


Introduction
During the Devonian, Iberia was situated in the northwestern margin of Gondwana and separated from Laurussia by the narrow northeastern-trending Rheic Ocean. As part of Gondwana, Iberia moved northward, reaching about 35°S during Givetian times (Scotese, Reference Scotese2000, Reference Scotese2001; Nance et al., Reference Nance, Gutiérrez-Alonso, Keppie, Linnemann, Murphy, Quesada, Strachan and Woodcock2012). Thus, the Devonian sediments were deposited in subtropical seas.
Devonian crinoids from the Cantabrian Zone in North Spain are well documented, and the first species were described by de Verneuil in the 19th century (de Verneuil, Reference Verneuil and de1850). Several authors described crinoids from this area (Oehlert, Reference Oehlert1897; Schmidt, Reference Schmidt1931; Almela and Revilla, Reference Almela and Revilla1950; Sieverts Doreck, Reference Sieverts Doreck1951), but the first comprehensive monograph on Spanish crinoids was Breimer (Reference Breimer1962). He described five new genera and sixteen new species, with fourteen of these taxa previously unreported from this area. Most of the crinoids studied by Breimer were from the Lower and Middle Devonian. He also described a small number of Carboniferous species. Since, only a few species have been reported either by professional paleontologists (Le Menn, Reference Le Menn1976; Webster, Reference Webster1976; Pidal, Reference Pidal1984, Reference Pidal2008; Kammer, Reference Kammer2001; Zamora et al., Reference Zamora, Álvaro, Arbizu, Colmenar, Esteve, Fernández-Marinez, Fernández, Gutiérrez-Marco, Andrés, Villas, Waters, Zamora and Rábano2015) or amateurs (e.g., Hauser, Reference Hauser2010, Reference Hauser2015; Hauser and Landeta, Reference Hauser and Landeta2007, Reference Hauser and Landeta2012, Reference Hauser and Landeta2013).
Apart from the well-known fauna of the Cantabrian Zone, other crinoids from the Spanish Devonian are unknown and only fragments have been reported in general works (Carls, Reference Carls1965). Le Menn (Reference Le Menn1985) mentioned the species Pterinocrinus tenuibrachiatus Le Menn, Reference Le Menn1976 in the Mariposas Formation. He also described Aragocrinus molinoesnsis Le Menn, Reference Le Menn1988, which corresponds to the distal holdfast from an unknown crinoid. Carls and Valenzuela-Ríos (Reference Carls, Valenzuela-Ríos, García-López and Bastida2002) also mentioned the presence of Seilloucrinus verneuili (Cailliaud, Reference Cailliaud1861) and Asperocrinus minumus LeMenn, Reference Le Menn1976 in the Mariposas Formation based apparently on isolated columnals that have been neither described nor figured. Lastly, Herrera and Villas (Reference Herrera and Villas2013) figured a rich collection of crinoid columnals from the Mariposas Formation, including Botryocrinus sp. and several morphotypes placed in open nomenclature.
In this paper, we describe a new crinoid fauna based on articulated specimens from the Mariposas Formation (Emsian), which crops out in the eastern Iberian Chain. A rich shelly fauna that includes trilobites, corals, mollusks, brachiopods, and crinoidal fragments (Carls, Reference Carls1965) has been reported previously from this area. Material described here is from large exposures provided by road construction. Five additional taxa are described from the Mariposas Formation, including three new species and two taxa left in open nomenclature. This fills an important gap in the knowledge of crinoids from Spain, and this fauna is compared to other Spanish faunas.
Geological setting
A complete Ordovician to Devonian succession crops out in the vicinity of Fombuena, Luesma, and Santa Cruz de Nogueras (Fig. 1.1), in the eastern Iberian Chain. The complete Devonian thickness is ~4000 m, 95% of which is comprised of siliciclastic rocks; but due to complicated tectonics, a complete Devonian section is lacking. Carls (Reference Carls1965) was a pioneer in describing these Devonian strata in detail, mapping the areas, and providing information on biostratigraphy. German and Spanish disciples of Carls have also worked in the area for the last half century and demonstrated the worldwide importance of Devonian strata around the axial depression of the Cámaras River. The best sequence of Rhenish faunas there is known from the Early Devonian (Carls and Valenzuela-Ríos, Reference Carls, Valenzuela-Ríos, García-López and Bastida2002).

Figure 1. (1) General geographic setting of the study area; (2) Paleozoic outcrops of the Iberian Chains, indicating the location of the studied area in the Herrera Unit (modified from Gozalo and Liñan, Reference Gozalo and Liñán1988); (3) geological map of the Santa Cruz de Nogueras area, with the location of the studied site (modified from Lendínez et al., Reference Lendínez, Ruiz and Carls1989).
The section studied herein crops out in a new road cut of the TE-V-1521 road from Santa Cruz de Nogueras to Bádenas villages (Fig. 1.2) where part of the Mariposas Formation is well exposed (Fig. 2). The coordinates of the outcrop are 41°6′31″N, 1°5′57″W. The Mariposas Formation is one of the most fossiliferous Devonian units in the Iberian Chains. It consists of a 200 m thick interval dominated by shales and carbonates. The faunas change from typical Rhenish facies of shallow water to Hercynian hemipelagic biofacies. The Mariposas Formation is subdivided into two members (d4a and d4b) (Fig. 2).
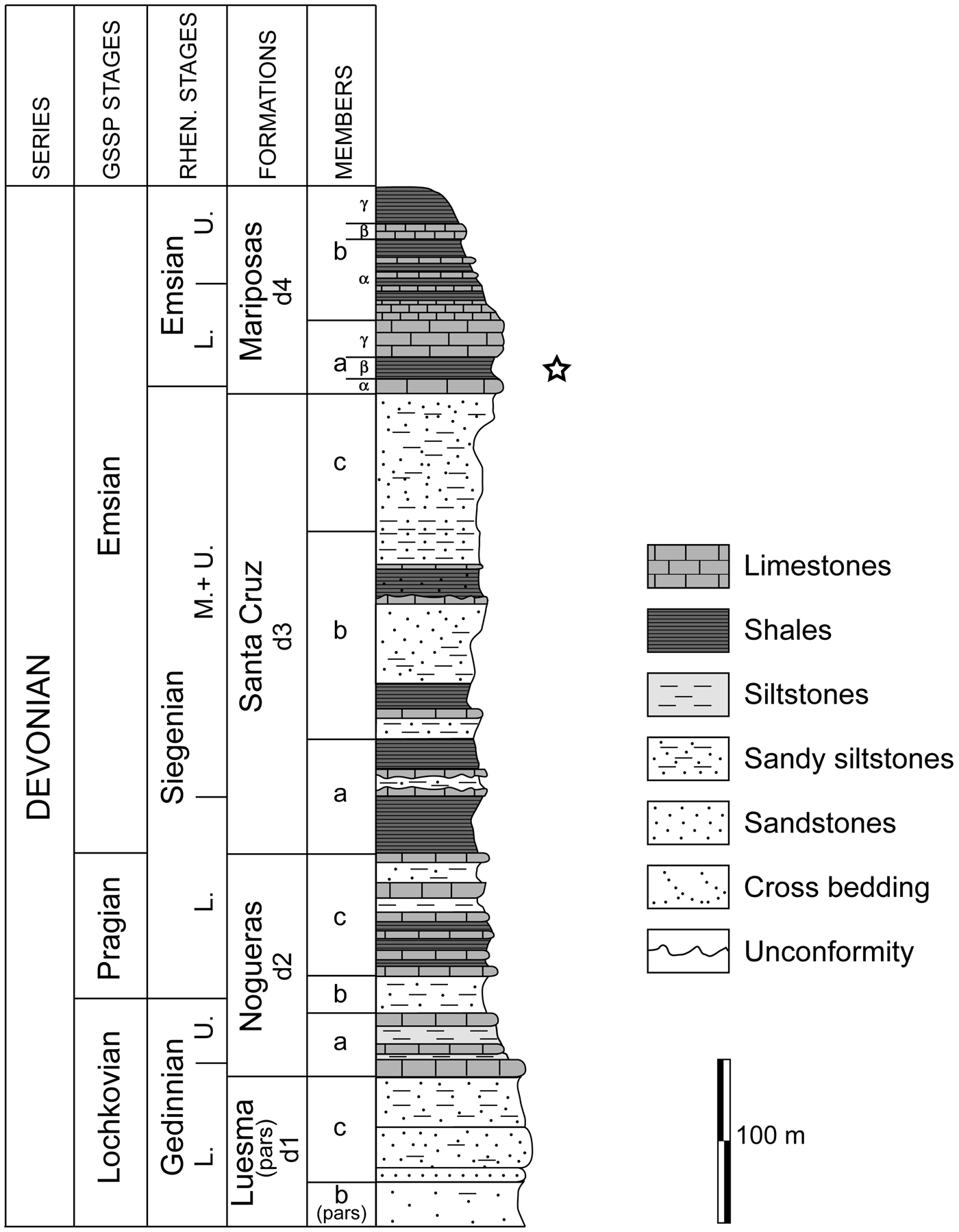
Figure 2. Stratigraphic section of the Lower Devonian of the Iberian Chains in the axial depression of the Cámaras River (simplified from Carls and Valenzuela-Ríos, Reference Carls, Valenzuela-Ríos, García-López and Bastida2002). Occurrence of crinoids noted.
Carls and Valenzuela- Ríos (Reference Carls, Valenzuela-Ríos, García-López and Bastida2002) provided a synthesis of the Devonian sequence from the Iberian Chains and indicated that the Mariposas Formation starts with an 8 m thick alternation of shelly limestones, bryozoan marls, and shales (submember d4aα), in which the lower boundary of the traditional German Emsian Stage is located (Carls, Reference Carls1987, Reference Carls, McMillan, Embry and Glass1988). Submember d4aβ is 20 m thick and contains Rhenish brachiopods, trilobites, and endemic conodonts of the genus Icriodus. There are rare Otarion and proetid trilobites, solitary rugose corals, thamnoporid and micheliniid tabulate corals, tentaculitoids, ostracodes, crinoids, and bivalves. Crinoids described in this paper have been collected from submember d4aβ. According to Villas et al. (Reference Villas, Colmenar, García-Alcalde, Carls, Herrera, Zamora, Valenzuela-Ríos, García-Joral, Villas and Baeza-Carratalá2018), the aforementioned interval yielding crinoids is dominated by brachiopods adapted to turbid waters. The low diversity of conodonts “Polygnathus” and abundance of “Icriodus” in the interbedded limestones indicate a shallow-water environment. The overlying submember (d4aγ) is 15–20 m thick, consists of a shelly crinoidal limestone, and contains a similar fauna to that of previous units, but also includes abundant atrypid brachiopods, some dacryoconarid tentaculitoids, and trilobites (scutellids and Phacops).
According to Carls and Valenzuela-Ríos (Reference Carls, Valenzuela-Ríos, García-López and Bastida2002), Rhenish faunas nearly disappear at the beginning of the d4b Member, with only a few trilobites (Asteropyginae) and brachiopods (Arduspirifer) present. The classic German Emsian begins 1 m below the pair of limestones beds in the middle of submember d4aβ, where the brachiopod Arduspirifer prolatestriatus (Mittmeyer, Reference Mittmeyer1973) marks the classical boundary in Germany (Villas et al., Reference Villas, Colmenar, García-Alcalde, Carls, Herrera, Zamora, Valenzuela-Ríos, García-Joral, Villas and Baeza-Carratalá2018). Thus, the levels containing the crinoids described herein are lower Emsian.
Materials and methods
Repositories and institutional abbreviations
New material described here is deposited in the Museo de Ciencias Naturales (University of Zaragoza) (MPZ) and the Museo Geominero, Madrid (MGM).
Systematic paleontology
Classification and terminology
The classification of crinoid higher taxa used here follows Ausich et al. (Reference Ausich, Kammer, Wright, Cole, Peter, Rhenberg, Zamora and Rábano2015), Cole (Reference Cole2017), Wright (Reference Wright2017), and Wright et al. (Reference Wright, Ausich, Cole, Peter and Rhenberg2017). Other aspects of suprageneric classification follow Moore and Teichert (Reference Moore and Teichert1978).
Morphologic terminology follows Ubaghs (Reference Ubaghs, Moore and Teichert1978). The plating of interrays is given in the standard abbreviated form as the number of plates in each range from the proximal-most plate to the last range before the tegmen. In the posterior interray, the primanal is indicated by “P” and the first interradial in regular interrays is indicated by “1.” The shorthand notation used to describe the nodal-internodal structure of heteromorphic columns is from Webster (Reference Webster1974). All measurements are in mm; * indicates that the measured feature is either incomplete or crushed. Other abbreviations include: ACH, aboral cup height; ACW, aboral cup width; AH, arm height; BH, basal circlet height; CH, column height; CrH, crown height; RH, radial plate height; RW, radial plate width; TH, tegmen height.
Class Crinoidea Miller, Reference Miller1821
Infraclass Eucamerata Cole, Reference Cole2017
Order Diplobathrida Moore and Laudon, Reference Moore and Laudon1943
Superfamily Rhodocrinitacea Roemer, Reference Roemer, Bronn and Geognostica1855
Family Rhodocrinitidae Strimple and Watkins, Reference Strimple and Watkins1955
Genus Acanthocrinus Roemer, Reference Roemer1850
Type species
Acanthocrinus longispina Roemer, Reference Roemer1850, by subsequent designation.
Other species
Acanthocrinus? benedettoi Haude, Reference Haude, Heinzeller and Nebelsick2004; A. brevispina Roemer, Reference Roemer1850; A. carlsi n. sp., A. faouensis Le Menn, Reference Le Menn1985; A. gracilior Jaekel, Reference Jaekel1895; A. gracilior var. vermicularis Schmidt, Reference Schmidt1942; A. heroldi Schmidt, Reference Schmidt1934; A. jaekeli Schmidt, Reference Schmidt1942; A. lingenbachensis Lehmann, Reference Lehmann1939; A. onondaga Goldring, Reference Goldring1923; A. rex Jaekel, Reference Jaekel1895; A. spinosus (Hall, Reference Hall1862) non Südkamp, Reference Südkamp2007; A. spinosus Südkamp, Reference Südkamp2007 non Hall, Reference Hall1862.
Occurrence
Devonian: Algeria, Argentina, Belgium, France, Germany, Spain, and United States.
Acanthocrinus carlsi new species
Figure 3.4
Holotype
MPZ2018/513.
Diagnosis
Acanthocrinus with short spines (as known), connection between interradial plating and the tegmen absent, ray ridges absent, anal tube absent (as known), permanently recumbent free arms, nodal columnals with prominent nodes, and a single internodal between nodals in the mesistele.
Occurrence
Mariposas Formation (Emsian), Iberian Chains, Spain.
Description
Calyx high vase shape. Calyx plating too poorly preserved to decipher many details. Downward projecting spine with rounded terminus from a more proximal calyx plate (perhaps a radial or basal plate).
Regular interrays not in contact; plating cannot be deciphered.
Free arms presumably 10 from large elliptical arm facets, permanently recumbent (Fig. 3.4), project downward more than twice the height of calyx. Free arms pinnulate, presumably branch (but this is not verified); brachials rounded biserial throughout free arms.

Figure 3. Emsian crinoids from the Mariposas Formation (NE, Spain). (1–3). Calyx and arms of Camerata indet.; specimen MPZ2018/518; (1) lateral view showing two articulated pinnulate arms; (2) lateral view showing a prominent spine on the tegmen; (3) lateral view showing arm articulations. (4) Acanthocrinus carsli n. sp. preserving complete calyx with arms folded down and stem; specimen MPZ2018/513. (5, 6) Platyhexacrinus santacruzensis n. sp. preserving complete calyx, proximal part of the arms, and stem; specimen MPZ2018/514. (7) Eucladid indet.; specimen MPZ2018/519. All specimens are latex casts whitened with ammonium chloride sublimate.
Low inverse cup-shaped tegmen projecting above position of arm openings; width of tegmen 50% of calyx width at position of arm openings; tegmen twice as wide as high. Tegmen plates small, polygonal; position of anal opening unknown.
Column circular, holomeric; heteromorphic N1 construction in medial stem. Distinctive spinose nodals (Fig. 3.4), nodals and internodals sutured along a high syzygial articulation prominent in lateral view. Proximal and distal stem, holdfast, and lumen unknown.
Etymology
After Peter Carls for his pioneering work in understanding the Devonian of the Iberian Chains.
Measurements
MPZ2018/513: CrH, 46.0; TH, 3.2; CH, 42.0*.
Remarks
Despite the relatively poorly preserved condition of the single Acanthocrinus carlsi n. sp. specimen from the Mariposas Formation, it has several features that make it distinct from other known species. Short spines (as known), connection between interradial plating and the tegmen absent, ray ridges absent, anal tube absent (as known), permanently recumbent free arms, nodal columnals with prominent nodes, and a single internodal between nodals in the mesistele render this taxon distinctive from other known species. Acanthocrinus carlsi n. sp. is compared to other species of Acanthocrinus in Table 1.
Table 1. Species-diagnosis table for Acanthocrinus.

Order Monobathrida Moore and Laudon, Reference Moore and Laudon1943
Superfamily Hexacrinitacea Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Family Hexacrinitidae Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Genus Platyhexacrinus Schmidt, Reference Schmidt1913
Type species
Platyhexacrinus inornatus Schmidt, Reference Schmidt1913; by subsequent designation.
Other species
Platyhexacrinus grandis Schmidt, Reference Schmidt1942; P. gurievskiensis Dubatolova, Reference Dubatolova1964; P. kegeli Schmidt, Reference Schmidt1931; P. nollenbachensis Hauser, Reference Hauser2015; P. ornatus (Schmidt, Reference Schmidt1913); P.? pisum Schmidt, Reference Schmidt1942; and Platyhexacrinus santacruzensis n. sp.
Occurrence
Devonian of France, Germany, Russia, and Spain.
Remarks
Platyhexacrinus is a relatively poorly known genus, due at least in part to taphonomy. This taxon has very thin aboral cup plates that are poorly sutured. Thus, members of this taxon disarticulated rapidly after death and were easily compressed during compaction, which may be contributing factor to this being a relatively rare crinoid.
Species characters for Platyhexacrinus are overall size, aboral cup plate sculpturing, presence or absence of impressed aboral cup plate sutures, relative height of basal circlet, height to width ratio of radial plates, radial facet width, and presence or absence of fixed primibrachials and secundibrachials.
Platyhexacrinus santacruzensis new species
Figures 3.5, 3.6, 4.1
- Reference Zamora, Álvaro, Arbizu, Colmenar, Esteve, Fernández-Marinez, Fernández, Gutiérrez-Marco, Andrés, Villas, Waters, Zamora and Rábano2015
camerate crinoid, Zamora et al., p. 245, fig. 24c, 24d.
Holotype
MPZ2018/514.
Diagnosis
Relatively small species of Platyhexacrinus, smooth aboral cup plate sculpturing, aboral cup sutures not impressed, basal circlet low, height to width ratio of radial plates ~1.0, radial facet ~33% of distal radial plate width, and fixed primibrachials and secundibrachials absent.
Occurrence
Mariposas Formation (Emsian), Iberian Chains, Spain.
Description
Calyx, small in size; only basal and radial plates in calyx that has a low cone shape; smooth plate sculpturing. Basal circlet visible in side view, sides concave, ~33% of aboral cup height, more than three times wider than high (Fig. 3.5). Basal plates five, ~1.4 times wider than high. Radial circlet ~67% of aboral cup height; radial plates presumably five, slightly higher than wide. Radial facets angustary, ~33% of distal radial plate width (Fig. 4.1), horseshoe shaped and protruding laterally.

Figure 4. Plate diagrams of Mariposas Formation crinoids. (1) Lateral view of Platyhexacrinus santacruzensis n. sp. calyx with primibrachials and two secundibrachials preserved in one ray; compare to Figure 3.5; scale bar, 1 mm (MPZ2018/514). (2, 3) Eucladid Indeterminate; compare to Figure 3.7 (MPZ2018/519); scale bar, 10 mm; (2) plate diagram of mostly disarticulated calyx and arm plates; (3) interpretation of plate homologies where possible: black filling, radial plates; E, E radial plate; CD, CD basal plate; D, D radial plate; DE, DE basal plate; IBr1, first primibrachial; IBr2, second primibrachial; RA, radianal plate.
Tegmen plates relatively large, but details unknown. Fixed interradial plates absent (Fig. 3.6), posterior interray unknown.
Free arms five, one arm division as known; first primibrachial free, tetragonal, approximately as high as wide, much smaller than radial plates and somewhat smaller than primaxil; second primibrachial axillary, pentagonal, 1.5 times higher than wide. Second secundibrachial axillary.
Proximal column circular, holomeric, heteromorphic with pattern of N212, nodals and priminternodals with thin, extended epifacet. Secundinternodals moderately convex laterally. Lumen unknown.
Etymology
Species dedicated to the locality of Santa Cruz de Nogueras where it was found.
Measurements
MPZ2018/514: CrH, 13.5*; ACH, 5.4; ACW, 6.1*; BH, 2.0; RH, 3.4; RW, 3.2; CH, 9.5*.
Remarks
Breimer (Reference Breimer1962) described several Devonian crinoids from Spain with very few plates fixed into the calyx, including species of Culicocrinus?, Cantharocrinus, Oenochoacrinus, and “Platycrinus.” Platyhexacrinus differs from all of these because the first primibrachial is free and no interradial plates are incorporated into a calyx. The previously described taxa all have one or two fixed brachials and at least one fixed primibrachial.
The small size, smooth sutures that are not impressed, low basal circlet, radial plate height/width of ~1.0, and no fixed primibrachials or secundibrachials distinguish this species of Platyhexacrinus. This new species is compared to other species in this genus in Table 2.
Table 2. Species-diagnosis table for Platyhexacrinus.

Superfamily Platycrinitacea Austin and Austin, Reference Austin and Austin1842
Family Hapalocrinidae Jaekel, Reference Jaekel1895
Genus Culicocrinus Müller, Reference Müller, Zeiler and Wirtgen1855
Type species
Platycrinus nodosus Müller, Reference Müller, Zeiler and Wirtgen1855, by original designation.
Other species
Culicocrinus breimeri n. sp.; C. confluentinus (Müller, Reference Müller, Zeiler and Wirtgen1855); C.? girardeauensis Brower, Reference Brower1973; C. inermis Jaekel, Reference Jaekel1895; C. inventriosus Schmidt, Reference Schmidt1942; C. inventriosus intemperans Schmidt, Reference Schmidt1942; C. nodosus confluentina (Müller, Reference Müller, Zeiler and Wirtgen1855); C. nodosus virgo (Schmidt, Reference Schmidt1942); C. rhenanus Follmann, Reference Follmann1891; C. rugosus de Koninck in Zeiler and Wirtgen, Reference Zeiler and Wirtgen1855; C. shotoriensis Webster et al., Reference Webster, Maples, Yazdi, Marcus and Waters2011; C. spinatus Jaekel, Reference Jaekel1895; C. spinosus Springer, Reference Springer1926.
Occurrence
Ordovician, United States; Silurian, United States; Devonian of Germany, Iran, and Spain.
Remarks
Webster and Webster (Reference Webster and Webster2014) list 10 species in Culicocrinus, two of which have subspecies. These taxa range in age from the Ordovician to the Pennsylvanian. The Devonian forms are morphologically distinct from the earlier and later forms (C.? girardeauensis, Ordovician; C. spinosus, Silurian; and C. shotoriensis, Pennsylvanian). The Devonian forms all have fixed brachials through at least a few secundibrachials. The non-Devonian species lack fixed secundibrachials and are tentatively not regarded as species within Culicocrinus for the discussion below. Also, two species listed by Webster and Webster (Reference Webster and Webster2014), C. rhenanus and C. rugosus, are nomina dubia; and C. confluentinus is too poorly understood to compare to other taxa. Thus, five species are considered for comparison to Culicocrinus breimeri n. sp.
Species level characters for Devonian Culicocrinus are the following: relative height of the basal circlet, nature of the rim of the basal concavity, nodes on the basal plates, nodes on the radial plates, number and size of interradial plates, and general character of the arms.
Culicocrinus breimeri new species
Figure 5
- Reference Breimer1962
Hapalocrinidae cf. Culicocrinus nodosus (Müller, Reference Müller, Zeiler and Wirtgen1855), Breimer, Reference Breimer1962, p. 122, pl. 11, figs. 1–4.
Holotype
MPZ2018/515.
Diagnosis
Culicocrinus with relatively high basal circlet, a short rim is present around the periphery of the basal concavity, two circular nodes project outward on the radial plates, more than one small interradial plate in regular interrays, and free arms relatively slender.
Occurrence
Breimer (Reference Breimer1962) figured a single specimen (also figured here) from an unknown locality and indicated that only one specimen was in the collection from Orzonaga (León) from levels near the Coladilla-Santa Lucía Formation boundary. The new material described herein comes from a known horizon (d4aβ) in the Mariposas Formation, so we prefer to choose this known horizon as the type level for Culicocrinus breimeri n. sp. These new specimens are from Santa Cruz de Nogueras (Teruel, NE Spain); Mariposas Formation (Emsian), Iberian Chains, Spain.
Description
Calyx medium globe shape, approximately as high as wide; smooth plate sculpturing other than nodes noted below; plate sutures flush with plate surface.
Basal circlet visible in side view, ~25% of calyx height, pentagonal outline from basal view; basal concavity outlined by short rim (Fig. 5.3). Basal plates three, unequal in size, azygous basal plate in AB interray. Circular nodes in an interradial position around basal circlet (Fig. 5.2, 5.5), which results in one or two circular nodes per plate depending on plate size; nodes project outward.

Figure 5. Emsian crinoids from the Coladilla-Santa Lucía formations boundary in the Cantabrian Zone (1–3) and Mariposas Formation (4–7) in the Iberian Chains, North Spain. (1–3) Original specimen of Culicocrinus breimeri n. sp. figured by Breimer (Reference Breimer1962) as Hapalocrinidae cf. Culicocrinus nodosus; unknown locality, horizon probably corresponds to the Coladilla-Santa Lucía Formation boundary (according to Breimer, Reference Breimer1962) (MGM2996D). (4, 5) Culicocrinus breimeri n. sp. from the Mariposas Formation; (4) lateral view showing calyx, arms, and proximal stem; (5) oblique view of the calyx showing plate boundaries and ornamentation (MPZ2018/515); latex cast whitened with ammonium chloride sublimate; (6, 7) Culicocrinus breimeri n. sp. MPZ2018/517 and MPZ2018/516.
Radial circlet uninterrupted by interradial plates, ~50% of calyx height; radial plates heptagonal, wider than high; much larger than basal plates; two circular nodes on each radial plate at mid-height, nodes project outward.
Regular interrays in contact with tegmen, not depressed (Fig. 5.2). One small, fixed interradial plate, higher than wide, sutured to narrow upper shoulders of subjacent radial plates; interradial oral surface plates sutured distally to fixed interradial plate of regular interrays.
Posterior interray with small primanal sutured to narrow upper shoulders of radial plates beneath, one small plate sutured to each side of primanal and to other plating distally (not preserved) that leads to anus.
Fixed brachials ~24% of calyx height. Two fixed primibrachials; first primibrachial approximately three times as wide as high; second primibrachial axillary, pentagonal. At least first secundibrachials fixed.
Ten free arms that branch once, pinnulate; brachials round biserial throughout free arms (Fig. 5.7).
Oral surface with exposed ambulacra and presumably moveable ambulacral cover plates (which are not preserved), five primary peristomial cover plates (Fig. 5.1–5.3, 5.6), and a few large interradial plates. Anal opening on posterior side of oral surface.
Proximal stem circular, holomeric; other details unknown.
Etymology
After Albert Breimer for his outstanding work increasing the knowledge of Spanish Devonian crinoids.
Materials
Paratypes: MGM2996D, MPZ2018/516 (part and counterpart), and MPZ2018/517.
Measurements
MPZ2018/515: ACH, 3.6*; ACW, 7.2*; BH, 1.4; RH, 2.0*; RW, 3.1; AH, 13.5; CH, 2.7*.
Remarks
The specimen illustrated by Breimer (Reference Breimer1962, pl. 11, figs. 1–4) is considered conspecific with the new material from the Mariposas Formation, and the description above incorporates the morphology of both Breimer's specimen and new material. This new species is distinct with a relatively high basal circlet, a short rim is present around the periphery of the basal concavity, two circular nodes projecting outward on the radial plates, more than one small interradial plate in regular interrays, and free arms relatively slender. Species of Culicocrinus are compared in Table 3.
Table 3. Species diagnosis Table for Devonian species of Culicocrinus.

Camerata indeterminate
Figure 3.1–3.3
Occurrence
Mariposas Formation (Emsian), Iberian Chains, Spain.
Description
Calyx small in size; fixed rays protruding with interradial regions depressed. Calyx plate sculpturing smooth. Number and character of aboral cup circlets unknown. One interray exposed (but its orientation is not known), presumably two large interradial plates, arranged 1–1, in contact with tegmen. Tegmen with large plates and one high, central spine (Fig. 3.2).
First secundibrachial distal-most fixed brachial (Fig. 3.2). Free arms presumably 10 and atomous; brachials round chisel biserial; pinnules long, slender (Fig. 3.1).
Column unknown.
Material
This taxon is represented by a single, poorly preserved specimen (MPZ2018/518, part and counterpart).
Remarks
Although portions of this crinoid are well known, key morphological features, such as the presence or absence of the infrabasal circlet and plating in interradial areas, are not known. Consequently, this camerate crinoid cannot be recognized with certainty as either a monobathrid or diplobathrid camerate and is referred to here as Camerata Indeterminate. Although the central tegmen spine is similar to Acanthocrinus carlsi n. sp., the arms are different. The general size and shape, tegmen plating, and fixed rays suggest a monobathrid camerate, but the only Devonian crinoid from Spain and surrounding countries that bears any resemblance to this new specimen is Macaracrinus(?) sp. (Breimer, Reference Breimer1962), which is a diplobathrid. Further, Macaracrinus(?) sp. has a different tegmen, more fixed brachials, and more plates in interrays. The long, central tegmen spine and protruding fixed rays of Camerata Indeterminate are distinctive characteristics. These and other characters should provide a ready means by which to compare new material, which is needed for a positive identification.
Infraclass Inadunata Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Magnaorder Eucladida Wright, Reference Wright, Ausich, Cole, Peter and Rhenberg2017
Eucladid indeterminate
Figures 3.7, 4.2, 4.3
Occurrence
Mariposas Formation (Emsian), Iberian Chains, Spain.
Materials
MPZ2018/519.
Remarks
A single incomplete and partially disarticulated eucladid was collected from the Mariposas Formation (Fig. 3.7). Although unidentifiable, it has many distinctive characters. Despite the fact that the specimen is incomplete with plates somewhat askew, the following plates can be identified (Fig. 4.2, 4.3): C and D infrabasal plates; BC, CD, and DE basal plates; D and E radial plates; radianal; and first and second primibrachials in the D ray. In addition other aboral cup, column, and brachials and/or anal sac plates are present.
Key diagnostic characters that are preserved are infrabasal plates visible in lateral view, the radianal is interpreted to be beneath and to the left of the C radial plate, the radianal separates the C and D radial plates, the anal X is interpreted to be directly above the radianal and only partially in the aboral cup, radial facets are peneplenary, primibrachials are rectangular uniserial and broadly convex, the second primibrachial is axillary, and deep pits are present at the triple junction of aboral cup plates. This specimen is superficially similar to Costalocrinus ibericus Kammer, Reference Kammer2001 (especially see Breimer, Reference Breimer1962, pl. 15, figs. 9, 10) and Bactrocrinites robustus Pidal, Reference Pidal2008. However, it differs from C. ibericus in the posterior plate arrangement, as presently understood, and in the relative height and width of the primibrachials. It differs from B. robustus by having peneplenary radial facets and the relative height of the infrabasal circlet is unknown on the Mariposas specimen. Until specimens are recovered that reveal the aboral cup shape and arm branching, it is prudent to leave this taxon in open nomenclature.
Devonian Spanish crinoids in time and space
Crinoids are an important component of Devonian faunas in North Spain with isolated columnals a common component in many marine strata, articulated specimens are only present in certain horizons. In order to analyze the occurrence of complete crinoids in Spain, we have updated a list with all occurrences based on previous publications (see Appendix 1). This listing includes all previously known valid crinoid taxa based on preserved crowns (column taxa are excluded) known from Spain. In the present study, only Mariposas crinoids were critically evaluated for generic and specific assignments. A thorough taxonomic re-evaluation of all Devonian crinoids from Spain is needed.
In terms of stratigraphic comparisons, we used the stratigraphic scheme (Fig. 6) of García-Alcalde et al. (Reference García-Alcalde, Carls, Pardo Alonso, Sanz López, Soto, Truyols-Massoni, Valenzuela-Ríos, Gibbons and Moreno2002) that takes into account main areas where complete crinoids have been collected, especially in the Cantabrian Zone (Asturias and León) and the Iberian Chains (herein). Apart from these areas, complete crinoids are limited to the Lochkovian of the Pyrenees where complete crinoids have been described or figured from both Catalan (Haude, Reference Haude1992) and Aragonesse (Zamora, Reference Zamora and Zamora2018) regions.

Figure 6. Chronostratigraphic chart of Devonian formations cropping out in the Cantabrian Mountains (Asturias, León, Palencia) and Iberian Ranges (Montalbán, Tabuenca). Formations providing complete crinoids are marked as gray. After García-Alcalde et al. (Reference García-Alcalde, Carls, Pardo Alonso, Sanz López, Soto, Truyols-Massoni, Valenzuela-Ríos, Gibbons and Moreno2002).
As presently understood, ~124 crinoid taxa (based on crown material) have been named from the Devonian of Spain (Appendix 1). Excluding taxa left in open nomenclature, 93 species have been described. Of these taxa, 76% (71 species) are restricted to Spain. In contrast, only 18% (9 of 51) of the genera are restricted to Spain. Many of these taxa with wider geographic distributions are also known from southern Europe, and several are cosmopolitan, being known from Laurentia, across Gondwana, or in both regions. The new species described here are known exclusively from the Mariposas Formation from Spain (with the exception of C. breimeri n. sp., which is also known from the Cantabrian Zone), but all Mariposas genera are cosmopolitan.
The stratigraphically oldest complete crinoids from the Devonian in Asturias are from the Nieva Formation (Lochkovian-Pragian) where Hauser and Landeta (Reference Hauser and Landeta2007, Reference Hauser and Landeta2012) described a crinoid assemblage from Punta de Llampero (Asturias) including an indeterminate flexible (not figured), Botryocrinus sp., and Pisocrinus asturianus Hauser and Landeta, Reference Hauser and Landeta2012. Faunas equivalent in age to the Mariposas Formation described herein include a low diversity fauna from the lower Emsian that precedes the peak diversity reached during the upper Emsian of the Cantabrian Zone. Compared with Cantabrian crinoids, those from the Iberian Chains are from very shallow marine and turbid environments, and have been preserved in dark shales with few carbonate intercalations. Other important lower Emsian faunas are those from the La Ladrona, Aguión, and Moniello formations. The La Ladrona Formation (Emsian) contains complete crinoids in Asturias. They have been described in the papers of Hauser (Reference Hauser2010) and Hauser and Landeta (Reference Hauser and Landeta2012, Reference Hauser and Landeta2013) and include Bactrocrinites rauffi Hauser in Hauser and Landeta, Reference Hauser and Landeta2012, Diamenocrinus sp., Eutaxocrinus patulus Schmidt Reference Schmidt1942, Oehlerticrinus anguliferus (Whidborne, Reference Whidborne1897), Thylacocrinus sitter Hauser and Landeta, Reference Hauser and Landeta2013, and Zenkericrinus asturianus Hauser in Hauser and Landeta, Reference Hauser and Landeta2012. All of the species are known exclusively from Spain, as is Zenkericrinus, but the remainder of the genera are cosmopolitan.
The Aguión and Moniello formations in Asturias and Valporquero, Coladilla, from the upper part of La Vid Group, and Santa Lucia formations in León record some of the most diversified crinoid assemblages from the Spanish Devonian. The La Vid Group was deposited on a carbonate ramp, and its stratigraphic succession has been tied to two 3rd order transgressive-regressive cycles (Keller and Grötsch, Reference Keller and Grötsch1990; Keller, Reference Keller1997). The shales of the Valporquero Formation are thought to record the highstand of the upper 3rd order cycle, and they correspond to one of the richest units in crinoids, especially in the famous locality of Colle and surrounding areas (Breimer, Reference Breimer1962; Zamora et al., Reference Zamora, Álvaro, Arbizu, Colmenar, Esteve, Fernández-Marinez, Fernández, Gutiérrez-Marco, Andrés, Villas, Waters, Zamora and Rábano2015). The Aguión crinoid fauna consists of nine species, all of which are known only from Spain.
Crinoids continued to flourish and were abundant in the carbonate platforms from the Emsian Moniello and Santa Lucía formations (Breimer, Reference Breimer1962; Hauser and Landeta, Reference Hauser and Landeta2007, Reference Hauser and Landeta2009; Zamora et al., Reference Zamora, Álvaro, Arbizu, Colmenar, Esteve, Fernández-Marinez, Fernández, Gutiérrez-Marco, Andrés, Villas, Waters, Zamora and Rábano2015). The Moniello Formation, in the classic locality of Arnao (Asturias), also has a rich diversified crinoid fauna. This peak diversity in crinoids from the Upper Emsian is probably related to several factors that have not been explored in detail and probably include among others the local development of reefs (Arbizu et al., Reference Arbizu, Méndez-Bedia and Soto1995), microbial mounds (Fernández et al., Reference Fernández, Nose, Fernández-Martínez, Méndez-Bedia, Schröeder and Soto2006), and different substrata available for crinoid attachment (Zamora et al., Reference Zamora, Álvaro, Arbizu, Colmenar, Esteve, Fernández-Marinez, Fernández, Gutiérrez-Marco, Andrés, Villas, Waters, Zamora and Rábano2015). Taphonomic conditions are also favorable for crinoid preservation in this interval with the alternation of high storm-induced energetic events and calm conditions plus high sedimentary rates. The Santa Lucía Formation represents a carbonate shelf with a well-developed lagoon, a reef belt, and an off-reef facies (sensu Hofmann and Keller, Reference Hofmann and Keller2006), but distribution of crinoids within this model remains to be studied.
Unlike the bryozoans described by Suárez Andrés and Wyse Jackson (Reference Suárez Andrés and Wyse Jackson2017), the upper Emsian crinoid faunas are dominantly endemic to Spain. Only 27% (3 of 11) of the crinoid species from the Moniello Formation co-occur in Germany (e.g., Amblacrinus rosaceus [Roemer, Reference Roemer1844], Gasterocoma antiqua Goldfuss, Reference Goldfuss1839, and Halocrinus nodosus [Sandberger and Sandberger, Reference Sandberger and Sandberger1856]). None occurs beyond the Rhenish-Bohemian Province of the Old World Realm (see Boucot et al., Reference Boucot, Johnson, Talent and Oswald1967, Reference Boucot, Johnson and Talent1969; Suárez Andrés and Wyse Jackson, Reference Suárez Andrés and Wyse Jackson2017). Similarly, only 7% (2 of 27) of species in the Santa Lucia Formation occur outside Spain (three from Germany and one from Laurentia). In contrast, of the 23 genera in the Moniello and Santa Lucia formations, 17.4% are only known from Spain, 34.7% are restricted to the Rhenish-Bohemian Province, and 47.8% occur in multiple realms and are regarded as cosmopolitan.
The youngest Devonian crinoid occurrences in Spain are from the Givetian Candás and Portilla formations in Asturias and León respectively, and the Givetian–Frasnian Cardaño Formation in Palencia. In Frasnian levels, Breimer (Reference Breimer1962) reported a single crinoid, Lenneocrinus ventanillensis Breimer, Reference Breimer1962 from the Frasnian portion of the Cardaño Formation (see García Alcalde et al., Reference García-Alcalde, Montesinos, Truyóls-Massoni, García-López, Arbizu and Soto1988), and larger faunas are known from both the Candás and Portilla formations. As elsewhere in the Rhenish-Bohemian Province, the Cupressocrinidae are common and diverse. This well-known group has recently been revised by Bohatý (Reference Bohatý2005, Reference Bohatý2009) and Bohatý and Herbig (Reference Bohatý and Herbig2010). Givetian crinoid faunas of Spain are substantially more cosmopolitan than older faunas. At the species level, 50% (6 of 12) of Candás Formation species and 38% (3 of 8) of Portilla species are known only from Spain. However, 69% (9 of 13) of genera are cosmopolitan. One other genus is restricted to Spain, and two are known from only the Rhenish-Bohemian Province. Despite the lack of a comprehensive revision of Spanish Devonian crinoids, it is clear that as continental collisions proceeded during the Devonian, Spanish crinoid faunas became progressively more cosmopolitan.
Acknowledgments
This work was supported by the Spanish Ministry of Science, Innovation and Universities (grant number CGL2017-87631) and co-financed by the European Regional Development Fund. SZ was funded by a Ramón y Cajal Grant (RYC-2012-10576) and project “Aragosaurus: recursos geológicos y paleoambientales” (ref: E18_17R) funded by the Goberment of Aragón. We thank S. Menéndez (Museo Geominero, IGME, Spain) for assistance with curated specimens, and J. Colmenar and D. Rubio for their help collecting specimens. The volunteers from the Museo de Ciencias Naturales of the University of Zaragoza helped curating specimens collected from the Mariposas Formation. B. Lefebvre, G. Sevastopulo, and G.D. Webster improved an earlier version of this manuscript.
Appendix 1.
List of Devonian crinoids from Spain. Assignment of species to genera follow Webster and Webster (Reference Webster and Webster2014) and the authors that they cite. For publications after those included in Webster and Webster (Reference Webster and Webster2014), the generic assignments are listed as published, with the acknowledgement that some taxa may require further study. Taxa based exclusively on columnals and pluricolumnals are excluded.
FRASNIAN
Cardaño Formation; 1 km SW Ventanilla, near Cervera de Pisuerga (Palencia) (García-Alcalde et al., Reference García-Alcalde, Montesinos, Truyóls-Massoni, García-López, Arbizu and Soto1988)
Lenneocrinus ventanillensis Breimer, Reference Breimer1962
GIVETIAN
Candás Formation
Codiacrinus globisus Hauser in Hauser and Landeta, Reference Hauser and Landeta2012
Cupressocrinites crassus Goldfuss, Reference Goldfuss1831
Cupressocrinites hibrida Hauser in Hauser and Landeta, Reference Hauser and Landeta2012
Cupressocrinites hieroglyphicus (Schultze, Reference Schultze1866)
Espanocrinus elongatus Hauser in Hauser and Landeta, Reference Hauser and Landeta2013
Halocrinites schlotheimii Steininger, Reference Steininger1831
Halocrinites sp. aff. H. townsendi in (Koenig, Reference Koenig1825)
Haplocrinites boitardi Rouault, Reference Rouault1847
Haplocrinites mespiliformis (Goldfuss, Reference Goldfuss1831)
Megaradialocrinus elongatus (Goldfuss, Reference Goldfuss1839)
Megaradialocrinus? macrotatus (Austin and Austin, Reference Austin and Austin1845)
Megaradialocrinus tuberculatus (von Koenen, Reference Koenen1886)
Stylocrinus tabulatus depressus (Müller, Reference Müller, Zeiler and Wirtgen1855)
Vasocrinus aff. turbinatus Kirk, Reference Kirk1929
Portilla Formation
Ammonicrinus wanneri Springer, Reference Springer1926
Cupressocrinites spp. in Breimer, Reference Breimer1962
Dolatocrinus cantabricus Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Halocrinites inflatus (Schultze, Reference Schultze1866)
Halocrinites sampelayoi (Almela and Revilla, Reference Almela and Revilla1950)
Halocrinites townsendi (Koenig, Reference Koenig1825)
Halocrinites sp. aff. H. townsendi (Koenig, Reference Koenig1825)
Lemennocrinus truyolsi Pidal, Reference Pidal1984
Myelodactylus spp. in Breimer, Reference Breimer1962
Storthingocrinus cf. hauhi Oehlert, Reference Oehlert1897
Vasocrinus conicus Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Unspecified Formation
Cupressocrinus spp. in Oehlert, Reference Oehlert1902
EIFELIAN
Arnao Limestone
Pyxidocrinus? bifrons (Schmidt, Reference Schmidt1932)
cultijugatus Biozone
Platyhexacrinus kegeli Schmidt, Reference Schmidt1931
Platyhexacrinus? pisum Schmidt, Reference Schmidt1942
Storthingocrinus labiatus Schmidt, Reference Schmidt1932
Peña Negra Formation
Monstrocrinus aliformis Le Menn, Reference Le Menn1990
Unspecified Formation
Griphocrinus ovetensis Breimer, Reference Breimer1962
Halocrinites townsendi (König, Reference Koenig1825)
Orthocrinus planus Schmidt, Reference Schmidt1932
Orthocrinus sp. in Breimer, Reference Breimer1962
EMSIAN OR EMSIAN-EIFELIAN BOUNDARY
Santa Lucia Formation
Alejecrinus doyagueae Hauser and Landeta, Reference Hauser and Landeta2013
Babiacrinites costulatus Hauser in Hauser and Landeta, Reference Hauser and Landeta2009
Babiacrinites pyramidalis Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Bactrocrinites aff. depressus (Schultze, Reference Schultze1866)
Bactrocrinites onondagensis Goldring, Reference Goldring1954
Cantharocrinus melladoae Hauser and Landeta, Reference Hauser and Landeta2012
Cantharocrinus simplex Breimer, Reference Breimer1962
Corocrinus? grandosensis Breimer, Reference Breimer1962
Diamenocrinus jouani Oehlert, Reference Oehlert1891
Eifelocrinus cantabricus Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Espanocrinus barrandei Hauser in Hauser and Landeta, Reference Hauser and Landeta2012
Eutaxocrinus rhenanus (Roemer, Reference Roemer, Bronn and Geognostica1851)
Myelodactylus spp. in Breimer, Reference Breimer1962
Oenochoacrinus galeatus Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Oenochoacrinus pileatus Breimer, Reference Breimer1962
Orthocrinus elongatus Breimer, Reference Breimer1962
Orthocrinus sp. aff. primaevus in Le Menn, Reference Le Menn1980
Orthocrinus robustus Mellado in Arenal, Reference Arenal1950
Pithocrinus abbreviatus Hauser in Hauser and Landeta, Reference Hauser and Landeta2013
Pithocrinus ovatus Breimer, Reference Breimer1962
Pithocrinus aff. P. ovatus Breimer, Reference Breimer1962
Pithocrinus spinosus Breimer, Reference Breimer1962
Pithocrinus waliszewskii (Oehlert, Reference Oehlert1897)
Platyhexacrinus kegeli Schmidt, Reference Schmidt1932 in Hauser, Reference Hauser2015
Pyxidocrinus collensis Breimer, Reference Breimer1962
Sphaerocrinus wolfgangschmidti Hauser and Landeta, Reference Hauser and Landeta2007
Storthingocrinus aff. haughi Oehlert, Reference Oehlert1897
Vasocrinus breimeri Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Verneuilicrinus ibericus (Kammer, Reference Kammer2001)
Emsian-Eifelian Boundary Beds
Gasterocoma antiqua Goldfuss, Reference Goldfuss1839
Unspecified Formation
Gennaeocrinus sp. cf. G. nyssa (Hall, Reference Hall1862)
Macarocrinus? sp. in Breimer, Reference Breimer1962
EMSIAN
Aguión Formation
Espanocrinus lemonei Webster, Reference Webster1976
Gasterocoma xivaresensis Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Oenochoacrinus princeps Breimer, Reference Breimer1962
Periechocrinus baylii (de Verneuil, Reference Verneuil and de1850)
Pterinocrinus decembrachiatus Breimer, Reference Breimer1962
Stamnocrinus intrastigmatus (Schmidt, Reference Schmidt1932)
Stamnocrinus sp. 1 in Breimer, Reference Breimer1962
Vasocrinus fernandolandetai Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Verneuilicrinus landetai Hauser in Hauser and Landeta, Reference Hauser and Landeta2013
Coladilla Formation
Bactrocrinites robustus Pidal, Reference Pidal2008
La Vid Group
Bactrocrinites sp. in Breimer, Reference Breimer1962
Cantharocrinus minor Breimer, Reference Breimer1962
Eutaxocrinus collantesi Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Lasiocrinus? sp. in Breimer, Reference Breimer1962
Oenochoacrinus scaber Breimer, Reference Breimer1962
Pradocrinus baylii de Verneuil, Reference Verneuil and de1850
Pterinocrinus tenuibrachiatus Le Menn, Reference Le Menn1976
Pyxidocrinus collensis Breimer, Reference Breimer1962
Pyxidocrinus latus Breimer, Reference Breimer1962
Situlacrinus costatus Breimer, Reference Breimer1962
Trybliocrinus collensis Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Verneuilicrinus consolidatus (Schmidt, Reference Schmidt1942)
Verneuilicrinus ibericus (Kammer, Reference Kammer2001)
Verneuilicrinus thymos (Kammer, Reference Kammer2001)
La Ladrona Formation
Bactrocrinites rauffi Hauser in Hauser and Landeta, Reference Hauser and Landeta2012
Diamenocrinus sp. in Hauser and Landeta, Reference Hauser and Landeta2013
Eutaxocrinus patulus Schmidt Reference Schmidt1942
Oehlerticrinus anguliferus (Whidborne, Reference Whidborne1897)
Thylacocrinus sitteri Hauser and Landeta, Reference Hauser and Landeta2013
Zenkericrinus asturianus Hauser in Hauser and Landeta, Reference Hauser and Landeta2012
Mariposas Formation
Acanthocrinus carlsi n. sp. Ausich and Zamora, present study
Culicocrinus breimeri n. sp. Ausich and Zamora, present study
Platyhexacrinus santacruzensis n. sp. Ausich and Zamora, present study
Pterinocrinus tenuibrachiatus Le Menn, Reference Le Menn1976
Camerata Indeterminate in Ausich and Zamora, present study
Eucladida Indeterminate in Ausich and Zamora, present study
Moniello Formation (mostly Emsian)
Amblacrinus rosaceous (Roemer, Reference Roemer1844)
Cantharocrinus aff. minor in Breimer, Reference Breimer1962
Codiacrinus sp. aff. C. granulatus Schultze, Reference Schultze1866
Espanocrinus arnaoiensis Hauser, Reference Hauser2008
Eutaxocrinus asturianus Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Gasterocoma antiqua Goldfuss, Reference Goldfuss1839
Hallocrinus nodosus (Sandberger and Sandberger, Reference Sandberger and Sandberger1856)
Moniellocrinus alcaldei Pidal, Reference Pidal1984
Oenochoacrinus galeatus Hauser and Landeta, Reference Hauser and Landeta2007
Orthocrinus planus Schmidt, Reference Schmidt1932 in Hauser and Landeta, Reference Hauser and Landeta2009
Pithocrinus knorri Hauser and Landeta, Reference Hauser and Landeta2012
?Platyhexacrinus sp. in Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Robustocrinites scaber Schultze, Reference Schultze1866
Sphaerocrinus wolfgangschmidti Hauser in Hauser and Landeta, Reference Hauser and Landeta2007
Unspecified Formation
Codiacrinus spp. in Breimer, Reference Breimer1962
Diamenocrinus spp. in Breimer, Reference Breimer1962
Gennaeocrinus sp. cf. G. nyssa (Hall, Reference Hall1862)
Halocrinites sp. cf. C. schlotheimi (Steininger, Reference Steininger1831)
Hapalocrinidae cf. Culicocrinus nodosus (Müller, Reference Müller, Zeiler and Wirtgen1855)
Orthocrinus sp. (nov.?) in Breimer, Reference Breimer1962
Pterinocrinus diensti Schmidt, Reference Schmidt1934
PRAGIAN
Nieva Formation (Rañeces Group)
Botryocrinus llamperensis Hauser in Hauser and Landeta, Reference Hauser and Landeta2012
Pisocrinus asturianus Hauser and Landeta, Reference Hauser and Landeta2012
LOCKHOVIAN
?Late Silurian, Early Devonian, Pridoli–Lochkovian (probably Lochkovian).
Camarocrinus n. sp.? 1 in Haude, Reference Haude1992
Camarocrinus n. sp.? 2 in Haude, Reference Haude1992
UNKNOWN HORIZON IN SPAIN
Orthocrinus sp. cf. O. elongatus in Breimer, Reference Breimer1962
Pleurocrinus sp. (ex. gr. coplowensis Wright, Reference Wright1938) in Breimer, Reference Breimer1962 [Devonian or Mississippian]
Pyxidocrinus sanmigueli (Astre, Reference Astre1925) [glacial drift; Silurian or Devonian]
Pyxidocrinus sp. in Ruhrmann, Reference Ruhrmann1971
Scyphocrinus sp. ind. in Waagen and Jahn, Reference Waagen, Jahn and Barrande1899 [Pridoli or Lockhovian]
Sphaerocrinus sp. indet. in Schmidt, Reference Schmidt1942
Stamnocrinus sp. 2 in Breimer, Reference Breimer1962
Storthingocrinus haugi Oehlert, Reference Oehlert1897
Trybliocrinus flatheanus (Geinitz, Reference Geinitz1867)
Vasocrinus sp. cf. V. sculptus Lyon, Reference Lyon1857
Vasocrinus stellaris (Schultze, Reference Schultze1866)
Vasocrinus turbinatus Kirk, Reference Kirk1929
Vasocrinus aff. V. turbinatus Kirk, Reference Kirk1929









