Ornithosuchidae is a group of terrestrial carnivorous archosaurs reported from the Upper Triassic of Argentina and Scotland. The name was coined by von Huene in 1908 and currently comprises three species: Ornithosuchus longidens (Huxley Reference Huxley1877), Riojasuchus tenuisceps Bonaparte, Reference Bonaparte1969, and Venaticosuchus rusconii Bonaparte, Reference Bonaparte1970.
Ornithosuchus longidens was the first ornithosuchid to be discovered, and is the only representative of the clade from the northern hemisphere. It is represented by 11 specimens, including juveniles and adults, from the Lossiemouth Sandstone Formation, Moray, Scotland (Walker Reference Walker1964). During the last century, the taxonomic status of Ornithosuchus underwent several changes, including its synonymy with Dasygnathoides (=“Dasygnathus”) longidens, a species based on very fragmentary materials. In addition, three species (Ornithosuchus longidens, Ornithosuchus woodwardi and Ornithosuchus taylori) were named, which were later synonymised by Walker (Reference Walker1964), resulting in the nomenclatural combination of Dasygnathoides longidens (Huxley Reference Huxley1877) and Ornithosuchus woodwardi Newton, Reference Newton1894, into the current species Ornithosuchus longidens (Huxley Reference Huxley1877).
The purpose of this work is to revisit the taxonomy of Ornithosuchus and test the synonymy between Ornithosuchus woodwardi Newton, Reference Newton1894 and Dasygnathoides (=“Dasygnathus”) longidens (Huxley Reference Huxley1877).
Institutional abbreviations: BP, Evolutionary Studies Institute (formerly Bernard Price Institute for Palaeontological Research), University of the Witswatersrand, Johannesburg, South Africa; CAMZM, University Museum of Zoology, Cambridge, UK; ELGNM, Elgin Museum, Elgin, UK; GSM, Geological Survey Museum, London, UK; MANCH, Manchester Museum, Manchester, UK; NHMUK PV R, Natural History Museum, London, UK; PVL, Paleontología de Vertebrados, Instituto Miguel Lillo, Tucumán, Argentina; RC, Rubidge Collection, Wellwood, Graaff-Reinet, South Africa; SMNS, Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany.
1. Taxonomic background
The holotype of Ornithosuchus (= “Dasygnathus”, = Dasygnathoides) longidens (Huxley Reference Huxley1877) is a natural mould of the medial surface of a right maxilla (ELGNM 1) found at the Findrassie Quarry near Elgin, Lossiemouth Sandstone Formation, Scotland. The moulds of a large isolated right pterygoid (ELGNM 15) and a possible detached articular (ELGNM 29) were also found associated in the same quarry and referred to the same individual (Walker Reference Walker1961, Reference Walker1964).
Huxley (Reference Huxley1859) originally interpreted ELGNM 1 as part of the lower jaw of Stagonolepis robertsoni, because there was no other reptile known from the Findrassie Quarry that matched its tooth implantation pattern. Subsequently, Huxley (Reference Huxley1869, p. 424) changed his previous interpretation and clarified that he was convinced that “the mandible with long curved teeth which I supposed might belong to Stagonolepis certainly appertains to some other animal”. Later, Huxley (Reference Huxley1877) considered ELGNM 1 “a dentigerous bone, insertae sedis” and found it convenient to establish a new name for the animal to which this “jaw” belonged, namely “Dasygnathus” longidens.
In Reference Newton1894, Newton published a study of two new genera and species of reptiles from the Spynie Quarry, Lossiemouth Sandstone Formation (formerly Elgin Sandstones), near Findrassie Quarry, where “Dasygnathus” longidens was discovered. One of these new taxa was an almost complete individual that Newton named Ornithosuchus woodwardi, in honour of Arthur Smith Woodward, who allowed him to describe this new specimen. The specimen consists of a skull (NHMUK PV R2409) and postcranium (NHMUK PV R2410) that were originally in the same slab, but were split in two to prepare the specimen. The skull of this specimen was not exposed when it was found in the quarry, resulting in its good three-dimensional preservation, unlike the rest of the skeleton, which is mostly eroded and preserved as moulds. During its preparation at the Natural History Museum (=British Museum), the skull was uncovered and the skeleton moulds were cleaned of bone fragments to make casts (Newton Reference Newton1894). NHMUK PV R2409 is currently the most complete and best preserved skull known for Ornithosuchus.
In 1903, Boulenger described two new reptilian specimens from the West Quarry of Lossiemouth (NHMUK PV R3142, NHMUK PV R3143). He recognised their resemblance to Ornithosuchus woodwardi, but these specimens are much larger (about 2 m in total body length) than the holotype of Ornithosuchus woodwardi (approximately 80 cm in length). He discussed whether they were ontogenetically older specimens of Ornithosuchus woodwardi or a different species, but finally favoured the former idea (Boulenger Reference Boulenger1903).
In 1913, Broom reanalysed the species Ornithosuchus woodwardi, adding novel information given by the discovery of the Triassic archosauriform Euparkeria capensis, proposed as a relative of Ornithosuchus woodwardi by him. Broom (Reference Broom1913) proposed that the larger specimens (NHMUK PV R3142, NHMUK PV R3143) described by Boulenger (Reference Boulenger1903) represented a new species, based on which he erected Ornithosuchus taylori, in honour of William Taylor, the discoverer of the specimens. The difference between the two species was based only on their size and robustness (Broom Reference Broom1913).
In Reference Kuhn1961, Kuhn realised that the generic name Dasygnathus, which was the name that Huxley (Reference Huxley1877) had assigned to ELGNM 1, was pre-occupied by an Australian dynastid coleopteran (Macleay Reference Macleay1819; Bisby et al. Reference Bisby, Roskov, Orrell, Nicolson, Paglinawan, Bailly, Kirk, Bourgoin, Baillargeon and Ouvrard2011), and he proposed the replacement name Dasygnathoides, resulting in the new combination Dasygnathoides longidens (Kuhn Reference Kuhn1961).
In Reference Walker1961, Walker reanalysed ELGNM 1 in the context of a detailed osteology of the aetosaur Stagonolepis robertsoni, because this specimen was once considered to belong to that species. He recognised that the material was not a lower jaw, but rather the mould of a left maxilla; although Walker (Reference Walker1961) at that time mistakenly interpreted it as the impression of its lateral surface.
In Reference Walker1964, Walker re-examined all specimens assigned to the genus Ornithosuchus and the specimen referred to Dasygnathoides (= “Dasygnathus”) longidens. Walker (Reference Walker1964) recognised that the size and robustness differences between Ornithosuchus woodwardi and Ornithosuchus taylori, indicated by Broom (Reference Broom1913), were insufficient to separate them as two species. Furthermore, his thorough study, re-preparation and reinterpretation of several elements of some specimens of Ornithosuchus allowed the identification of more cranial features shared amongst all specimens of Ornithosuchus woodwardi and Ornithosuchus taylori. He also identified minor differences amongst individuals, which could be explained by their different ontogenetic stages, combined with intraspecific variation and deformation (Walker Reference Walker1964); concluding that the two species were synonymous. In the same study, Walker (Reference Walker1964) reinterpreted the holotype of Dasygnathoides (=“Dasygnathus”) longidens as the mould of the medial surface of a right maxilla based on the presence of interdental plates and a palatal process. He also considered Dasygnathoides (=“Dasygnathus”) longidens to be indistinguishable from Ornithosuchus woodwardi. This interpretation was justified by the shared presence of a “rear forking” on the posterior end of the maxillae, the presence of only nine maxillary teeth, and the association of the right pterygoid ELGNM 15, which Walker (Reference Walker1964) also considered similar to that of Ornithosuchus woodwardi. Therefore, Walker (Reference Walker1964) assigned the fossils previously referred to Dasygnathoides (=“Dasygnathus”) longidens (ELGNM 1 and ELGNM 15) to the genus Ornithosuchus, and proposed the nomenclatural combination Ornithosuchus longidens (Huxley Reference Huxley1877), which has been used since. Accordingly, Ornithosuchus woodwardi and Ornithosuchus taylori were regarded by Walker (Reference Walker1964) as subjective junior synonyms of Ornithosuchus longidens.
2. Studied materials
For this study, the following materials have been studied first-hand: ELGNM 1, ELGNM 15, ELGNM 29, unnumbered (formerly E1978-570.4, E1978.716A–B); NHMUK PV R2409-R2410, NHMUK PV R3142, NHMUK PV R3143, NHMUK PV R3149, NHMUK PV R3152, NHMUK PV R3561, NHMUK PV R3562, NHMUK PV R3916.
3. Reassessment of Dasygnathoides longidens and its synonymy with Ornithosuchus woodwardi
According to Walker (Reference Walker1961, Reference Walker1964), Dasygnathoides (=“Dasygnathus”) longidens (Huxley Reference Huxley1877) is represented by natural moulds of right maxilla and pterygoid, preserved on fine-grained sandstone slabs (ELGNM 1 and ELGNM 15 respectively). These specimens have other incomplete moulds/slabs associated to them: a partial vertebra (ELGNM 1); a phalanx and a haemal arch (ELGNM 15); an articular (ELGNM 29); and an osteoderm (ELGNM unnumbered).
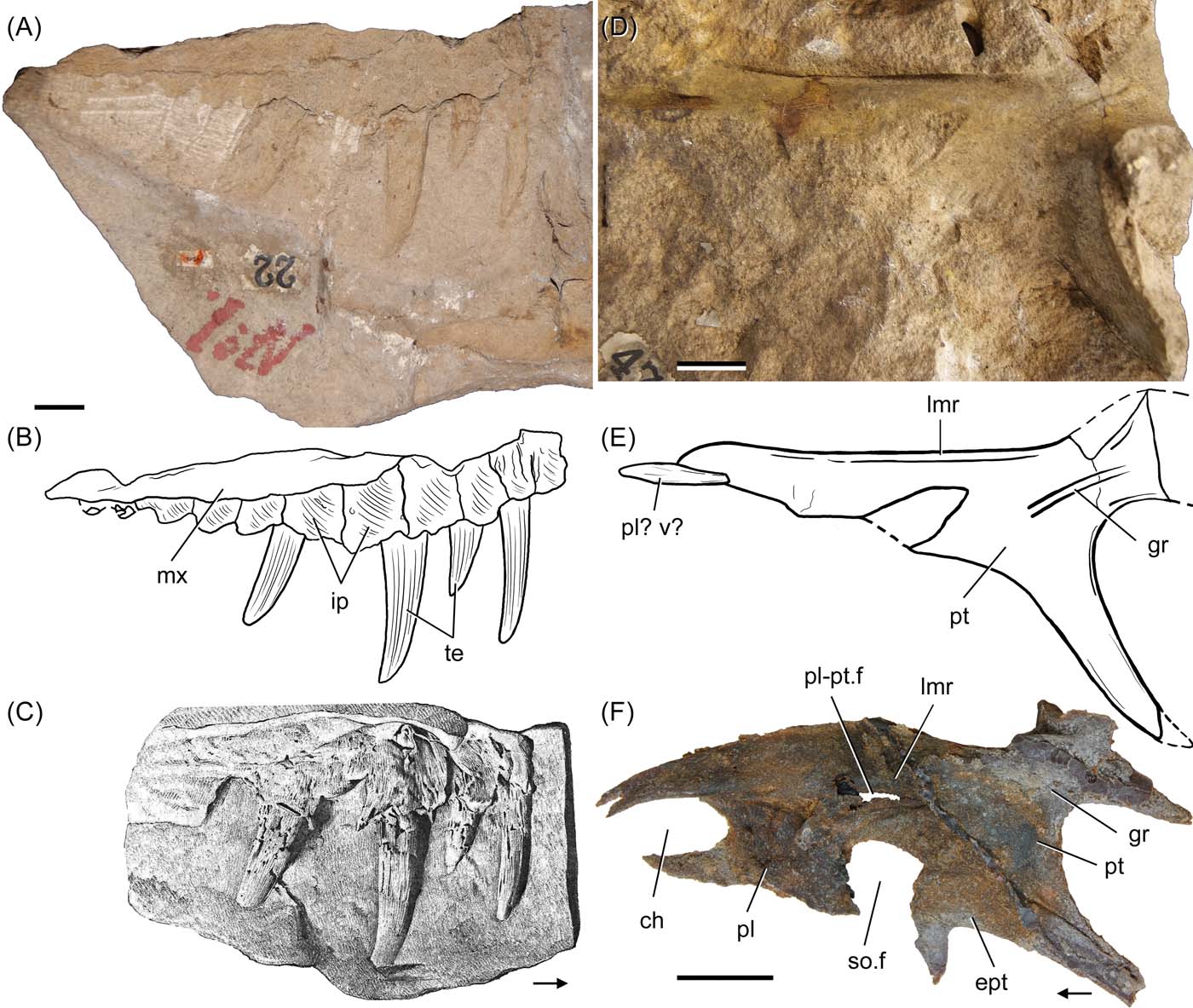
ELGNM 1 is the natural mould of the medial side of the ventral half of a right maxilla, with a total preserved length of 195 mm (Figs 1A, 2A–C). In this impression, four teeth and nine pentagonal-shaped interdental plates are recognised. All four teeth were in their natural position, although probably slightly displaced out of their alveoli. Tooth crowns were posteriorly curved and had sharp edges (Fig. 1A: te), resembling the shape seen in most carnivorous archosauriforms (e.g., Erythrosuchus africanus: NHMUK PV R3592, BP/1/5207; Riojasuchus tenuisceps: PVL 3828; Saurosuchus galilei: PVL 2062; Gracilisuchus stipanicicorum: PVL 4597). The mould of the longest tooth is 53.5 mm long and that of the shortest one is 29.5 mm long. The interdental plates were separated from one another and probably formed the medial wall of the alveoli. These plates were pentagonal to triangular in shape, had a rugose medial surface, and were separated from the rest of the maxilla by an anterodorsally curved shelf (Fig. 2B: ip). This is a widespread condition that resembles that of most archosauriforms (e.g., Erythrosuchus africanus: NHMUK PV R3592; Asperoris mnyama: NHMUK PV R36615; Saurosuchus galilei: PVL 2062; Batrachotomus kupferzellensis: SMNS 52970; Gracilisuchus stipanicicorum: PVL 4216). The ascending process is not preserved and the palatal process that Walker (Reference Walker1964) identified in ELGNM 1 cannot be identified on the specimen. As it is presently preserved, the partial mould of a vertebra is also seen in ELGNM 1, which can be identified as a dorsal or anterior caudal vertebra because of the well-developed, laterally oriented transverse processes. A possible base of the neural spine can also be recognised, but no other diagnostic feature can be seen on this poorly preserved mould (Fig. 1A: tp, ve).

Figure 1 (A) ELGNM 1 (holotype), mould of partial maxilla and vertebra; (B) ELGNM 15, moulds of pterygoid, vertebra, haemal arch, phalanx; (C) mould of the ventral surface of an osteoderm, ELGNM unnumbered; (D) mould of indeterminate element, ELGNM 29; (E) cast of ELGNM 29; and (F) mould of indeterminate ulna and radius. Abbreviations: ha = haemal arch; ip = interdental plates; lmr = longitudinal medial ridge; mx = maxilla; ol = olecranon; ph = phalanx, pl? v? = possible palatine or vomer; pt = pterygoid; ra = radius; te = teeth; tp = transverse process; ul = ulna; ve = vertebra. Scale bars = 20 mm.
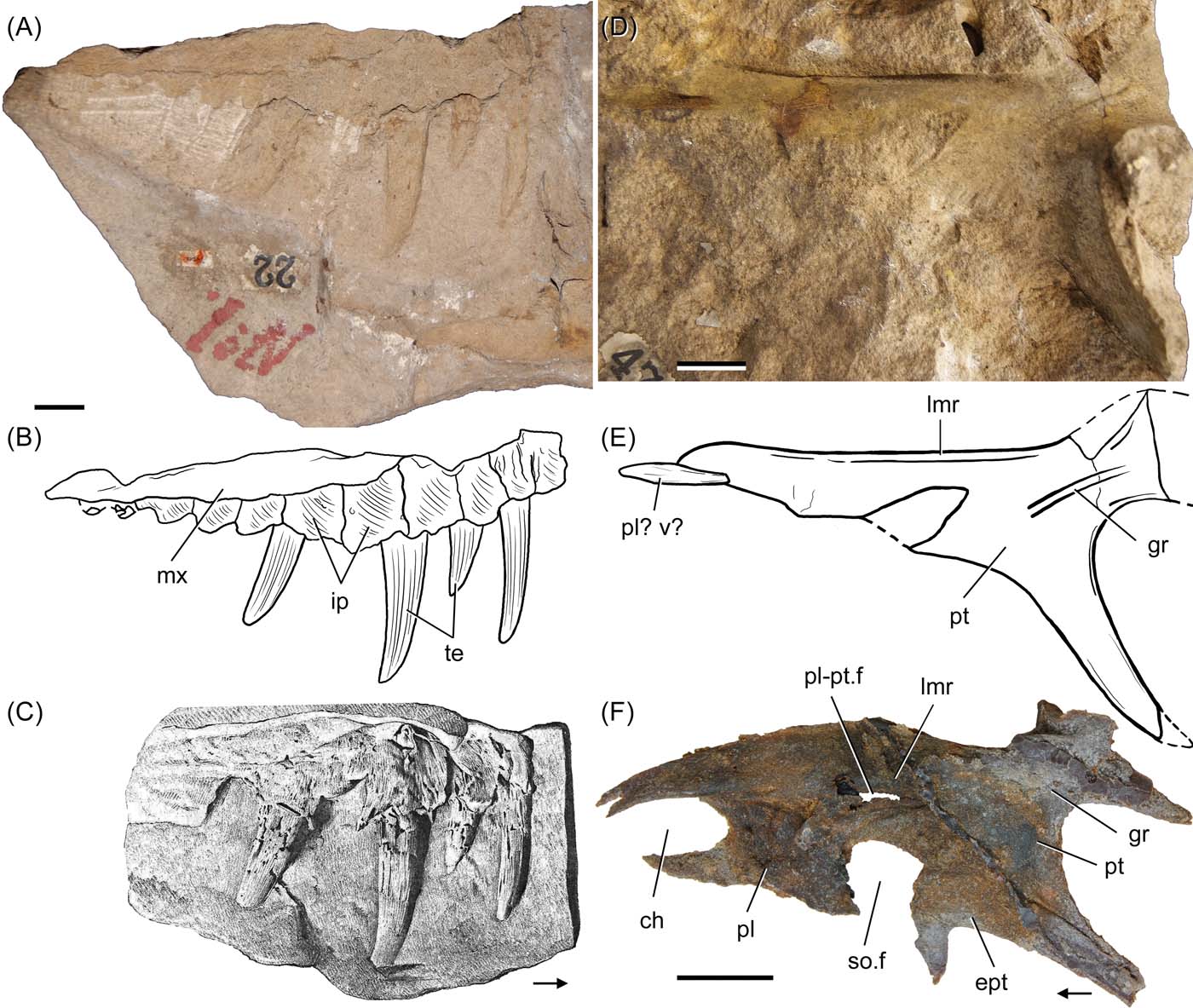
Figure 2 (A) Partial mould of the medial surface of the right maxilla of ELGNM 1, with (B) its interpretative drawing and (C) the original drawing of the cast of ELGNM 1 from Huxley Reference Huxley1877; (D) mould of the dorsal surface of the right pterygoid of ELGNM 15, with (E) its interpretative drawing; (F) cast of right pterygoid of Ornithosuchus woodwardi NHMUK PV R3143, in dorsal view. Abbreviations: ch = choana; ept = ectopterygoid; gr = groove; ip = interdental plates; lmr = longitudinal medial ridge; mx = maxilla; pl = palatine; pl–pt.f = palatine–pterygoid fenestra; pl? v? = possible palatine or vomer; pt = pterygoid; so.f = suborbital fenestra; te = teeth. Arrows indicate anterior direction. Scale bars = 20 mm.
Walker (Reference Walker1964) compared ELGNM 1 with NHMUK PV R3143, and believed that the posterior end of both maxillae had a “rear forking”, considered as a unique feature shared by Dasygnathoides longidens and Ornithosuchus woodwardi. However, the “forking structure” of ELGNM 1 does not correspond to that seen near the posteroventral margin of the maxilla of NHMUK PV R3143. In ELGNM 1, the structure appears as some kind of a ventral projection on the alveolar margin of the maxilla, probably corresponding to the impression of the last preserved interdental plate. On the other hand, the “forking process” of NHMUK PV R3143 is located on the lateral surface of the posterior process of the maxilla and does not project ventrally. Moreover, this “rear forking” of NHMUK PV R3143 cannot be identified on any other specimen of Ornithosuchus (NHMUK PV R2409, R3149, R3562) and is probably an artefact related to the sediment covering the posterior process of the maxilla.
ELGNM 15 includes the natural mould of the dorsal surface of a large right pterygoid, connected to what seems to be a small part of the anteromedial end of the palatine or posterior end of the vomer (Figs. 1B, 2D, E: pt, pl? v?). It is 164 mm long, smooth and convex on its surface. The anterior ramus of the pterygoid is transversely narrow and tapers anteriorly, similar to that of pseudosuchians in general (including phytosaurs), but differing from the broader anterior ramus of the pterygoids of erythrosuchids, proterosuchids, proterochampsids, doswelliids and dinosauriforms (Liu et al. Reference Liu, Butler, Sullivan and Ezcurra2015; Ezcurra Reference Ezcurra2016). The medial margin of the pterygoid was thick, leaving a deep furrow on the mould. The lateral margin of the pterygoid mould is clearly recognised, but is slightly broken on the area of the border of the suborbital fenestra. The quadrate ramus of the pterygoid was almost completely preserved, only lacking its distal tip; it was almost as wide as long and dorsally curved. It also bears a groove extending anterolaterally (Fig. 2E: gr), as seen in the dorsal surface of the base of this process in several other archosauriform pterygoids (e.g., Proterosuchus fergusi: RC 846; Ornithosuchus woodwardi: NHMUK PV R3143; Saurosuchus galilei: PVL 2062). The transverse ramus of the pterygoid was complete and its main axis was posterolaterally oriented, with a ventral component.
When comparing the pterygoid of ELGNM 15 with those of NHMUK PV R2409 and NHMUK PV R3143, the assignment of the former to Ornithosuchus woodwardi is not straightforward. The palatine-pterygoid fenestra, which is a synapomorphy of Ornithosuchidae, is placed adjacent to the longitudinal medial ridge of the anterior ramus of the pterygoid in Ornithosuchus woodwardi (NHMUK PV R2409, NHMUK PV R3143). By contrast, the bone surface immediately lateral to the homologous ridge in ELGNM 15 is complete and lacks such an opening. The subtriangular broken area in the anterior ramus of ELGNM 15 is placed more laterally than expected for the palatine–pterygoid fenestra (Fig. 2E–F). Therefore, the morphology of the pterygoid referred to Dasygnathoides longidens provides evidence against the synonymy with Ornithosuchus woodwardi as proposed by Walker (Reference Walker1964, p. 64).
ELGNM 29 is an indeterminate element that Walker (Reference Walker1964) interpreted as the mould of the medial surface of a left articular (Fig. 1D, E). The bone that left this natural impression was 44.5 mm long and 20 mm high; its surface was smooth, and had two slightly concave areas separated from one another by an oblique change in slope. These concavities do not correspond in shape or position to the glenoid fossa and medial surface of the articular bone of archosaurs in general. In addition, there is no evidence of a retroarticular process or medial processes in this element. Due to the lack of unambiguous evidence supporting the identification of ELGNM 29 as an articular, the bone is considered indeterminate.
Two other isolated elements have been referred to Dasygnathoides longidens. E1978-570.4, identified by Walker (Reference Walker1964) as “ELGNM unnumbered”, is a mould previously interpreted as that of an isolated right nasal (Walker Reference Walker1964) (Fig. 1C). This mould is 36.5 mm long, subrectangular, supposedly anteroposteriorly elongated, smooth and convex, with one of its sides steeper than the other (Fig. 1C). Its anterior margin has a subtle, symmetric concavity that was interpreted by Walker (Reference Walker1964) as the margin of the external naris, but it does not correspond in shape to the other nasals known for Ornithosuchus woodwardi (NHMUK PV R2409). This element better matches the shape of the ventral surface of an osteoderm, resembling the anteroposteriorly elongated dermal elements of Ornithosuchus (NHMUK PV R3142) and several other archosaurs. Yet, it differs from the transversely elongated or square-shaped paramedian or lateral osteoderms of aetosaurs (e.g., Stagonolepis robertsoni: ELGNM 27, ELGNM 40). E1978-716A–B is the mould of an articulated radius and ulna and its counterpart, which were found amongst the holotype of Dasygnathoides longidens at the Elgin Museum during this study (Fig. 1F). The radius and ulna were very slender, with a total length of 31 mm and 36 mm, respectively. The moulds are very simple, but the ulna possesses a distinctly well proximally developed olecranon (Fig. 1F: ra, ol, ul). The presence of a well-developed olecranon differs from the condition seen in two of the small Elgin diapsids, namely Scleromachlus taylori (Benton Reference Benton1999) and Saltopus elginensis (Benton & Walker Reference Benton and Walker2011), which apparently lack such a structure. The morphology of the proximal end of the ulna is unknown in the other small diapsid of the Elgin assemblage, Erpetosuchus walkeri, and the morphology of the ulna is unknown in the other known erpetosuchid species known so far, i.e., Parringtonia gracilis from the late Middle Triassic of Tanzania (NHMUK PV R3139).
E1978-570.4 does not have the original red and black numbers that identify the materials of Dasygnathoides longidens, but its label corresponds to the same old numbering sequence as the other specimens referred to the taxon (e.g., ELGNM 1 = E1978-570.1, ELGNM 15 = E1978-570.2). On the contrary, E1978-716A–B does not match that numbering sequence. Furthermore, the size of the ulna and radius does not correspond to that of the maxilla of Dasygnathoides longidens, which is comparatively much larger. Therefore, that specimen cannot be interpreted as part of the same individual as ELGNM 1 and ELGNM 15 (which compares in size to ELGNM 1). The absence of diagnostic features hampers a precise taxonomic identification of this specimen.
4. Affinities of Dasygnathoides longidens
No autapomorphies of Ornithosuchus woodwardi or synapomorphies shared by ornithosuchids can be recognised in the material assigned to Dasygnathoides longidens (e.g., palatine–pterygoid fenestra) and, as a consequence, its previous assignment to this group has no support. ELGNM 1 can be interpreted as an archosauriform, based on the presence of interdental plates and a probable thecodont tooth implantation (because the teeth are not fused to the interdental plates). In addition, the interdental plates of ELGNM 1 differ from those of proterosuchids and some erythrosuchids, because these taxa have an ankylothecodont tooth implantation (Nesbitt Reference Nesbitt2011; Reference EzcurraEzcurra 2016). It also differs from erythrosuchids, because its interdental plates are higher than anteroposteriorly long, as occurs in Euparkeria capensis (CAMZM T692) and other eucrocopod archosauriforms in general (sensu Ezcurra Reference Ezcurra2016). Thus, this feature indicates that Dasygnathoides longidens could represent a large predatory eucrocopod archosauriform (e.g., Asperoris mnyama, Batrachotomus kupferzellensis, Herrerasaurus ischigualastensis). In particular, the tooth count and morphology of ELGNM 1 is clearly different from those of aetosaurs (e.g., Stagonolepis robertsoni), erpetosuchids (e.g., Erpetosuchus granti), silesaurids (e.g., Silesaurus opolensis), sauropodomorphs (e.g., Eoraptor lunensis) or ornithischians (e.g., Lesothosaurus diagnosticus, Heterodontosaurus tucki).
Considering the information provided by the pterygoid, the presence of a transversely narrow anterior ramus indicates that Dasygnathoides longides could be a pseudosuchian (Liu et al. Reference Liu, Butler, Sullivan and Ezcurra2015; Ezcurra Reference Ezcurra2016), excluding ornithodiran (e.g., Scleromochlus taylori, Saltopus elginensis) and non-archosaurian affinities.
Dasygnathoides and Dasygnathus longidens are here designated as nomina dubia, because they are based on undiagnostic fragmentary material that does not have autapomorphies or a unique combination of character-states. As a result, the revision conducted here has the following taxonomic implications.
5. Systematic palaeontology
Archosauria Cope, Reference Cope1871 sensu Gauthier & Padian, Reference Gauthier, Padian, Hecht, Ostrom, Viohl and Wellnhofer1985
Pseudosuchia Zittel, Reference Zittel1887–1890 sensu Gauthier & Padian, Reference Gauthier, Padian, Hecht, Ostrom, Viohl and Wellnhofer1985
“Dasygnathoides longidens” (Huxley Reference Huxley1877)
=Dasygnathus longidens Huxley, Reference Huxley1877 non Macleay, Reference Macleay1819
=Ornithosuchus longidens (Huxley Reference Huxley1877) sensu Walker, Reference Walker1964
Type specimen. ELGNM 1, natural mould of the medial surface of a right maxilla.
The following specimens were found associated to ELGNM 1 and are considered to be part of the same individual: ELGNM 15 (right pterygoid); ELGNM 29 (indeterminate bone); unnumbered (osteoderm) (Walker Reference Walker1961, Reference Walker1964).
Status. Nomen dubium.
Ornithosuchidae Huene, Reference Huene1908 sensu Sereno, Reference Sereno1991
Ornithosuchus Newton, Reference Newton1894
Ornithosuchus woodwardi Newton, Reference Newton1894
= Ornithosuchus taylori Broom, Reference Broom1913 junior synonym of Ornithosuchus woodwardi
Type specimen. NHMUK PV R2409–2410. Almost complete skull and postcranium belonging to a small-sized individual (see Walker Reference Walker1964).
Referred specimens. NHMUK PV R3142, NHMUK PV R3143, NHMUK PV R3149, NHMUK PV R3561, NHMUK PV R3562, NHMUK PV R3916, NHMUK PV R8170; GSM 91072–78, GSM 91080–81, GSM 91085–86; MANCH 8271 (see Walker Reference Walker1964).
Status. Valid.
Emended diagnosis. Small pseudosuchian that can be distinguished from other archosauriforms, based on the following unique combination of character-states (autapomorphies marked with an asterisk): subtle ornamentation on skull roof; downturned premaxilla not extending ventrally beyond the ventral-most portion of the alveolar margin of the maxilla; postorbital with strong central horizontal crest; jugal with longitudinal ridge on the lateral surface above the ventral margin of the bone; palatine–pterygoid fenestra oval and strongly anteroposteriorly elongated, being approximately three times narrower than the suborbital fenestra*; ventral margin of the posterior third of the lower jaw concave and placed dorsally to the rest of the hemimandible in lateral view*; posterior surangular foramen positioned near surangular–angular suture; anterior spur on the neural spine of anterior to middle caudal vertebrae; femur with longitudinal groove on its proximal surface; and metatarsal V without hooked proximal end.
6. Discussion and conclusion
Even though the interpretation of “Dasygnathoides longidens” has been changed, and it is here rejected as a synonym of Ornithosuchus woodwardi, it is important to note that this taxonomic modification does not affect the current character scorings of Ornithosuchus in most recent phylogenies (Brusatte et al. Reference Brusatte, Benton, Desojo and Langer2010, Nesbitt Reference Nesbitt2011, Baczko et al. Reference Baczko, Desojo and Pol2014; Butler et al. Reference Butler, Sullivan, Ezcurra, Liu, Lecuona and Sookias2014; Ezcurra Reference Ezcurra2016), which are mostly based on NHMUK PV R2409–2410.
The diagnosis of Ornithosuchus woodwardi was here emended after rejecting the mentioned synonymy. Sereno (Reference Sereno1991) was the first to provide a diagnosis for “Ornithosuchus longidens” (extended by monotypy to the genus Ornithosuchus) and more recent authors followed this diagnosis without changes (e.g., Nesbitt Reference Nesbitt2011, Ezcurra Reference Ezcurra2016). That autapomorphy list includes two cranial character states, one of which concerns the rear forking of the maxilla, described as follows (Sereno Reference Sereno1991, p. 12): “Maxilla with free posterior prong: An unusual prong-shaped process of the maxilla projects posteriorly from the alveolar margin and is present in the holotype specimen (Elgin Museum ELGNM 1 R; Walker, Reference Walker1964, p. 64).” On reassessing ELGNM 1, we believe that this feature was misinterpreted (see description above), and is, therefore, not diagnostic for Ornithosuchus. Other cranial autapomorphies proposed by Sereno (Reference Sereno1991) cannot be seen in ELGNM 1 because they refer to elements not preserved in this specimen (i.e., skull roof ornamentation, horizontal crest on the postorbital). Therefore, ELGNM 1 cannot be assigned with certainty to Ornithosuchus.
ELGNM 1 is the largest specimen referred to Ornithosuchidae but, under the taxonomic interpretation proposed here, the upper size limit of ornithosuchids is reduced by approximately 40 % (Fig. 3). The shortest skull is represented by the referred specimen of Ornithosuchus woodwardi NHMUK PV R8170 (= 50 mm) and the largest is the referred specimen of Riojasuchus tenuisceps PVL 3828 (= 259 mm). The skull length of the Argentinean ornithosuchids, Riojasuchus tenuisceps and Venaticosuchus rusconii, currently closely approaches the maximum skull size sampled for Ornithosuchus woodwardi (Table 1). The total body length previously estimated for the Scottish ornithosuchid was 4 m (Walker Reference Walker1964), but with the new interpretation it is reduced to 2.2 m, resembling the total body size of the known Argentinean ornithosuchids (Baczko & Ezcurra Reference Baczko and Ezcurra2013).

Figure 3 Histogram showing the frequency of specimens along a distribution of total skull lengths measured or estimated for Ornithosuchus woodwardi (light grey), Riojasuchus tenuisceps and Venaticosuchus rusconii (dark grey), and “Dasygnathoides longidens” (ELGNM 1) (black).
Table 1 Total skull lengths used in the histogram of Figure 3.

Measurements (in mm) for Ornithosuchus woodwardi and ELGNM 1 taken from Walker (Reference Walker1964); those for Riojasuchus tenuisceps from Baczko & Desojo (Reference Baczko and Desojo2016).
* denotes estimated measurement.
In conclusion, the pseudosuchian represented by the holotype of “Dasygnathoides longidens” (ELGNM 1), plus the referred pterygoid ELGNM 15, does not belong to any of the known archosauriforms sampled from the reptiliferous Lossiemouth Sandstone Formation; namely, Stagonolepis robertsoni, Erpetosuchus granti, Scleromochlus taylori, Saltopus elginensis and Ornithosuchus woodwardi. ELGNM 1 represents the largest predatory tetrapod so far recorded in the Triassic rocks of Scotland.
7. Acknowledgements
We are indebted to D. J. Gower, R. J. Butler, F. Holwerda and, especially, M. J. Benton for their kindness searching for Victorian papers about the Elgin fossil fauna. We thank L. Steel and S. Beardmore for allowing us to study the Elgin specimens under their care, and the following curators and collection managers who allowed the study of comparative specimens: J. Powell, B. Rubidge, F. Abdala, B. Zipfel, J. Choiniere, and R. Schoch. We appreciate the enlightening feedback given by J. B. Desojo, D. E. Perez, M. J. Álvarez, and M. B. Santelli. We are grateful for helpful comments and suggestions of the reviewers M. C. Langer and S. L. Brusatte, and for the editorial work of V. Hammond.