichard, Roger K.
Churcher, Thomas S.
and
Zhou, Xiao-Nong
2012.
A Research Agenda for Helminth Diseases of Humans: Modelling for Control and Elimination.
PLoS Neglected Tropical Diseases,
Vol. 6,
Issue. 4,
p.
e1548.
Basáñez, María-Gloria
French, Michael D.
Walker, Martin
and
Churcher, Thomas S.
2012.
Paradigm lost: how parasite control may alter pattern and process in human helminthiases.
Trends in Parasitology,
Vol. 28,
Issue. 4,
p.
161.
Cobey, Sarah
and
Lipsitch, Marc
2013.
Pathogen Diversity and Hidden Regimes of Apparent Competition.
The American Naturalist,
Vol. 181,
Issue. 1,
p.
12.
Yakob, Laith
Williams, Gail M
Gray, Darren J
Halton, Kate
Solon, Juan Antonio
and
Clements, Archie CA
2013.
Slaving and release in co-infection control.
Parasites & Vectors,
Vol. 6,
Issue. 1,
Shrestha, Sourya
Foxman, Betsy
Weinberger, Daniel M.
Steiner, Claudia
Viboud, Cécile
and
Rohani, Pejman
2013.
Identifying the Interaction Between Influenza and Pneumococcal Pneumonia Using Incidence Data.
Science Translational Medicine,
Vol. 5,
Issue. 191,
Yakob, Laith
Soares Magalhães, Ricardo J.
Gray, Darren J.
Milinovich, Gabriel
Wardrop, Nicola
Dunning, Rebecca
Barendregt, Jan
Bieri, Franziska
Williams, Gail M.
and
Clements, Archie C.A.
2014.
Modelling parasite aggregation: disentangling statistical and ecological approaches.
International Journal for Parasitology,
Vol. 44,
Issue. 6,
p.
339.
Fowler, A. C.
and
Déirdre Hollingsworth, T.
2016.
The Dynamics of Ascaris lumbricoides Infections.
Bulletin of Mathematical Biology,
Vol. 78,
Issue. 4,
p.
815.