


Introduction
End-Permian Mass Extinction and Recovery
The end-Permian mass extinction saw the extinction of 78% of marine genera (Alroy et al. Reference Alroy, Aberhan, Bottjer, Foote, Fürsich, Harries, Hendy, Holland, Ivany and Kiessling2008) and more than 95% of marine species (Raup Reference Raup1979) and the ecological restructuring of marine communities (e.g., Schubert and Bottjer Reference Schubert and Bottjer1995; Dineen et al. Reference Dineen, Fraiser and Sheehan2014). This restructuring included the initiation of the eventual decline of brachiopod ecological importance in favor of bivalve dominance in the later Mesozoic (Fraiser and Bottjer Reference Fraiser and Bottjer2007; Greene et al. Reference Greene, Bottjer, Hagdorn and Zonneveld2011). Environmental instability following the initial extinction pulse led to a series of failed recovery attempts in some benthic marine environments (Song et al. Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan and Niu2011; Grasby et al. Reference Grasby, Beauchamp, Embry and Sanei2013; Pietsch and Bottjer Reference Pietsch and Bottjer2014) throughout the 5 Myr interval. This environmental instability has been linked to temperature fluctuation (Joachimski et al. Reference Joachimski, Lai, Shen, Jiang, Luo, Chen, Chen and Sun2012; Sun et al. Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012; Romano et al. Reference Romano, Goudemand, Vennemann, Ware, Schneebeli-Hermann, Hochuli, Brühwiler, Brinkmann and Bucher2013), reoccurring instances of oxygen minimum zone expansion onto shallow-shelf settings (Algeo et al. Reference Algeo, Chen, Fraiser and Twitchett2011; Song et al. Reference Song, Wignall, Chu, Tong, Sun, Song, He and Tian2014), and changing marine productivity and nutrient availability (Wei et al. Reference Wei, Shen, Schoepfer, Krystyn, Richoz and Algeo2014). Many workers have found evidence of intervals of benthic faunal recovery relatively early after the extinction event in which oxygen or temperature conditions are more favorable (Twitchett et al. Reference Twitchett, Krystyn, Baud, Wheeley and Richoz2004; Hofmann et al. Reference Hofmann, Goudemand, Wasmer, Bucher and Hautmann2011), which subsequently ended due to onset of unfavorable conditions. Recovery of pelagic taxa appears to have progressed more rapidly than in benthic environments, with groups such as ammonoids and conodonts experiencing rapid booms in diversity shortly after the extinction event (Brayard et al. Reference Brayard, Bucher, Escarguel, Fluteau, Bourquin and Galfetti2006; Orchard Reference Orchard2007), some of which also experienced busts in diversity shortly afterward (Stanley Reference Stanley2009).
Early Triassic Disaster Taxa
The disaster taxa of the Early Triassic are a group of benthic invertebrate taxa that are observed to become dominant and widespread in the aftermath of the end-Permian mass extinction. This group was first recognized by Hallam and Wignall (Reference Hallam and Wignall1997) and included the four bivalve genera Claraia, Unionites, Eumorphotis, and Promyalina (Fig. 1). Rodland and Bottjer (Reference Rodland and Bottjer2001) later observed the ecological dominance of the inarticulate brachiopod “Lingula” (herein Lingularia sensu Posenato et al. [Reference Posenato, Holmer and Prinoth2014]) in Lower Triassic western U.S. fossil assemblages (Fig. 1C), which was later added to the group of Early Triassic disaster taxa in Benton (Reference Benton2003). These five taxa are observed in Lower Triassic marine sections worldwide (Schubert and Bottjer Reference Schubert and Bottjer1995; Boyer et al. Reference Boyer, Bottjer and Droser2004; Fraiser and Bottjer Reference Fraiser and Bottjer2007; Komatsu et al. Reference Komatsu, Huyen and Jin-Hua2008), and commonly one or many of these taxa are the most abundant within an assemblage (e.g., assemblages collected by Schubert and Bottjer Reference Schubert and Bottjer1995). The term “disaster species” was first coined by Fischer and Arthur (Reference Fischer and Arthur1977) to refer to an organism that becomes more abundant and widespread in response to a biotic crisis, whereas it was only a minor component of communities before the crisis (Harries et al. Reference Harries, Kauffman and Hansen1996). Numerical dominance within a community can have large ecological implications, as the most dominant organisms tend to control many ecological processes, such as energy flow through a food web (Clapham et al. Reference Clapham, Bottjer, Powers, Bonuso, Fraiser, Marenco, Dornbos and Pruss2006). Kauffman and Harries (Reference Kauffman and Harries1996) define disaster taxa as opportunistic, r-strategist organisms uniquely adapted to the harsh conditions during and after a biotic crisis and therefore able to proliferate in environments where other taxa are excluded.

Figure 1 Images of the five genera of Early Triassic disaster genera as summarized by Benton (Reference Benton2003). (A) Unionites cf. canalensis, (B) Promyalina cf. putiatinensis, (C) Lingularia borealis, (D) Eumorphotis cf. virginensis, and (E) Claraia aurita. Scale bar, 1 cm.
Lingularia and Claraia have been hypothesized to be resilient to low-oxygen environments, a condition that was reoccurring in some Early Triassic settings (Wignall and Hallam Reference Wignall and Hallam1993), based on the occurrence of these taxa in dysoxic sediment types throughout the fossil record (Allison et al. Reference Allison, Wignall and Brett1995) and physiological studies (Hammen et al. Reference Hammen, Hanlon and Lum1962) on modern representatives of the group in the case of Lingularia. The modern lingulid brachiopod Lingula, deemed by some a “living fossil” due to the morphological similarity of fossil and living forms (see Emig [Reference Emig2003] for discussion), is physiologically resilient in low-oxygen conditions. Modern lingulids live infaunaully in organic-rich muddy nearshore environments that are susceptible to oxygen depletion (Craig Reference Craig1952; Worcester Reference Worcester1969), where resilience to low oxygen levels is advantageous. Experiments by Hammen et al. (Reference Hammen, Hanlon and Lum1962) and Shumway (Reference Shumway1982) suggested that modern Lingula consume less oxygen while sustaining normal metabolic functions when compared with mollusks and articulate brachiopods, which likely lends them an advantage during low-oxygen events. This potential physiological resilience may have allowed fossil lingulid brachiopods to thrive in low-oxygen conditions in the past as well. Lingulaform brachiopods occur in dysaerobic settings throughout the Phanerozoic, including within Cambrian matground facies (e.g., wrinkle structures [Bailey et al. Reference Bailey, Corsetti, Bottjer and Marenco2006]) and Paleozoic black shales (Craig Reference Craig1952; Ferguson Reference Ferguson1963), and seem to be restricted to either dysaerobic basinal settings or organic-rich intertidal settings during these time intervals (Rodland and Bottjer Reference Rodland and Bottjer2001). In the Triassic, Lingularia are categorized as shallow infaunal suspension feeders (Hofmann et al. Reference Hofmann, Hautmann, Brayard, Nützel, Bylund, Jenks, Vennin, Olivier and Bucher2014), similar in life mode to modern Lingula (Craig Reference Craig1952), and are found abundantly in Panthalassic and Tethyan collections (Wignall and Hallam Reference Wignall and Hallam1993; Rodland and Bottjer Reference Rodland and Bottjer2001; Peng et al. Reference Peng, Shi, Gao, He and Shen2007; Zonneveld et al. Reference Zonneveld, Beatty and Pemberton2007; Posenato et al. Reference Posenato, Holmer and Prinoth2014). Posenato et al. (Reference Posenato, Holmer and Prinoth2014) found postextinction Lingularia with smaller shells and larger lophophoral cavities relative to pre-extinction forms, likely enabling resistance to low-oxygen and high-temperature conditions due to a decreased body mass to lophophore surface area ratio. Lingulaform brachiopods have long been a component of low-oxygen fauna throughout the Phanerozoic (Allison et al. Reference Allison, Wignall and Brett1995), and their prevalence in these environments indicated an evolutionarily long-lived physiological resilience to these conditions.
The paper pecten Claraia shares a common morphology with other thin-walled bivalves that tend to also occur in dysoxic sediment in the fossil record (Wignall Reference Wignall1993). These include the bivalves Halobia and Paracyclas, which are also found in poorly oxygenated sediment throughout the Phanerozoic (Allison et al. Reference Allison, Wignall and Brett1995; McRoberts Reference McRoberts2000; Van Iten et al. Reference Van Iten, Tollerton, Ver Straeten, De Moraes Leme, Simoes and Rodrigues2013). Additionally, Claraia shared a remarkably similar morphology with the Toarcian bivalves Bositra and Pseudomytiloides, which act as opportunistic disaster taxa following the Early Jurassic ocean anoxic event (Caswell et al. Reference Caswell, Coe and Cohen2009; Caswell and Coe Reference Caswell and Coe2013). Claraia, a byssally attached epifaunal suspension feeder (Allison et al. Reference Allison, Wignall and Brett1995), is found throughout Panthalassic, Tethyan, and Boreal Lower Triassic collections in both carbonates and shales (e.g., Yin Reference Yin1981; Boyer et al. Reference Boyer, Bottjer and Droser2004; Posenato et al. Reference Posenato2008). Claraia appears to become locally extinct in South China at the Smithian–Spathian boundary (see Sun et al. Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012), but other workers (Tozer Reference Tozer1961; Fraiser and Bottjer Reference Fraiser and Bottjer2007; McGowan et al. Reference McGowan, Smith and Taylor2009) have reported a number of Claraia from Panthalassic Spathian collections.
No specific physiological survival strategy has been proposed for the remaining disaster genera Unionites, Eumorphotis, and Promyalina, but it is generally believed that a eurytopic life strategy allowed them to proliferate in environments heavily affected by extinction and thus to outcompete other survivors (Schubert and Bottjer Reference Schubert and Bottjer1995; Kashiyama and Oji Reference Kashiyama and Oji2004). It is possible these taxa possessed advantageous adaptations, such as a resistance to low oxygen levels, high temperature, or elevated acidity, but current understanding of the physiology of these extinct clades is limited. A recent revision of the life modes of these genera by Hautmann et al. (Reference Hautmann, Smith, McGowan and Bucher2013) and Hofmann et al. (Reference Hofmann, Hautmann, Brayard, Nützel, Bylund, Jenks, Vennin, Olivier and Bucher2014) categorizes Eumorphotis as a byssally attached epifaunal suspension feeder, similar to the bivalve Leptochondria, which is also found frequently in Lower Triassic carbonate deposits (Schubert and Bottjer Reference Schubert and Bottjer1995). Promyalina has been described as an epifaunal or semi-infaunal byssally attached suspension feeder (McRoberts and Newell Reference McRoberts and Newell2005; Hofmann et al. Reference Hofmann, Hautmann and Bucher2013a). These relatively “simple,” low-energy life modes may have lent an advantage to these taxa in deleterious conditions (Aberhan and Baumiller Reference Aberhan and Baumiller2003). Unionites is categorized as a shallow infaunal suspension feeder (Hautmann et al. Reference Hautmann, Smith, McGowan and Bucher2013), similar to Lingularia and modern-day Lingula (Craig Reference Craig1952).
Quantifying Recovery
Several workers have focused on different metrics to gauge the extent of recovery to pre-extinction levels of community complexity in Early Triassic assemblages. Some metrics estimate recovery extent based on the reappearance of complex communities consisting of diverse, multitiered trophic webs and the presence of ecosystem-engineering organisms (Payne et al. Reference Payne, Lehrmann, Wei and Knoll2006; Song et al. Reference Song, Wignall, Chen, Tong, Bond, Lai, Zhao, Jiang, Yan and Niu2011; Chen and Benton Reference Chen and Benton2012; Pietsch and Bottjer Reference Pietsch and Bottjer2014). Other metrics base recovery on the reappearance of diverse ichnocoenoses that include deep-tiering and low-oxygen-sensitive ichnotaxa such as Thalassinoides (Pruss and Bottjer Reference Pruss and Bottjer2004; Hofmann et al. Reference Hofmann, Goudemand, Wasmer, Bucher and Hautmann2011). Hofmann et al. (Reference Hofmann, Hautmann, Wasmer and Bucher2013b) determined intervals of recovery in Panthalassic assemblages based on changes in alpha and beta diversity. Functional diversity, or the disparity of life modes represented in assemblages, has also been used as an indicator of ecological recovery following the end-Permian mass extinction (Dineen et al. Reference Dineen, Fraiser and Sheehan2014; Foster and Twitchett Reference Foster and Twitchett2014). All of these metrics are similar in that the aim is to quantify the ecological complexity, and thereby the recovery progression, of ancient communities. Communities that are highly uneven, low in diversity, and highly simplistic in life mode are more susceptible to disturbances, temporally unstable, and are indicative of an ecosystem under physical stress (Hillebrand et al. Reference Hillebrand, Bennett and Cadotte2008). Increasing community complexity, with a decrease of highly dominant taxa and increasing evenness, can be used to quantify progression of recovery; thus, the measure of disaster-taxon dominance is expected to track metrics of community complexity. An abundance of disaster taxa in assemblages may indicate: (1) that overall evenness of the community is low, with lower richness and alpha diversity and potentially less functional diversity and trophic-web complexity represented; and (2) that certain abiotic stressors, such as low oxygen levels, may be present and promoting the expansion of disaster taxa by excluding other taxa. This study aims to explore the correlation of disaster-taxon occurrences and abundance with metrics for ecosystem complexity and to assess the pace of recovery of complexity in benthic communities in the Early Triassic.
Methods
For this study, a quantitative analysis of dominance in Early Triassic marine benthic communities in Panthalassa and Tethys was undertaken using abundance data from fossil collections downloaded from the Paleobiology Database (2014). Entries with abundance data reported for brachiopod, bivalve, and gastropod genera were used to represent invertebrate assemblages. Further filtering of collections was applied; entries were deemed suitable for inclusion in this study if abundance counts of all bivalves, brachiopods, and gastropods were reported (e.g., brachiopod-only studies were excluded) and more than 20 individuals were counted. The cutoff criteria of 20 individuals or more per collection was chosen to reduce noise introduced by smaller collections and was found to increase statistical significance in the analysis. Other biomineralizing benthic marine invertebrates groups, such as echinoderms, were not included, due to the difficulty of assessing accurate counts of living individuals from disarticulated bioclast counts and their lack of representation in Paleobiology Database abundance reports. Supplementary Table S1 summarizes all collection references used. Western U.S. and western Canada collections were used to represent the Panthalassic region, and collections from Italy, Pakistan, and South China were used to represent the Tethys (Fig. 2). The data set was analyzed for three time bins representing the Early Triassic stage or substage intervals: the Induan stage (including the Griesbachian and Dienerian substages), the Smithian substage, and the Spathian substage. Griesbachian and Dienerian substage collections were combined into the single Induan stage time bin due to the limited number of collections available in some cases. The Tethyan analysis included abundance counts from collections made by the authors and collaborators (Pietsch and Bottjer Reference Pietsch2015). These counts were extracted using bulk sampling methods whereby an equal volume of fossiliferous rock was collected and analyzed with the aid of a dissecting microscope, and invertebrate taxa were identified to the genus level. A total of 179 abundance collections reporting 19,217 individuals were used in this analysis, with 140 collections representing Panthalassa and 39 representing Tethys. Smithian and Spathian Tethyan collections were underrepresented compared with Panthalassa, but the large number of individuals reported from this region (n=2155) makes it possible to measure true biological signals when normalizing metrics such as relative abundance are used.

Figure 2 Simplified Early Triassic map of Pangea showing regions where abundance collections reported in the Paleobiology Database were made, including western United States, western Canada, Italy, Pakistan, and South China. Modified from Scotese (Reference Scotese2001) and Pietsch and Bottjer (Reference Pietsch and Bottjer2014).
Total abundance and average relative abundance were calculated for each collection, and occurrence frequencies, or the percent of collections in which a genus occurs, were calculated per time bin and region for all genera present. Clapham et al. (Reference Clapham, Bottjer, Powers, Bonuso, Fraiser, Marenco, Dornbos and Pruss2006) advocated the use of abundance data in addition to presence–absence data for evaluating ecological trends in the fossil record, as taxonomic dominance (high diversity) is not necessarily reflective of ecological dominance (high abundance). Ecological dominance of a taxon can be quantitatively identified by high average relative abundance and a high occurrence frequency, and both were considered in identifying periods of disaster-taxon dominance. Traditionally, abundance data has not been used in paleoecological studies due to concerns regarding the fidelity of the abundance record and the time-consuming nature of bulk-sampling methods. However, many actuopaleontological studies have shown that time-averaged death assemblages accurately reflect generalized abundance relationships between taxa in living communities (Kidwell and Flessa Reference Kidwell and Flessa1995; Kidwell Reference Kidwell2002). Here, we assume relative abundance of fossils accurately approximated the time-averaged abundance distributions of the living benthic communities.
Simpson’s dominance index (D) (calculated using PAST, Version 1.89 [Hammer et al. Reference Hammer, Harper and Ryan2009]) and evenness (E 1/D) were used as a metric of overall community evenness–dominance independent of disaster-taxon abundance. Simpson’s D is a measure of taxonomic diversity that is sensitive to highly abundant taxa, more so than other diversity indices such as Shannon’s H (Hammer and Harper Reference Hammer and Harper2008). Evenness (E 1/D) is calculated by normalizing the reciprocal of Simpson’s D to taxonomic richness and thus represents evenness independent of diversity (Magurran Reference Magurran2004). These two metrics are calculated as follows, where p i is the relative abundance of the ith taxon in a given collection, and N is the total number of taxa in that collection:
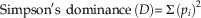 $${\rm Simpson' s \;\,dominance}\,{\rm (\it D){\equals}}\mathop{\Sigma}{(p_{i} )^{2} } $$
$${\rm Simpson' s \;\,dominance}\,{\rm (\it D){\equals}}\mathop{\Sigma}{(p_{i} )^{2} } $$
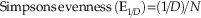 $${\rm Simpson \rsquo s}\,{\rm evenness}\,{\rm (E}_{{1\!/\!D}} {\rm ){\equals}(1\!/\!}D{\rm )\!/}N$$
$${\rm Simpson \rsquo s}\,{\rm evenness}\,{\rm (E}_{{1\!/\!D}} {\rm ){\equals}(1\!/\!}D{\rm )\!/}N$$
Results
Dominance and evenness in Early Triassic benthic assemblages exhibit some significant differences between time bins and regions. Median Simpson’s dominance index is high in Induan Panthalassic assemblages (Fig. 3A) but decreases significantly in the Smithian (Mann-Whitney U-test: p=0.034, a=0.05). Median dominance remains low in the Spathian of Panthalassa when compared with the Induan, but a larger portion of the collections exhibit a dominance of 0.5 or greater. However, no significant difference is observed between Smithian and Spathian dominance values. Within Tethyan time bins, median dominance decreases between Induan and Smithian assemblages, though not significantly (Fig. 3B). However, there is a significant increase in dominance between Tethyan Smithian and Spathian assemblages, though sample size for these time bins is small (n=4 and n=8 collections, respectively). When dominance is normalized to taxon richness (E 1/D), there is no significant difference observed between any time bins in either region (Figs. 3B and 4B), implying that changes in richness significantly affect dominance in these time bins. Time bin comparisons between Panthalassa and Tethys show significantly lower dominance in Tethyan Induan and Smithian assemblages when compared with contemporaneous Panthalassic assemblages (Fig. 5A,B; Mann-Whitney U-test: p=0.007 and p=0.013, a=0.05). Overall, Panthalassic assemblages exhibit declining levels of dominance, whereas Tethyan assemblages sustain similar levels of dominance throughout the three time bins. Tethyan assemblages are generally more even (less dominance) then contemporaneous Panthalassic assemblages, except in the Spathian, though this difference is not statistically significant and is likely skewed by the small number of collections in the Tethys (n=8) versus Panthalassa (n=59) at this time.

Figure 3 Simpson’s dominance index (D) calculated using PAST software (Hammer et al. Reference Hammer, Harper and Ryan2009) for the Induan, Smithian, and Spathian of (A) Panthalassic and (B) Tethyan collections. Mann-Whitney U-test p-values are reported between neighboring time bins. Bold values indicate significant change between time bins (a=0.05). Width of box plot represents relative number of collections used in the analysis.

Figure 4 Evenness (E 1/D) calculated using PAST software (Hammer et al. Reference Hammer, Harper and Ryan2009) for the Induan, Smithian, and Spathian of (A) Panthalassic and (B) Tethyan collections. Mann-Whitney U-test p-values are reported between neighboring time bins. Bold values indicate significant change between time bins (a=0.05). Width of box plot represents relative number of collections used in the analysis.

Figure 5 Regional comparison of dominance (D) and evenness (E 1/D) between Panthalassic and Tethyan assemblages. Dominance is shown in (A) Induan, (B) Smithian, and (C) Spathian assemblages, and evenness is shown in (D) Induan, (E) Smithian, and (F) Spathian assemblages, with width of box plot representing relative number of collections used in the analysis. Mann-Whitney U-test p-values are reported between regions. Bold values indicate significant change between regions (a=0.05).
Abundance tallies in Early Triassic assemblages show high relative abundance of the previously hypothesized disaster-taxon genera relative to other biomineralizing marine benthic invertebrates in the Induan of Panthalassic and Tethyan assemblages, with these disaster taxa representing 79.1% and 54.6% percent of individuals reported, respectively (Fig. 6). Following this high abundance in the Induan, the Smithian and Spathian assemblages exhibit a dramatic decline in disaster-taxon relative abundance in both regions. This decline is most severe in Panthalassa between the Induan and Smithian time bins, where the relative abundance is reduced to 9.0%. Tethyan assemblages also exhibit a similar decline, with Smithian disaster taxa only representing 26.2% of individuals. Spathian assemblages in both regions show low disaster-taxon relative abundance, with 20.5% in Panthalassa and 25.4% in Tethys. Interestingly, Smithian Panthalassic assemblages exhibit overall lower disaster-taxon abundance than is found in the Spathian. This decline in disaster-taxon dominance after the Induan stage of the Early Triassic coincides with a decrease of overall dominance (increased evenness) in Panthalassa. This pattern is not as apparent in Tethyan assemblages, as dominance does not significantly change between time bins, even though a decrease in disaster-taxon relative abundance is observed.

Figure 6 Overall relative abundance of all individuals reported from (A) Panthalassic and (B) Tethyan collections. The relative abundance of disaster taxa is shown in black, with all other taxa in white.
Average relative abundances and occurrence frequencies of the disaster taxa reflect this pattern of decreasing importance throughout the Early Triassic both within assemblages and regionally. In Panthalassic Induan collections, the disaster taxa Unionites, Lingularia, and Eumorphotis show the highest occurrence frequency and relative abundance of taxa present (Fig. 7A,B), with the exception of the bivalves Pteria and Leptochondria, which are also highly abundant in some Panthalassic Induan collections. In the Smithian, Unionites is the most abundant disaster taxon but is outranked in abundance and occurrence frequency by Leptochondria, Permophorus, and some gastropod genera (Fig. 7C,D). In the Spathian, we see a return of high abundance and occurrence frequency of a few disaster-taxon genera, with Eumorphotis and Promyalina ranking highly (Fig. 7 E,F). However, other taxa, including the articulate brachiopods Protogusarella and Piarorhynchella, are also highly abundant (Fig. 7F). The disaster taxa occur in 20–70% of Panthalassic collections in the Induan (Fig. 7B) but are more limited in extent in the Smithian (Fig. 7D). The Spathian of Panthalassa sees the return of widely distributed Eumorphotis and Promyalina (Fig. 7C), reflecting a reoccurrence of disaster-taxon importance following the Smithian.

Figure 7 Occurrence frequency and average relative abundance of each genus for Panthalassic assemblages. Panthalassic occurrence frequency of genera is shown in the (A) Induan, (B) Smithian, and (C) Spathian, and Panthalassic average relative abundance is shown in the (D) Induan, (E) Smithian, and (F) Spathian. Disaster taxa are shown with open marks and all other genera are shown with solid marks.
In Tethyan collections, the bivalve Unionites is the most frequently occurring and abundant disaster taxon in the Induan time bin (Fig. 8A,B), whereas in the Smithian and Spathian, Eumorphotis also gains some importance. However, in the Smithian and Spathian, both are outranked in terms of abundance by the bivalve Neoschizodus. In both Panthalassic and Tethyan collections, Claraia and Lingularia only occur in high abundance during the Induan time bin (Figs. 7B and 8B), after which they become only minor components of assemblages. Compared with Panthalassic Spathian assemblages, where Eumorphotis and Promyalina are more prevalent than other disaster taxa in Tethyan assemblages, Unionites appears more important at this time. The patterns of changing relative abundance of the disaster taxa are reflected in their regional distributions in Panthalassa and Tethys; as these taxa become less frequently occurring regionally, they also become less abundant in those assemblages in which they do occur.

Figure 8 Occurrence frequency and average relative abundance of each genus for Tethyan assemblages. Tethyan occurrence frequency of genera is shown in the (A) Induan, (B) Smithian, and (C) Spathian, and Tethyan average relative abundance is shown in the (D) Induan, (E) Smithian, and (F) Spathian. Disaster taxa are shown with open marks and all other genera are shown with solid marks.
Discussion
There is a significant decrease in Simpson’s dominance in Panthalassic assemblages between the Induan and Smithian time bins coincident with a dramatic decrease in the relative abundance of the disaster taxa as a group, as well as an overall decline in dominance over time in Panthalassa. A similar, though not as dramatic, decline in disaster-taxon abundance is observed in Tethyan assemblages. The abundance of the so-called disaster taxa appears to track dominance, a metric of community complexity, most clearly in Panthalassic assemblages.
While individual disaster taxa reoccur in high abundance at other times in the Early Triassic, it is only in the Induan that we see the disaster taxa rank highest in terms of relative abundance and occurrence frequency as a group; most clearly in Panthalassic collections. Tethyan Induan assemblages are more even than contemporaneous Panthalassic collections, and only the disaster taxon Unionites is highly abundant and widespread. Following the Induan stage, disaster taxa overall become less important in terms of abundance and occurrence frequency in both regions, as other taxa become more ecologically important and widespread. These more abundant non–disaster taxa are associated with an overall decrease in community dominance and an increase in diversity. Claraia and Lingularia occur in later stages of the Early Triassic, but the peak of their ecological importance is limited to the Induan stage. Of all the disaster taxa, Claraia and Lingularia are the only two with clear preferences for occurring in low-oxygen conditions (Allison et al. Reference Allison, Wignall and Brett1995). Their prevalence in the Induan of Panthalassa may indicate an increased role of low-oxygen conditions in biotic suppression in these environments, which subsequently subsides in prevalence. Their dominance may also be a reflection of the direct effects of the extinction event allowing these two physiologically adept genera to become dominant and widespread, while the later dominance of Eumorphotis, Promyalina, and Unionites may be a response to onset of different environmental conditions or normal competitive interaction. Whatever the reason for the decline in Claraia and Lingularia abundance after the Induan, it is clear that these two genera exhibit dissimilar patterns of dominance from the other three taxa but act as true disaster taxa as defined by Kauffman and Harries (Reference Kauffman and Harries1996) following the Permian–Triassic extinction. Further work is needed to clarify the status of the remaining three genera as true disaster taxa.
The decline of disaster-taxon abundance after the Induan stage tracks increasing community complexity in benthic assemblages, as overall dominance declines and diversity increases. As recovery progressed following the Permian–Triassic extinction, rebounds and resets in recovery of community complexity have been observed by many workers looking at discrete sections or regions (Pietsch and Bottjer Reference Pietsch and Bottjer2014). As some of these reported failed recovery attempts occur within single substages, the temporal resolution of the data set used for this study does not allow for direct identification of these intervals. However, the averaging of these short-term recovery signals over the entirety of the Early Triassic reveals a pattern of overarching recovery, starting in the Smithian and continuing into the Spathian. As this study samples Panthalassic and Tethyan assemblages collected over a wide range of benthic environments with varying energy levels, depositional settings, and oxygen conditions, the observed decrease in dominance after the Induan stage reflects an averaged signal of recovery of ecosystem complexity across these environments.
As complexity increased over the Early Triassic, larger-scale ecological restructuring was already underway (Fraiser and Bottjer Reference Fraiser and Bottjer2007). The end-Permian extinction marked the beginning of the end for brachiopod ecological and taxonomic importance in marine communities, but the transition from brachiopod-dominated Paleozoic Fauna assemblages to bivalve-dominated Modern Fauna assemblages was not instantaneous. A rebound of Paleozoic faunal components was observed in Panthalassic Middle Triassic assemblages by Greene et al. (Reference Greene, Bottjer, Hagdorn and Zonneveld2011) and is attributed to the decoupling of taxonomic and ecological restructuring starting at the Paleozoic–Mesozoic transition. In this study, an interval of increased articulate brachiopod ecological importance is observed in the Spathian of Panthalassa, with reports of high abundance of Protogusarella and Piarorhynchella. Rebounds in ecological importance of Paleozoic faunal components such as brachiopods and crinoids (Boyer et al. Reference Boyer, Bottjer and Droser2004) in the Spathian of the Early Triassic lends further evidence to the asynchronous nature of the Paleozoic to Modern faunal transition.
Conclusion
Dominance of disaster taxa in marine benthic assemblages reflects the status of ecosystem complexity and therefore of progression of recovery following the end-Permian mass extinction. We find that the Induan stage in both Panthalassic and Tethyan assemblages is when disaster taxa occur at their highest abundance and widest distribution. This stage is also when overall community dominance is at its peak in Panthalassic assemblages. Following the Induan stage, we see a reduction of overall community dominance as well as disaster-taxon abundance and distribution. The disaster taxa Claraia and Lingularia exhibit the most extreme decrease in importance, and highly abundant occurrences of these two taxa are limited to the Induan stage. Eumorphotis, Promyalina, and Unionites are found to occur in high abundance in some Smithian and Spathian collections, but other non–disaster taxa join their ranks as well, including articulate brachiopods in Panthalassa. The general decrease of disaster-taxon abundance observed tracks an averaged signal of gradual recovery of benthic marine community complexity through the Early Triassic.
Acknowledgments
We thank R. Twitchett, F. Corsetti, and W. Berelson for helpful discussions, as well as M. Clapham and an additional anonymous reviewer for their insightful comments. We are indebted to the contributors, editors, and authorizers of the Paleobiology Database and all those who make this resource freely accessible for research. We would like to thank M. Clapham and J. Thompson for assistance with all R code used. Additionally, we would like to thank C. Pietsch for collaboration both in the field and in the lab. Finally, we would like to acknowledge the funding sources that made this research possible: the Graduate Student Research Award from the Geological Society of America, the Paleontological Society Student Research Award, the SEPM Student Research Grant, and funding sources made available through the University of Southern California Earth Sciences Department. This article is Paleobiology Database official publication number 241.
Supplementary Material
Supplemental materials deposited at Dryad: doi:10.1017/pab.2015.47








