



Introduction
Climate change is predicted to increase the frequency and severity of agricultural droughts (periods of soil moisture deficit) across most of the United States (USGCRP 2017). These trends are likely to occur even in regions like the Northeast, which will also receive increased precipitation due to increased temperatures (Hayhoe et al. Reference Hayhoe, Wake, Huntington, Luo, Schwartz, Sheffield, Wood, Anderson, Bradbury, DeGaetano, Troy and Wolfe2007; USGCRP 2017). The vulnerability of food production systems to drought is dependent on a combination of biological factors (e.g., drought resistance in crop species) and management factors (e.g., irrigation capacity; Chaves and Oliveira Reference Chaves and Oliveira2004; Lu et al. Reference Lu, Carbone, Huang, Lackstrom and Gao2020; Wolfe et al. Reference Wolfe, DeGaetano, Peck, Carey, Ziska, Lea-Cox, Kemanian, Hoffmann and Hollinger2018). Another determinant of vulnerability, the effect of drought on competition from agricultural weeds, combines biological and management components (DiTommaso et al. Reference DiTommaso, Zhong and Clements2014; Ramesh et al. Reference Ramesh, Matloob, Aslam, Florentine and Chauhan2017; Varanasi et al. Reference Varanasi, Prasad and Jugulam2016). Understanding the relationship between drought stress and weed pressure is crucial to understanding the risks posed by climate change (Korres et al. Reference Korres, Norsworthy, Tehranchian, Gitsopoulos, Loka, Oosterhuis, Gealy, Moss, Burgos, Miller and Palhano2016; Ramesh et al. Reference Ramesh, Matloob, Aslam, Florentine and Chauhan2017).
Corn is the world’s leading cereal crop (United Nations FAO 2020) and a major commodity in New York State, accounting for more than 400,000 harvested hectares (45% silage; USDA-NASS 2020). Corn production is threatened by direct effects of climate change, including drought (Korres et al. Reference Korres, Norsworthy, Tehranchian, Gitsopoulos, Loka, Oosterhuis, Gealy, Moss, Burgos, Miller and Palhano2016; McFadden et al. Reference McFadden, Smith, Wechsler and Wallander2019; Prasad et al. Reference Prasad, Gunn, Rotz, Karsten, Roth, Buda and Stoner2018). In New York State, where field crops (including corn) are largely unirrigated, Sweet et al. (Reference Sweet, Wolfe, DeGaetano and Benner2017) surveyed field crop growers after a major drought and found that most had experienced substantial yield losses. Corn in the Northeast is also likely to suffer worsening weed problems as troublesome weeds from warmer southern states shift their ranges northward due to climate change (McDonald et al. Reference McDonald, Riha, DiTommaso and DeGaetano2009). In addition to limiting species ranges, climatic factors can influence the intensity and outcome of weed-crop competition (Patterson Reference Patterson1995; Ramesh et al. Reference Ramesh, Matloob, Aslam, Florentine and Chauhan2017; Ziska and Dukes Reference Ziska and Dukes2011). However, it is often difficult to predict how increased abiotic stress will affect competition between corn and associated weed species.
Burcucumber (Figure 1, A and B) is a summer annual vine in the family Cucurbitaceae, native throughout the eastern United States and invasive across much of Europe and Asia (CABI 2019; USDA-NRCS 2021). It is listed as noxious in Delaware, Indiana, and Kentucky (USDA-NRCS 2021). Burcucumber is also a serious weed in the Northeast (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a; Messersmith et al. Reference Messersmith, Curran and Lingenfelter1998) and has been reported to be one of the most troublesome corn weeds in Pennsylvania (DiTommaso et al. Reference DiTommaso, Buck, Riha and McDonald2012). The most important trait contributing to burcucumber weediness is its vining habit, which allows the weed to climb on crop plants, interfere with harvesting, and even pull crops to the ground (VanGessel and Johnson Reference VanGessel and Johnson2019; Webb and Johnston Reference Webb and Johnston1981). Control of burcucumber is complicated by its continuous germination and rapid growth (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a, 2001b; Messersmith et al. Reference Messersmith, Curran, Hartwig, Orzolek and Roth1999; Smeda and Weller Reference Smeda and Weller2001). Despite these advantages, burcucumber may suffer severe reductions in growth and fecundity due to competition from corn (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a). This observation implies that increased crop competitiveness could contribute to burcucumber control.

Figure 1. Design of a 2-yr field study testing the effects of burcucumber competition and drought on silage corn in Ithaca, NY (A) Burcucumber and corn on June 29, 2018. (B) Burcucumber and corn on September 13, 2018. (C) Rainout shelters on June 29, 2018.
Burcucumber appears to thrive in moist environments. Before it became a serious agricultural problem, it was found “along river banks and in moist places” (Britton and Brown Reference Britton and Brown1913 p. 293). It still exhibits a preference for habitats with high rainfall (Gibson et al. Reference Gibson, Johnson and Hillger2005; Smeda and Weller Reference Smeda and Weller2001), reflected at a larger scale in its global distribution (CABI 2019). One explanation for these observations is the stimulatory effect of moisture on burcucumber germination (Önen et al. Reference Önen, Farooq, Tad, Özaslan, Gunal and Chauhan2018; Smeda and Weller Reference Smeda and Weller2001). Burcucumber germination is more sensitive to moisture deficit than corn germination (Mann et al. Reference Mann, Rieck and Witt1981; VanGessel and Johnson Reference VanGessel and Johnson2019). After germination, burcucumber develops a shallow root system that has been characterized as fibrous or as a branched taproot (Virginia Tech CALS n.d.). Burcucumber roots may be shallower than corn roots, which are also fibrous but often reach depths of 1.5 m or more (Archontoulis and Licht Reference Archontoulis and Licht2017; Feldman Reference Feldman1994; Ordóñez et al. Reference Ordóñez, Castellano, Hatfield, Helmers, Licht, Liebman, Dietzel, Martinez-Feria, Iqbal, Puntel, Córdova, Togliatti, Wright and Archontoulis2018). Corn and burcucumber also differ in that corn uses the C4 photosynthetic pathway, which promotes water use efficiency relative to the ancestral C3 pathway used by burcucumber (Ehleringer and Monson Reference Ehleringer and Monson1993; Leakey et al. Reference Leakey, Ferguson, Pignon, Wu, Jin, Hammer and Lobell2019).
The objective of this study was to evaluate the effects of simulated drought and increasing burcucumber density on silage corn in New York State. We hypothesized that 1) both stressors would decrease silage yield, but 2) the adverse effect of burcucumber on silage yield would decrease under drought. Our second hypothesis reflected the assumption that burcucumber, which frequently grows in moist environments, would be severely inhibited by drought.
Materials and Methods
The experiment was conducted in 2018 and 2019 at the Caldwell Field Research Facility in Ithaca, NY (42.45N, 76.46W). The soil was a Williamson very fine sandy loam (USDA-NRCS 2018) and the field site had been used for corn in previous years. In the study years, the field was plowed, disked, and harrowed, then corn was planted on May 28, 2018, and May 21, 2019. A 95-d glyphosate-tolerant (Roundup Ready®) silage corn hybrid (DKC 45-66) was planted in 76-cm rows at 71,660 plants ha−1. Fertilizer (10-20-20 N-P-K, 224 kg ha−1) was applied at planting. No irrigation was provided. Weeds other than burcucumber were controlled with regular hand weeding, interrow cultivation (June 21, 2018, and July 3, 2019), and glyphosate. Glyphosate (GlyStar Plus®, 1.12 kg ai ha−1) was applied prior to burcucumber transplanting (week of June 16, 2019, only).
Burcucumber seeds, purchased from Prairie Moon Nursery in Winona, MN, were cold stratified for 90 d before pretransplant germination in a greenhouse. Germination in 2018 was poor, so burcucumber seedlings were collected from a field in Barton, NY (47 km from the field site), on May 29, 2018. Seedlings selected for transplant were 15 cm or shorter. In 2019, seedcoats were clipped after stratification, which improved greenhouse germination. Burcucumber seedlings were grown under greenhouse conditions until transplanting into the corn rows on June 8, 2018, and June 25, 2019, when burcucumber and corn seedlings were both approximately 15 cm tall (1 to 3 leaves).
Plots were arranged according to a randomized complete block design with four blocks (replicates) separated by eight corn rows. Each block contained four plots, corresponding to burcucumber transplanting densities of 0, 0.5, 2, and 3 plants m−2 within rows. There were 320 cm (rows 1, 3) or 213 cm (rows 2, 4) between burcucumber seedlings in the 0.5 plants m−2 treatment, 64 cm between burcucumber seedlings in the 2 plants m−2 treatment, and 43 cm between burcucumber seedlings in the 3 plants m−2 treatment. These densities were selected because they represent the range of burcucumber densities observed in New York State crop fields (A DiTommaso, personal observation). Each plot was 3 m (four 76-cm corn rows) wide and 6 m long. Plots were separated by 4.5 m within blocks. Each plot was split into two subplots (3 by 3 m), corresponding to drought and no-drought treatments. The drought stress was imposed on June 22, 2018, and July 11, 2019, by covering drought subplots with rainout shelters (Figure 1C), which were 2.7 m wide by 3 m long by 1.5 m tall. The shelters were constructed from steel electrical conduit frames with 6 MIL high-clarity greenhouse film (Sun Master® Pull and Cut) forming a roof and extending partway down the sides. Gutters attached to drainage tile were used to remove water from subplots. We also constructed soil berms to reduce aboveground water flow, so plants under shelters relied primarily on stored water and belowground diffusion. Rainout shelters, including the shelters used in this study (KM Averill, unpublished data; Hunter et al. Reference Hunter, Kemanian and Mortensen2021), can impose drought stress with minimal microclimate effects (see also Gray et al. Reference Gray, Strellner, Puthuval, Ng, Shulman, Siebers, Rogers and Leakey2013; Kant et al. Reference Kant, Thoday-Kennedy, Joshi, Vakani, Hughes, Maphosa, Sadler, Menidis, Slater and Spangenberg2017). The shelters were removed from the drought subplots on July 30, 2018, and July 31, 2019.
Available soil moisture (the percentage of soil water available to plant roots) was estimated in each subplot using fixed-in-place gypsum block electrodes (Delmhorst GB-1; Towaco, NJ) in the center of each subplot (15 cm depth). We used the conversion chart in the manual to estimate available soil moisture from the (unitless) meter readings. These measurements were taken on 12 dates between July 3, 2018, and September 7, 2018, and on 11 dates between July 15, 2019, and September 10, 2019. Corn and burcucumber aboveground plant material were harvested on September 12 to 14, 2018, and September 9 to 10, 2019. Vegetative growth was complete by harvest, but seeds were still ripening. Aboveground plant material was harvested from 1.14-m2 quadrats in the center of each subplot. Creeping burcucumber stems were also harvested from the half-interrow areas to each side of the quadrats. We recorded the number of corn stems in each quadrat, which ranged from 4 to 17 (depending on establishment and mortality rates). Harvested corn was immediately weighed to determine fresh silage yield. All harvested corn and burcucumber was dried to determine silage moisture content, dry silage yield, and dry burcucumber biomass. Corn was dried for 14 d in a greenhouse (15-h photoperiod, 25/20 C) and burcucumber was dried in a drying oven (55 C).
Data analysis was carried out in R (4.0.2, R Core Team 2021). Unless otherwise noted, average values are presented as mean ± standard error. Factors affecting available soil moisture, corn fresh silage yield, corn dry silage yield, and burcucumber dry biomass were analyzed with mixed-effects analysis of variance (packages lme4, lmerTest). For biomass models, we began with full models and selected reduced models by backward elimination of random-effect terms followed by backward elimination of fixed-effect terms (Satterthwaite’s method, α = 0.1 for random effects and α = 0.05 for fixed effects, package lmerTest). The full models included the fixed effects of drought treatment, burcucumber planting density, year, number of corn stems in the sampled quadrat, and all interactions, in addition to the random effect of block. All residuals were normal and homoscedastic without data transformation. Graphs (packages ggplot2, ggpubr) show linear regressions with 95% confidence intervals.
Results and Discussion
Weather and Soil Conditions
At the field site, the 30-yr average rainfall (1981 to 2010) from June to September was 385 mm (Northeast Regional Climate Center 2021). Rainfall from June to September was 416 mm in 2018 and 401 mm in 2019 (Northeast Regional Climate Center 2021), so water was unlikely to be limiting in the no-drought treatments. The 30-yr average temperature from June to September was 18.4 C (Northeast Regional Climate Center 2021). In 2018 and 2019, the average temperatures were 19.5 C and 18.3 C, respectively (Northeast Regional Climate Center 2021). These data suggest that temperature conditions were fairly typical for the field site in both years, although it is possible that slightly warmer temperatures in 2019 impacted production.
Available soil moisture as measured indirectly using gypsum blocks was lower in drought plots (45% ± 2% in 2018 and 43% ± 2% in 2019) than no-drought plots (73% ± 1% in 2018 and 69% ± 2% in 2019) over the period between shelter placement and shelter removal. Over the period between shelter placement and harvest, available soil moisture remained lower in drought plots (47% ± 1% in 2018 and 52% ± 2% in 2019) than no-drought plots (69% ± 1% in 2018 and 68% ± 1% in 2019). Available soil moisture was lower in the drought treatment (P < 0.001) regardless of which period was considered. Therefore, we conclude that rainout shelters successfully imposed a drought stress under the study conditions (Figure 2).
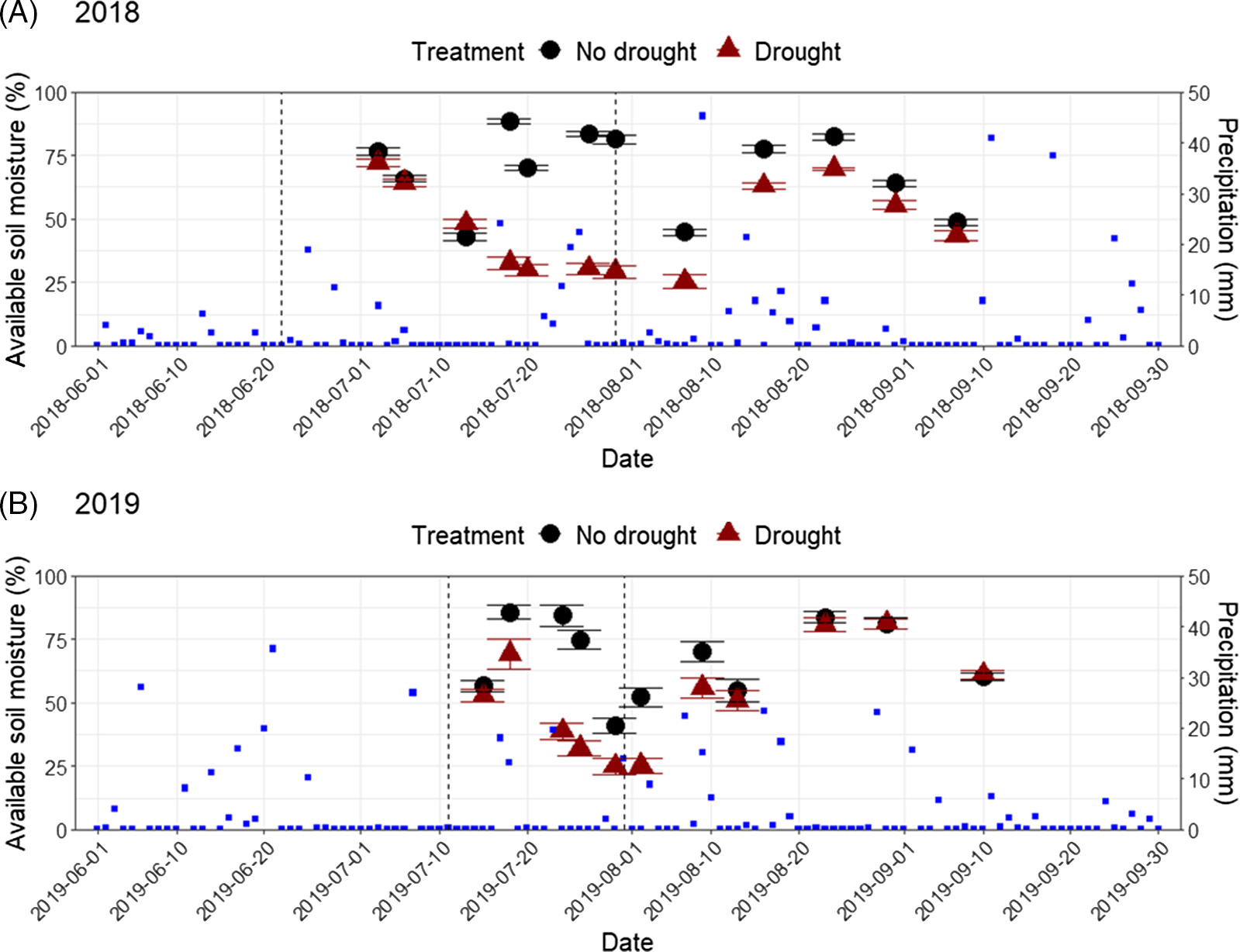
Figure 2. Precipitation and available soil moisture (mean ± SE, as measured indirectly using gypsum blocks) in (A) 2018 and (B) 2019 in Ithaca, NY. The no-drought control (black circles) received 416 mm (2018) and 401 mm (2019) rainfall between June and September (blue squares; Northeast Regional Climate Center 2021). In the drought treatment (red triangles), rainout shelters were placed in the field on June 22, 2018, and July 11, 2019, and removed on July 30, 2018, and July 31, 2019 (dashed lines).
Silage Corn Yield and Burcucumber Biomass
As hypothesized, we found negative effects of both burcucumber planting density and drought on silage corn yield. The reduced model of fresh silage yield contained the fixed effects of burcucumber planting density (P = 0.008), drought treatment (P < 0.001), year (P = 0.1), the interaction between drought treatment and year (P = 0.007), and number of corn stems in the sampled quadrat (P < 0.001), in addition to the random effect of block. Increasing burcucumber planting density decreased fresh silage yield (Figure 3A). With other factors held constant, fresh silage yield was reduced by approximately 1,700 kg ha−1 per burcucumber plant m−2.
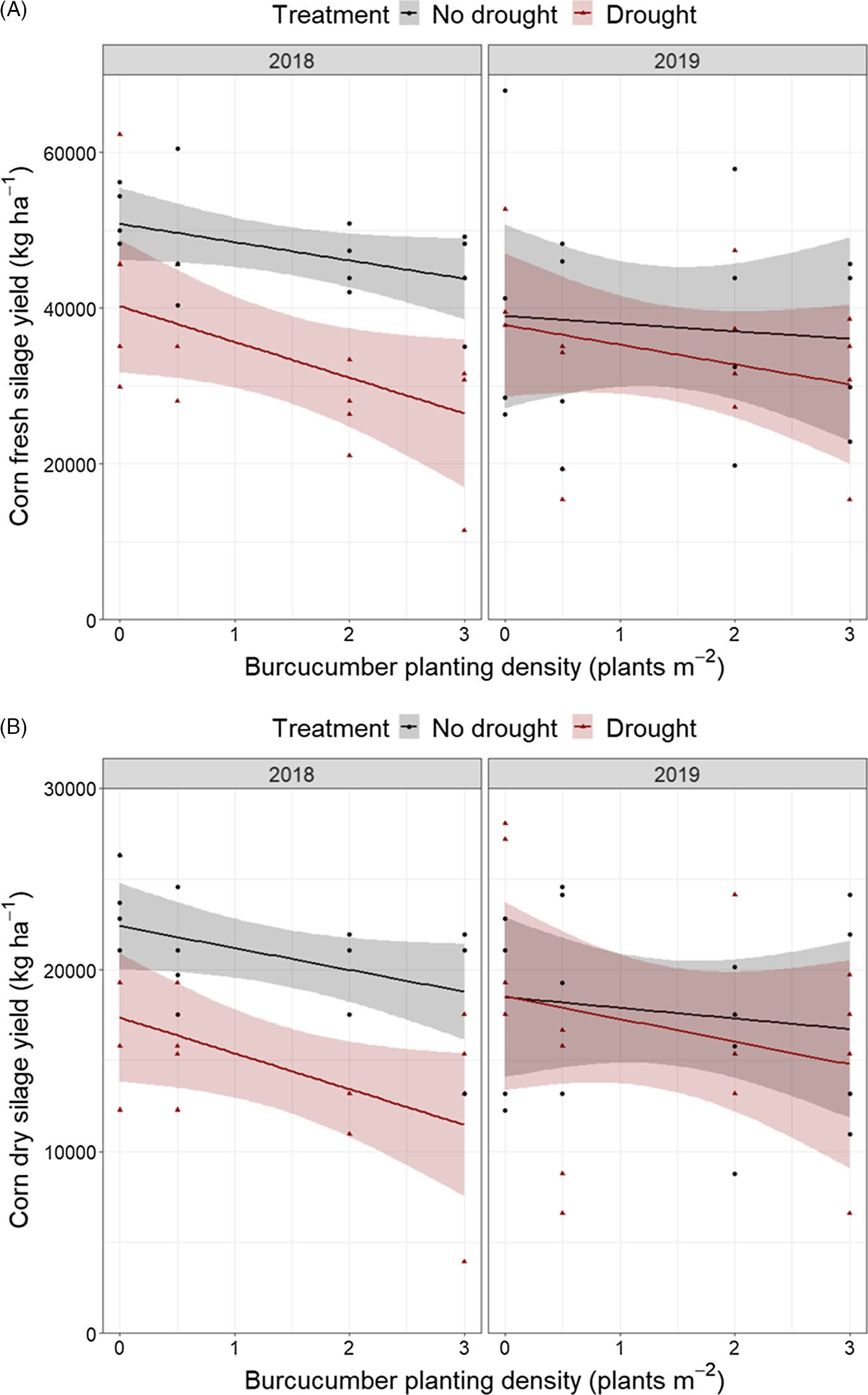
Figure 3. Effects of burcucumber planting density and drought treatment on (A) corn fresh silage yield and (B) corn dry silage yield in Ithaca, NY, in 2018 (left) and 2019 (right). Silage was harvested on September 14, 2018, and September 9 to 10, 2019. Fitted lines in red (drought) and black (no drought) represent linear models with 95% confidence intervals.
Fresh yield also decreased with drought. In 2018, fresh yield was 29% lower in the drought treatment (34,000 ± 3,000 kg ha−1) relative to the no-drought control (48,000 ± 2,000 kg ha−1). In 2019, fresh yield was 9% lower in the drought treatment (34,000 ± 3,000 kg ha−1) relative to the no-drought control (38,000 ± 3,000 kg ha−1). This interannual variation was consistent with the greater disparity in available soil moisture (between drought and no-drought treatments over the period from shelter placement to harvest) in 2018 vs. 2019. We were unable to determine whether higher yields in the no-drought treatment in 2018 vs. 2019 reflected weather conditions or other factors. The number of corn stems in the sampled quadrat (no-drought treatment) averaged 14.4 ± 0.4 in 2018 and 9.1 ± 0.7 in 2019, suggesting a difference in establishment or mortality rates. The interaction between burcucumber planting density and drought treatment was not included in the reduced model. This finding was incompatible with the hypothesis that drought would reduce the negative effect of burcucumber on yield. Instead, this finding suggests that burcucumber and drought have additive effects on corn yield and can be considered independently.
We observed similar trends in dry silage yield, as expected from the low variation in moisture content (54% ± 1% moisture; Figure 3B). The reduced model of dry silage yield contained the fixed effects of burcucumber planting density (P = 0.005), drought treatment (P < 0.001), year (P = 0.008), the interaction between drought treatment and year (P = 0.002), and number of corn stems in the sampled quadrat (P < 0.001), in addition to the random effect of block. Drought decreased dry yield by 29% in 2018 (21,000 ± 1,000 kg ha−1 to 15,000 ± 1,000 kg ha−1) and by 5% in 2019 (18,000 ± 1,000 kg ha−1 to 17,000 ± 2,000 kg ha−1).
Lastly, we tested whether burcucumber planting density, drought treatment, year, number of corn stems in the sampled quadrat, or their interactions affected dry burcucumber biomass. The reduced model contained the fixed effects of burcucumber planting density (P < 0.001), drought (P = 0.003), year (P = 0.03), number of corn stems (P = 0.7), and the interaction between year and number of corn stems (P = 0.02). Burcucumber biomass increased with increased planting density (Figure 4). Burcucumber biomass also increased in the drought treatment. Burcucumber biomass tended to be higher in 2019, but (in 2019 only) decreased with increasing numbers of corn stems in the sampled quadrat.
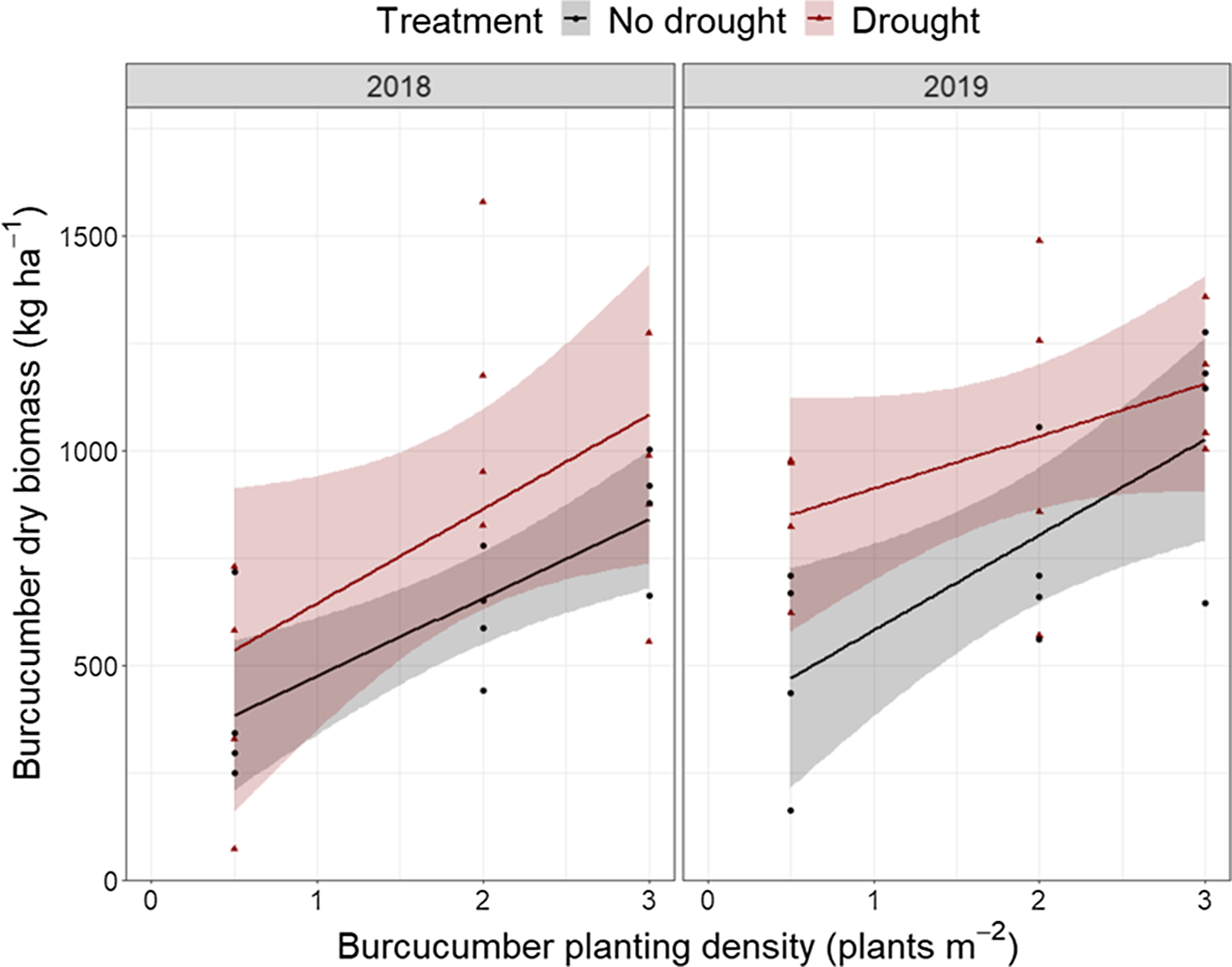
Figure 4. Effects of burcucumber planting density and drought treatment on dry burcucumber biomass in Ithaca, NY, in 2018 (left) and 2019 (right). Biomass was harvested on September 12 to 13, 2018, and September 9 to 10, 2019. Fitted lines in red (drought) and black (no drought) represent linear models with 95% confidence intervals.
Although the negative effect of burcucumber planting density on silage corn was modest, it was statistically significant. This finding is consistent with previous studies in grain corn. For example, Esbenshade et al. (Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001b) reported that the combination of glufosinate and atrazine reduced burcucumber density (4.9 to 1.2 plants m−2) and burcucumber biomass (94% lower), resulting in increased corn grain yield (26% higher). Uncontrolled burcucumber reduces grain yield by competing for resources and/or interfering with harvesting (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001b; Messersmith et al. Reference Messersmith, Curran, Roth, Hartwig and Orzolek2000; VanGessel and Johnson Reference VanGessel and Johnson2019; Webb and Johnston Reference Webb and Johnston1981). We observed that many plants were highly entangled with vines, which could complicate mechanical harvesting even in silage corn. Data from hand-harvested plots may be considered a conservative estimate of potential yield losses (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001b).
An important difference between silage and grain corn is that silage is harvested earlier. In this study, the silage harvest was completed by mid-September. According to 5-yr average data on crop progress in New York State, silage corn is 65% harvested by October 11 while grain corn is only 10% harvested (Fike Reference Fike2020). The difference in harvest timing allows growers to salvage burcucumber-infested fields through an early silage harvest (Messersmith et al. Reference Messersmith, Curran and Lingenfelter1998). This approach works because it reduces the number of seeds returned to the field and because ensiling kills immature (but not mature) burcucumber seeds (Messersmith et al. Reference Messersmith, Curran and Lingenfelter1998). More generally, it relies on the fact that burcucumber seeds mature fairly late in the season (Messersmith et al. Reference Messersmith, Curran and Lingenfelter1998), a characteristic that may be associated with the continuous germination of this annual species (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a, 2001b; Messersmith et al. Reference Messersmith, Curran, Hartwig, Orzolek and Roth1999; Smeda and Weller Reference Smeda and Weller2001). Late burcucumber establishment tends to reduce seed production without preventing it altogether (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a). Taken together, these observations suggest that burcucumber could have a greater effect on grain corn and that its effect on silage corn could change if warming temperatures lengthen the growing season in the Northeast (USGCRP 2017).
The finding that drought decreased silage yield (Figure 3) is consistent with the regional and global risk of corn yield losses due to soil moisture deficit (Korres et al. Reference Korres, Norsworthy, Tehranchian, Gitsopoulos, Loka, Oosterhuis, Gealy, Moss, Burgos, Miller and Palhano2016; McFadden et al. Reference McFadden, Smith, Wechsler and Wallander2019; Prasad et al. Reference Prasad, Gunn, Rotz, Karsten, Roth, Buda and Stoner2018; Sweet et al. Reference Sweet, Wolfe, DeGaetano and Benner2017). More surprisingly, the absence of an antagonistic interaction between drought and burcucumber density (Figure 3) indicates that drought did not make burcucumber less competitive against corn. This conclusion is also supported by the finding that drought increased burcucumber aboveground biomass (Figure 4). This finding was unexpected because burcucumber tends to inhabit moist environments (Britton and Brown Reference Britton and Brown1913; Gibson et al. Reference Gibson, Johnson and Hillger2005; Smeda and Weller Reference Smeda and Weller2001). However, it is entirely possible that burcucumber growth would have been inhibited by a more severe drought stress or a stress imposed at a more vulnerable developmental stage.
Multiple factors may jointly account for burcucumber’s competitive effect on corn. Competition for water may have contributed to this effect. However, the absence of an interaction between burcucumber planting density and drought treatment, in combination with the fact that burcucumber does not have deep roots (European and Mediterranean Plant Protection Organization 2010; Rashid et al. Reference Rashid, Uddin, Sarkar, Parveen and Asaeda2019), suggests that competition for water was not the only mechanism underlying crop yield losses. Some authors have reported that interference with harvesting (i.e., burcucumber pulling crop plants to the ground) was the main mechanism underlying crop yield losses because burcucumber plants were not very competitive for light or nutrients (European and Mediterranean Plant Protection Organization 2010; Webb and Johnston Reference Webb and Johnston1981). Other authors have reported that shading is an important mechanism by which burcucumber suppresses competitors (Farooq et al. Reference Farooq, Tad, Onen, Gunal, Caldiran and Ozaslan2017; Hartzler Reference Hartzler2015). In general, shallow lateral roots are useful for acquiring immobile nutrients (Lynch Reference Lynch1995). It is possible that water, light, and nutrients are all relevant to competition between corn and burcucumber (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a, 2001b). Although we did not observe corn being pulled all the way to the ground, we did find that burcucumber climbed on and over corn plants throughout the growing season. It therefore seems likely that competition for light accounted for much of the competitive effect of burcucumber on corn in this experiment. A greenhouse experiment might help distinguish competition for light from competition for water and nutrients.
The fact that drought increased burcucumber biomass could indicate that burcucumber growth in the no-drought treatment was limited by competition from corn for factors other than water. Esbenshade et al. (Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a) reported a very strong competitive effect of corn on burcucumber: competition reduced burcucumber dry matter by 96% and seed production by 98%. When competitive crops prevent weeds from acquiring resources, weed pressure decreases (Buhler Reference Buhler2002; Swanton and Weise Reference Swanton and Weise1991). In our experiment, competition from corn plants may have limited the availability of light and/or nutrients to burcucumber in the no-drought treatment but had a weaker impact in the drought treatment. A similar explanation was proposed by Mojzes et al. (Reference Mojzes, Ónodi, Lhotsky, Kalapos and Kröel-Dulay2020), who suggested that horseweed [Conyza canadensis (L.) Cronquist] exhibited improved performance in drought plots because the drought stress alleviated competitive pressure from perennial grasses. However, we cannot explain why corn yield losses due to drought were stronger in the first study year but the effect of drought on burcucumber biomass was similar between years. Although rainout shelters are a proven method of imposing drought with minimal microclimate effects (Hunter et al. Reference Hunter, Kemanian and Mortensen2021; see also Gray et al. Reference Gray, Strellner, Puthuval, Ng, Shulman, Siebers, Rogers and Leakey2013; Kant et al. Reference Kant, Thoday-Kennedy, Joshi, Vakani, Hughes, Maphosa, Sadler, Menidis, Slater and Spangenberg2017), it is possible that the drought and no-drought treatments differed in some unplanned way. Future work should consider including a set of burcucumber monoculture plots, which would allow statistical testing for an interaction between the presence of corn and drought treatment on burcucumber biomass.
Major findings of this experiment include the negative effects of burcucumber competition and drought on fresh and dry silage yield. We did not find an interaction between these main effects, indicating that drought does not modify the competitive effect of burcucumber. Although burcucumber is often found in wet areas, New York State growers should not expect that burcucumber will be impeded by soil water deficit or eliminated under climate change. This damaging annual vine should be controlled in both silage and grain corn, regardless of water availability.
Acknowledgments
We thank Gene Sczepanski and other Cornell Farm Services employees who completed field preparation, planting, fertilizing, cultivating, and spraying. We also thank the many undergraduates who contributed to various aspects of the project including Danilo Pivaral, Robert Galbraith, Yuan Li, Maria Paula Osuna Barreto, and Julie Mudasumbwa. Lastly, we thank the associate editor and reviewers for their helpful comments on this manuscript. This work was supported by the USDA–National Institute of Food and Agriculture, Hatch project #1013464. No conflicts of interest have been declared.




