Introduction
Pearl millet [Pennisetum glaucum (L.) R. Br.] is one of the important cereal crops grown in a wide range of latitudes (35°S to 35°N of the equator). It is an important grain crop in Africa and Asia and a fodder crop in other arid and semi-arid parts of the world (Andrews and Anand Kumar, Reference Andrews and Anand Kumar1992). It is mainly cultivated in Niger, Nigeria, Burkina Faso, Togo, Ghana, Mali, Senegal, Central African Republic, Cameroon, Sudan, Botswana, Namibia, Zambia, Zimbabwe and South Africa (Africa) and India, Pakistan and Yemen (Asia) (Upadhyaya et al., Reference Upadhyaya, Reddy, Irshad Ahmed and Gowda2010).
Latitude, which is the measurement of the distance of a location on earth from the equator, is the primary factor that affects unequal heating of the earth's atmosphere. Latitudinal gradient in species diversity has been recognized for nearly a century, and some of these polar–equatorial trends have been discussed in detail elsewhere (Darlington, Reference Darlington1959; Fischer, Reference Fischer1960). Plants experience considerable variations in natural selection across their latitudinal range for local adaptation, resulting in geographic differentiation of populations (Joshi et al., Reference Joshi, Schmid, Caldeira, Dimitrakopoulos, Good, Harris, Hector, Huss-Danell, Jumpponen, Minns, Mulder, Pereira, Prinz, Scherer-Lorenzen, Siamantziouras, Terry, Troumbis and Lawton2001; Streisfeld and Kohn, Reference Streisfeld and Kohn2005; Springer, Reference Springer2007). Since solar irradiance, temperature and photoperiod changes are known to influence many aspects of plant life (Berry and Raison, Reference Berry, Raison, Lange, Nobel, Osmond and Ziegler1981), widely distributed plant species may be expected to show phenotypic variability across latitudes. Genetically based latitudinal variations in phenology (Weber and Schmid, Reference Weber and Schmid1998), growth (Chapin and Chapin, Reference Chapin and Chapin1981) and sexual reproduction (Aizen and Woodcock, Reference Aizen and Woodcock1992) have been reported in terrestrial species. The overall pattern of diversity of crops including pearl millet largely depends on natural and human selection and sensitivity to environmental factors such as day lengths, minimum and maximum temperatures, rainfall patterns, soils, etc.
The world collection of pearl millet germplasm (22,211 accessions) conserved at the ICRISAT genebank, Patancheru, India is obtained from regions at a wide range of latitudes (0°–34.37° on both sides of the equator). Due to its largest collection, the study of latitudinal patterns of diversity in the collection of pearl millet germplasm becomes very important for its targeted use in crop improvement programmes. The availability of a geographic information system and climate data of collection sites have opened up avenues in understanding latitudinal patterns of diversity. Therefore, in the present study, patterns of diversity for important morpho-agronomic traits in pearl millet landraces from different latitudes were assessed and discussed in relation to the available information on climate in different latitudinal ranges on either side of the equator.
Materials and methods
The passport information and characterization data of pearl millet germplasm (22,211 accessions) conserved at the ICRISAT genebank, Patancheru, India were used in the present study. The collection material consisted of 19,063 landraces, 2269 breeding materials, 129 improved cultivars and 750 wild accessions belonging to 24 species of the genus Pennisetum. Passport information of the landraces, particularly the location of collection sites and corresponding geographic coordinates, was updated by referring to all related records, collection reports and catalogues. Using Microsoft EncartaR, an electronic atlas (MS EncartaR Interactive World Atlas, 2000), geographic coordinates were retrieved for accessions without coordinates to fill the gaps for landraces having location information. The accuracy of coordinates was verified by plotting all the landraces on a political map of world. Finally, a set of 15,904 landraces having geographic coordinates was used in the present study. The collection material includes landraces from regions at latitudes ranging between 0° and 34.37° on both sides of the equator, covering almost all major pearl millet-growing countries across the world.
Pearl millet landraces were characterized in batches of 500 to 1000 every year at ICRISAT, Patancheru (17.53°N latitude, 78.27°E longitude and 545 m.a.s.l), in alfisols during the rainy and post-rainy seasons from 1974 to 2011. These two different seasonal conditions are typical to semi-arid regions (Reddy et al., Reference Reddy, Kameshwara Rao and Irshad Ahmed2004). During the rainy season, the accessions were sown in the month of June and harvested in the month of October/November. In contrast, during the post-rainy season, the accessions were sown in the month of November and harvested in the month of March of the subsequent year. At Patancheru, the day length decreases from 13.10 h (in June) to 11.40 h (in November) in the rainy season and increases from 11.10 h (in December) to 12.00 h (in March) in the post-rainy season. The monthly mean minimum temperature varies from 23.6°C (in June) to 16.0°C (in November) and the monthly mean maximum temperature ranges from 34.4°C (in June) to 28.9°C (in November) in the rainy season. During the post-rainy season, the monthly mean minimum temperature increases from 12.9°C (in December) to 19.3°C (in March) and the mean maximum temperature increases from 27.9°C (in December) to 35.2°C (in March). The mean annual rainfall is 908 mm. Each accession was grown in two rows of 4 m length each with a spacing of 75 cm between the rows and 10 cm between the plants within a row, accommodating a total of 80 plants in two rows. The accessions were randomized in all the evaluations. Fertilizers were applied at the rate of 100 kg N and 40 kg P2O5/ha. Need-based irrigations were applied during the rainy season, while the crop was irrigated at regular intervals during the post-rainy season. The crop was protected from weeds, pests and diseases. By the end of 2011, almost all landraces were characterized for 19 morpho-agronomic characteristics following the descriptors mentioned previously (IBPGR and ICRISAT, 1993). Days to 50% flowering, plant height, panicle length and thickness were recorded during both rainy and post-rainy seasons, whereas number of total and productive tillers, panicle shape, spikelet density, bristle length, seed yield potential, green fodder yield potential and overall plant aspect were recorded only during the rainy season. For panicle density, bristle length, seed yield potential, green fodder yield potential and overall plant aspect, landraces were visually scored on a scale of 1 to 9, where 1 indicates most undesirable and 9 most desirable. Observations on grain characteristics, such as 1000-seed weight, seed shape and colour, were recorded after harvesting during the post-rainy season. Emergence of stigma in 50% of plants in a plot (accession) was considered as days to 50% flowering (IBPGR and ICRISAT, 1993). The height from the base to the tip of the panicle was recorded as plant height (cm). In each plot, the number of total and productive tillers per plant was counted and the mean of five plants was recorded. The mean panicle length (cm) over five plants was recorded, while the width of the panicle (mm) was recorded at the maximum thickness of the panicle. Weight of 1000 seeds (g) drawn from the plot harvest was recorded. The green fodder yield potential of accessions was scored on the scale of 1–9, considering plant height, tillering and leafiness of accessions, while the seed yield potential of accessions was scored on the basis of the number of productive tillers, spikelet density, panicle length and thickness and seed size. The overall plant aspect was recorded based on important agronomic traits.
Frequencies were estimated for all qualitative traits under study for the total collection, Northern Hemisphere (NH) and Southern Hemisphere (SH) and for each latitude range with an interval of 5.00° in both NH and SH. Climatic data, such as monthly mean minimum and maximum temperatures (over the past 30 years), rainfall patterns and day lengths for each collection site, were retrieved from the website http://www.worldclim.org/current using the spatial analyst extension in ArcGIS® software in June 2011 (Hijmans et al., Reference Hijmans, Cameron, Parra, Jones and Jarvis2005). High-resolution (1 km) interpolated climate surface data are useful for studying the spatial relationship between environmental variables and the vegetation existing at that location. The lowest and highest monthly mean minimum and maximum temperatures and day lengths for the entire collection and for the NH and SH, as well as lower ( < 20°) and higher (>20°) latitudes and each latitude range with an interval of 5.00° starting from 0.00° to 35.00° on either side of the equator, were estimated.
Agronomic data were analysed using the residual maximum likelihood procedure in Genstat 14 release (http://www.vsni.co.uk) to partition genotypic variance into between hemispheres, lower ( < 20°) and higher (>20°) latitudes, and latitude ranges within hemispheres. The respective standard errors were estimated and used to determine the significance of variance components. Means, ranges and variances were calculated for 11 quantitative characteristics, for the entire collection, hemispheres, lower and higher latitudes and latitude ranges within each hemisphere. The mean values of different traits were compared using the Newman–Keuls procedure (Newman, Reference Newman1939; Keuls, Reference Keuls1952). The homogeneity of phenotypic variances was tested by Levene's (Reference Levene and Olkin1960) test. The Shannon and Weaver (Reference Shannon and Weaver1949) diversity index (H′) was used to measure and compare the phenotypic diversity of all the 11 traits for the entire collection, hemispheres, lower and higher latitudes and all latitude ranges within hemispheres. Measured values of individual accessions for a given trait were standardized by subtracting the mean value of the trait from each observation and subsequently dividing by its standard deviation. This resulted in standardized values for each trait with an average value of 0 and a standardized deviation of 1 or less. The principal component analysis (PCA) was carried out in Genstat 14 release using these standardized values. The cluster analysis (Ward, Reference Ward1963) was carried out using the scores of the first three principal components (PCs) to cluster different latitudinal ranges.
Results
Frequency distribution
Pearl millet landraces used in the present study were collected from regions at a wide range of latitudes from 33.00° in the SH to 34.37° in the NH. The frequency distribution of the landraces indicated 80.5% (12,808) of landraces from the NH and 19.5% (3096) of landraces from the SH (Table 1). In both hemispheres, more landraces were found at lower latitudes ( < 20°; 9573 in the NH and 2687 in the SH) than at higher latitudes (>20°; 3235 in the NH and 409 in the SH). The frequency distribution over the entire collection and for each latitude range on either side of the equator indicated that the latitude range of 10°–15° on the northern side and 15°–20° on the southern side of the equator were found to be important source regions for the prevalence of pearl millet with 39.6% and 13.1% accessions, respectively. The proportion of accessions at latitudes close to the equator (0°–5°) and at higher latitudes (30°–35°) is very low (~1%) (Table 1).
Table 1 Range of variation in various traits of pearl millet germplasm from different latitudes on both sides of the equator, evaluated at ICRISAT, Patancheru, India

Lat, latitude; DFLR, days to 50% flowering in the rainy season; DFLPR, days to 50% flowering in the post-rainy season; PHTR, plant height (cm) in the rainy season; PHTPR, plant height (cm) in the post-rainy season; TTR, total tillers/plant in the rainy season; PTR, productive tillers/plant in the rainy season; PLR, panicle length (cm) in the rainy season; PLPR, panicle length (cm) in the post-rainy season; PWR, panicle width (mm) in the rainy season; PWPR, panicle width (mm) in the post-rainy season; SDWT, 1000-seed weight; EC, entire collection; NH, Northern Hemisphere; SH, Southern Hemisphere; NH-HL, higher-latitude Northern Hemisphere (>20°); NH-LL, lower-latitude Northern Hemisphere ( < 20°); SH-HL, higher-latitude Southern Hemisphere (>20°); SH-LL, lower-latitude Southern Hemisphere ( < 20°).
Diversity for qualitative traits
Panicle shape
Panicle shape recorded on the basis of the majority of panicles in the plot is useful for classifying the landraces. Nine panicle shapes (cylindrical, conical, spindle, club, candle, dumb-bell, lanceolate, oblanceolate and globose) are recognized in the collection (IBPGR and ICRISAT, 1993) (Supplementary Table S2, available online). Landraces producing candle-shaped panicles were more common in the entire collection, both hemispheres and all latitude ranges except for 25°–30° in the NH and 0°–15° and 20°–25° in the SH. The latitude range of 25°–30° in the NH and 10°–15° in the SH were found to be important source regions for pearl millet with cylindrical-shaped panicles, with 43.4% and 48.9% accessions, respectively. Landraces producing lanceolate-shaped panicles were predominant in the latitude ranges of 0°–10° and 20°–25° in the SH.
Seed colour
Seed colour is an important trait that differentiates landraces. Ten seed colours (ivory, cream, yellow, grey, deep grey, grey brown, brown, purple, purplish black and mixture of grey and white) were found in the collection, and 50% of the total landraces produced grey colour seeds and 32% of the landraces produced grey-brown colour seeds (IBPGR and ICRISAT, 1993) (Supplementary Table S2, available online). All other colours were negligible ( < 8%) in the collection. Both hemispheres and all latitude ranges were found to be predominant source regions for grey to grey-brown seeds. Seed colours such as ivory and cream were more predominant at latitudes of 0°–15°N and 10°–20°S. Seeds with yellow pericarp were predominant at latitudes of 5°–15°N.
Seed shape
Five seed shapes (obovate, oblanceolate, elliptical, hexagonal and globular) were found in the collection (IBPGR and ICRISAT, 1993) (Supplementary Table S2, available online). A maximum of 25.5% of landraces produced globular-shaped seeds followed by 23.8% elliptical-, 18% oblanceolate-, 17.3% obovate- and 15.5% hexagonal-shaped seeds. The NH was the predominant source for globular-, hexagonal- and elliptical-shaped seeds when compared with the SH, which was found as the important source for obovate- and oblanceolate-shaped seeds. The latitude ranges of 10°–15°N (39.2%) and 5°–10°N (46.5%) were found to be important source regions for globular-shaped seeds. In the SH, the latitudes of 0°–15°, 15°–20° and 20°–25° were found to be important source regions for obovate-, oblanceolate- and globular-shaped seeds, respectively.
Spikelet density
Spikelet density or panicle density is an important trait contributing to the yield of pearl millet. A high score of 7 to 9 indicated high panicle density, which is desirable. Only 12.8% of the landraces in the collection had a high score (7–9). There were 215 (1.35%) landraces in the collection that scored 8 and five (0.03%) landraces scored 9 for this trait. The NH was found to be the predominant source for pearl millet landraces with a high score for this trait. Among the latitude ranges, 5°–20° on both sides of the equator was found to be important source regions for high-scoring landraces for this trait (Supplementary Table S2, available online).
Bristle length
A high score for bristle length indicates long bristles, which is a desirable trait. The long bristles of the panicle will penetrate into the eye of the bird, acting as a self-defence mechanism to scare birds. More than 96% of the total landraces produced short bristles (bristles below the level of the apex of the seed). In contrast, 0.47% (75 landraces) of the total collection produced bristles longer than 2 cm above the seed and scored 7–9. All landraces scoring 9 were from the SH. Within the SH, the latitudes of 10°–15° and 20°–25° were the important source regions for pearl millet with long bristles (Supplementary Table S2, available online).
Green fodder yield potential
This trait determines the fodder yield potential of landraces. In the entire collection, about 41% of the landraces had scored 7, whereas 12% scored 8 and 0.7% scored 9 for green fodder yield potential. More than 50% of the landraces from both hemispheres had scored more than 6. Within the NH, a maximum of 4.0% of the landraces from the latitudes of 5°–10° had scored 9 (Supplementary Table S2, available online).
Seed yield potential
This is an important trait contributing to the seed yield of pearl millet. A maximum of 43% of the landraces scored 6, followed by 14% of the landraces scoring 7, 0.96% scoring 8 and 0.01% scoring 9. All the landraces that scored 8 and 9 were from the NH. All the landraces scoring 9 were from the latitudes of 10°–15°N and 20°–25°N (Supplementary Table S2, available online).
Overall plant aspect
This trait indicates the overall agronomic acceptability of landraces (Supplementary Table S2, available online). None of the landraces in the collection scored 9. In the entire collection, only 0.54% landraces scored 8. Only 0.66% landraces from the NH and 0.03% from the SH scored 8. However, in the NH, landraces scoring 8 occurred at all latitude ranges.
Diversity for quantitative traits
Range and means
A large variation was observed in the collection for all quantitative traits under study. Landraces from the NH varied widely for all traits except for panicle width when compared with those from the SH (Table 1). Landraces from lower latitudes on either side of the equator varied widely for all traits. Among the latitude ranges, 10°–15°N showed a high variation for all traits under study except for panicle width. Earliest flowering landraces were from the latitude of 20°–25°N. Very late flowering (>150 d in the rainy season and >130 d in the post-rainy season), tall (>480 cm in the rainy season and >420 cm in the post-rainy season), high-tillering (35 tillers per plant in the rainy season and >17 tillers in the post-rainy season), long (>100 cm) and thick panicle (>50 mm) and large seed (>18 g/1000 seeds) landraces were from the latitude of 10°–15°N.
The Newman–Keuls (Newman, Reference Newman1939; Keuls, Reference Keuls1952) test of significance for mean values indicated significant differences between the landraces from the two hemispheres for all traits except for days to 50% flowering in the rainy season (Table 2). Landraces from the NH flowered early in the post-rainy season, grew short, produced more tillers and had small, thin panicles and large seeds when compared with those from the SH. Higher and lower latitudes within each hemisphere also differed significantly for all traits. Landraces from higher latitudes in the NH flowered early in both seasons, grew short in the rainy season and tall in the post-rainy season, and produced more tillers with small panicles and small seeds. In contrast, in the SH, landraces from higher latitudes flowered early in the rainy season and late in the post-rainy season, grew short, and produced more tillers with small panicles and small seeds. Significant differences were observed between the latitude ranges in the NH for all traits. Landraces from the latitude of 5°–10°N flowered late and grew tall in both seasons and produced more tillers. Landraces from the latitude of 10°–15°N produced few tillers and had long and thick panicles with larger seeds. Landraces from higher latitudes (30°–35°N) flowered early and grew to a short height in the rainy season (Table 2).
Table 2 Meana values for various traits of pearl millet germplasm from different latitudes on both sides of the equator, evaluated at ICRISAT, Patancheru, India

Lat, latitude; DFLR, days to 50% flowering in the rainy season; DFLPR, days to 50% flowering in the post-rainy season; PHTR, plant height (cm) in the rainy season; PHTPR, plant height (cm) in the post-rainy season; TTR, total tillers/plant in the rainy season; PTR, productive tillers/plant in the rainy season; PLR, panicle length (cm) in the rainy season; PLPR, panicle length (cm) in the post-rainy season; PWR, panicle width (mm) in the rainy season; PWPR, panicle width (mm) in the post-rainy season; SDWT, 1000-seed weight; EC, entire collection; NH, Northern Hemisphere; SH, Southern Hemisphere; NH-HL, higher-latitude Northern Hemisphere (>20°); NH-LL, lower-latitude Northern Hemisphere ( < 20°); SH-HL, higher-latitude Southern Hemisphere (>20°); SH-LL, lower-latitude Southern Hemisphere ( < 20°).
a Means were tested by Newman–Keuls test and means followed by different letters are significantly different at P= 0.05.
Variances
The homogeneity of variances of the hemispheres and latitude ranges within each hemisphere was tested for all the 11 quantitative traits by Levene's (Reference Levene and Olkin1960) test (Supplementary Table S1, available online). The variances for hemispheres were heterogeneous (P≤ 0.0001) for all traits under study and for all traits in the lower and higher latitudes on either side of the equator. Variances were heterogeneous (P ≤0.0001) for all traits, except for total and productive tillers per plant, at all latitude ranges in the NH and for all traits in the SH.
Phenotypic diversity
The Shannon–Weaver diversity index (H′) (Shannon and Weaver, Reference Shannon and Weaver1949) was calculated for the entire landrace collection, collections from the two hemispheres, lower and higher latitudes in each hemisphere and latitude ranges within hemispheres, to compare the phenotypic diversity of the 11 quantitative traits (Table 3). A low H′ value indicates extremely unbalanced frequency classes for an individual trait and a lack of genetic diversity in the collection or species. In the entire collection, the H′ value ranged from 0.427 ± 0.020 for total tillers per plant to 0.632 ± 0.020 for plant height in the post-rainy season. Among the hemispheres, mean diversity over all the traits was more (H′ = 0.569 ± 0.019) in the landraces from the NH than those from the SH (H′ = 0.563 ± 0.027). In the NH, diversity was high at higher latitudes for all the traits under study except for flowering in the post-rainy season, panicle width in the rainy season and 1000-seed weight. Within each hemisphere, mean diversity over all the traits was high at higher latitudes when compared with that at lower latitudes. Among the latitude ranges, the diversity index value for all the traits ranged from 0.571 ± 0.026 in the latitude of 10°–15° to 0.606 ± 0.009 in the latitude of 25°–30° in the NH and from 0.276 ± 0.001 in the latitude of 30°–35° to 0.583 ± 0.017 in the latitude of 20°–25° in the SH.
Table 3 Shannon–Weaver diversity index (H′) for various traits of pearl millet germplasm from different latitudes on both sides of the equator, evaluated at ICRISAT, Patancheru, India

Lat, latitude; DFLR, days to 50% flowering in the rainy season; DFLPR, days to 50% flowering in the post-rainy season; PHTR, plant height (cm) in the rainy season; PHTPR, plant height (cm) in the post-rainy season; TTR, total tillers/plant in the rainy season; PTR, productive tillers/plant in the rainy season; PLR, panicle length (cm) in the rainy season; PLPR, panicle length (cm) in the post-rainy season; PWR, panicle width (mm) in the rainy season; PWPR, panicle width (mm) in the post-rainy season; SDWT, 1000-seed weight; EC, entire collection; NH, Northern Hemisphere; SH, Southern Hemisphere; NH-HL, higher-latitude Northern Hemisphere (>20°); NH-LL, lower-latitude Northern Hemisphere ( < 20°); SH-HL, higher-latitude Southern Hemisphere (>20°); SH-LL, lower-latitude Southern Hemisphere ( < 20°).
Cluster analysis
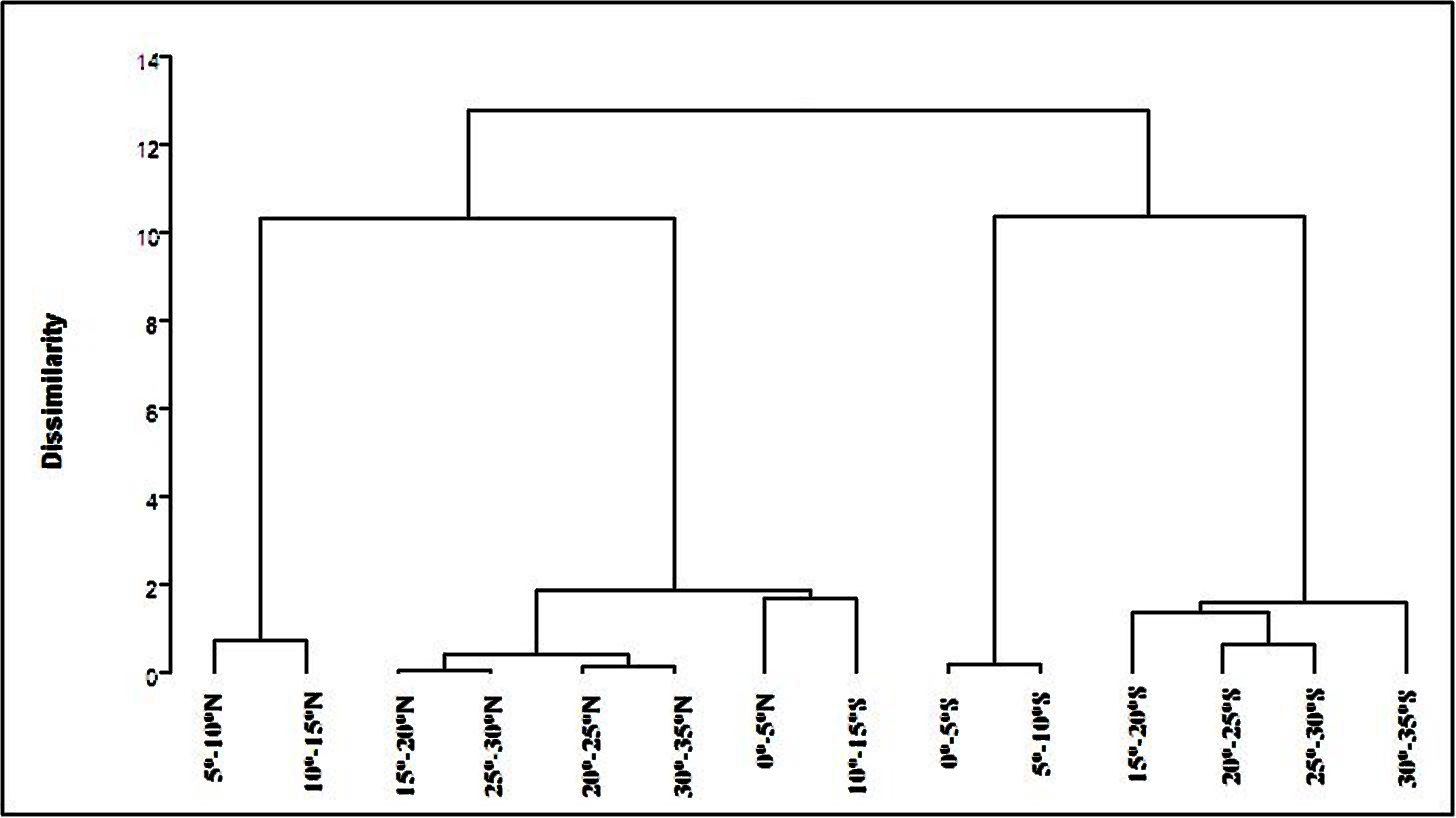
The PCA carried out using the standardized data of the 11 quantitative traits captured 90% of the total variation from the first three PCs. A hierarchical cluster analysis conducted on the scores of the first three PCs resulted in four clusters of latitude ranges. Landraces from the latitude of 5°–15°N formed cluster 1, those from 0°–5°N, 15°–35°N and 10°–15°S formed cluster 2, those from 0°–10°S formed cluster 3 and those from 15°-35°S formed cluster 4 (Supplementary Fig. S1, available online).
Climate
Day length
The mean annual day length over all the collection sites was 12.12 h. The mean annual day length in the NH (12.13 h) differed significantly from that of the SH (12.07 h). Significant differences were observed in day length at higher and lower latitudes in both hemispheres. In the NH, day length was high (12.15 h) at higher latitudes than at lower latitudes (12.12 h). It was observed that in both hemispheres, as the latitude range increased, the minimum, maximum and mean of the lowest monthly mean for day length decreased, whereas the minimum, maximum and mean of the highest monthly mean increased.
Rainfall
The mean annual total rainfall over all the collection sites was 737 mm. Collection sites in the NH received a significantly higher (753 mm) rainfall than those in the SH (673 mm) (Table 4). Lower latitudes in both hemispheres received a significantly higher rainfall (772 mm in the NH and 686 mm in the SH) than higher latitudes (697 mm in the NH and 580 mm in the SH). No clear patterns exist in the rainfall of different latitudinal ranges in both hemispheres. Only the latitude of 0°–10°N received more than 1000 mm rainfall. All latitude ranges in the SH received < 1000 mm rainfall with a maximum of 982 mm in the latitude of 10°–15°.
Table 4 Range of variation of climatic variables in different latitudinal ranges on either side of the equator

LMM, lowest monthly mean; HMM, highest monthly mean; N, Northern Hemisphere; S, Southern Hemisphere.
Temperature
Minimum temperature
The mean annual minimum temperature over all the collection sites was 19.0°C (Table 4). The mean annual minimum temperature (20.1°C) in the NH was significantly higher than that of the SH (14.6°C). It was less (19.0°C in the NH and 12.9°C in the SH) in the higher latitudes than in the lower latitudes (20.5°C in the NH and 14.9°C in the SH). No clear pattern exists for different latitudinal ranges in both hemispheres for the minimum temperature. However, the minimum lowest monthly mean ( − 17.6°C) was observed in the latitude range of 30°–35°N. Furthermore, there was an abrupt reduction in the minimum of the lowest and highest monthly mean minimum temperature in the latitude range of 10°–15°N. The maximum and mean of the lowest monthly mean minimum temperature decreased with the increase in latitude in both hemispheres.
Maximum temperature
The mean annual maximum temperature over all the collection sites was 32.4°C (Table 4). It was significantly higher (33.3°C) in the NH than in the SH (28.9°C). Lower latitudes were significantly warmer (33.6°C in the NH and 29.2°C in the SH) than higher latitudes (32.4°C in the NH and 27.2°C in the SH). In different latitudinal ranges of both hemispheres, the minimum of the lowest and highest monthly mean maximum temperature did not show any pattern. There was an abrupt reduction in the minimum of the lowest and highest monthly mean maximum temperature in the latitude ranges of 10°–15°N and 15°–20°S. The maximum and mean of the highest monthly mean maximum temperature increased with an increase in latitude range in the NH, and no such pattern was observed in the SH.
Discussion
Pearl millet is well adapted to areas characterized by drought, low soil fertility and relatively high temperature. It also performs well in soils with high salinity. Because of its tolerance to various stress conditions, it can be grown in areas where other cereal crops, such as maize or wheat, do not perform well. Adaptation of crops to the challenges of climate change will involve the exploitation of the continually developing technologies, resources and expertise. The world collection of pearl millet landraces used in the present study is obtained from a wide range of latitudes (34.37°N to 33.00°S). The frequency distribution of landraces indicated the unequal prevalence of pearl millet across the latitudes. The main factors influencing the adaptation of pearl millet include natural and human selection, photoperiods, temperatures, soils and rainfall patterns. The results of the present study revealed that the higher mean annual day length (12.13 h), minimum (20.1°C) and maximum (33.3°C) temperature and rainfall (753.6 mm) in the NH compared with those in the SH (minimum temperature 12.1°C and maximum temperature 28.9°C and rainfall 672 mm) has led to a high prevalence of pearl millet in the NH, resulting in a larger number of landraces. Because of a wide variation in climatic factors, landraces from the NH are highly variable for all traits except for panicle thickness and was found to be a good source for early as well as late maturity, short and tall, high-tillering landraces. Bidinger and Rai (Reference Bidinger and Rai1989) reported early flowering in pearl millet under a 12 h photoperiod and a delay in flowering under a long photoperiod (14 h). Kipp (Reference Kipp2007) reported the direct effect of heat stress on plant growth and development. Ong (Reference Ong1983) reported 12°C as the base temperature, 30–35°C as the optimum temperature and 45°C as the lethal temperature for pearl millet.
A higher proportion of landraces (9573 in the NH and 2687 in the SH) from lower latitudes than from higher latitudes (3235 in the NH and 409 in the SH) indicate a high prevalence of pearl millet and an intensive collection of germplasm from lower latitudes. These results may be attributed to the near-optimum and significantly higher annual minimum (20.5°C in the NH and 14.9°C in the SH) and maximum (33.6°C in the NH and 29.2°C in the SH) temperature and rainfall (772 mm in the NH and 686 mm in the SH) in lower latitudes (Ong, Reference Ong1983; Erskine et al., Reference Erskine, Ellis, Summerfield, Roberts and Hussain1990). A high range of variation in landraces for all traits from lower latitudes can be attributed to a highly variable mean annual rainfall (647–1595 mm) and near-optimum minimum and maximum temperature at collection sites in addition to the variation due to natural and human selection over the years (Tables 1 and 4). The pearl millet landraces adapted to low mean annual temperatures, inconsistent rainfall patterns and longer days at higher latitudes are expected to flower late and grow tall. Due to the near-optimum minimum (20.8°C) and maximum (30.6°C) temperature and shorter days at the Patancheru location (latitude 17.53°) than at collection sites, landraces from higher latitudes flowered early and grew short (Hijmans et al., Reference Hijmans Robert, Guarino and Rojas2002). Near the equator, day length is constantly around 12 h and facilitates crop growth all the year around. Pearl millet is a facultative short-day species, flowering at all photoperiods, although much earlier with short days (Ong, Reference Ong1983). The critical photoperiod and temperature required to trigger flowering is species and cultivar specific. Almost all cultivars show some response to temperature and photoperiod according to their specific geographical adaptation, particularly the latitude (Ong, Reference Ong1983; Bidinger and Rai, Reference Bidinger and Rai1989; Joshi et al., Reference Joshi, Pandya, Mathukia and Dangaria2005). Wareing and Phillips (Reference Wareing and Phillips1981) reported that in many plant species, a day-length change as short as 15–20 min will have a significant effect on flowering. Pearson and Coaldrake (Reference Pearson and Coaldrake1983) reported the delay in flowering by 4–5 d per degree latitude. McIntyre et al. (Reference McIntyre, Flower and Riba1993) reported a 2-d reduction in the length of the growth period of pearl millet for each degree rise in temperature. The results of the present study are in conformity with those of Ong and Everard (Reference Ong and Everard1979) who reported delayed flowering due to long days and that each short day results in a reduction of 1.4 d to reach anthesis, leading to early flowering in pearl millet.
Climate will not be homogeneous over a relatively larger area of the hemisphere. Therefore, the assessment of diversity patterns in relation to the crop ecology of more homogeneous and smaller latitudinal ranges reveals the patterns of diversity in a better way (Upadhyaya et al., Reference Upadhyaya, Reddy, Gowda, Irshad Ahmed and Singh2007). The prevalence of diverse pearl millet in smaller regions such as different latitudinal ranges in the present study can be attributed to the differences in adaptation levels of cultivars/genotypes to the prevailing day length, minimum and maximum temperatures, soils, rainfall patterns, duration of rainfall and natural and human selection in each latitude range. The high prevalence of landraces in the latitude ranges of 10°–15°N and 15°–20°S can be imputed to the near-optimum minimum of the lowest and highest monthly mean minimum and maximum temperature prevailing in the region, indicating the minimum temperature at the collection sites as an important factor in determining the adaptation patterns of pearl millet (Table 4). Hellmers and Burton (Reference Hellmers and Burton1972) found a substantial effect of temperature on the flowering of pearl millet. A deviation in climate from the latitude ranges of 10°–15°N and 15°–20°S is reflected in the reduced prevalence of pearl millet and less number (~1%) of landraces. This was evidenced in latitudes close to the equator (0°–5°) and in higher latitudes (30°–35°). These results indicate a stronger association of latitude and the factors responsible for patterns of diversity (Ong, Reference Ong1983; Erskine et al., Reference Erskine, Ellis, Summerfield, Roberts and Hussain1990; Upadhyaya et al., Reference Upadhyaya, Reddy, Irshad Ahmed, Dronavalli and Gowda2012). Ashraf and Hafeez (Reference Ashraf and Hafeez2004) reported an optimum temperature of 33–34°C for pearl millet and its growth retarded when the temperature is too high or too low.
Because of favourable climate for pearl millet, the latitude range of 10°–15°N was found to be the good source for almost all panicle shapes, seed colours and shapes. Long-bristled landraces, which were considered to be bird resistant, are relatively more in the latitudes of 10°–15°S and 20–25°S. When a species or crop has a wide range of distribution, there is a considerable difference in the latitude between its northern and southern limits, resulting in different ecotypes, which differ in their response to temperature and day length (Wareing and Phillips, Reference Wareing and Phillips1981; Santamaria, Reference Santamaria2003). Small deviations in the patterns of diversity in the present study can be attributed to the varying local conditions, elevations, date of sowings, soils, etc. in different latitudes. Natural selection pressure for adaptation to different latitudes coupled with farmers' selection for cultivation of a specific type of material in adverse conditions in some areas might have also accounted for the observed patterns of diversity.
Clustering of different latitudinal ranges of both hemispheres was in agreement with natural geographic regions (hemispheres), except for the deviation of 10°–15°S into cluster 2 along with 0°–5°N and 15°–35°N. This deviation can be attributed to the near similarity in climate for collection sites, especially mean annual rainfall, which was maximum in the latitude range of 0°–5°N in the NH and 10°–15°S in the SH.
The genetic resource base for developing new climate change-ready crop cultivars has not been adequately explored or shared. Scientists have already embarked on a hunt to identify climate change-ready cultivars of crops in germplasm collection. Studies on different crops have shown that a careful investigation of climatic variables can lead to the identification of germplasm with useful and predictable attributes (Rick, Reference Rick and Srb1973; Klebesadel and Helm, Reference Klebesadel and Helm1986). Exploitation of the germplasm for developing cultivars suitable for specific locations in a wide range of latitudes is possible only when the information on temperature and photoperiod responses, patterns of geographical adaptation and agronomic performance of the parental material is available. Late-maturing, tall and high-tillering landraces from lower latitudes ( < 20°) are better sources for fodder production (Burton and Powell, Reference Burton and Powell1968). Early-maturing photoperiod-insensitive landraces producing long and thick panicles with large seeds from mid-latitudes (15°–20°) in both hemispheres are useful in developing cultivars suitable for diverse latitudes (Upadhyaya et al., Reference Upadhyaya, Reddy, Irshad Ahmed, Dronavalli and Gowda2012). Latitudes of 5°–15°N, which were an important source for landraces with light-coloured seeds, high scoring for spikelet density, fodder yield potential, seed yield potential and overall plant aspect, may be explored. Recording location-specific geo-reference data while collecting the germplasm is critical in studying the agronomic performance of pearl millet landraces.
Using the latitudinal patterns of diversity evidenced in pearl millet landraces under study, cost-effective missions may be launched to explore areas of high-diversity, under-collected and threatened areas for the collection of materials of interest. Although the evaluation data used in the present study are preliminary in nature and recorded over years, these data reflect the differences in the genetic make-up of the landraces in terms of clear patterns of diversity (Upadhyaya et al., Reference Upadhyaya, Reddy, Gowda, Irshad Ahmed and Singh2007). Furthermore, the present study helps in identifying suitable sites for the regeneration and evaluation of trait-specific germplasm and the introduction of appropriate pearl millet into different regions, and emphasizes similar studies on germplasm collections of other crops for their enhanced use in crop improvement programmes.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S1479262113000348
Acknowledgements
The authors sincerely acknowledge all former and present staff of the genebank at ICRISAT, Patancheru, India for their contribution to the collection, assembly, characterization and conservation of pearl millet genetic resources. The authors appreciate D. Bapa Rao, G. Dasaratha Rao, G. Ram Reddy, Shaik Babu and Mohd Ismail (all research technicians) for their help in recording observations, documentation and retrieval of geo-reference and climate data.