


Introduction
Macquarie Island is a geographically isolated, sub-Antarctic island that has never been connected or close to any other landmass. The island is relatively recent in origin, being above sea level for fewer than 600,000 years (Greenslade Reference Greenslade2006). It is 34 km long, up to 5 km wide, occupies an area of 120 km2 and is situated at 54° 45′S and 159° 0′E, some 1000 km south of Tasmania and New Zealand (Fig. 1). The climate, topography, flora and fauna are described in detail by Selkirk and others (Reference Selkirk, Seppelt and Selkirk1990) and by Greenslade (Reference Greenslade2006). Macquarie Island is a World Heritage Area and a Tasmanian National Park. Management is currently directed toward removal of feral vertebrates and minimisation of human impacts (Anon 2003).

Fig. 1 Map of Macquarie Island showing its location in the Southern Hemisphere (lower inset). The rectangle centred on the northern region of the island (left) indicates the sampling area where the distributions of the two exotic terrestrial crustaceans were mapped at intervals over twelve years between 1992 to 2004. The areas searched intensively for both Puhuruhuru patersoni and Styloniscus otakensis are shown by an ‘A’ (where the ANARE station is located on the isthmus; Fig. 3) and ‘B’ (Fig. 4).
Two exotic invertebrates were discovered on Macquarie Island in the early 1990s. The first, a terrestrial isopod, Styloniscus otakensis (Chilton, 1901) (Styloniscidae), was recorded on Macquarie Island for the first time by van Klinken and Green (Reference van Klinken and Green1992) and the second, a terrestrial amphipod, Puhuruhuru patersoni (Stephensen, 1938) (Talitridae), was first recorded by Richardson and Jackson (Reference Richardson and Jackson1995). See Greenslade (Reference Greenslade2006) for further information and images. Each species was recorded from a single but different locality near the Australian National Antarctic Research Expeditions’ (ANARE) Station (van Klinken and Green Reference van Klinken and Green1992; Richardson and Jackson Reference Richardson and Jackson1995). Since their initial discovery, detailed surveys of the populations of these two species have been conducted in four of the twelve years between 1994 and 2004.
The distributions of the two species, as found on the different surveys, together with any changes over time, are recorded in this paper. In addition, evidence for the possible routes of their introduction to Macquarie Island is discussed. The potential impacts of these species on the island's ecosystem are considered and the management implications of their current distributions, with regard to present activities on the island are discussed. The relative merits of conducting further research on the species or attempting to remove the species from the island, are considered.
Study area
The island is located half way between Tasmania and the Antarctic continent and consists largely of a raised plateau surrounded by a narrow coastal platform (Fig. 1). At the northern end, the plateau ends in a narrow isthmus with a further, smaller and lower raised plateau at the extreme northern end. The ANARE Station is located on the isthmus which covers an area of about 500 m by 200 m (11 ha), at the northern end of Macquarie Island (Fig. 1). The island lacks trees or shrubs and the vegetation consists of tussock grassland and low herbland with some bare areas with patchy mosses. The climate is equable, being consistently cool and moist and totally icefree. The physical attributes of the island are described in more detail in Greenslade (Reference Greenslade2006).
Methods
As the earliest records of both species were within the bounds of, or close to, the ANARE station, this area was surveyed most intensely and field work extended southwards from there until neither species was encountered. As both species are up to 10 mm long (males 5.5 to 9 mm, females 7.5 to 11 mm, average 8.5 mm for both sexes), visual methods were sufficient for detection. More detailed survey methods are described below.
Puhuruhuru patersoni
Before the systematic surveys began in summer 1993/94, opportunistic searches for invertebrates were conducted in 1992 and 1993 (Richardson and Jackson Reference Richardson and Jackson1995). The amphipod was first only recorded in 1993 from a small area near the fuel dump on the isthmus. Later, detailed surveys were carried out from 9 to 11 March 1994, in November 1996, in February 2003 and between 24 January and 29 February 2004. More superficial checks on population distributions were made (March 1996 and 29 November 1996, 11 November 1999), but not all areas were surveyed at these times. All work was carried out during the summer months.
The first detailed mapping (summer 1993/4) divided the isthmus into 25 m by 25 m grid cells and searches were made by hand using the method described in Richardson and Jackson (Reference Richardson and Jackson1995). Each grid cell was searched for six person minutes within an area of roughly 4 m2 at the centre of a cell. A compass and tape measure were used to establish the sampling points. In several areas, the resolution of the survey was increased by dividing each grid cell into four and searching each smaller grid cell (12.5 m by 12.5 m) in the same way. A total of 263 grid cells were searched (Fig. 2). Densities of the amphipod were not determined because this animal appears to prefer the bases of tussocks and, since destructive sampling of whole tussocks would be required to quantify abundance, there was a risk of spreading the species to new locations. The apparent centre of the population was determined from this survey.

Fig. 2 Map of the isthmus (showing ANARE station buildings) mapped for Puhuruhuru patersoni in summer 1993/1994 using 25 × 25 m grid cells (smaller cells were 12.5 × 12.5 m). Shaded squares indicate cells where amphipods were present, empty cells indicate those that were searched but amphipods were not found.
In November 1996, only the most westerly and largest population was mapped, but in considerable detail. Presence of amphipods and isopods was recorded at 5 m intervals along eight compass points radiating from the point thought to be the centre of the population. Three additional transects were also surveyed to the south and east of the main survey area (Fig. 3). The approximate number of amphipods observed was recorded as either few (1–2), moderate (3–5) or numerous (more than 5).

Fig. 3 Map of the station area on the isthmus showing the current distribution of Puhuruhuru patersoni and the northern distribution of Styloniscus otakensis (see Fig. 4) from the 2004 survey. Transects to the west of the biology laboratory that indicated changes from the November 1996 survey for either the amphipod or isopod distributions are indicated using lowercase letters (a-n).
In January 2003, a limited survey was conducted using two of the transects used in 1996, and presence and absence of P. patersoni was recorded. At this time vegetation cover and species composition was recorded in detail. The most recent and most extensive survey was carried out between 24 January and 29 February 2004. In this survey, ArcMap™ 9.1(ESRI®) was utilised and all known populations were examined using the transect intervals and methodology from the 1996 survey, in addition to a re-examination of the most easterly and southernmost population adjacent to the fuel dump (Fig. 3).
Styloniscus otakensis
The original record of this species was made in summer 1990/1991 from a single site south of the living quarters, Southern Aurora dormitory (Fig. 3) at the station. A general survey for the isopod was carried out during the summer of 1991/1992, 1992/1993 and 2004 south of this site. The method used was to search the organic matter debris under Poa foliosa by brushing aside about 10 cm by 10 cm of the loose soil and leaf litter with a trowel and then watching the harder soil surface for movement. If the isopod were present it would become mobile after a few minutes. Stems and seed heads of Stilbocarpa plants were also examined as the isopod had been observed on them. Because of the small-scale patchy distribution of this species, each negative record was confirmed by searching several plants at each site. The search was continued every 20 m to 50 m until no more individuals were found.
In order to make an estimate of the density/activity and sex ratio of the isopod, in the summer of 1992/1993, pitfall traps were set in two areas, outside the Southern Aurora dormitory and on Razorback Ridge (Figs 3, 4), where isopod populations were most dense. The traps, 1.8 cm in diameter, were part filled with 70% alcohol. Also, densities of the isopod were quantitatively determined at two sites, Razorback Ridge and Halfway Hill (summer 1992/1993, 5 January and 8 February) and at four sites in February 1994 (Fig. 4). Four soil cores (diameter 50 mm by depth 40 mm) were taken close together from each of a pair of plots 25 cm by 25 cm in dimensions, several metres apart, one being an area of isopod presence and the other of absence. Several pairs of plots were sampled on each date. Soil moisture and soil density were measured gravimetrically and cores were hand sorted for isopods on white trays (extraction by placing soil cores in Tullgren funnels also was tried but was unsuccessful). In addition, 25 randomly located cores were taken on 12 March 1994. For all samples, mean densities were calculated by first restricting the data to non-zero values and, as suggested by an examination of the residuals, the isopod counts were log transformed. Since the sampling design was unbalanced and included both fixed and random effects, a mixed-effects model was fitted by restricted maximum likelihood (Payne Reference Payne2002) treating date, site, soil moisture, and soil density as fixed effects and plot as a random effect. Significance of an effect was determined using a likelihood ratio test (Payne Reference Payne2002). The cores taken in 1994 were considered as being taken from 25 plots.
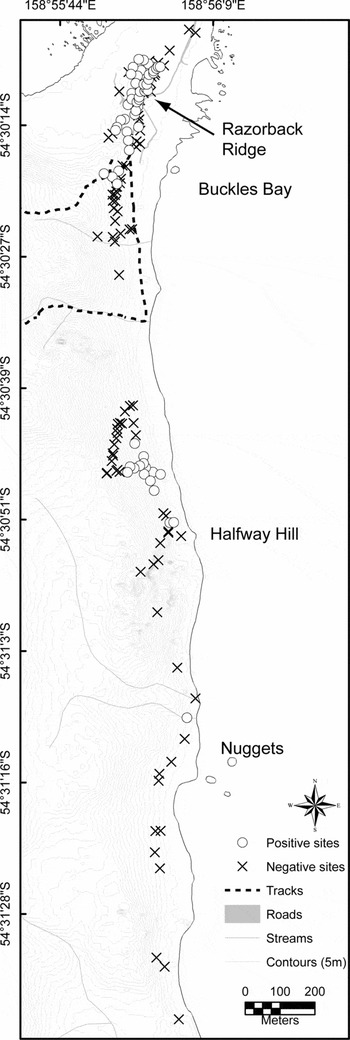
Fig. 4 Map of the distribution of Styloniscus otakensis from Razorback Ridge (where most dense populations were found) to the most southern known record at the Nuggets (previously at Halfway Hill) found in 2004 (map continues from Fig. 3).
The 24 January to 29 February 2004 survey examined all known populations for presence or absence using the same method as during the summers of 1991–1993 and these data were mapped into ArcMap™ 9.1(ESRI®).
The distribution of potential habitat (Poa foliosa tussock) was also investigated. The modern distribution of tussock on the isthmus was determined from photographs taken in 2004, and compared visually with the distribution of tussock in the early 1900s from photographs of the isthmus taken in about 1912 by members of the Australasian Antarctic Expedition led by Douglas Mawson (Fig. 5a). To determine possible sources of introduction, a search was conducted among historical accounts, principally those of Cumpston (Reference Cumpston1968), A. Hamilton (Reference Hamilton1894) and the unpublished diary of H. Hamilton. Early photographs were used to determine the location of tracks and huts.

Fig. 5 Photographs indicating vegetation cover (primarily Poa foliosa) on the isthmus, Macquarie Island: (A) 1911–1914, from Mawson (Reference Mawson1942), Plate 1. ‘A panoramic winter prospect from the summit of Wireless Hill’ (Sandell); (B) 1963 (P. Whyte); (C) 1988 (M. Whittle); (D) 2004 (M. Stevens). The area of dense Poa foliosa on isthmus in which the densest populations of the amphipod is found is conspicuous on all dates (see circle indicating the stack that now represents the approximate centre of the amphipod population) and lacks snow in winter (B and C). Sealers’ hut to the east of Razorback Ridge (see arrow, A) where densest populations of isopods are present. A comparison of the 1911–1914 and 2004 photographs shows marked erosion across the isthmus (in foreground).
Results
Puhuruhuru patersoni
As a result of the detailed survey in 1994, a further population of Puhuruhuru patersoni was discovered about 50 m northwest of the population documented by Richardson and Jackson (Reference Richardson and Jackson1995) (Fig. 2). This second population was located in Poa foliosa tussock both east and west of the biology laboratory (Fig. 3). This area was not searched in 1993 (J. Jackson, personal communication). This population was more extensive than at the fuel dump and the animals appeared to be at higher densities. The approximate total area covered by this population in 1994 was 1.45 ha.
The second survey in 1996 showed that the centre of the population had moved about 15 m in a south- southwesterly direction (Fig. 3), the centre of the population in summer 1994 being now well north of the centre (see point ‘m’, Fig. 3). The total area of this population was now about a third of a hectare. In the survey of 20 January to 29 February 2004 when all known populations were re-examined as well as much of the surrounding area, the populations appear to have similar boundaries as were found previously in the 1996 survey, but densities appeared much greater within the previous limits. There were also a few instances (indicated by lowercase letters in Fig. 3) in which presence or absence of the amphipod in the 2004 survey was different to the situation revealed by the 1996 survey. There is some indication that the main populations are now centred around a stack that can be seen in Fig. 5 (the stack lies next to the ‘n’ in Fig. 3).
The vegetation survey of the most westerly population of P. pattersoni in 2003 indicated that the species was only associated with Poa tussocks and was absent from other plants or from bare ground. The modern distribution of tussock on the isthmus is shown in the photograph in fig. 5d and the distribution around 1912 is shown in fig. 5a. There seems to have been little, if any, change in the distribution of Poa tussock over 100 years although what was originally a continuous stand on the isthmus has been fragmented by the station buildings. Also, a visual comparison of fig. 5a with Fig. 5B–D shows that there has been considerable coastal erosion over the last 100 years especially on the western side. The specific human activities that resulted in the introduction of P. patersoni to the exact location at which it is now found have not been identified.
Styloniscus otakensis
Additional populations of S. otakensis were discovered in a survey of 1991/1992 by P. Serov. These were additional to that at the Southern Aurora dormitory discovered during the previous year. The distributions of the new populations were mapped in 1991/1992 and largely confirmed both in 1992/1993 and in 2004 and are shown in Figs 3 and 4. A large population was found at Razorback Ridge and other populations were found further south with a limit at the Nuggets which is about 2 km south from the previous record. The species was also recorded in 1996 adjacent to the most westerly population of P. patersoni, west of the science buildings but the species was not found at any other locality on the island despite searching some 500 additional sites away from these populations in 2004 (see Greenslade and others in press). Where the isopod occurs, it had a very patchy distribution. It was not found where humidity was low, leaf litter depth was shallow or plant cover was absent. It was observed on the tops of Poa tussocks, in root mats, occasionally on Stilbocarpa stems and on seed heads, in moss adjacent to streams, in the base of Festuca clumps and in short Epilobium-Acaena-Agrostis vegetation adjacent to the beach. It was found from just above the supralittoral zone to about 45 m altitude. The preferred soil types ranged from peat to almost pure sand.
Isopod densities were not significantly different between locations or between dates apart from on the eastern slope of Razorback Ridge where populations were more continuous. This site, where a sealers’ hut (seen in fig. 5a) was located at the base of the ridge, is considered to be the origin of the population. Where isopods were present, there was no effect of soil moisture or soil density on abundance, although there may be a threshold value for soil moisture of about 30%, below which isopods are not found (Fig. 6). Where isopods were present, mean isopod density per core was 10.3 individuals and the 95% confidence interval for the geometric mean was (8.5, 12.4). This equates approximately to 4000 to 6000 isopods per square metre. Individuals were not well collected in pitfalls. Standard hand searches showed that the sex ratio was about equal (23M: 22F) in summer 1993.

Fig. 6 Relationship of isopod abundance to soil moisture. Two sites were sampled on two dates in 1993. Razorback Ridge: open symbols. Halfway Hill: filled symbols. January: circles. February: triangles. Each point is the sum of four soil cores. The curve is a cubic smoothing spline with 3 degrees of freedom (P = 0.35).
Richardson and Jackson (Reference Richardson and Jackson1995) mapped six records of S. otakensis (Richardson and Jackson Reference Richardson and Jackson1995 Fig. 2) close to station buildings in 1992/1993, three north of the Southern Aurora dormitory. These three sites were checked twice during the 1996 summer survey and no isopods were found there. It appears that these three additional records were marked on the map in error (A.M.N. Richardson personal communication) and that Styloniscus is only known from the Southern Aurora dormitory (Fig. 3) and also from near the biology laboratory.
All known populations south to the Nuggets and surrounding areas were examined between 20 January and 29 February 2004. Styloniscus may have expanded southwards from Halfway Hill since surveyed in 1992 as a single record was found at Nuggets in 2004 (Fig. 4) but this may have been missed on the earlier survey. Apart from expanding southwards from the Southern Aurora dormitory since 1991, it had not otherwise spread significantly during the previous twelve years.
Discussion
Distributions of the two species
Puhuruhuru patersoni has only been found in all years surveyed within an area of current intensive use (Fig. 3) and also coincides with previous heavy use of the site by foot parties (Fig. 5). It probably originally had a continuous distribution across the isthmus within Poa dominated areas but the population has been fragmented by station buildings. No expansion or contraction was noted between 1994 and 2004 but the centre of the population has shifted a few metres to the south. Similarly Styloniscus otakensis populations also seem to have been fairly stable from 1991/1992 until 2004 although it may have spread by pedestrian traffic within the station.
P. patersoni has only been recorded south of latitude 45°S in New Zealand, despite apparently extensive collecting (Duncan Reference Duncan1994). The type locality is Bench Island, near Stewart Island and the species has also been collected from about 200 km to the north and south of Dunedin, and from the Snares, Stewart and South Cape Islands but not from Auckland or Campbell Islands (Duncan Reference Duncan1969, Reference Duncan1994). The species varies morphologically with latitude (Duncan Reference Duncan1994). According to Richardson and Jackson (Reference Richardson and Jackson1995), the Macquarie Island specimens most closely resemble the Snares Island specimens but are not identical. Stewart Island specimens have not been examined.
The distribution of S. otakensis in New Zealand is somewhat unresolved (van Klinken and Green Reference van Klinken and Green1992) but appears to be similar to P. patersoni in that it is recorded from near Dunedin, except that it is, in addition, recorded from the Chatham and Auckland Islands (Stephensen Reference Stephensen1927). So far, the species is not recorded from Campbell Island. It is not known if either the isopod or amphipod is present at The Bluff, the port of Invercargill, or if the isopod is on Stewart Island.
Route of introduction
As, apart from Macquarie Island, both crustacean species are otherwise only known from southern New Zealand and some outlying islands, any introduction is most likely to have originated from this area. It is extremely unlikely that the introduction of either species to Macquarie Island was a natural colonisation event. The organisms are too large to have been transported by wind or by birds and transport by flotsam is unlikely because ocean currents are unsuitable (Richardson and Jackson Reference Richardson and Jackson1995). Alternatively, the evidence for an accidental introduction by humans is very strong as the distribution of both species on Macquarie Island coincides with sites of intensive human activity on the isthmus over the last 200 years. Early photographs, as noted earlier, show that the population of S. otakensis at Razorback Ridge coincides with the location of a hut (Fig. 5a) that was used as living accommodation by oiling parties in the late 19th and early 20th centuries and this is supported by archaeological evidence (Townrow Reference Townrow1989). A recent introduction of another isopod, Porcellio scaber Latreille, 1804, to Marion Island is assumed to have arrived on building materials (Slabber and Chown Reference Slabber and Chown2002) and this could be the route by which S. otakensis was introduced. Only the southern population of S. otakensis appears to be isolated from a site of human habitation, although it is situated on a well-used coastal pedestrian route (Fig. 1). The most northerly population of S. otakensis is restricted to an area beside the present day Southern Aurora dormitory (Fig. 3) where it could have been transported on muddy boots from a site further south and this could have been the route by which individuals were transported further south along the coast to the Nuggets.
It is unlikely that either species was introduced after 1920 since there have been few opportunities for introductions from southern New Zealand as exploitation from there ceased in 1919. Since then, virtually all voyages have originated directly from Australia, with only a handful of exceptions, and quarantine controls are now stringent. Only two ships visited the island from New Zealand in the 1970s, one bringing supplies, but both originated from Christchurch (G.R. Copson personal communication), which is now known to be the northern distributional limit of both species (M. Stevens, unpublished data). Only recently, from the mid 1980s, have tourist ships visited the island, some of which come from New Zealand, but tight controls are imposed on them to prevent the introduction of alien organisms. There are now sometimes up to a dozen tourist ships a year coming from New Zealand. Several airdrops, some using ballast, also were made from the mid-1970s to the early 1980s but again they originated from Christchurch and usually dropped cargo on the Plateau (G.R. Copson personal communication) or on the isthmus (Macquarie Island ANARE station video archives). Given that the location of the second amphipod population is adjacent to the biology and science buildings, one possibility is that the amphipod was accidentally introduced more recently by a scientist (Greenslade and Farrow Reference Greenslade and Farrow1994). However, there have been no arrivals of personnel or equipment via the Dunedin region or further south since records began in the 1960s. In addition, no live plants have been introduced from New Zealand (K. Kerry personal communication, G. Copson personal communication) although some were brought by H. Hamilton in 1912 from the Hobart Botanic Gardens. The evidence therefore strongly suggests that both species were introduced before 1919, rather than as a result of more recent activities on the island.
Immediately upon its discovery in 1810, Macquarie Island was exploited for fur seal oil and skins, and later in the century for elephant seal and penguin oil. Oiling parties typically stayed for periods of several months, sometimes for up to two years, and brought with them a considerable quantity of cargo. Cargo that could serve as a vehicle for introductions included sacks of potatoes (often stored in straw) firewood, timber as building material, barrels containing coal and possibly terraria or live plants in pots with soil. Terrestrial crustaceans would be able to persist, alive, in damp soil for at least a week while the journey from the source area could take as little as two days. Another potential carrier would have been ships’ ballast, especially from ships that were wrecked at the isthmus. However, since there was no wharf, jetty or landing beach, ballast from wrecked vessels would most likely have been dumped in seawater. Although most of the earliest voyages (1810–1830) to Macquarie Island originated from Sydney, many went via ports in the southern New Zealand region (Cumpston Reference Cumpston1968) and even called in for shelter at harbours on southern islands of New Zealand en route to Macquarie Island during inclement weather, where, no doubt, they could have replenished their stores of firewood (Cumpston Reference Cumpston1968). During this early period the isthmus was almost permanently inhabited and more than a dozen huts were erected (Cumpston Reference Cumpston1968). So, an alternative view to that of van Klinken and Green (Reference van Klinken and Green1992) is that introductions occurred in the early rather than late 19th century. Even so, the most intense and continuous period of activity of harvesting penguin and elephant seal oil was from 1874 to 1919 and most voyages during this period originated from the southern New Zealand ports of Port Chalmers (Dunedin) or The Bluff (Invercargill) (Cumpston Reference Cumpston1968). Port Patterson on Stewart Island was a common last port of call and it was likely that firewood was replenished there (P. Johns personal communication). The landing site on the isthmus, where stores were often left for some time, was the nearest small bay to the Razorback Ridge hut (Fig. 5a) and some distance from where the amphipod is currently found.
Impact on the island's ecosystems
There are no terrestrial crustaceans or other large detritivores native to Macquarie Island, so the introduction of these species could have a significant impact on ecosystem processes and native species, as has been suggested for the two introduced, predatory flatworms (Greenslade and others in press). Both crustaceans are present in high numbers but so far no changes are observable in the structure of the vegetation or underlying peat and soil layers although the density of the isopod appears to be in the highest range of reported densities for terrestrial isopods (Petersen and Luxton Reference Petersen and Luxton1982). Terrestrial amphipods and isopods accelerate decomposition processes, as do many other soil and litter invertebrates (Seastedt Reference Seastedt1984), largely by stimulating the activity of fungi and bacteria (Richardson and Morton Reference Richardson and Morton1986; Gunnarsson and others Reference Jones, Lawton and Shachak1988; Griffiths and others Reference Griffiths, Wood and Cheshire1989; van Wensem Reference van Wensem1989; Teuben and Roelofsma Reference Teuben and Roelofsma1990) rather than by directly contributing to decomposition (compare Petersen and Luxton Reference Petersen and Luxton1982). In microcosm experiments, amphipods and isopods have been shown to increase microbial activity by 10% to 50% (Richardson and Morton Reference Richardson and Morton1986; van Wensem Reference van Wensem1989). Moreover, the action of these two species together may increase microbial activity even further (Richardson and Morton Reference Richardson and Morton1986). If one or both species became more widespread on the island, a major change in the landscape could possibly result by shifting the deposition/decomposition equilibrium thus giving shallower surface peat. This ‘ecosystem engineering’ (Jones and others Reference Jones, Lawton and Shachak1994) could, in turn, affect native plants and invertebrates, perhaps even leading to extinctions. For instance, the effect of feeding by the alien chironomid Limnophyes minimus on Marion Island ecosystems has been found to be significantly higher than the dominant native detritivore (Hänel and Chown Reference Hänel and Chown1998). The effect on these crustacean populations of the degradation of the landscape by the recent increase in rabbit populations, after cats were extirpated on the island, is unknown but is unlikely to increase their rate of spread.
A crucial issue is whether there is a danger of these species spreading on Macquarie Island and at what rate. At present, factors that limit the distribution of either species are virtually unknown. The isopod appears to be restricted by sparse plant cover, shallow leaf litter and low soil moisture, factors that are positively correlated, as it occurs only in soils with a moisture greater than 30% (Fig. 6), although more data are required to confirm this observation. Richardson and Jackson (Reference Richardson and Jackson1995) suggested that the amphipod was absent from other sites on the isthmus due to the presence of seals and the resulting compaction of the soil. However, the most westerly population (near the biology laboratory) is an area heavily used by elephant seals, so this hypothesis is unlikely. It does appear to be restricted by bare ground to the south and north, because there is unoccupied, apparently suitable, tussock habitat a few metres away to the south at least (Fig. 5d). Puhuruhuru patersoni is described as being a fairly inactive species (Duncan Reference Duncan1994) and there is some evidence that this is particularly so for females, which might explain the current lack of spread (Richardson and Jackson Reference Richardson and Jackson1995). Some other possible mechanisms restricting the distribution of both species on the isthmus are the intensity of salt spray (Richardson Reference Richardson1993; Richardson and others Reference Richardson, Swain and McCoull2001) or the occurrence of big winter storms, which cause waves to break over low lying areas of the isthmus and the presence of snow in winter. Predators such as birds and possibly spiders, could also be a factor. Monitoring in future years is required to establish any expansion of either species but it does not seem to be fast given that each species has probably been on the island for at least 80 years. The reason that collectors of terrestrial invertebrates did not detect them earlier is probably because they are likely to have ignored the amphipod, assuming it to be part of the marine littoral fauna. The isopod probably avoided earlier detection because of its cryptic habits. It is not extracted by Tullgren funnels nor does it fall into pitfall traps and hand observations have to be made using specific methods in particular habitats.
Assuming that the original site of introduction of the isopod was at Razorback Ridge, the most likely means of dispersal to the other two sites was by human activity, probably on the soles of boots as noted earlier. If so, the rate of spread is likely to increase with increased human visitation (Chown and others Reference Chown, Hull and Gaston2005) and possibly with climate change because the New Zealand distributions of both species is in warmer climates than that currently experienced by Macquarie Island.
Ethical aspects of removing a species accidentally introduced by human activities from a World Heritage Area must be considered as, if these species become widespread on the island, there is a risk of concomitant loss of native species (Farnsworth and Rosovsky Reference Farnsworth and Rosovsky1993; Hampicke Reference Hampicke1994). Slabber and Chown (Reference Slabber and Chown2002) recommend removal of an exotic isopod from Marion Island. By analogy, introduced vertebrates, such as the weka, cat, rabbit, rat and mouse have all been extirpated, or recommended for it, for the same reason. However the removal of either crustacean from the island appears impractical, as it would require removal of considerable areas of vegetation. Future work should consist of not only monitoring distribution, but also controlling spread caused by human visitation.
Acknowledgements
This work was carried out as part of two research projects on terrestrial invertebrates (ANARE 104 and ASAC 2397). ASAC 2397 was undertaken at the Department of Genetics at La Trobe University from 2003–2005, and at the School of Biological Sciences at Monash University since 2005. We thank the Australian Antarctic Division for financial and logistic support and are grateful for the support of the ANARE staff and use of the facilities on Macquarie Island. We would also like to thank Geoff Copson (Department of Primary Industries, Water and Environment), Peter Cusick and Noel Carmichael (Department of Tourism, Parks, Heritage and the Arts), Paul Sunnucks (Monash University), David Rounsevell (Griffiths University), Pat Selkirk (Macquarie University), Geoff Dannock, Murray Price, Pep Turner and Bruce Hull (Australian Antarctic Division) for valuable information of different kinds. Clive Crossley, Bruce Kemp, Peter Serov, Jean Jackson, and Robert Edwards all assisted with fieldwork. Robert Gibb provided much needed assistance with ArcMap to produce Figs. 1, 3, and 4, Nathalie Loussert provided Fig. 5, and Simon Joly helped with file transformations. We also thank two anonymous referees for helpful comments on the original manuscript.






