


INTRODUCTION
Excavation History
Friesack 4 is one of the most important Early Mesolithic sites in Northern Europe. It was situated on a small rise at a lakeshore in the Rhinluch fen landscape, in the Elbe-Oder ice-margin valley, ca. 50 km west of Berlin (Scholz Reference Scholz1962: 64; Figure 1). After its discovery in 1910, it was excavated in 1916–1925 by M. Schneider (Reference Schneider1932). In 1940, H. Reinerth demonstrated that stratigraphically distinct undisturbed layers were preserved. Intensive amelioration accompanied by groundwater level reduction led to renewed excavations by B. Gramsch, in 1977–1989 and 1998. In 2000–2001, S. Wenzel (Reference Wenzel2002) excavated some remaining areas (Gramsch Reference Gramsch2002, Reference Gramsch2016).
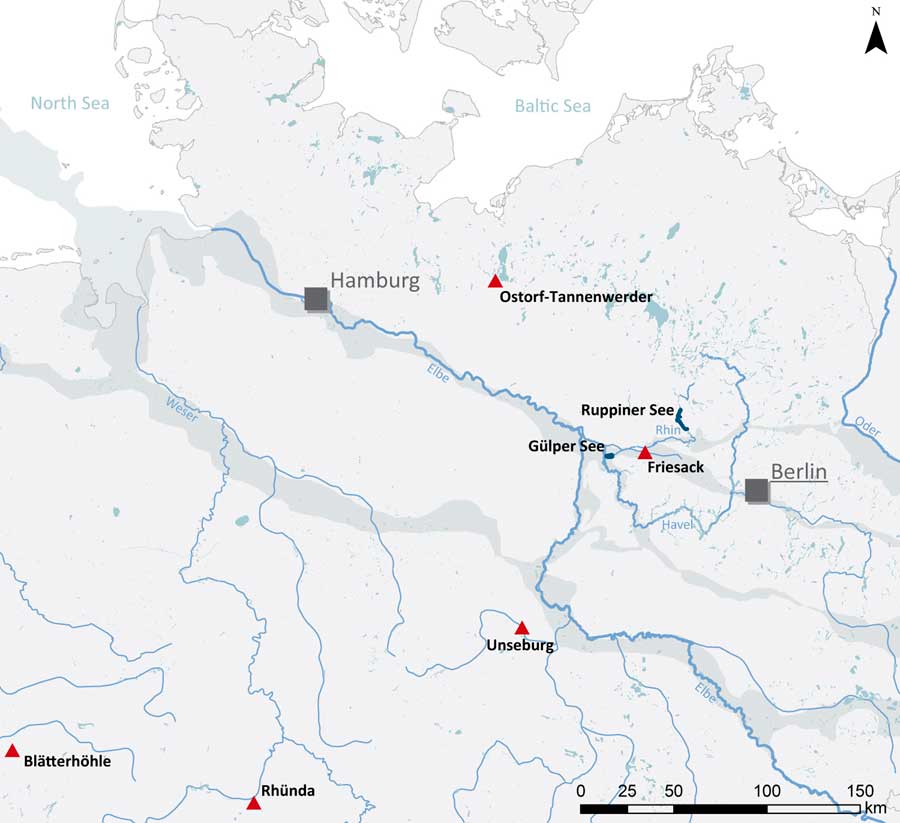
Figure 1 Sites and lakes mentioned in the text are highlighted. Rivers and lakes are depicted in blue. Ice margin valleys are indicated by grey shading. (Please see electronic version for color figures; data source: www.eea.europa.eu).
Although historic sand quarrying had destroyed much of the dryland settlement, a well-preserved assemblage, including organic remains, was recovered during Gramsch’s excavations of the former lakeshore area. The stratigraphy here consisted of different bands of sand that followed the slope of the underlying glacial sands, gyttja and sandy gyttja or humic sands (Kobusiewicz and Kabaciński Reference Kobusiewicz and Kabaciński1993: 14–17; Gramsch Reference Gramsch2002: 60; Groß Reference Groß2017: 63). This layering formed through the erosion of the sandy dryland settlement area during occupation episodes, which interrupted the continuous deposition of gyttja in the adjacent shore area. It was thus possible to distinguish more than 100 different Mesolithic occupations, which were grouped into chronologically sequential Complexes I to IV, and subsequent Neolithic occupation episodes, grouped into Complex V (Gramsch Reference Gramsch2002: 61–63; Reference Gramsch2016: 16–19). Radiometric 14C dates on 87 bulk charcoal samples date the Mesolithic sequence to ca. 11,000–7000 cal BP (Görsdorf and Gramsch Reference Görsdorf and Gramsch2004)Footnote 1 .
Holocene Environment
During its prehistoric occupation, Friesack 4 was located in a highly productive wetland, providing easy access to terrestrial and aquatic resources for the Mesolithic hunter-gatherer-fisher populations. Initially, the location provided good access to open water, but by the latest Mesolithic phase (Boreal), overgrowing processes partly enclosed the island on the north and east (Kloss Reference Kloss1987a: 125–129; Groß Reference Groß2017: 68–73).
Vegetation development followed the general trend in northeastern Germany: light pine-birch forest in the Preboreal (ca. 11,600–10,640 cal BP) (Litt et al. Reference Litt, Brauer, Goslar, Merkt, Bałaga, Müller, Ralska-Jasiewiczowa, Stebich and Negendank2001), followed by denser vegetation during the Boreal (ca. 10,640–9200 cal BP) (Litt et al. Reference Litt, Brauer, Goslar, Merkt, Bałaga, Müller, Ralska-Jasiewiczowa, Stebich and Negendank2001), with a constant presence of hazel. Marsh and aquatic plant pollen was most abundant during the Boreal, indicating extensive overgrowing (Jahns et al. Reference Jahns, Gramsch and Kloss2016, 30). In the Atlantic chronozone (ca. 9200–5600 cal BP) (Dörfler et al. Reference Dörfler, Feeser, van den Bogaard, Dreibrodt, Erlenkeuser, Kleinmann, Merkt and Wiethold2012), mixed-oak forests formed in dry areas and alder forest in moist surroundings (Kloss Reference Kloss1987a: 123–126; Reference Kloss1987b: 112–114; Jahns et al. Reference Jahns, Gramsch and Kloss2016: 27–31). The groundwater level reached its prehistoric minimum in the Late Boreal (Gramsch Reference Gramsch2002: 191; Jahns et al. Reference Jahns, Gramsch and Kloss2016: 28) but rose at the end of the Atlantic, so that formerly dry spots were covered with alder. The groundwater level fell again in the Early Sub-Boreal (ca. 5600–2600 cal BP) (Gramsch Reference Gramsch2002; Jahns et al. Reference Jahns, Gramsch and Kloss2016: 28).
Subsistence Economy
Over 9200 mammal remains from Complexes I–V have been identified (Benecke Reference Benecke2016; Schmölcke Reference Schmölcke2016); large game, particularly cervids, dominate in each phase (Figure 2). Numerically, roe deer (Capreolus capreolus) predominates, but red deer (Cervus elaphus) was more important in terms of meat weight. Fur-bearing animals, particularly beaver (Castor fiber), occur frequently in all phases. Domestic cattle (Bos taurus), undifferentiated pig (Sus sp.), sheep (Ovis aries), and goat (Capra hircus) occur regularly in Complex V, but together account for only 10% (number of identified specimens, hereafter NISP) of mammal remains in this phase. Metrical and morphological analyses of juvenile animal bones permitted their attribution to narrow age classes, which reflect the seasonality of hunting and thus the seasonality of site occupation. During the Early Mesolithic Friesack 4 was probably a summer camp; in winter, people probably lived in forest hunting camps on neighboring moraines (Benecke Reference Benecke2016; Schmölcke Reference Schmölcke2016).

Figure 2 Mammal assemblage composition, Complexes I–V, with NISP at the right of each bar (Benecke Reference Benecke2016; Schmölcke Reference Schmölcke2016).
Fish remains from Complexes I–IV were analyzed recently; 7520 specimens were examined, of which 1733 could be identified to family or species (Robson Reference Robson2016). Eight species from six families, all freshwater taxa, were recorded. The fish were probably caught during late spring and early summer, using an array of methods (see Robson Reference Robson2016). Northern pike (Esox lucius) predominated in Complexes I to III, but Wels catfish (Siluris glanis) became more important in Complex IV (Figure 3).

Figure 3 Fish species composition, Complexes I–IV, with NISP at the right of each bar (Robson Reference Robson2016).
As no caches of plants or human coprolites were found, evidence of plant food consumption at Friesack 4 is limited, but charred hazelnuts (Corylus avellana), berries (e.g. Fragaria sp.; Rubus sp.), water-lily seeds (Nymphaea sp.) and chenopods (Chenopodioideae) demonstrate the potential use of these plants. Bulrush (Typhaceae) rhizomes were probably also used. Water chestnut (Trapa natans) was only identified once, and may not have been important (Wolters Reference Wolters2016). The contribution of plant resources to overall human diets in each occupation phase is unknown.
Human Remains
Twenty human remains (15 bones and 5 isolated teeth) were recovered during Gramsch’s excavations. These remains represent secondary deposition, as there was no evidence of grave cuts or articulation. Four perforated teeth were interpreted as ornaments, including an unerupted permanent molar, perforated post mortem (Ullrich and Gramsch Reference Ullrich and Gramsch2015). To date, only one human bone from Friesack, a skull fragment found during dredging in 1980, has been subjected to both radiocarbon (14C) dating and carbon (δ13C) and nitrogen (δ15N) stable isotope analyses (Terberger et al. Reference Terberger, Gramsch and Heinemeier2012).
Research Questions
Mesolithic human remains, particularly from stratified contexts, are extremely rare in northern central Europe; the largest published corpus of dietary stable isotope data is from disarticulated bones dredged from former land surfaces on the floor of the North Sea (van der Plicht et al. Reference van der Plicht, Amkreutz, Niekus, Peeters and Smit2016). The remains from Friesack, with a detailed archaeological and palaeoenvironmental context, are therefore especially valuable to understanding Early-Middle Holocene subsistence strategies. We have now sampled all human bones from Friesack 4 in order to: (i) reconstruct individual diets, (ii) estimate each individual’s calendar date, (iii) relate human bone dates to the site chronology, and (iv) observe dietary patterns from the Early Mesolithic to the Early Neolithic. To make our reconstruction more accurate, we have also sampled 182 archaeozoological remains to provide a local isotopic baseline for freshwater and terrestrial foods, and six modern fish to estimate the potential magnitude of dietary 14C reservoir effects.
MATERIALS AND METHODS
Sampling
All 15 bones identified as human (Ullrich and Gramsch Reference Ullrich and Gramsch2015) were sampled for 14C, δ13C and δ15N analyses (Table 1)Footnote 2 . Within a larger study of Early Holocene mammal ecology in the North European plain (Hegge in prep.), 153 adult elk (Alces alces), red deer, roe deer and wild boar (Sus scrofa) bones from Complexes I–V at Friesack 4 and the nearby Mesolithic site Friesack 27a were sampled for δ13C and δ15N (Table 2). In addition, δ13C and δ15N analyses of 29 mammal and fish bones from Friesack 4 (4 aurochs, 5 European beaver, 8 northern pike, 4 European perch (Perca fluviatilis), 4 Wels catfish and 4 European eel [Anguilla anguilla]) were undertaken for the present study (Table 2).
Table 1 Analytical results from human remains. The following human remains were sampled but could not be dated due to a lack of collagen: 1977:7/M05 IV Z F4S 4 (adult thoracic vertebra), 1977:7/M07 IV A A5 4 (diaphysis fragment, male right ulna), 1977:7/M09 IV A A6 4 (fibula(?) fragment), 1977:7/M14 IV Z E4S 31c (adult cranium, 26 small fragments) (all failed in Kiel), and 1977:7/M04 III A+C (mandible, >40 yr) (failed at Aarhus).

§ Incorporating dietary reservoir-effect corrections (see text and Appendix S2), 95% probability, rounded outwards to 50 years.
* First published by Terberger et al. (Reference Terberger, Gramsch and Heinemeier2012).
# First published by Ullrich and Gramsch (Reference Ullrich and Gramsch2015).
Table 2 Summary statistics for the sampled faunal remains (mean±standard deviation).

In April 2017, six fish were caught in two small lakes on the River Rhin, ca. 40 km upstream (Ruppiner See) and ca. 20 km downstream (Guelper See) of Friesack (Figure 1). Northern pike, a piscivorous fish, and common bream (Abramis brama), which consumes low-trophic level aquatic species, were taken at both sites (two of each at Ruppiner See). Their sizes suggest that all six fish were several years old (Table 3).
Table 3 AMS measurements of freeze-dried flesh from fresh fish.

§ Conventional 14C age (sensu Stuiver and Polach Reference Stuiver and Polach1977), t=-8033×ln (F14C/1.000). The denominator 1.000 in the age calculation corresponds to the reference atmospheric 14C activity in AD 1950.
† Apparent 14C age=–8033×ln (F14C /1.015), i.e. calculation based on atmospheric 14C activity of 1.015±0.005 F14C, the average of May–August 2014–2016 atmospheric data (Northern Hemisphere mid-latitude peak growing season) (Hammer and Levin Reference Hammer and Levin2017).
AMS Dating and EA-IRMS Analysis of Human Bones
Four human bones were dated at the AMS 14C Dating Centre, Aarhus University, Denmark. Collagen extraction followed a modified Longin protocol (Olsen et al. Reference Olsen, Heinemeier, Lübke, Lüth and Terberger2010). An aliquot of collagen was sealed for AMS dating by a collaborating laboratory (Zoppi et al. Reference Zoppi, Crye, Song and Arjomand2007), and a second aliquot was analyzed by elemental analysis isotope ratio mass spectrometry (EA-IRMS) at Aarhus, by combustion in a EuroVector elemental analyzer coupled to an IsoPrime stable isotope ratio mass spectrometer. All δ13C and δ15N results are reported per mil (‰) relative to the international standards for δ13C (V-PDB) and δ15N (AIR).
At the Leibniz-Laboratory, Christian-Albrechts-University, Kiel, Germany, ca. 1 g of raw flesh from each fish was freeze-dried; ca. 6 mg of dry flesh was then combusted, graphitized and dated (Nadeau et al. Reference Nadeau, Grootes, Schleicher, Hasselberg, Rieck and Bitterling1998). Bone collagen was extracted following a modified Longin protocol (Grootes et al. Reference Grootes, Nadeau and Rieck2004), and filtered through a 0.45 μm silver filter before freeze-drying. An aliquot of collagen was sealed for AMS dating in Kiel, and a second aliquot was sent to the School of Life Sciences, University of Bradford, United Kingdom, for duplicate EA-IRMS measurements, using a Thermo Flash 1112 elemental analyzer and a Thermo Delta plus XL mass spectrometer. Laboratory and international standards were analyzed simultaneously. Typical uncertainties of±0.2‰ are quoted for both δ13C and δ15N.
Faunal Stable Isotope Analysis
Collagen from the elk, red deer, roe deer, and wild boar samples was extracted at the Leibniz-Laboratory (following Grootes et al. Reference Grootes, Nadeau and Rieck2004, but using a 5-μm cellulose nitrate filter), and analyzed by EA-IRMS at the Museum für Naturkunde, Berlin, Germany, using a Thermo Flash 1112 elemental analyzer coupled to a Thermo/Finnigan MAT V isotope ratio mass spectrometer; δ13C and δ15N measurements of peptone standards have standard deviations <0.15‰.
Collagen from the aurochs, beaver, and fish samples was extracted and analyzed at the BioArCh research center, University of York, United Kingdom. Bone was coarsely ground and demineralized in 0.6 or 0.1 M HCl in a cold room (4°C), rinsed with distilled H2O and gelatinized (pH3 [0.001M] HCl, 80°C, 48 hr). The supernatants were subsequently ultrafiltered (30kDa, Amicon® Ultra-4 Centrifugal Filter Units, Millipore, Billerica, MA, USA), frozen and lyophilized. Collagen was analyzed in duplicate by EA-IRMS using a Sercon GSL analyzer coupled to a Sercon 20-22 mass spectrometer. Accuracy was <0.3‰, based on results of standards (IAEA-600, IAEA-N2).
Statistical Modeling
Individual diets were estimated using FRUITS (Fernandes et al. Reference Fernandes, Millard, Brabec, Nadeau and Grootes2014), assuming three food groups (terrestrial animals and plants, freshwater fish; Appendix S1). The same parameter values were applied to each individual, as there is no evidence that baseline isotope values changed over the period of interest. FRUITS output includes probability distributions for each food group’s contribution to collagen δ13C and δ15N. The fish contribution to δ13C (and thus also to 14C) permits dietary reservoir-effect correction, if the fish freshwater reservoir effect (FRE) is known, using the Mix_Curves command in OxCal (Bronk Ramsey Reference Bronk Ramsey2001; Meadows et al. Reference Meadows, Bērziņš, Brinker, Lübke, Schmölcke, Staude, Zagorska and Zariņa2016). Appendix S2 includes the OxCal CQL (chronological query language) code used to calibrate human bone 14C ages.
A Bayesian chronological model for Mesolithic occupation at Friesack 4 was created using the 87 published bulk charcoal dates (Görsdorf and Gramsch Reference Görsdorf and Gramsch2004). To reduce the influence of large wood-age offsets, OxCal’s Charcoal Outlier_Model function (Dee and Bronk Ramsey Reference Dee and Bronk Ramsey2014) was applied to all these dates. Instead of using the highly constrained prior information applied by Görsdorf and Gramsch (Reference Görsdorf and Gramsch2004), our model (Appendix S2) simply consists of four sequential bounded phases, Complexes I–IV. A more detailed chronological model is unnecessary here, as faunal data are aggregated by sedimentary complex, and human remains were all redeposited.
RESULTS
Archaeological Samples
Of 197 samples extracted, 137 (69.5%) yielded enough collagen for reliable measurement and had atomic C:N values in the accepted range, 2.9–3.6 (DeNiro Reference DeNiro1985; Szpak Reference Szpak2011).
Nine of the 15 human bones yielded enough collagen, giving a wide range of 14C ages, δ13C and δ15N values (Table 1; Figure 4). The four bones dated in Aarhus were poorly preserved, yielding 0.8–3.0% collagen by weight, while one sample failed. Yields of the five samples dated in Kiel were better (3.1–10.0%), but four more samples failed, with yields ≪1%. The lower yields for samples dated at Aarhus thus reflect variable collagen preservation, not the minor differences in extraction protocols. The lower-yielding bones also gave lower δ13C and higher δ15N values, but this is probably coincidental, as yields and isotopic data are not correlated with C:N values. There is, moreover, a clear correlation between δ13C and δ15N (Pearson’s r=–0.833, p=0.005), which cannot easily be explained by diagenesis, as the observed pattern would require a hypothetical contaminant with either much higher δ15N and much lower δ13C than human bone collagen, or much lower δ15N and much higher δ13C; these isotopic signatures should not occur in early Holocene Northern Europe. The pattern must instead reflect dietary differences, with lower δ13C and higher δ15N reflecting greater intake of δ13C-depleted and δ15N-enriched freshwater fishFootnote 3 .

Figure 4 δ13C and δ15N on fish, mammal, and human bone collagen from Friesack. Point and whisker symbols represent the mean and standard deviations from multiple samples of the same species.
Of 162 mammal bones sampled, 124 yielded enough well-preserved collagen for reliable measurement (Table 2). The herbivores (elk, red deer, roe deer, aurochs, and beaver) gave almost identical average isotope values, while the omnivorous wild boar had slightly elevated δ15N values (Figure 4). As the mammal assemblage is dominated by cervids in every phase (Figure 2), and as elk, red deer, roe deer, and wild boar δ13C and δ15N values did not change over time (Hegge in prep.), the averaged mammal δ13C and δ15N values (–21.4‰, 5.0‰) should apply throughout the prehistoric occupation.
Only four of the 20 fish bones, from two species (eel and perch), produced enough well-preserved collagen for analysis (Table 2). Their isotopic values are clustered, and were on average δ13C 4.1‰ lower than the mammals, and δ15N 4.4‰ higher (Figure 4).Footnote 4
Freshwater Reservoir Effects
Our modern fish results did not show any 14C age difference between species or sampling locations (Table 3). As the atmospheric 14C level is higher than assumed in conventional 14C age calculation, we used the average 14C level during the previous three growing seasons (Hammer and Levin Reference Hammer and Levin2017) to calculate apparent 14C ages, which correspond to FREs in archaeological samples. If these values were applicable in the past, the average FRE in fish available at Friesack was ca. 1210 14C years. We cannot test whether the FRE was higher or lower prehistorically, as there are no identifiable pairs of exactly contemporaneous aquatic and terrestrial organisms that might be dated, but the FRE in modern fish is large enough to imply that human bone dates will be subject to significant dietary reservoir effects if fish was consumed regularly.
Diet Reconstruction
Figure 5 shows part of the FRUITS output. Plants are estimated to provide most of the energy in all diets, while animal foods provide much of the protein consumed by individuals with low-fish diets, and are only minor protein sources in high-fish consumers. Detailed results are given in Appendix S1.
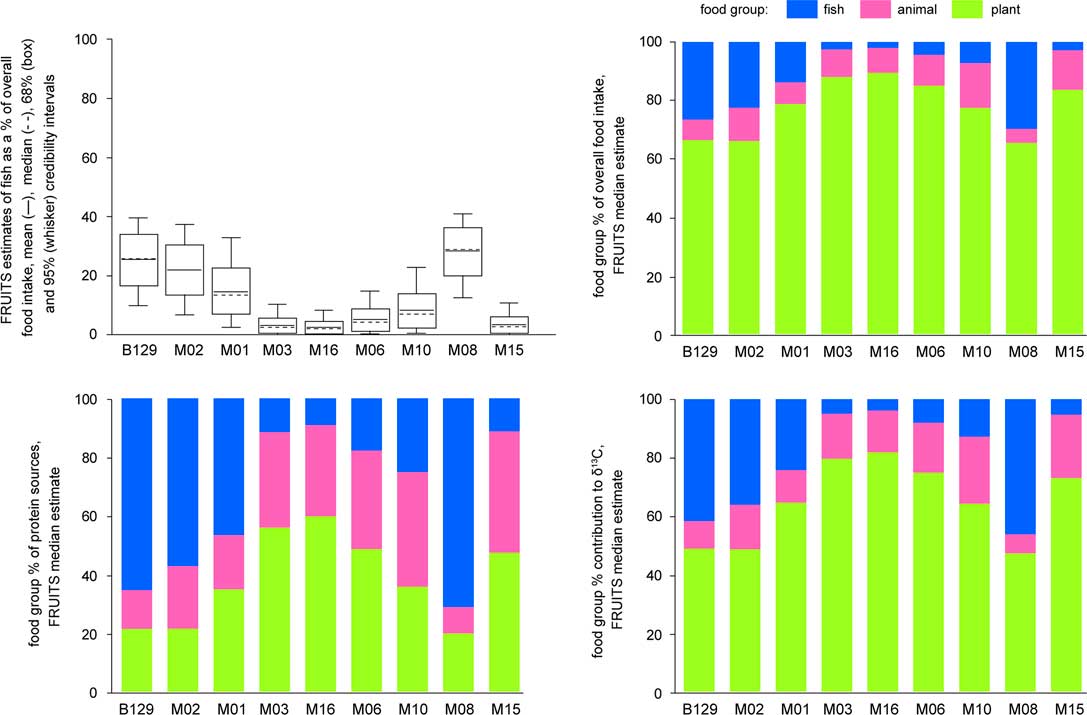
Figure 5 FRUITS individual diet reconstructions (for detail see Appendix S1). Individual sample codes correspond to those used in Table 1 and the main text.
Dietary Reservoir Effects
As fish is assumed to be rich in protein, its estimated share of protein intake is much higher than its share of overall diet. This means that a disproportionate share of carbon in collagen is fish-derived, as collagen is built mainly from dietary protein, although fats and carbohydrates also contribute carbon to collagen, particularly in low-protein diets (Fernandes et al. Reference Fernandes, Nadeau and Grootes2012). Combining median estimates of fish contribution to δ13C (Figure 5) with the modern fish FRE (1210 14C years), human 14C ages may embody dietary reservoir effects of between 60 (M16) and 600 years (M08). The accuracy of these estimates cannot be tested, as the human bones were redeposited, but there is no reason to think that the Early Holocene FRE at Friesack was lower than it is today. For calibration (Table 1; Appendix S2), we include uncertainty by using the mean±standard deviation of the fish contribution to δ13C (Appendix S1), and the standard error on the mean FRE of modern fish (±30).
Site Chronology
Figure 6 shows part of the OxCal model output, including the date ranges of charcoals from Complexes I–IV and diet-corrected dates of human bones. Overall, and within each excavation area (X, B, A & C, D, Z), the bulk charcoal dates fit the basic stratigraphic sequence. Complex I (early 11th millennium cal BP) and II (mid-11th millennium) were each relatively short phases, separated by a brief hiatus; Complex III lasted from the late 11th until the mid-10th millennium. After a hiatus of 200–400 years, Complex IV lasted from the late 10th to the mid-8th millennium cal BP.

Figure 6 Aspects of the OxCal model output (for detail see Appendix S2). Blue: average dates of bulk charcoal samples from Mesolithic sedimentary Complexes I–IV. Black: simple calibration of human bone 14C ages, with no correction for dietary reservoir effects. Red: estimated dates of these individuals, based on the diet reconstructions (Appendix S1) and modern fish 14C ages (Table 3).
DISCUSSION AND CONCLUSION
Individual Results
Figure 5 summarizes individual diet reconstructions. The stacked bar charts may be misleading, as they show median estimates without uncertainties, but the box-and-whisker plot indicates wide variations in fish consumption. In high-fish diets (e.g. B129), the high protein content of fish restricts the possible animal food intake, due to physiological limits to how much protein humans can metabolize (e.g. Speth and Spielmann Reference Speth and Spielmann1983); all other estimates are therefore more precise. Lower fish intake (e.g. M03) is associated with larger uncertainties in the importance of plant and animal foods, whose parameter values (isotopes and protein contents) are relatively similar; thus plant-food intake may actually have been similar (e.g. 60–70% of calories) for all individuals. In 14C terms, whether low-fish diets were dominated by plant or animal foods is irrelevant, but the local FRE is so high that even moderate uncertainties in the fish contribution to δ13C (e.g.±10%) make the diet-corrected dates far less precise than uncorrected calibrated dates (Figure 6).
Human Remains and Stratigraphic Complexes
All the human bones were disarticulated, and their association with any of the occupation complexes at Friesack is therefore debatable. Correction for dietary reservoir effects (implicitly assuming that these individuals were local residents) excludes some potential associations. The stray find, B129, is clearly Preboreal (after 11,700 cal BP; uncorrected, it dates to the Younger Dryas), but still appears to pre-date Mesolithic occupation at Friesack 4. M02 (from Complex II), which had appeared to pre-date Complex I, could be associated with Complex I or II, or the hiatus between them. M01 and M03 both appear to date to the start of Complex III, despite having been found in Complex II, and having had different diets. M16 dates either to the start of Complex IV, where it was found, or to the hiatus between Complex III and IV.
The other Complex IV bones are much more recent, however, dating to the end of the Early Neolithic (late 4th millennium cal BC). These samples (M06, M10, M08) could be exactly contemporaneous, despite their dietary differencesFootnote 5 . Finally, M15 dates to the earlier Bronze Age, not the expected Early Neolithic period. Indeed, even without correction for dietary reservoir effects, many human bones appear to be intrusions into older sediment layers.
Dietary Patterns at Friesack
Despite limited direct evidence of plant consumption, our reconstructions (Figure 5) show that overall diets were probably always plant-based, but that the main protein source alternated, between terrestrial mammals and freshwater fish. It is difficult to discern paleodiet patterns using faunal NISP values, due to differences in taphonomic filters and potential changes in seasonality of occupation, but in any case faunal data (particularly from Complexes I–III) do not show strong trends (Figures 2 and 3). Although fish remains were not as abundant at Friesack 4 as at other Mesolithic sites (e.g. Boethius Reference Boethius2017; Boethius et al. Reference Boethius, Storå, Hongslo Vala and Apel2017) and may even represent a taphocoenosis, our results show that some Early Mesolithic humans (B129, M02, M01) relied mainly on fish for their protein intake, while others (M03, M16) had almost fully terrestrial diets. We can tentatively associate a shift to more terrestrial diets with the Preboreal-Boreal transition, which coincided with reduced access to open water at Friesack 4 (above). The Early Neolithic individuals had contrasting diets: M06 and M10 were close to the terrestrial end of the spectrum, but M08 had the most aquatic diet of the Friesack humans. The Bronze Age individual (M15) had a mainly terrestrial diet, based on the C3 photosynthetic pathwayFootnote 6 .
Friesack in Context
Considering their date range, the number of individuals analyzed is too small to reveal robust paleodiet patterns at Friesack 4. Even assuming that these individuals were all local residents (which is difficult to demonstrate, given the scattered and fragmentary nature of these finds), differences between their diets may be interpreted as evidence either of temporal shifts in subsistence strategies, or of dietary diversity within a single economy (perhaps due to differences in seasonal mobility, or to social factors governing dietary preferences).
To better understand the situation at Friesack, we may compare our results to those of other paleodiet studies in this region (Figure 7), at the coarse temporal resolution permitted by our diet-corrected dates, and by the lack of reservoir-effect correction (or of 14C ages) for other assemblages.

Figure 7 δ13C and δ15N isotope data obtained on human bone collagen from Friesack (circles) compared with data from contemporaneous human remains (squares) throughout Northern Europe (after Grünberg Reference Grünberg2000; Fischer et al. Reference Fischer, Olsen, Richards, Heinemeier, Sveinbjörnsdóttir and Bennike2007a, Reference Fischer, Richards, Olsen, Robinson, Bennike, Kubiak‐Martens and Heinemeier2007b; Olsen et al. Reference Olsen, Heinemeier, Lübke, Lüth and Terberger2010; Terberger et al. Reference Terberger, Gramsch and Heinemeier2012; Bollongino et al. Reference Bollongino, Nehlich, Richards, Orschiedt, Thomas, Sell, Fajkošová, Powell and Burger2013; Drucker et al. Reference Drucker, Rosendahl, Van Neer, Weber, Görner and Bocherens2016; van der Plicht et al. Reference van der Plicht, Amkreutz, Niekus, Peeters and Smit2016).
The protein intake of the Rhünda human, a stray find from central Germany, was dominated by freshwater fish (Drucker et al. Reference Drucker, Rosendahl, Van Neer, Weber, Görner and Bocherens2016); with even a minor reservoir-effect correction, this individual (GrA-15947, 10,200±60 BP) would date to the early Preboreal, like B129 (AAR-15034, 10,277±28 BP) from Friesack. A Preboreal cranium from the North Sea (GrA-42700, 10,070±50 BP; van der Plicht et al. Reference van der Plicht, Amkreutz, Niekus, Peeters and Smit2016) also has isotopic values indicating a freshwater-fish-based diet.
About a dozen more North Sea finds dated to the later Preboreal or early Boreal have isotopic values consistent with heavy reliance on freshwater fish (high δ15N and/or low δ13C); none indicates a mainly terrestrial diet (van der Plicht et al. Reference van der Plicht, Amkreutz, Niekus, Peeters and Smit2016), although two individuals of similar date from Unseburg, ca. 100 km southwest of Friesack, appear to have had more mixed diets (Grünberg Reference Grünberg2000), as does an individual from Koelbjerg, in Denmark (Fischer et al. Reference Fischer, Olsen, Richards, Heinemeier, Sveinbjörnsdóttir and Bennike2007a; Figure 7). Thus the Friesack individuals dated to this period (M02 and M01) are again typical, in relying more on freshwater fish than on terrestrial protein sources.
The mainly terrestrial diets of M03 and M16 at Friesack are consistent with the low δ15N and moderate δ13C values in the three later Boreal individuals at Blätterhöhle, in western Germany (δ15N 7.6–8.2‰; Bollongino et al. Reference Bollongino, Nehlich, Richards, Orschiedt, Thomas, Sell, Fajkošová, Powell and Burger2013), and in one from Tømmerupgård in Denmark (δ15N 8.3‰; Fischer et al. Reference Fischer, Olsen, Richards, Heinemeier, Sveinbjörnsdóttir and Bennike2007a). However, most North Sea and Danish individuals of the same period had much more aquatic diets (freshwater [high δ15N and/or low δ13C] or marine [high δ15N and high δ13C]; van der Plicht et al. Reference van der Plicht, Amkreutz, Niekus, Peeters and Smit2016; Fischer et al. Reference Fischer, Olsen, Richards, Heinemeier, Sveinbjörnsdóttir and Bennike2007a; Figure 7).
There are practically no dietary stable isotope studies of Late Mesolithic human remains in northern central Europe (corresponding to Complex IV at Friesack 4), other than the Argus Bank remains (Fischer et al. Reference Fischer, Richards, Olsen, Robinson, Bennike, Kubiak‐Martens and Heinemeier2007b), and the Groß Fredenwalde inhumations (Terberger et al. Reference Terberger, Kotula, Lorenz, Schult, Burger and Jungklaus2015). At Argus Bank, a coastal site in southeastern Denmark, all 4 individuals clearly depended mainly on marine sources of protein. The Groß Fredenwalde situation is more complex, as stable isotope values are quite tightly clustered, but the 14C ages of 7 or 8 apparently contemporaneous individuals imply a wide range of dietary FRE, from negligible to ~500 14C years. Without reference data for potential FRE in local fish, it is unclear whether the variation in human 14C ages is due more to differences in diet or residence.
For the later Early Neolithic, there are many more comparanda from stratified sites, such as the burial ground at Ostorf-Tannenwerder, ca. 140 km northwest of Friesack. Long after the start of farming in the region, this community relied mainly on freshwater fish protein (Olsen et al. Reference Olsen, Heinemeier, Lübke, Lüth and Terberger2010; Fernandes et al. Reference Fernandes, Grootes, Nadeau and Nehlich2015). At Blätterhöhle, it is argued that in the later 4th millennium cal BC there were two subsistence strategies, linked to population ancestries, one based on freshwater fish and one on terrestrial (domestic) resources (Bollongino et al. Reference Bollongino, Nehlich, Richards, Orschiedt, Thomas, Sell, Fajkošová, Powell and Burger2013). Thus the fish-based diet of M08 and the terrestrial diets of the contemporaneous M06 and M10 hint at the coexistence of different groups at Friesack, some relying on fish and other wild resources, and others presumably farmers or pastoralists.
Overall, then, the small human bone assemblage from Friesack 4 reinforces our impression that inland Early Mesolithic individuals, particularly in the Preboreal, depended heavily on fishing, although by the late Boreal there were also some who relied almost entirely on wild terrestrial foods. The coexistence of Neolithic farmers and hunter-gatherer-fishers, already known from Blätterhöhle and coastal Scandinavia (Lidén et al. Reference Lidén, Eriksson, Nordqvist, Götherström and Bendixen2003), may be a more widespread phenomenon in inland Northern Europe than hitherto recognized (see also Terberger et al. submitted). Our results strengthen the case for 14C and stable isotope analyses of all human remains to complement other specialist studies, and illustrate the need to quantify dietary reservoir effects for each individual analyzed.
ACKNOWLEDGMENTS
Analyses at Aarhus of four human bones were funded by the German Research Foundation (DFG) grant TE 259/3-2, to TT. 153 faunal remains were analyzed within CH’s PhD research at Christian-Albrechts-University, Kiel, funded by DFG grant SO 861/2-1, to Dr Robert Sommer. Other analyses were funded through the Centre for Baltic and Scandinavian Archaeology (ZBSA)’s Man and Environment research theme, including subproject B2: Transitions of specialized foragers of the Collaborative Research Centre Scales of Transformation: Human-environmental Interaction in Prehistoric and Archaic Societies (DFG-SFB 1266). We thank all the laboratory staff responsible for these measurements.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/RDC.2018.69












