


Introduction
Every pest management tactic potentially impacts the overall biological community and produces unintended or nontarget effects. As highlighted by recent scrutiny of Bt-insecticidal crops and conventional insecticide applications (Dively, Reference Dively2005; Marvier et al., Reference Marvier, McCreedy, Regetz and Kareiva2007; Wolfenbarger et al., Reference Wolfenbarger, Naranjo, Lundgren, Bitzer and Watrud2008), the salient question for guiding agricultural decision-making is how the unintended effects of a technology compare to those of the practices it can supplant. Another question is how relevant those nontarget effects actually are to agroecosystem health and sustainability. Minor reductions in the abundance of predators observed in Bt-cotton fields, for instance, were determined to have “little ecological meaning” for the natural regulation of pest populations (Naranjo, Reference Naranjo2005a, Reference Naranjob). These dimensions of nontarget assessment are probably relevant well beyond such high profile scenarios as genetically-modified organisms and broad spectrum insecticides.
In managed turfgrass systems, carbamate and organophosphate insecticides have been shown, through diverse studies, to negatively impact the abundance of natural enemies (Cockfield & Potter, Reference Cockfield and Potter1983; Terry & Potter, Reference Terry and Potter1987; Vavrek & Niemczyk, Reference Vavrek and Niemczyk1990), as well as their function in the regulation of pest populations (Cockfield & Potter, Reference Cockfield and Potter1983; Terry et al., Reference Terry, Potter and Spicer1993; Zenger & Gibb, Reference Zenger and Gibb2001; Rogers & Potter, Reference Rogers and Potter2003). In the last decade, there has been a shift toward reliance on newer chemistries that feature lower vertebrate toxicity, reduced water solubility and more selective modes of action (Potter, Reference Potter1998). Foremost among these are the neonicotinoids, including acetamiprid, clothianidin, dinotefuran, imidacloprid and thiamethoxam (Maienfisch et al., Reference Maienfisch, Angst, Brandl, Fischer, Hofer, Kayser, Kobel, Rindlisbacher, Senn, Steinemann and Widmer2001; Tomizawa & Casida, Reference Tomizawa and Casida2003; Wakita et al., Reference Wakita, Kinoshita, Yamada, Yasui, Kawahara, Naoi, Nakaya, Ebihara, Matsuno and Kodaka2003; Jeschke & Nauen, Reference Jeschke and Nauen2008). While these still retain a relatively broad spectrum of insecticidal activity (Elbert et al., Reference Elbert, Nauen, Leicht, Ishaaya and Degheele1998), their systemic nature, like that of plant-induced toxins (e.g. Bt-transgenic plants), is considered to have fewer nontarget effects due to reduced ecological exposure (Smith & Krischik, Reference Smith and Krischik1999). It is, therefore, reasonable to expect that the magnitude of adverse effects on beneficial fauna like the natural enemies of pests might be diminished under control programs that favor these compounds. On the other hand, the relatively long-residual of neonicotinoids might result in increased environmental exposure to nontargets because of their persistence in the field.
Imidacloprid is probably the most widely used intervention product for turf-infesting white grubs (Coleoptera: Scarabaeidae) in the United States. Given the ideal application window at the time of oviposition, imidacloprid is applied preventively (Potter, Reference Potter1998), meaning that application decisions are made before it is feasible to scout pest populations and to assess thresholds. Turf managers are, therefore, more likely to make preventive applications because there is no method to gauge infestation levels in advance. One ramification is that applications may be made against non-damaging populations; another is that repeated yearly applications might be promoted in risk-adverse settings, such as on golf course playing surfaces.
Studies, therefore, are needed to address whether there are discernible nontarget effects on beneficial arthropods from neonicotinoid insecticides, whether those impacts are relevant to turfgrass health, and, if so, how they might be mitigated. This approach is further justified by evidence that links imidacloprid to pollinator declines (Chauzat et al., Reference Chauzat, Faucon, Martel, Lachaize, Cougoule and Aubert2006; Halm et al., Reference Halm, Rortais, Arnold, Tasei and Rault2006; Abbott et al., Reference Abbott, Nadeau, Higo and Winston2008) and by the extensive areas managed for production and amenity turf. There are an estimated 200,000 km2 of cultivated turfgrass in the US, providing a broad range of aesthetic, economic, recreational and environmental services (Beard & Green, Reference Beard and Green1994; NTF, 2003). In addition to production grass seed and sod farms, turf systems include residential home lawns, golf courses, athletic fields, parks and other public areas. In New York State, for instance, turfgrass covers 13,800 km2 and contributes $5.1 billion annually to the state's economy (NASS, 2004). Across these systems, white grubs are the most widespread and damaging insect pest complex, and their management would benefit from more informed decision-making as to the selection and application of control interventions.
In previous studies with white grub control options like trichlorfon, halofenozide and entomopathogenic nematodes, only imidacloprid harbored discernible nontarget impacts on the abundance of soil-active invertebrates in turfgrass (Peck, Reference Peck2009a). Repeated preventive applications reduced populations of certain nontarget taxa, including Coleoptera, Collembola, Hemiptera and Thysanoptera, the effects of which persisted over all six years of applications (Peck, Reference Peck2009b). Population reductions were expressed in the soil-active fauna recovered through Tullgren funnel extractions of soil cores, but not in the surface-active fauna captured from pitfall traps. Among the affected taxa are beneficials that probably play a role in stabilizing the turfgrass ecosystem through nutrient cycling and the natural regulation of pest populations. In turfgrass, the activities of decomposers such as earthworms and springtails contribute to thatch decomposition, while the activities of generalist natural enemies, such as ants, ground beetles and spiders, contribute to suppression of pest outbreaks (Potter, Reference Potter, Racke and Leslie1993, Reference Potter and Leslie1994). The extent to which selective pest management tactics can protect the services of these groups is key to maintaining healthy turf. Although imidacloprid is implicated in suppressing the populations of these potential beneficial soil-dwelling organisms, it is unknown if the magnitude of that effect has any relevance for their function in ecological processes, i.e. whether the amount of beneficial population reduction induced by imidacloprid will significantly impact pest management. Balanced with the positive attributes of a preventive imidacloprid regime in limiting the impact of white grub outbreaks, do we need to care that beetles and other groups suffer collateral damage?
This study examines the impact of single applications of neonicotinoid insecticides on the regulation of pest populations by predators. The objective was to detect and measure the magnitude and persistence of neonicotinoid impacts on the removal rates of Japanese beetle (Popillia japonica Newman) eggs. The egg was chosen because it is a known prey item for generalist natural enemies, it has been previously used to gauge the impacts of other insecticides, and it is present when preventive applications are made. The study was conducted as a series of three experiments. In a first experiment, data were obtained that confirmed the potential of a single application of imidacloprid to suppress the abundance of nontarget fauna and the rate of egg removal. A second experiment was conducted to compare the impact of four neonicotinoid insecticides on egg removal. Finally, a third experiment was conducted to measure changes in egg removal when imidacloprid was applied to target white grubs at three different times over the season. The working hypothesis was that a single application of neonicotinoid insecticide can lead to lower rates of egg predation and that the effect is temporary and does not vary with respect to active ingredient or month of application.
Methods
Study site
The experiments were conducted at the Crittenden North research farm at Cornell University's New York State Agricultural Experiment Station, Geneva, NY. Study plots were situated on a 1.3-ha site managed to simulate a low maintenance home lawn. The experimental area received no irrigation, fertilization, pesticides or overseeding since its conversion in 1997 from an orchard fallow. Maintenance involved weekly mowing (cutting height 10.4 cm) with clippings left on the surface. Starting in 2006, there was an autumn aeration using a tractor-pulled core aerator. A single broadcast application of urea fertilizer (0.48 kg N 100 m−2) was made in June 2007. Turf composition was fine fescue (38.0%), perennial ryegrass (22.7%), weedy grass species (0.8%) and broadleaf weeds (37.9%). Soil was silty clay loam (12.5% sand, 55.0% silt, 32.5% clay) with 5.7% organic matter and pH 6.5. All experimental plots were 10×10 m, separated from one another by a 10-m untreated buffer.
Treatment applications
Most insecticide treatments were applied in liquid formulation using a pressurized CO2 sprayer system (R and D Sprayers, Opelousas, LA). Applications were made at 30 psi, using a 1.2-m boom fitted with four flat fan nozzles (Teejet® 8002VS, TeeJet Technologies, Wheaton, IL) for a final coverage width of 1.5 m. Application volume was standardized to 1.2 liters 100 m−2. Rate of application matched the standard labeled rates for turfgrass application, or, in the case of dinotefuran (not labeled for use on turf), the maximum labeled rate for ornamental plants. Four neonicotinoid insecticides were included in these studies based on the following products and rates: clothianidin (Arena 50 WDG, 0.21 kg AI ha−1, Valent, Richardson, TX), dinotefuran (Safari 20 SG, 0.59 kg AI ha−1, Valent), imidacloprid (Merit 2F, 0.44 kg AI ha−1, Bayer Environmental Science, Montvale, NJ) and thiamethoxam (Meridian 25 WG, 0.31 kg AI ha−1, Syngenta, Wilmington, DE). Nothing was applied to the untreated check plots. Experiment 1 (see below) was an exception because imidacloprid was applied in a granular formulation (Merit 0.2G, 0.44 kg AI ha−1, Bayer CropScience, Research Triangle Park, NC) with a drop spreader (Model #D, Gandy, Owatanna, MN).
Post-application irrigation is part of the strategy to improve how insecticides target white grubs that reside at the soil/thatch interface. This practice may not be very common, however, given the lack of explicit information on product labels and the prevalence of non-irrigated turf systems. Therefore, our reliance on natural precipitation represents a common – but not ideal – application practice for neonicotinoids in white grub control. In order to fully document application conditions, we recorded daily precipitation over a one-week period after each date of application along with other environmental parameters at the time of application (table 1). Overall, ≥2.0 mm of precipitation occurred within three days after each application date, with the exception of experiment 1, where it was delayed until the fifth day.
Table 1. Environmental conditions at time of application and precipitation post-application.

Japanese beetle egg predation
As one measure of natural predation rates, protocols were adapted from Terry et al. (Reference Terry, Potter and Spicer1993) to gather data on Japanese beetle egg removal. Newly laid, viable eggs were obtained from Japanese beetle adults that were collected in the vicinity using standard vane traps (Tanglefoot, Grand Rapids, MI) fitted with pheromone/floral lures (Spectracide Bag-a-bug floral lures, Spectrum Brands, Atlanta, GA). Beetles were provisioned with fresh wild grape leaves, Vitis vinifera L., and were held in wood-framed screen cages (0.5×0.5×0.5 m) with sandy-loam soil for oviposition. Newly laid eggs were separated from the soil using a coarse sieve and/or flotation in a 3M (17.4% w/v) solution of NaCl.
Predation trays were designed that would allow eggs to be implanted as naturally as possible into the soil profile of experimental field plots. Trays were constructed from the bottom of 50-mm diameter Petri dishes. A 45-mm diameter portion of the bottom was replaced with aluminum window screen (mesh size 1.13×1.30 mm) held in place with silicone caulk. Lined with a 42.5-mm diameter piece of filter paper, the resulting unit provided a stable surface for the placement of eggs, air and water exchange between the soil and eggs through the filter paper- and screen-lined bottom, and free access to soil-inhabiting arthropods through the open top. To measure egg removal rates, ten eggs were placed in each tray after moistening the filter paper with distilled water to promote adhesion. To implant a tray into the soil profile, a 5-cm diameter core was removed using a tubular turf-plugger (Turf Tec, Tallahassee, FL) set to a depth of 5 cm. One tray was placed flat onto the bottom of the hole left by the soil core, and then the core was gently replaced back into the hole without crushing the eggs or disrupting the tray. Five (2005) or six (2006, 2007) trays were implanted per experimental plot. In 2005, the insertion sites were approximately plot center and 2 m from each corner on the diagonal axes. In 2006 and 2007, however, the trays were placed at locations that were selected at random. After exposure for two days, trays were recovered to count the number of eggs remaining.
Impact on arthropods
To measure the activity-density of surface-active arthropods, five permanent pitfall traps were established in each plot. One trap was located in the center of the plot, the others one meter from each corner on the diagonal axes. Traps consisted of 1.1-l plastic cups (11 cm top diameter, 15 cm deep) buried flush with the surface in holes opened up by removing a 10.4-cm diameter core with a golf course cup cutter. Trapping was conducted over two-day intervals. A removable 0.54-l cup (11 cm top diameter, 7.5 cm deep) was placed inside the first buried cup to serve as the capture cup. The removable capture cup was filled with ≈3 cm of propylene glycol to retain and preserve arthropod captures. Traps were covered with tight fitting plastic lids during the intervals between sampling. The content of each trap was transferred to 70% ethyl alcohol for storage until taxonomic assessment.
To measure the density of soil-active arthropods, a series of 15 soil cores was extracted haphazardly from the surface of each plot with a 5-cm diameter turf plugger set to a depth of 5 cm. A 1-m border was excluded. Cores from each plot were grouped together, grass side down, in modified Tullgren funnels, each fitted with a 40 W light bulb. Samples were held three days under those conditions for extraction of invertebrates into 70% ethyl alcohol.
Invertebrates captured from pitfall traps and extracted from soil cores were sorted and enumerated. All were identified to class, hexapods to order, and some to family or other taxonomic grouping.
Short-term effects of imidacloprid on egg removal and nontarget arthropods (experiment 1)
A first experiment was conducted in 2005 to confirm protocols and substantiate the idea that a single application could impact egg removal rates. While no attempt was made to establish which taxa might be involved with egg removal, a suppressive effect of imidacloprid on nontarget taxa needed verification. Therefore, we also measured impacts on the incidence of soil- and surface-active fauna.
Treatments were applied 23 August 2005 to experimental plots arranged in a randomized complete block design with six repetitions of untreated and imidacloprid-treated plots. Egg predation, soil-active predators and surface-active predators were assessed at ten and 20 days after treatment (DAT). The egg predation protocols diverged from later studies in that trays were lined with a 1-mm layer of soil rather than filter paper. Furthermore, to deploy trays, a 10.4-cm diameter soil was removed to a depth of 15 cm; a horizontal slit was made in the side, just below the thatch layer; the tray was inserted; and the core was replaced. Egg removal rates were analyzed with analysis of variance (ANOVA) to test for an effect of treatment, and assessed with a type I error rate of 5% (P=0.05). Given departures from the assumption of normality, data on the abundance of soil-active arthropods and the abundance-activity of surface-active arthropods were analyzed with the nonparametric Wilcoxon rank-sum test. Given an a priori expectation of declines in the abundance of taxa in treated plots (Peck, Reference Peck2009a, Reference Peckb), a type I error rate of 10% (P=0.10) was used to assess all treatment-related fixed effects for the soil core captures. The two sampling dates were analyzed separately. In order to measure loss of eggs due to experimental manipulation, a series of ten additional trays were implanted just outside the experimental plots, left in place for 5 min, and then recovered. For this study and the following, statistical calculations were conducted with JMP Statistical Discovery software (SAS Institute, 2003) and means were reported with their standard error.
Comparative impacts of four neonicotinoids (experiment 2)
A second experiment was conducted in 2006 and 2007 to compare the impacts of clothianidin, dinotefuran, imidacloprid and thiamethoxam on egg removal rates. Field work was completed over a one-month interval in 2006 and repeated again in 2007 on a different set of plots. Application dates were 24 July 2006 and 31 July 2007. Egg predation was assessed at 1, 8, 15 and 22 DAT in each year, hereafter referred to as one, two, three and four weeks after treatment (WAT). Arthropods were not sampled in this experiment. Thiamethoxam was not available for inclusion at one WAT in 2006.
This study was conducted each year as a completely randomized design with six repetitions, and analyzed with ANOVA to test for an effect of neonicotinoid treatment with respect to WAT. Following a significant effect of WAT, differences among treatments were explored for each WAT using the least significant difference (LSD) means separation procedure. Analyses were assessed with a type I error rate of 5% (P=0.05). Given the magnitude of differences in egg removal rates between years, data were analyzed separately for each year.
Seasonal variation in the impact of imidacloprid (experiment 3)
A third experiment was conducted in 2006 and 2007 to compare the impact of imidacloprid when the application was made at different times in the season. While application timing is recommended for the period of scarab beetle oviposition, field efficacy has been demonstrated as early as late April (e.g. Heller & Kline, Reference Heller and Kline2007) and as late as early September (e.g. Heller & Kline, Reference Heller and Kline2005; Swier & Rollins, Reference Swier and Rollins2006). Fieldwork was conducted over a three-month interval in 2006 and repeated again in 2007. Applications were made in July, August and September, corresponding to 17 July, 14 August and 11 September in 2006, and 31 July, 28 August and 25 September in 2007. Each trial was conducted on a different set of plots. Egg removal rates were assessed at one, two, three and four WAT. Arthropods were not sampled in this experiment. Each month of application was arranged and analyzed in a randomized complete block design with six repetitions. Data were analyzed separately for each year to test for an effect of imidacloprid treatment with respect to WAT and month of application. The overall mixed model ANOVA included block as a random effect, with treatment, WAT, month and their interactions as fixed effects. Following a significant effect of treatment, further differences among fixed effects were explored using multiple means comparisons. Based on results from 2005 (experiment 1), a type I error rate of 10% (P=0.10) was used given an a priori expectation of declines in egg removal rates.
Results
Short-term effects of imidacloprid on nontarget arthropods (experiment 1)
From the pitfall traps, a total of 1448 individuals were captured, representing seven hexapod orders and three other arthropod classes. A partial listing of the numerically dominant taxa is provided in table 2. Several other taxonomic groups were detected but not included in the statistical analysis because of low numbers (n<20 pooled across both DAT). These included the hexapod orders Coleoptera (Scarabaeidae, Staphylinidae), Diptera, Hemiptera, Hymenoptera and Lepidoptera, plus the classes Diplopoda and Gastropoda (snails, slugs).
Table 2. Number of captures from pitfall traps in untreated and imidacloprid plots at ten and 20 d after treatment (DAT).

1 Each sample consisted of captures from five 11-cm diameter pitfall traps per plot, opened for two days.
2 Significant differences are highlighted with asterisks at P⩽0.10 (*), P⩽0.05 (**) and P⩽0.01 (***).
3 Suborder classification.
There was a significant effect of treatment on the number of total pitfall captures at ten DAT (S=21, Z=−2.81, P=0.0049) and 20 DAT (S=28, Z=−1.69, P=0.092). At ten DAT, the mean number of total captures was 30.5% lower in the imidacloprid plots (78.2±10.40) compared to the untreated plots (112.5±14.0). At 20 DAT, the mean number of total captures was 21.0% lower in the imidacloprid plots (22.3±2.0) compared to the untreated plots (28.3±5.0).
Among the individual taxa, at ten DAT, there was a significant reduction in arthropod abundance in imidacloprid plots compared to the untreated plots for captures of Symphypleona (95.3% reduction), Entomobryomorpha (83.3%), Porcellionidae (61.2%), non-myrmicine Formicidae (55.6%) and Orthoptera (20.3%) (table 2). In contrast, captures were significantly higher in the imidacloprid plots for non-lycosid Araneae and Carabidae. At 20 DAT, there was a significant reduction in arthropod abundance in imidacloprid plots for captures of Carabidae (71.4%) and Orthoptera (37.7%) but a significant increase in captures of non-lycosid Araneae, non-myrmicine Formicidae and Myrmicinae.
From the soil cores, a total of 8331 individuals were extracted, representing nine hexapod orders and five other arthropod classes. A partial listing of the numerically dominant taxa is provided in table 3. Several other taxonomic groups were detected but not included in the statistical analysis because of low numbers (n<20 pooled across both DAT). These included the hexapod orders Coleoptera (Bruchidae, Chrysomelidae, Curculionidae, Elateridae, Lathridiidae, Nitidulidae), Collembola (Symphypleona), Lepidoptera, Protura and Psocoptera, plus the classes Chilopoda, Diplopoda, Gastropoda (snails), Malacostraca (Isopoda), Oligochaeta (Enchytraeidae, Lumbricidae) and Symphyla. Considering only the untreated check, the extraction rates for ten and 20 DAT, respectively, were 14,811 and 11,827 individuals m−2 of turf and associated topsoil.
Table 3. Number of captures extracted from soil cores in untreated and imidacloprid plots at ten and 20 d after treatment (DAT).

1 Each sample consisted of Tullgren funnel extractions from 15 soil cores (diameter and depth 5 cm) per plot.
2 Significant differences are highlighted with asterisks at P⩽0.10 (*), P⩽0.05 (**) and P⩽0.01 (***).
3 Suborder classification.
As with pitfall traps, there was a significant effect of imidacloprid on the mean number of total captures at ten DAT (S=21, Z=−2.80, P=0.0051), but an effect was not apparent at 20 DAT (S=36, Z=−0.40, P=0.689). At ten DAT, the number of total captures was 43.4% lower in the imidacloprid plots (246.7±35.9) compared to the untreated plots (436.0±52.0). A difference was no longer detected at 20 DAT, when captures were nearly identical between treatments (357.7±96.8 imidacloprid, 348.2±66.8 untreated).
Among the individual taxa, at ten DAT, there was a significant reduction in arthropod abundance in imidacloprid plots compared to the untreated plots for Proisotoma (97.2%), Lasius (78.9%), Onychiurus (69.6%), Collembola (63.8%), Formicidae adults (62.3%), Pseudococcoidea (60.4%), total Hemiptera (58.7%), Oribatidae (41.7%) and Pseudosinella (30.0%). In contrast, captures of Araneae were significantly higher in the imidacloprid plots. At 20 DAT, there was a significant reduction in imidacloprid plots for Solenopsis molesta (79.8%), Aphididae (70.0%), Formicidae adults (69.5%) and Proisotoma (45.0%) but a significant increase for Oribatidae.
Short-term effects of imidacloprid on Japanese beetle egg removal (experiment 1)
For the two sampling dates, mean two-day egg removal rates in the untreated check were 62.7±8.2% at ten DAT and 93.0±4.4% at 20 DAT, or 77.8% when averaged across both DAT. While it was impossible to confirm that each egg was lost due to the action of predators, the loss due to experimental manipulation was close to negligible. In the test of implantation procedures, 99% of eggs were recovered.
There was a significant effect of imidacloprid on egg removal at ten DAT (F=8.3; df=1, 4; P=0.045) but not 20 DAT (F=0.0076; df=1, 5; P=0.934). At ten DAT, the mean removal rate in the untreated plots (62.7±8.2%) was almost twice that of the imidacloprid plots (32.8±5.2%). A difference was no longer detected at 20 DAT when the mean removal rates were nearly identical between treatments (93.0±4.4% untreated, 92.3±3.8% imidacloprid).
Comparative impacts of four neonicotinoids (experiment 2)
Egg removal rates were significantly lower in 2006 than in 2007 (F=14.9; df=1, 46; P=0.0003). Averaged across the four sampling dates, mean two-day egg removal rate in the untreated check was 41.3±4.8% in 2006 and 67.2±4.7% in 2007. In 2006 (fig. 1), there was a significant effect of week (F=29.71; df=3, 76; P<0.0001), treatment (F=5.05; df=3, 76; P=0.003), and week×treatment (F=2.25; df=9, 76; P=0.027). There was no difference in egg removal across the experimental plots at one WAT (F=1.23; df=3, 20; P=0.326). By two WAT, however, egg removal was significantly lower in all insecticide plots (F=3.88; df=4, 23; P=0.015). Clothianidin, dinotefuran, imidacloprid and thiamethoxam treatments reduced egg removal by 62.0, 43.5, 55.2 and 63.0% compared to the untreated plots. A significant egg removal effect remained at three WAT (F=5.73; df=4, 23; P=0.0024), but it was only expressed in imidacloprid and thiamethoxam plots, which reduced removal by 76.1 and 40.4%, respectively. By four WAT, there was no longer any effect of treatment on egg removal (F=0.76; df=4, 25; P=0.56). Based on the difference between the highest and lowest egg removal rates, variation among insecticides was only 7.0, 11.1, 24.4 and 17.2% at one, two, three and four WAT.

Fig. 1. Mean (±SE) removal of Japanese beetle eggs assessed in weeks one, two, three and four after treatment (WAT) in untreated plots and those treated with clothianidin (Cloth), dinotefuran (Dino), imidacloprid (Imi) and thiamethoxam (Thia) in 2006 and 2007. Thiamethoxam was not evaluated at one WAT in 2006. For each time, columns with different letters are significantly different (least-squares means comparisons, P⩽0.05).
In 2007 (fig. 2), there was a significant effect of week (F=37.72; df=3, 100; P<0.0001) and treatment (F=6.05; df=4, 100; P=0.0002), but not week×treatment (F=1.24; df=12, 100; P=0.27). A treatment effect on egg removal was already expressed at one WAT (F=3.57; df=4, 25; P=0.020). Dinotefuran, imidacloprid and thiamethoxam reduced removal by 41.8, 37.4 and 43.6%, compared to the untreated plots. At two WAT (F=3.52; df=4, 25; P=0.021) and three WAT (F=2.91; df=4, 25; P=0.042), respectively, clothianidin, dinotefuran and thiamethoxam reduced egg removal by 29.6, 31.5, 36.2% and 22.9, 16.5, 44.5%. There was an effect of imidacloprid at one WAT, but not at two or three WAT. As in 2006, by four WAT, there was no egg removal effect detected among treatments (F=0.24; df=4, 25; P=0.915). Based on the difference between the highest and lowest egg removal rates, variation among insecticides was only 12.5, 17.3, 9.7 and 8.5% at one, two, three and four WAT.

Fig. 2. Mean (±SE) removal of Japanese beetle eggs, with respect to year, week after treatment (WAT: 1–4) and treatment, pooled across three application dates (July, August, September). White bars are imidacloprid treated plots and black bars are untreated plots. For each WAT, significant differences are indicated with asterisks at P⩽0.10 (*), P⩽0.01 (**) and P⩽0.001 (***) (least-squares means comparisons).
Seasonal variation in the impact of imidacloprid (experiment 3)
As in experiment 2, egg removal rates were significantly lower in 2006 than 2007 (F=19.6; df=1, 143; P<0.0001). Averaged across the four WAT in 2006, mean two-day removal rates in the untreated check were 45.1±4.0% in July, 34.5±3.7% in August and 54.7±3.2% in September. Corresponding values in 2007 were 67.2±4.7, 59.1±4.1 and 53.6±4.2%.
The effect of imidacloprid on egg removal rates did not vary across month of application as there was no significant month×treatment interaction in either 2006 or 2007 (table 4). Pooled across WAT in 2006, egg removal in the imidacloprid plots compared to the untreated plots was suppressed 16.0, 7.5 and 28.3% in July, August and September, respectively. The corresponding values in 2007 were 16.7, 27.4 and 29.5%.
Table 4. Results from a multi-level ANOVA examining the effect of imidacloprid (treatment) on Japanese beetle egg predation with respect to month of application (July, August, September) and week after treatment (WAT: 1, 2, 3, 4) over two years.
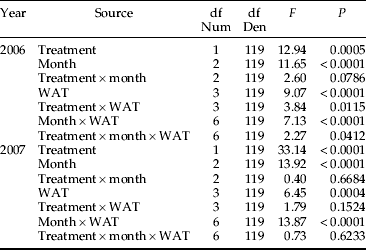
In contrast, there was a significant effect of treatment, month, WAT and month×WAT in both 2006 and 2007 (table 4). Unlike the results from experiment 2, egg removal was significantly reduced at one, two, three and four WAT (fig. 2). The only exception was three WAT in 2006. Relative to the untreated check, egg removal rates in the imidacloprid plots were reduced 39.2, 21.4, (−9.8) and 14.6% across the four WAT in 2006. The corresponding values in 2007 were 39.2, 25.8, 14.3 and 17.0%. The change in degree of reduction diverged from the trend revealed in experiment 2, where the impact of imidacloprid was strongest at two and three WAT and absent at four WAT.
Discussion
The main objective of this study was to determine if applications of neonicotinoid insecticides influence the rates of predation on Japanese beetle eggs. A difference in predation activity was expected because imidacloprid applications were recently documented as reducing the abundance of certain nontarget beneficial fauna in turfgrass (Peck, Reference Peck2009a,b) and because white grub eggs are included in the diet of generalist predators. As predicted, under the conditions of this study, the rate of egg removal decreased after a single neonicotinoid application, and the effect persisted for as long as 3–4 weeks. The results also matched predictions that the detrimental effects on predation would not vary with neonicotinoid chemistry nor with month of application.
Magnitude and persistence of impacts
Results confirm the idea that the natural regulation of pest populations is meaningful to the suppression of pest outbreaks and, thereby, favors the sustainability of turf habitats (Potter, Reference Potter, Racke and Leslie1993). In untreated plots, the natural rates of two-day egg removal varied from 41–78% when sampling dates were pooled across the five studies conducted from 2005 to 2007. Those rates were similar to other turfgrass studies that used implanted grub eggs in stands of Kentucky bluegrass. López & Potter (Reference López and Potter2000) measured 45–55% removal over 12 h of exposure on golf course roughs (mowing height 5 cm) and 2–38% on fairways (mowing height 3 cm). Kunkel et al. (Reference Kunkel, Held and Potter1999) measured 21–48% removal over three days, and Terry et al. (Reference Terry, Potter and Spicer1993) measured 35–75% removal over two days in experimental lawns (mowing height 5 cm). After an imidacloprid application, however, declines in egg removal rates were detected in all five studies conducted from 2005 to 2007, ranging as high as 28.3–76.1%. Egg removal was disrupted as soon as one WAT and was expressed for at least four WAT; and, because sampling was discontinued at four WAT, it is possible disruption could have lasted longer. There is a degree of incongruence in the long residual of neonicotinoids and the instances when the disruptive effect was brief, such as in 2005 when egg predation rates recovered in imidacloprid plots between ten and 20 DAT. This is more consistent with a short-term acute response mediated through some form of direct versus systemic exposure. The mobility of generalist predators, such as ants, ground beetles and rove beetles, means they could respond relatively rapidly to acute insecticidal effects through dispersal between treated and untreated areas. While the magnitude of disruption on egg removal rates measured here is probably meaningful for the regulation of pest populations, ascertaining the practical relevance for turf management will depend on future studies.
Seasonal variation
No significant variation was detected in the effect on egg removal when applications were made at different times in the season, i.e. from July to September. Although recommended for application during the period of oviposition, imidacloprid has a fairly forgiving window of treatment because of its long residual. In the Northeast, it can be effective when applied from early June to mid August, but some individual studies do show efficacy as a Japanese beetle curative as late as early September (e.g. Heller & Kline, Reference Heller and Kline2005; Swier & Rollins, Reference Swier and Rollins2006). Regardless, due to the long residual of neonicotinoids, there is the possibility of modifying the timing of applications in order to promote control efficacy as well as to mitigate detrimental outcomes such as nontarget effects. In their study on the impact of imidacloprid to a specialist parasitoid, Rogers & Potter (Reference Rogers and Potter2003) showed that early preventive treatments significantly interfered with the activities of Tiphia vernalis (Hymenoptera: Tiphiidae), but late preventive treatments did not. A group of generalist predators, not individual species of specialist enemies, are the likely agent of egg removal in our studies (Terry et al., Reference Terry, Potter and Spicer1993; Kunkel et al., Reference Kunkel, Held and Potter1999; López & Potter, Reference López and Potter2000). If predator abundance and activity varies relatively little over the application season, it could explain the lack of a seasonal effect, and make it less likely that adjustments in the timing of neonicotinoid applications could mitigate the disruptive effects on egg predation.
Neonicotinoid variation
No meaningful or consistent variation was detected among the impacts of clothianidin, dinotefuran, imidacloprid and thiamethoxam on egg removal. Across WAT and years, variation among insecticides only ranged from 7.0 to 24.4%. A treatment effect was detected as early as one WAT for dinotefuran, imidacloprid and thiamethoxam in 2007, but not until two WAT for all other trials. A treatment effect only persisted through one WAT for imidacloprid in 2007 and two WAT for clothianidin and dinotefuran in 2006, but through three WAT for all other trials. Beyond physical and toxicological attributes of their chemical class, functional similarities among these products include systemicity and relatively long residual (Tomizawa & Casida, Reference Tomizawa and Casida2003; Jeschke & Nauen, Reference Jeschke and Nauen2008). These neonicotinoids, however, do vary in attributes like translocation and translaminar activity (Buchholz & Nauen, Reference Buchholz and Nauen2001; Elbert et al., Reference Elbert, Nauen, Leicht, Ishaaya and Degheele1998). Based on our results, differences among those chemistries may have no meaningful relevance for the degree to which pest suppression is disrupted. Whether that translates to similar population level impacts on the nontarget arthropods that contribute to egg removal is unclear. Oliver et al. (Reference Oliver, Mannion, Klein, Moyseenko and Bishop2005) revealed a negative effect of soil-incorporated thiamethoxam, but not imidacloprid, on Tiphia parasitoid development. Although scarce, studies from other agroecosystems reveal differences in the impacts of neonicotinoid chemistries on the abundance of nontarget arthropods (Kilpatrick et al., Reference Kilpatrick, Hagerty, Turnipseed, Sullivan and Bridges2005).
Identity of egg predators
This study did not identify which nontarget taxa contributed to the removal of Japanese beetle eggs. Formicidae (ants) and the beetle families Carabidae, Cicindelidae, Histeridae and Staphylinidae are confirmed predators of grub eggs (Terry et al., Reference Terry, Potter and Spicer1993; López & Potter, Reference López and Potter2000). One study on home lawns identified 17 species of carabids alone (Rochefort et al., Reference Rochefort, Shetlar and Brodeur2006), indicating the potential diversity of natural enemies in turf systems. Our results did reveal some negative impacts on Carabidae and Formicidae, but the overall evidence was mixed and inconclusive. Based on pitfall sampling, in imidacloprid plots the abundance-activity of Carabidae was higher at ten DAT but lower at 20 DAT. In contrast, non-myrmicine Formicidae was lower at ten DAT, but myrmicine and non-myrmicine Formicidae were higher at 20 DAT. Egg scavenging might occur in two other groups negatively impacted by imidacloprid. Porcellionidae (sowbugs) was lower at ten DAT, while Orthoptera (largely Gryllidae) was lower at both DAT. Based on soil core extractions, population effects were consistently negative among the only group of affected potential predators, Formicidae. Abundance was lower for total ant adults and Myrmicinae at both DAT and for Lasius spp. at ten DAT. Thysanoptera might also contribute to egg removal as scavengers or facultative predators; their abundance was lower at ten DAT.
Previous work documented a similar density of soil-active arthropods extracted from soil cores (7616 to 25,826 individuals m−2) but did not reveal any impacts of single imidacloprid applications (Peck, Reference Peck2009a). In that study, however, a relatively long post-application sampling interval (2–3 wk) could have obscured any response. Over the long-term, six consecutive seasons of imidacloprid applications led to a significant reduction in two largely predaceous beetle families (Carabidae and Staphylinidae) but not in two largely phytophagous families (Chrysomelidae and Curculionidae) (Peck, Reference Peck2009b). Therefore, it was proposed that an imidacloprid-mediated decrease in Collembola populations led to a decrease in predatory beetle populations through the removal of food resources. While the results presented here do not confirm a definitive imidacloprid effect on the Carabidae, they do indicate an effect on the Collembola. As measured through soil cores extractions, the abundance of Onychiurus, Proisotoma, Pseudosinella and Symphypleona was 3.3-, 28.0-, 1.4- and 5.0-fold lower at ten DAT. Abundance recovered by 20 DAT, with the exception of Proisotoma 9.3-fold lower.
The specific identity of predators remains a black box and an obvious subject of future research, as is how a neonicotinoid effect is channeled through them to impact egg removal rates. It is likely that a suite of generalist natural enemies will have to be addressed. The fact that removal rates did not vary across season is consistent with activity from a complex of generalist predators that would be less variable through time, rather than from a few individual species.
Post-application irrigation
Our study was not designed to establish how the timing, amount and variation in post-application rainfall influenced the outcome of our results. Given the overall decline in precipitation from 2006 to 2007, however, it was feasible to assess whether differences in treatment effects may have been attributable to differences in post-application precipitation (table 1). In experiment 3, the week after application in the July trials the total rainfall declined considerably from 2006 (57.5 mm) to 2007 (2.5 mm); however, the suppressive effect of imidacloprid on egg predation was nearly identical (16.0 and 16.7%, respectively). In contrast, a similar decline in precipitation from 2006 (18.0) to 2007 (0.5 mm) in the August trials was associated with a higher degree of variation in egg predation suppression (7.5 and 27.5%, respectively). In experiment 2 (fig. 1), post-application rainfall also declined from 2006 (18.6 mm) to 2007 (2.5 mm); but, again, there was no discernible trend that linked variation in rainfall data with the detection or magnitude of treatment effects on egg predation rates.
Implications
We recommend that future studies examine not only how application timing and irrigation affect insecticide performance, but how they might be manipulated to mitigate the negative consequences posed by long-residual neonicotinoid insecticides on the abundance and function of nontarget arthropods. When insecticides target soil insects in turf, a general recommendation is to follow with irrigation so material moves from the foliage to the target zone before any degradation due to wind and sun exposure (Potter, Reference Potter1998). As post-application irrigation is linked to improved performance against white grubs for at least imidacloprid, irrigation might also influence the scope of nontarget effects. Kunkel et al. (Reference Kunkel, Held and Potter2001), for instance, showed that post-application irrigation reduced the deleterious behavioral effects of granular imidacloprid on surface-active generalist predators because of decreased insecticide exposure. Under the conditions of our study, post-application irrigation was uncontrolled as it relied on natural precipitation. Similar application scenarios may be widespread in the northeast US. On the labels of all five products evaluated in our study, recommendations on post-application irrigation are variably inexplicit, which may contribute to poor adoption. Moreover, this practice may not even be achievable under many circumstances given the prominence of non-irrigated turf systems. In New York state, for instance, 85% (9740 km2) of residential lawns are non-irrigated, as well as 62–63% of turf on golf courses and sod farms and 81–87% on park and school grounds (NASS, 2004).
Neonicotinoids (at least imidacloprid) are recommended for application at the time of Japanese beetle oviposition, which puts their unintended effects (disruption of egg predation) in direct conflict with their intended effects (mortality of neonate white grubs). We speculate on one scenario whereby the role of natural enemies could be capitalized within a management program that relies on intervention with long-residual insecticides. Delaying imidacloprid applications means more chances to scout, assess thresholds and spot-treat smaller areas. The concomitant decline in efficacy with increasing white grub instar could be buffered by conservation of predation on early life stages. Early application, on the contrary, may not be a promising approach. While imidacloprid can be highly effective when applied up to eight weeks before oviposition (e.g. Heller & Kline, Reference Heller and Kline2007), the marginal increase due to predation might be inconsequential.
Under scenarios that include a diverse local fauna and low pest pressure, it might be reasonable to expect that the activities of natural enemies could supplant much of the reliance on preventive insecticides. Using insecticides in a sound and effective manner, consistent with IPM principles, is a priority for environmental stewardship in turf. It will conserve the role of beneficials, reduce groundwater contamination, proactively assuage public concerns over usage, and safeguard the long-term use of neonicotinoids as effective and low risk alternatives for insect pest management. Refining our understanding of the balance between the intended and unintended consequences of preventive insecticides will enhance opportunities for pest management practitioners to harness the benefits of natural pest control.
Acknowledgements
We thank A. Corichi, E. Jennings, A. Mazo, A. Morales, K. Nelson and C. Ospina for assistance in the laboratory and field. For agronomic advice and support, we thank B. Arehart, S. Gordner, F. Rossi and M. Thurne. Ant species were initially identified by C. Klass. Three anonymous reviewers and G. Loeb made comments that greatly improved the manuscript.






