



Introduction
The order Trigoniida arose in the late Silurian, maintaining a low diversity until the Permian when it experienced a radiation (Newell and Boyd, Reference Newell and Boyd1975). It was highly successful during Mesozoic times, when it was the most conspicuous and diverse group of shallow-burrowing bivalves (Stanley, Reference Stanley1977). It was considered a highly resilient taxon (Ros and Echevarría, Reference Ros Franch and Echevarría2011) that swiftly recovered after being strongly affected by the end-Permian and end-Triassic mass extinctions. On the other hand, after the end-Cretaceous extinction event, trigoniids were confined to epicontinental seas of Australia and New Guinea, where they are nowadays represented by a single genus (Darragh, Reference Darragh1986).
During most of the Early Jurassic the Trigoniida seems to have been largely restricted to the Pacific coast of the Americas and to Japan (Ziegler, Reference Ziegler1971; Aberhan, Reference Aberhan, Crame and Owen2002, appendices 1, 2; Francis and Hallam, Reference Francis and Hallam2003; Ros Franch et al., Reference Ros Franch, Márquez Aliaga and Damborenea2014, fig. 41; Echevarría and Ros Franch, Reference Echevarría and Ros Franch2019); although some authors mentioned the presence of lower Early Jurassic trigoniids elsewhere (e.g., Deecke, Reference Deecke1925; Hallam, Reference Hallam1977, appendix, p. 72). By the late Early Jurassic–early Middle Jurassic they experienced a wide dispersion, reaching the European Tethys (Francis, Reference Francis2000; Francis and Hallam, Reference Francis and Hallam2003) and South East Asian Tethys (Wandel, Reference Wandel1936; Hayami, Reference Hayami1972) by the Toarcian, the high latitudes of Antarctica by the Pliensbachian–Toarcian (Kelly, Reference Kelly1995), New Zealand by the Aalenian (Fleming, Reference Fleming1987), and eastern Africa by the Bajocian (Cox, Reference Cox1965). After the Triassic/Jurassic extinction event, the Trigoniida are well recorded and relatively abundant in South American Early Jurassic deposits (Leanza, Reference Leanza1942; Pérez and Reyes, Reference Pérez and Reyes1977; Ishikawa et al., Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983; Leanza, Reference Leanza1993; Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008). The present paper deals with this particularly interesting place and time-span for the study of Trigoniida systematics and evolution, and documents the rapid radiation of the group.
The systematics of the group has been variably treated, but a stable taxonomic framework has remained elusive. The genus Trigonia was first described by Bruguière (Reference Bruguière1789) and characterized mostly by its particular hinge; soon, several species were recognized within it and the genus was subdivided into sections (Agassiz, Reference Agassiz1840; Lycett, Reference Lycett1872–1879), which were the base for the recognition of many new genera (Bayle, Reference Bayle1878; van Hoepen, Reference van Hoepen1929; Crickmay, Reference Crickmay1930; Dietrich, Reference Dietrich1933). Cox (Reference Cox1952, see also Cox et al., Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969) differentiated the family Trigoniidae from the family Myophoriidae on the basis of hinge characters, particularly the broad and prominent median tooth of the left valve with transverse ridges on both occluding surfaces in the former. Later authors (Kobayashi, Reference Kobayashi1954; Saveliev, Reference Saveliev1958; Poulton, Reference Poulton1979; Leanza, Reference Leanza1993; Francis, Reference Francis2000) maintained that scheme of a single family, subdividing it into several subfamilies. Although Newell and Boyd (Reference Newell and Boyd1975) suggested a polyphyletic origin for trigoniid hinge characters, few authors followed their scheme. Boyd and Newell (Reference Boyd and Newell1997) themselves returned to the distinction between Myophoriidae and Trigoniidae.
Cooper (Reference Cooper1991) considered that, given the high variability within the Trigoniida, the systematic scheme was inadequate. He proposed two suborders within the order Trigoniida: Trigoniina and Myophorellina. The first one included the superfamilies Myophoriacea (equivalent to Myophoriidae sensu Cox, Reference Cox1952) and Trigoniacea. The second one included the superfamilies Myophorellacea and Megatrigoniacea. Many of the subfamilies were raised in rank to families.
Because the Early Jurassic stock is the basis for the main Mesozoic radiation of the group, the detailed study of the faunas here presented is fundamental in order to understand the evolution (and, as a consequence, the systematics) of the order. To infer evolutionary relationships among the studied species we employed a stratophenetic approach (Gingerich, Reference Gingerich, Cracraft and Eldredge1979) whenever possible, based mostly on a qualitative assessment of characters. We tried to maintain a phylogenetic criterion on supraspecific taxa, avoiding polyphyletic groups. In contrast, since ancestor-descendant relationships were inferred in some cases, paraphyly could not be avoided.
The systematics of Early Jurassic Trigoniida still needs to be supported by a phylogenetic analysis of the Triassic representatives (work in progress, but out of the scope of this contribution). The systematic arrangement followed here is thus tentative, and is based on Bieler et al. (Reference Bieler, Carter and Coan2010) and Carter et al. (Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011), with some modifications. We accept the superfamily Trigonioidea in the sense of Bieler et al. (Reference Bieler, Carter and Coan2010) and Carter et al. (Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011), including, among others, the families Groeberellidae, Trigoniidae, and Prosogyrotrigoniidae, and the family Myophoriidae as the ancestral stock for the others. The superfamily Myophorelloidea includes, in this report, the families Frenguelliellidae and Myophorellidae. We regard Frenguelliellidae as ancestral to Myophorellidae. Since we were not able to thoroughly analyze the later diversification of the suprageneric taxa here included, we refrained from providing proper diagnoses for them. The subfamily category sometimes has been used in excess in this group, resulting in lots of monogeneric subfamilies. Therefore, we prefer to avoid the use of subfamilies until further systematic revision is carried out.
The oldest paleontological papers describing Early Jurassic marine macrofossils from Argentina contain records of Trigoniida (Behrendsen, Reference Behrendsen1891; Burckhardt, Reference Burckhardt1902; Jaworski, Reference Jaworski1915), even though a few of the earliest described species were erroneously assigned stratigraphically to the Early Cretaceous (Jaworski, Reference Jaworski1915) or the Late Triassic (Groeber, Reference Groeber1924). Further descriptions were included in papers dealing with diverse invertebrate faunas (Weaver, Reference Weaver1931; Feruglio, Reference Feruglio1934; Carral-Tolosa, Reference Carral-Tolosa1942; Leanza, Reference Leanza1942), and later in papers specifically concerning the Trigoniida (Lambert, Reference Lambert1944; Levy, Reference Levy1966, Reference Levy1967; Leanza and Garate-Zubillaga, Reference Leanza, Garate-Zubillaga and Volkheimer1987; Leanza et al., Reference Leanza, Pérez and Reyes1987; Leanza, Reference Leanza1993). Early Jurassic Trigoniida from Chile were recently revised by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
We present here a comprehensive systematic analysis based on collections gathered during decades, by detailed sampling of a geographically extensive area across the entire Early Jurassic stratigraphy. Specimens from old collections made in Argentina are scattered in several European repositories; these specimens were examined whenever possible. The large amount of material available enabled us to pay particular attention to morphologic variability within many of the taxa described, an aspect somewhat neglected in the past, although its importance has been highlighted by some authors (e.g., Francis, Reference Francis2000; Schneider et al., Reference Schneider, Fürsich and Werner2011).
Geological setting
Marine Lower Jurassic beds in Argentina were widely deposited in the west-center of the country from southern San Juan to Chubut provinces (Fig. 1). Two main basins, probably interconnected, developed (i.e., Neuquén and Chubut). They both contain widespread deposits exhibiting a variety of paleoenvironments. The Aconcagua-Neuquén depocenter (Riccardi, Reference Riccardi, Moullade and Nairn1983) developed during the Late Triassic–Eocene (Howell et al., Reference Howell, Schwarz, Spalletti, Veiga, Veiga, Spalletti, Howell and Schwarz2005; Lanés, Reference Lanés2005) between 30° and 41°S latitude, with an eastward expansion towards the southeast (the Neuquén Embayment) that can be regarded as an epeiric sea. The onset of this basin is related to a rifting phase from the Middle Triassic to the Sinemurian (Ramos, Reference Ramos1992; Manceda and Figueroa, Reference Manceda, Figueroa, Tankard, Suárez Soruco and Welsink1995) that led to the evolution of a series of narrow and isolated depocenters (Uliana and Biddle, Reference Uliana and Biddle1988; Legarreta and Uliana, Reference Legarreta and Uliana1996). Marine sediments of this first stage crop out only in the Atuel River region (southern Mendoza) (Riccardi et al., Reference Riccardi, Damborenea, Manceñido and Ballent1988), close to the so-called Curepto Strait, which was one of the main connections of the basin with the Paleopacific (Vicente, Reference Vicente2005). After that, a sag stage developed from late early Sinemurian to Toarcian, causing the coalescence of the initial depocenters during the late Sinemurian–Pliensbachian, and thus enlargement of the area under marine influence (Legarreta and Gulisano, Reference Legarreta, Gulisano, Chebli and Spalletti1989; Legarreta and Uliana, Reference Legarreta and Uliana1996). From this time on, the Neuquén Basin developed as a back-arc basin related to circum-Pacific convergence (Legarreta and Uliana, Reference Legarreta and Uliana1996), with pyroclastic and volcanic content especially common in Lower Jurassic deposits. During the early Pliensbachian the basin expanded southward, reaching its maximum extension in the late Pliensbachian. Although the basin was partially barred from the open ocean, the well-diversified faunas indicate that free oceanic connections occurred through gaps in the arc (Legarreta and Uliana, Reference Legarreta and Uliana1996). By Middle Jurassic times, proximal deltaic and alluvial facies started to prograde, reducing the depositional area (Legarreta and Uliana, Reference Legarreta, Uliana and Caminos1999, p. 405).
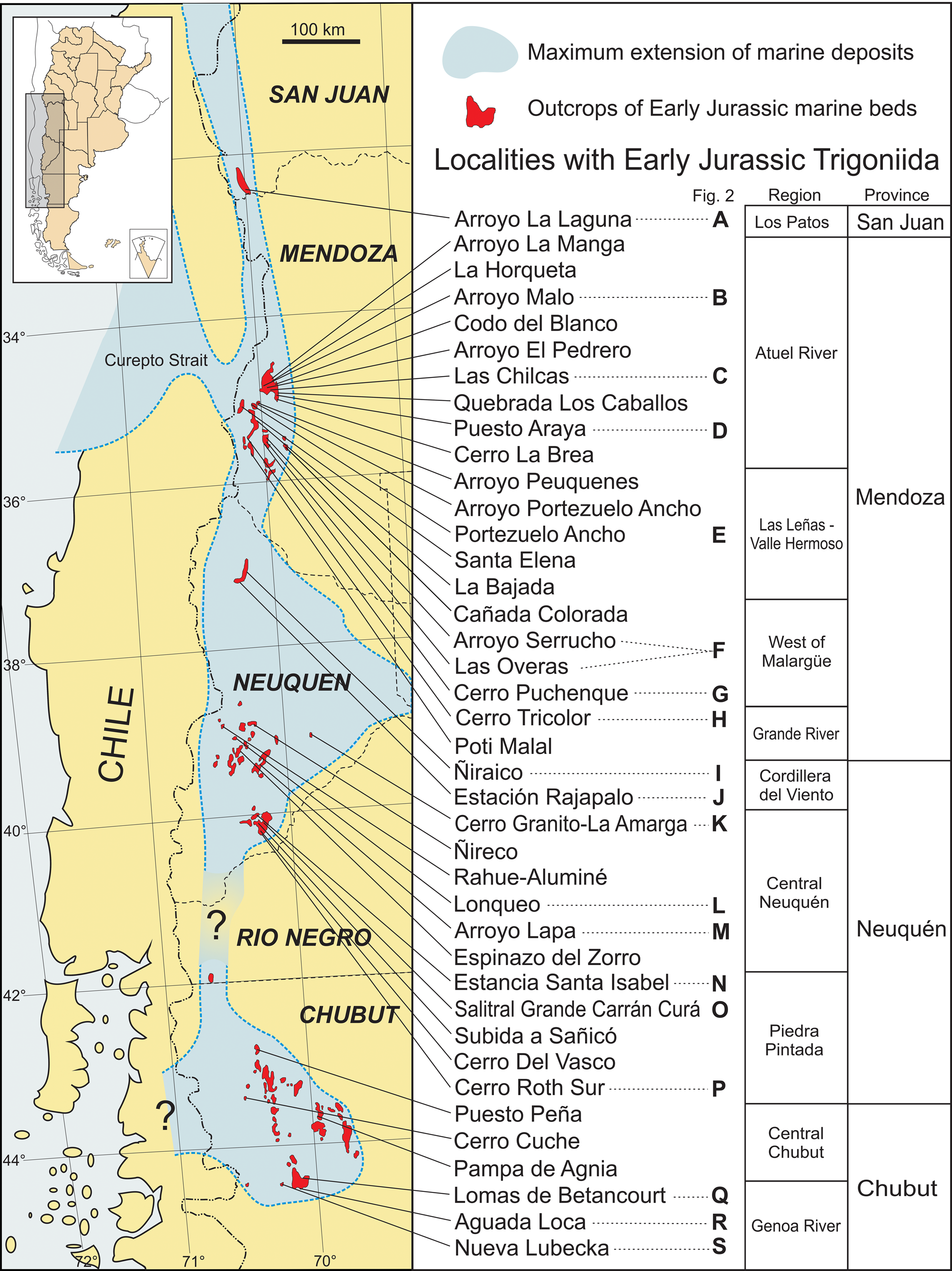
Figure 1. Map of the localities mentioned in the text, in the context of general Pliensbachian–Toarcian paleobiogeography, compiled from various sources, mainly Legarreta and Uliana (Reference Legarreta and Uliana1996), and Vicente (Reference Vicente2005). Localities are grouped by region and province as used in the text. Stratigraphical sections depicted in Figure 2 are indicated by capital letters.
The Chubut Basin is a NNW-SSE elongated depocenter (Fig. 1) with marine and continental sedimentary deposits of Early Jurassic age, mainly exposed in the western region of Chubut Province and extending northwards into Río Negro Province (41°00’–46°30′S) (Suárez and Márquez, Reference Suárez and Márquez2007). The sedimentary succession of the Chubut Basin accumulated under an extensional tectonic regime (Lizuain, Reference Lizuain and Caminos1999; Uliana and Legarreta, Reference Uliana, Legarreta and Caminos1999), starting with continental deposits overlain by shallow-marine and continental successions. To the east, these marine beds interfinger with mainly pyroclastic continental facies (Franchi et al., Reference Franchi, Panza, De Barrio, Chebli and Spalletti1989). Early Jurassic marine sediments lie unconformably over late Paleozoic rocks, but contrary to Carral-Tolosa's (Reference Carral-Tolosa1942, p. 22, 67) assumption of a complete Early Jurassic marine sequence in Chubut, most marine beds seem to have been deposited during the short interval from the late Pliensbachian to the early Toarcian (Riccardi Reference Riccardi2008a, Reference Riccardib).
Thus, by late Pliensbachian times, there was an elongate marine encroachment with two expanded basins (Neuquén and Chubut) that were connected with the Pacific Ocean (Legarreta and Uliana, Reference Legarreta and Uliana1996). Their sedimentary successions maintained local variations in thickness and lithology due to an uneven basin floor related to active rifting and a variety of depositional environments (Legarreta and Uliana, Reference Legarreta, Uliana and Caminos1999). The basinal successions are punctuated by turbiditic strata and by shallower water deposits (lowstand wedges). Syntheses of paleogeographic reconstructions for different time intervals were published by Legarreta and Uliana (Reference Legarreta and Uliana1996, fig. 9, Reference Legarreta, Uliana and Caminos1999, figs. 5–7), Vicente (Reference Vicente2005, Reference Vicente2012), and Arregui et al. (Reference Arregui, Carbone, Martínez, Leanza, Arregui, Carbone, Danieli and Vallés2011, figs. 5, 6), among others.
Ammonite faunas are abundant and diverse in both the Neuquén and Chubut basins. A detailed local biostratigraphy, which is well correlated to the standard zonation, was developed by Riccardi (Reference Riccardi2008a, Reference Riccardib) and Riccardi et al. (Reference Riccardi, Damborenea, Manceñido, Leanza, Leanza, Arregui, Carbone, Danieli and Vallés2011). This is the age framework used as reference in this paper (as shown in Figs. 2, 3).

Figure 2. Selected sections arranged from north to south, with the distribution in space and time of the genus-group taxa treated in this paper. Biostratigraphic framework from Riccardi (Reference Riccardi2008a, Reference Riccardib) and Riccardi et al. (Reference Riccardi, Damborenea, Manceñido, Leanza, Leanza, Arregui, Carbone, Danieli and Vallés2011). All sections sketched to the same scale and leveled to the base of the Toarcian.

Figure 3. Stratigraphic distribution in Argentina of species described. Local ammonite biozonation and its equivalence to the standard zonation from Riccardi (Reference Riccardi2008a, Reference Riccardib).
Materials and methods
Stratigraphic sections were logged at most of the localities listed in Figure 1, but only the most relevant to this study are sketched in Figure 2. The entire fauna was recorded and/or sampled at each fossiliferous bed, but only those horizons containing Trigoniida are indicated in the figure. A synthesis of the stratigraphic range for each species is depicted on Figure 3.
Terminology
We follow Carter et al. (Reference Carter, Harries, Malchus, Sartori, Anderson, Bieler and Bogan2012) for general morphologic terminology (Fig. 4). Accordingly, we use “costae” or “ribs” for shell surface ornamentation structures that are not expressed in the interior of the shell, and “folds” for undulations that affect the entire thickness of the shell. Some of the terms used here need explanation. For instance, the marginal and escutcheon carinae are typical characters of the group, to the point that sometimes they have been described even when the shell lacks an actual carina. Because these differences may bear importance in the phylogenetic study of the group, we distinguish and apply the following terms: (1) “carina” when a protruding elevation is developed (Fig. 4.3), sometimes (though not always) as a fold in the shell; (2) “angulation” (sensu Cooper, Reference Cooper1989) when there is a sharp change in orientation between the area and the other surfaces (Fig. 4.4), developing a linear structure equivalent in position to the carina, but not a carina strictly speaking; and (3) “bend” (see Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 66 under “Description” for Prosogyrotrigonia tenuis Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) for a gentle change in orientation between the area and the other surfaces (Fig. 4.5). The term “stepped angulation” is used when the boundary between the area and the escutcheon is an angulation, but because the escutcheon is depressed, a small step between both surfaces is developed (Fig. 4.4). Similar to the carinae, an antecarinal sulcus occasionally has been described, even when there is no actual depression. Thus, the term “antecarinal sulcus” is restricted to a true depression anterior to the marginal carina, angulation, or bend (Fig. 4.2), and “antecarinal space” is used for flat surfaces (leveled with the flank) showing an abrupt difference in ornamentation relative to the flank (Fig. 4.1).

Figure 4. Morphology and terminology. (1, 2) Main descriptive terminology for trigoniid shell features; (3–5) transverse sections of valve, showing different types of contact between escutcheon (es) and area (a), and between area and flank (fl): (3) prominent escutcheon and marginal carinae, (4) stepped escutcheon angulation and marginal angulation, (5) escutcheon and marginal bends; (6, 7) trigonian-grade hinge in left and right valves, showing hinge notation, m.b. = myophoric buttress.
In addition to the usual kinds of costae orientation in shell ornamentation (e.g., radial, commarginal, and oblique), the term “sub-commarginal” is here used to refer to flank costae where the costal segment on the central part of the flank is commarginal, but on the anterior part, it cuts across growth lines and meets the anterior margin at high angles (see Poulton, Reference Poulton1979, p. 12, text-fig. 4, characterized as “pseudoconcentric”; Fig. 4.1).
Some genera may develop an “internal radial ridge” (sensu Cox, Reference Cox1952, p. 57–58, under Prorotrigonia, and p. 59–60, under Pterotrigonia) on the posterior part of the inner surface of the shell, which is approximately coincident with the midline of the area. According to Gould and Jones (Reference Gould and Jones1974), this internal radial ridge may have helped these bivalves separate inhalant from exhalant currents.
Dentition notation is the most generally used for the group (Fig. 4.6, 4.7). Trigonian-grade hinge has been characterized by the presence of a prominent, subtriangular, and extremely broad median tooth on the left valve (tooth 2 on Fig. 4.6), with a concave ventral surface giving it a bifid aspect, and with anterior and posterior faces bearing strong transverse ridges or striae (Cox, Reference Cox1952; Newell and Boyd, Reference Newell and Boyd1975; Poulton, Reference Poulton1979; Boyd and Newell, Reference Boyd and Newell1997). The two main teeth of the right valve (teeth 3a and 3b; Fig. 4.7) are widely divergent and more or less symmetrically disposed, also with transverse ridges (Cox, Reference Cox1952; Newell and Boyd, Reference Newell and Boyd1975; Poulton, Reference Poulton1979). Other characters usually mentioned are a gap or hiatus in the right valve hinge plate and the presence of a myophoric buttress (“m.b.” on Fig. 4.6, 4.7) posterior to the anterior adductor muscle scar. This myophoric buttress is fused with tooth 3a in the right valve and with the hinge plate, which forms the socket for this tooth in the left valve (Cox, Reference Cox1952; Fleming, Reference Fleming1964; Newell and Boyd, Reference Newell and Boyd1975).
In contrast, the myophorian-grade hinge (see Newell and Boyd, Reference Newell and Boyd1975, fig. 12.C, 12.D) has been characterized as having a simple (Cox, Reference Cox1952) or bifid tooth 2, which is not particularly prominent (Cox, Reference Cox1952; Fleming, Reference Fleming1964; Newell and Boyd, Reference Newell and Boyd1975). Teeth 3a and 3b are more unequally and asymmetrically arranged than in the trigonian-grade hinge, and are placed on a hinge plate (Cox, Reference Cox1952; Fleming, Reference Fleming1964; although Newell and Boyd, Reference Newell and Boyd1975, mentioned that some species may have a hiatus). Dental striations are variably developed in the myophorian-grade hinge, although the striation is less conspicuous than that of the trigonian-grade hinge (Cox, Reference Cox1952; Newell and Boyd, Reference Newell and Boyd1975). Newell and Boyd (Reference Newell and Boyd1975) and Boyd and Newell (Reference Boyd and Newell1997) also mentioned, as a distinctive character of myophorian-grade hinge, the elongation of the posterior limb of the tooth 2, which functioned as a second inner pseudolateral tooth.
Although the trigonian-grade hinge definition provided above fits well for the hinges of most, if not all, post-Triassic Trigoniida, that is not the case for Triassic representatives, where the different elements may be variably combined (e.g., Boyd and Newell, Reference Boyd and Newell1997; Hautmann, Reference Hautmann2003). This mosaic combination of characters may be indicative of a polyphyletic origin for the trigonian-grade hinge, as suggested by Newell and Boyd (Reference Newell and Boyd1975). Although the subject is beyond the scope of this study, when discussing some Triassic representatives of the group, we follow Boyd and Newell (Reference Boyd and Newell1997) and consider that the minimum requirements for a hinge to be defined as trigonian-grade are: (1) to have well-developed striation on both surfaces of teeth 2 and 3a (and on the corresponding sockets), and (2) the absence of the elongate posterior limb on tooth 2.
Shell size is characterized by length according to the following scheme: length <20 mm = very small shell; 20 to <40 mm = small shell; 40 to <60 mm = medium-sized shell; 60 to <80 mm = large shell; length ≥80 mm = very large shell. Measurements are given in mm, and the following abbreviations are used: L = length, H = height, W = width of both valves, V = width of a single valve.
Repositories and institutional abbreviations
Specimens collected by the authors are housed in the following repositories: IANIGLA-PI = Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Mendoza, Argentina; MCF-PIPH = Museo Municipal Carmen Funes, Plaza Huincul, Argentina; MLP = División Paleontología Invertebrados, Museo de Ciencias Naturales de La Plata, La Plata, Argentina; MOZ-PI = Museo Provincial de Ciencias Naturales “Dr. Prof. Juan A. Olsacher”, Zapala, Argentina; and MPEF-PI, Museo Paleontológico Egidio Feruglio, Chubut, Argentina. Further specimens examined are housed in: BMNH = Burke Museum of Natural History, Seattle, USA; CPBA = Geology Department, Universidad de Buenos Aires, Buenos Aires, Argentina; SIRAME-SEGEMAR and DNGM = Dirección Nacional de Geología y Minería, Buenos Aires, Argentina; GSC = Geological Survey of Canada, Calgary, Canada; IGPB = Institute of Geosciences, Paleontology Section, Bonn, Germany; NHMB = Naturhistorisches Museum Basel, Basel, Switzerland; SNGM = Servicio Nacional de Geología y Minería, Santiago, Chile; MNHN = Muséum Nationale d’ Histoire Naturelle, Paris, France. A list of Argentinian Trigoniida specimens included in this study by species, locality, repository abbreviation, and local zonation is provided in Appendix 1.
The synonymy lists were prepared according to Matthews (Reference Matthews1973) to indicate the degree of confidence in allocation of each entry. They include only published records.
Systematic paleontology
Order Trigoniida Dall, Reference Dall1889
Superfamily Trigonioidea Lamarck, Reference Lamarck1819
Remarks
According to Bieler et al. (Reference Bieler, Carter and Coan2010) and Carter et al. (Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011), this superfamily includes members representing the three hinge grades (schizodian-grade, myophorian-grade, and trigonian-grade). Of interest for this report are the families Groeberellidae, Trigoniidae, and Prosogyrotrigoniidae, all with a trigonian-grade hinge, and the family Myophoriidae, with a myophorian-grade hinge and without any Jurassic representatives, but probably ancestral to the other three.
Family Groeberellidae Pérez, Reyes, and Damborenea, Reference Pérez, Reyes and Damborenea1995
Remarks
The family Groeberellidae was characterized by Pérez et al. (Reference Pérez, Reyes and Damborenea1995) as having a myophoriform shell (i.e., subquadrate, inflated, orthogyrate to slightly prosogyrate), trigonian-grade hinge and a flank ornamentation dominated by radial costae. They inferred an origin from myophorid ancestors independent from that of other trigoniids.
General shell morphology in Groeberella corresponds well to that of some Triassic species with myophorian-grade hinges, and for this reason Groeber (Reference Groeber1924) assigned a Triassic age to the beds bearing the type material. Costatoria, for example, is characterized by the presence of well-developed radial costation, in some species with wide concave interspaces (see Hautmann, Reference Hautmann2001, figs. 26.10, 26.11, 27.1–27.11 for some examples). Although being most abundant in the Tethys and in Japan (Ros Franch et al., Reference Ros Franch, Márquez Aliaga and Damborenea2014, p.120, fig. 42), there are also a few mentions from the American Pacific margin (Chong and Hillebrandt, Reference Chong and von Hillebrandt1985, Costatoria sp., Norian or Rhaetian from Quebrada San Juan, Antofagasta Region, North Chile; Damborenea and González-León, Reference Damborenea and González-León1997, Costatoria? sp., Upper Triassic, probably Norian, of Sierra del Álamo, Sonora, Mexico). The record from Mexico shows poorly developed striation on hinge teeth not covering the entire occluding surfaces, but, as in Groeberella, striation does not develop on the posterior face of 3b (Damborenea and González-León, Reference Damborenea and González-León1997, p. 192, fig. 8.3, 8.4).
In contrast, some authors (e.g., Levy, Reference Levy1967; Leanza, Reference Leanza1993) related Groeberella to the Minetrigoniinae, with a trigonian-grade hinge, flank sculpture of intersecting radial and commarginal costae, and trellised ornament on the area (Kobayashi, Reference Kobayashi1954; Fleming, Reference Fleming1987).
Genus Groeberella Leanza, Reference Leanza1993
Type species
Myophoria neuquensis Groeber, Reference Groeber1924, Pliensbachian, Neuquén, Argentina.
Diagnosis
Subquadrangular, inequivalve, palmate shell. Orthogyrate to slightly prosogyrate umbos. Prominent escutcheon and marginal carinae, and few widely spaced radial costae on the flank. Intercostal (and intercarinal) spaces concave. Trigonian-grade hinge.
Remarks
The diagnosis of the genus is adapted from Pérez et al. (Reference Pérez, Reyes and Damborenea1995). Besides the type species, early Sinemurian material from Chile was referred to Groeberella sp. (Pérez et al., Reference Pérez, Reyes and Damborenea1995). Also, an early Pliensbachian left valve from Mexico was described as Groeberella sp. A (Scholz et al., Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008), and a fragmentary specimen from Neuquén referred to the Bajocian was likewise left in open nomenclature as Groeberella sp. by Leanza (Reference Leanza1993, p. 19).
Groeberella neuquensis (Groeber, Reference Groeber1924)
Figure 5.1–5.8
- v* Reference Groeber1924
Myophoria neuquensis Groeber, p. 92, pl. 1, figs. a, b.
- v Reference Windhausen1931
Myophoria neuquensis; Windhausen, p. 179, fig. 71. [reproduced from Groeber, Reference Groeber1924]
- v Reference Carral-Tolosa1942
Myophoria neuquensis; Carral-Tolosa, p. 59, pl. 6, figs. 3a–c.
- v Reference Levy1967
Myophorigonia neuquensis; Levy, p. 14, figs. 1a–f.
- Reference von Hillebrandt and Schmidt-Effing1981
Myophorigonia neuquensis; Hillebrandt and Schmidt-Effing, p. 10.
- Reference Pérez1982
Myophorigonia aff. M. neuquensis; Pérez, p. 40, pl. 15, figs. 2–5, appendix 1.
- v Reference Damborenea and Manceñido1992
‘Myophorigonia’ neuquensis; Damborenea and Manceñido, p. 134, pl. 1, fig. 5a.
- v Reference Damborenea and Manceñido1992
Myophorigonia neuquensis; Damborenea et al., pl. 115, fig. 14.
- Reference Leanza1993
Groeberella neuquensis; Leanza, p. 18, pl. 1, figs. 2, 3.
- Reference Pérez, Reyes and Damborenea1995
Groeberella neuquensis; Pérez et al., p. 147, pl. 1, figs. 1–3, 5–13, 15, 16, 18–22.
- Reference Pérez and Reyes1997
Groeberella neuquensis; Pérez and Reyes, p. 573.
- v Reference Pérez and Reyes2008
Groeberella neuquensis; Pérez and Reyes in Pérez et al., p. 58, pl. 1, figs. 1–13, 15, 17–18.
- v Reference Riccardi, Damborenea, Manceñido, Leanza, Leanza, Arregui, Carbone, Danieli and Vallés2011
Groeberella neuquensis; Riccardi et al., fig. 7.3.
Type materials
Holotype: DNGM 7337 (= casts SNGM 7366, MLP 24324); right valve from the Pliensbachian of Puruvé-Pehuén, Neuquén Province, Argentina. Figured by Groeber (Reference Groeber1924, pl. 1, figs. a, b), Damborenea and Manceñido (Reference Damborenea and Manceñido1992, pl. 1, fig. 5a), Damborenea et al. (Reference Damborenea, Polubotko, Sey, Paraketsov and Westermann1992, pl. 115, fig. 14), Pérez et al. (Reference Pérez, Reyes and Damborenea1995, pl. 1, fig. 1), and Figure 5.1–5.3 herein.
Occurrence
The type locality is Puruvé-Pehuén (somewhere between Ñireco and Lonqueo in central Neuquén). Also recorded from: Las Chilcas, Puesto Araya, Quebrada de los Caballos (Atuel River region); La Bajada, Portezuelo Ancho (Las Leñas/Valle Hermoso region); Estancia Santa Isabel, Salitral Grande Carrán Curá, Cerro Roth Sur (Piedra Pintada region); and Nueva Lubecka (Genoa River region). Leanza (Reference Leanza1993, p. 10) mentioned the presence of this species at La Amarga, near Rincón del Águila (central Neuquén), but he listed neither this locality when describing the species (p. 19) nor this species among the fauna from that locality (p. 68). In Argentina, its stratigraphical range is late Sinemurian to late Pliensbachian (Fig. 3). This species seems to have had a wider stratigraphical range in Chile, extending at least to early Aalenian according to Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 55).
Description
Medium-sized, subquadrangular, orthogyrate to slightly prosogyrate shell. Umbos anteriorly located, slightly displaced from each other (Fig. 5.8). Escutcheon margin feebly convex; posterior margin slightly concave. Ventral margin palmate due to the presence of two radial ribs (Fig. 5.4). Anterior margin slightly convex. Escutcheon wide, smooth or with growth lines; escutcheon carina prominent. Area smooth (or with growth lines), slightly concave (Fig. 5.1, 5.4); prominent marginal carina. Area and flank meet at an angle of ~90°. Flank with two prominent radial ribs; spaces between the ribs (and marginal carina) wide and concave. Flank ribs and carinae as folds of the shell, shell thicker on the costae than on the interspaces. Costae and carinae on the left valve with transverse crenulations (Fig. 5.4, 5.5); intercostal spaces on the flank with thin commarginal costellae, more densely arranged than the crenulations. Costae sharper on the right valve (Fig. 5.1–5.3), lacking transverse ornamentation.
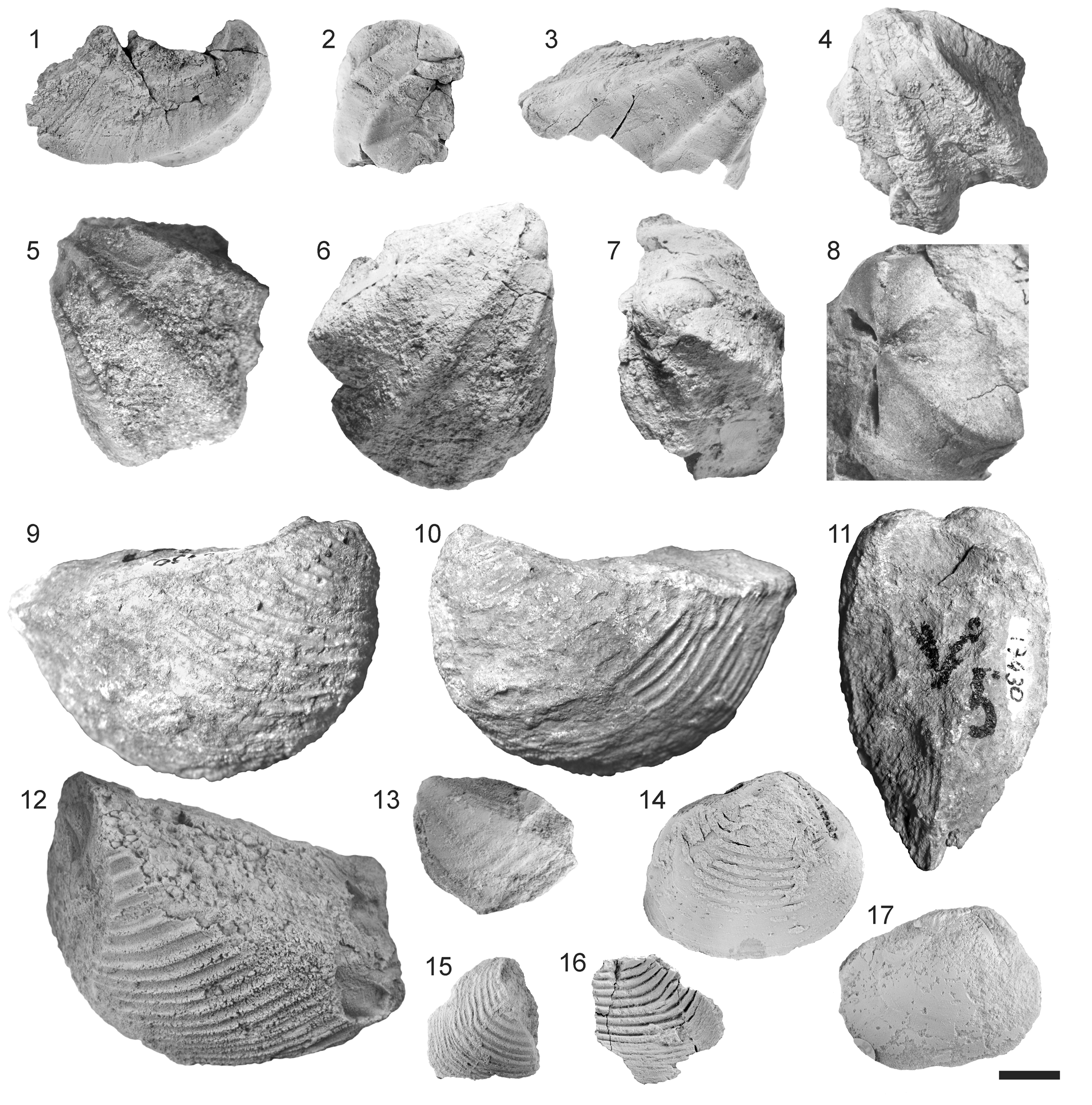
Figure 5. Trigoniida from the Early Jurassic of Argentina. Scale bar = 10 mm. (1–8) Groeberella neuquensis (Groeber): (1–3) holotype, DNGM 7337, right valve, dorsal, anterior and right lateral views, Pliensbachian, Puruvé-Pehuén; (4) DNGM 7340a, left valve, lateral view, Pliensbachian, Nueva Lubecka; (5) MPEF-PI 6401, left valve, Pliensbachian, Nueva Lubecka; (6, 7) DNGM 7339, right valve, lateral and dorsal views, Pliensbachian, Nueva Lubecka; (8) MLP 27844, internal molds of both valves open in butterfly position, early Pliensbachian, Puesto Araya; (9–11) Trigonia? sp. 2, CPBA 17430, right lateral, left lateral and dorsal views, early Toarcian, Arroyo La Laguna; (12, 13) Trigonia? sp. 1, early Toarcian, Arroyo La Laguna: (12) MLP 36312a, left valve; (13) MLP 36311, juvenile? left valve; (14–17) Prosogyrotrigonia tenuis Pérez and Reyes, early Sinemurian, Arroyo Malo: (14) MLP 32789, composite mold with shell remains, right lateral view; (15) MLP 32802, postero-ventral fragment of right valve composite mold; (16) MLP 32828, postero-ventral fragment of right valve external mold; (17) MLP 32805, internal mold, right lateral view.
Trigonian-grade hinge (sensu Boyd and Newell, Reference Boyd and Newell1997). Tooth 2 broad and conspicuous, ventrally bifid; teeth 4a and 4b relatively short. Tooth 3a thicker than tooth 3b. Few strong ridges on both occluding surfaces of 2 and 3a, on the anterior face of 3b, and on the posterior face of 4a. Nymph short.
Materials
Thirty-five specimens were examined: the holotype and DNGM 7338–7340, MLP 16387, 17478, 17724, 27246–27253, 27844, 27931, 28683, 36194; MPEF-PI 2913, 2918, 2993, 3392, 6397, 6435; IANIGLA-PI 3344; plus the specimen MOZ-PI 4060 figured by Leanza (Reference Leanza1993).
Measurements
Holotype (DNGM 7337): L = 42 mm (broken ventrally); DNGM 7339: L = 47 mm, H = 44 mm; DNGM 7340: H = 47 mm.
Remarks
The species is endemic to South America (Damborenea et al., Reference Damborenea, Echevarría and Ros-Franch2013).
Family Trigoniidae Lamarck, Reference Lamarck1819
Remarks
The Trigoniidae were described by Cooper (Reference Cooper1991) as having a trigonal to rhomboidal shell shape, a trigonian-grade hinge, subcommarginal flank ornamentation, a prominent marginal carina, and an area with radial ornamentation (at least on early stages). Some previous authors (e.g., Kobayashi, Reference Kobayashi1954; Poulton, Reference Poulton1979), provided a similar characterization for the subfamily Trigoniinae.
The earliest representatives of this family are included in the genus Primatrigonia (defined as a subgenus of Trigonia by Repin in Paevskaya et al., Reference Paevskaya, Polubotko, Repin, Rozanov and Shevyrev2001; see Echevarría et al., Reference Echevarría, Ros Franch and Manceñido2018 for a nomenclatural revision of this taxon). All the species included in Primatrigonia share a subdued ornamentation pattern, with weak radial costellae on the area and very subtle commarginal costellae or prominent growth lines on the flank (nearly smooth in some cases). Trigonia tabacoensis Barthel, Reference Barthel1958 (see also Pérez and Reyes, Reference Pérez and Reyes2008, fig. 1) from the Anisian of Chile, and Trigonia n. sp. A of Fleming (Reference Fleming1987), from the Anisian of New Zealand, may well be included in Primatrigonia. The Chilean species is the oldest within the order with a recognizable trigonian-grade hinge. Other Late Triassic species are Primatrigonia yunnanensis (Guo, Reference Guo1985), from southwestern China, Primatrigonia zlambachiensis (Haas, Reference Haas1909), from Austria, Iran, and Vietnam (Hautmann, Reference Hautmann2001), and Primatrigonia gaytani (von Klipstein, Reference von Klipstein1843) from Austria. The hinge shows some variability among these species (Hautmann, Reference Hautmann2001, p. 120–122). The genus Trigonia probably evolved from Primatrigonia by the development of a stronger ornamentation pattern.
Genus Trigonia Bruguière, Reference Bruguière1789
Type species
Venus sulcata Hermann, Reference Hermann1781, pl. 4, figs. 2–4, 9, 10 (see ICZN, 1955), late Early Jurassic (Toarcian), Gundershoffen, Alsace, France.
Diagnosis
Shell trigonal to trigonally ovate, inequilateral, opisthogyrate. Area bipartite, with reticulate ornament, composed of radial and commarginal costellae. Prominent marginal carina; antecarinal sulcus present on the left valve. Flank with sub-commarginal costae. Escutcheon smooth or with striae.
Remarks
The diagnosis of the genus was compiled from Crickmay (Reference Crickmay1932), Leanza (Reference Leanza1993), and Francis (Reference Francis2000). If Primatrigonia is regarded as a different genus, then the oldest Trigonia representative is the species Trigonia senex Kobayashi and Mori, Reference Kobayashi and Mori1954, from the Hettangian of Japan. During most of the Early Jurassic, the genus occurred in the Pacific (Kobayashi and Mori, Reference Kobayashi and Mori1954; Poulton, Reference Poulton1976, Reference Poulton1979, Ishikawa et al., Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983; Pérez and Reyes, Reference Pérez and Reyes1991; Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008). By the Toarcian it reached the European Tethys (Hermann, Reference Hermann1781; Agassiz, Reference Agassiz1840; Fürsich et al., Reference Fürsich, Berndt, Scheuer and Gahr2001, Francis and Hallam, Reference Francis and Hallam2003).
Trigonia? sp. 1
Figure 5.12, 5.13
Occurrence
Arroyo La Laguna (Los Patos region), San Juan Province. Early Toarcian (D. hoelderi Biozone [≈ Serpentinum Biozone]) (Fig. 3).
Description
Medium-sized, slightly opisthogyrate shell. Escutcheon poorly preserved, apparently wide. Area wide, ~1/3 of shell surface (Fig. 5.12), with mid radial stepped angulation and thin radial costellae intersected by thin commarginal costellae (Fig. 5.12, 5.13). Marginal carina poorly preserved, prominent (at least in juvenile stages). Flank with sharp sub-commarginal costae, more densely arranged as shell grows.
Materials
Three left valves, MLP 36311, 36312a, 36313.
Measurements
MLP 36311, composite mold: L = 25 mm, H = 24 mm; MLP 36313, composite mold: L = 28 mm, H = 23 mm; MLP 36312a, single valve: L = 55 mm, H = 45 mm, V = 18 mm.
Remarks
Poor preservation of the material prevents a specific taxonomic assignment. Neuquenitrigonia Leanza and Garate-Zubillaga, Reference Leanza, Garate-Zubillaga and Volkheimer1987 was distinguished from Trigonia mainly due to flank ornamentation (oblique to growth lines), but Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) later included material with sub-commarginal flank costae. Unfortunately, they did not discuss the diagnosis for the genus. Yet, based on the material they included, the main difference from Trigonia seems to be the transverse costellae on the escutcheon that occur in Neuquenitrigonia. Escutcheon ornamentation is not preserved in our material; hence, an assignment to Neuquenitrigonia cannot be ruled out. Only two species of Neuquenitrigonia are known: N. hunickeni (Leanza and Garate-Zubillaga, Reference Leanza and Garate-Zubillaga1985), from the Bajocian of Neuquén, Argentina, and middle Toarcian–early Aalenian of Atacama, Chile; and N. plazaensis Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, from the middle Toarcian of Atacama, Chile. The second one seems closer in morphology to the material here described. However, Trigonia? sp. 1 is larger, has a wider area, and is somewhat more elongate. Trigonia sp. 1 in Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), from the middle Toarcian of Atacama (Chile), is also similar in morphology, although it is smaller and with middle flank costae more densely arranged; the area ornamentation pattern, on the other hand, is strikingly similar in both taxa. The Toarcian unfigured material from northern Chile referred to Trigonia aff. T. bella Lycett, Reference Lycett1877 (Möricke, Reference Möricke1894, p. 48; see also Pérez and Reyes, Reference Pérez and Reyes1977, p. 11) may be related to this record, according to that broad description. Trigonia sp. B of Ishikawa et al. (Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983), from the Lower Jurassic of south central Perú, is a rather fragmentary specimen, but the observable characters agree with those of Trigonia? sp. 1.
Most Middle Jurassic Trigonia species from the Neuquén Basin can be distinguished easily from Trigonia? sp. 1. Trigonia stelzneri Gottsche, Reference Gottsche and Stelzner1878, has a narrower area, a more clearly opisthogyrate shell, and is relatively shorter. Trigonia corderoi Lambert, Reference Lambert1944, is larger, much more opisthogyrate, and with stronger and blunter flank costae. Trigonia mollensis Lambert, Reference Lambert1944, is more similar to Trigonia? sp. 1 in general shell shape, but the flank costae show some undulations that are absent in the Toarcian material. Trigonia losadai Leanza, Reference Leanza1993 has stronger and more sparsely arranged flank costae.
Trigonia senex from the Hettangian of Japan (Kobayashi and Mori, Reference Kobayashi and Mori1954) bears a large area and an ornamentation pattern similar to that in Trigonia? sp. 1. According to its original description, the Japanese species is particular in having an insignificant marginal carina; unfortunately, the marginal carina is not preserved in any of our specimens. Trigonia sp. described by Poulton (Reference Poulton1976), based on probably Pliensbachian material from south-western British Columbia (Canada), is similar in general shell shape (though slightly smaller); the most noticeable differences are a closer spacing of flank costae and a coarser commarginal ornamentation on the area in the North American taxon. The type species, Trigonia sulcata (Hermann, Reference Hermann1781), and its probable synonym, Trigonia similis Agassiz, Reference Agassiz1840 (see also Bayle Reference Bayle1878, pl. 119, figs. 3–5, as Lyriodon simile; Cox et al., Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969, fig. D66.1), both from the late Early Jurassic of Alsace, are similar in general ornamentation pattern, but they differ from Trigonia? sp. 1 by their more opisthogyrate shell shape.
Trigonia? sp. 2
Figure 5.9–5.11
Occurrence
Arroyo La Laguna (San Juan Province), early Toarcian (D. hoelderi ? Biozone [≈ Serpentinum Biozone]) (Fig. 3).
Description
Large, clearly opisthogyrate shell. Escutcheon wide, poorly preserved. Area narrow, ~1/4 of shell surface (Fig. 5.11), and almost at right angles to the flank; with thin radial costellae intersected by thin commarginal costellae. Mid radial stepped angulation on the area. Marginal carina poorly preserved, seemingly prominent. Antecarinal sulcus narrow. Semicircular flank margin, though in one of the shells the ventral margin is clearly longer than the anterior one. Flank with sub-commarginal sharp costae, less closely spaced as shell grows (8–9 costae/cm on umbonal region, 3–5 costae/cm in middle flank), in late growth stages dense again (6 costae/cm).
Materials
Two shells, CPBA 17430, 17478.
Measurements
CPBA 17430; L = 65 mm, H = 44 mm, W = 36 mm.
Remarks
Although similar to Trigonia? sp. 1, Trigonia? sp. 2 shows some differences that suggest a different species. In Trigonia? sp. 2, the shell is larger and more opisthogyrate; besides, the area is narrower, and area and flank surfaces are almost perpendicular. Both taxa share a pattern of flank costae growing closer at later growth stages; anyhow, this is an age-related character.
As in Trigonia? sp. 1, the assignment to Neuquenitrigonia cannot be ruled out. Trigonia? sp. 2 is similar to Neuquenitrigonia plazaensis, but larger and more opisthogyrate. Trigonia sp. 1 in Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), from the middle Toarcian of Atacama (Chile), is smaller and less opisthogyrate.
Trigonia stelzneri Gottsche, Reference Gottsche and Stelzner1878, from the Bajocian of Paso del Espinacito (San Juan Province), seems close to Trigonia? sp. 2, though it has a subtriangular flank; it is also relatively shorter. Besides, flank costae in T. stelzneri tend to bend ventrally at their anteriormost portion. Trigonia corderoi Lambert, Reference Lambert1944, also has a subtriangular flank with stronger and blunter costae. Trigonia mollesensis Lambert, Reference Lambert1944, bears flank costae with some undulations absent in the Toarcian material. The two species described by Lambert also have a relatively narrower area with stronger radial costellae. Trigonia losadai Leanza, Reference Leanza1993, has stronger and more sparsely spaced flank costae.
This taxon seems close to Trigonia sulcata, although it appears to have a narrower area and, if the illustrations of the European species (Hermann, Reference Hermann1781, pl. 4, figs. 2–4, 9, 10) are to be trusted, Trigonia? sp. 2 differs also by its area orthogonal to the flank. The specimen from Normandy illustrated by Hermann (Reference Hermann1781, pl. 4, figs. 13, 14) as intermediate between T. sulcata and T. dubia (Hermann, Reference Hermann1781) is quite similar to Trigonia? sp. 2.
Family Prosogyrotrigoniidae Kobayashi, Reference Kobayashi1954
Remarks
Kobayashi (Reference Kobayashi1954) characterized the Prosogyrotrigoniidae as lacking a prominent marginal carina and having smooth shell or ornamented by commarginal costae. Cooper (Reference Cooper1991) also considered the prosogyrate umbo and rounded posterior margin as characteristic of the family, and he assumed an origin from the Trigoniidae. Nevertheless, other authors (e.g., Kobayashi and Tamura, Reference Kobayashi and Tamura1968, p. 129–130, table 7) considered the possibility of an independent origin from myophorid genera.
Genus Prosogyrotrigonia Krumbeck, Reference Krumbeck and Wanner1924
Type species
Prosogyrotrigonia timorensis Krumbeck, Reference Krumbeck and Wanner1924, p. 245, by monotypy. Late Triassic, Timor.
Diagnosis
Subovate shell with prosogyrate beaks, trigonian-grade hinge, commarginal ornamentation throughout the shell surface, and with marginal bend, usually associated with a change in costae density between the flank and the area.
Remarks
The diagnosis is modified from Cox (Reference Cox1952) and Cox et al. (Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969). The type species, Prosogyrotrigonia timorensis, was recorded from the Late Triassic of Timor (late Norian, according to Hasibuan, Reference Hasibuan2010); it shows the typical prosogyrate shell shape, commarginal ornamentation, and marginal bend of other species (Kobayashi and Mori, Reference Kobayashi and Mori1954). Prosogyrotrigonia iranica Fallahi et al., Reference Fallahi, Gruber, Tichy and Zapfe1983, from the Norian–Rhaetian of Iran and Late Triassic of Yunnan in China (Hautmann, Reference Hautmann2001, fig. 11), has a subtriangular shell shape, thin and dense commarginal costae, and a rounded marginal bend.
Early Jurassic species included in Prosogyrotrigonia show few differences from Triassic representatives. In this sense, Prosogyrotrigonia can be considered a conservative lineage during the Triassic/Jurassic transition. Early Jurassic records include the species P. tenuis Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, from southern South America, and P. inouyei (Yehara, Reference Yehara1921), from the Hettangian or early Sinemurian of Japan (Kobayashi and Mori, Reference Kobayashi and Mori1954). Frebold and Poulton (Reference Frebold and Poulton1977) and Poulton (Reference Poulton1991, p. 45, 48) also reported P.? cf. P. inouyei from the early Hettangian of Yukon, in northern Canada. From the early Sinemurian of Sonora, Mexico, Scholz et al. (Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, p. 292–293, figs. 10.L, 10.M) illustrated Prosogyrotrigonia sp. A, comparable to both species mentioned above. Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) also mentioned three other undetermined species of Prosogyrotrigonia, mostly from the Sinemurian of Chile, two of which have the commarginal costae broken into irregular tubercles. Considering this appearance of tubercles in the genus, the unusual Hettangian Quadratojaworskiella acarinata Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 78, from Chile, is here referred to Prosogyrotrigonia (see Remarks under Quadratojaworskiella).
Prosogyrotrigonia tenuis Pérez and Reyes in Pérez et al., Reference Pérez and Reyes2008
Figure 5.14–5.17
- Reference Pérez and Reyes1997
Prosogyrotrigonia sp.; Pérez and Reyes, p. 574.
- *v Reference Pérez and Reyes2008
Prosogyrotrigonia tenuis Pérez and Reyes in Pérez et al., p. 64, pl. 3, figs. 1–3, 5–7, 11, 12, pl. 4, figs. 1–12, 14, 15, pl. 5, figs. 2, 5.
- v Reference Damborenea and Echevarría2015
Prosogyrotrigonia tenuis; Damborenea and Echevarría, appendix 2.
- v Reference Damborenea, Echevarría and Ros-Franch2017
Prosogyrotrigonia tenuis; Damborenea et al., p. 101, fig. 4, table 2.
Type material
Holotype: SNGM 486 (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, pl. 4, figs. 11, 12), right valve with ventral margin incomplete; hinge partly visible. From Cerros de Cuevitas, northern Chile.
Occurrence
Arroyo Malo section (Atuel River region), Mendoza Province. Early Sinemurian (Coroniceras-Arnioceras Biozone [≈ Bucklandi-Semicostatum Biozone]) (Fig. 3). The species was first described from Chile, where it occurs in the late Hettangian west of Quillagua and late Hettangian and earliest Sinemurian of Cerros de Cuevitas, Antofagasta (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Poorly preserved material. Small, weakly inflated, orthogyrate to slightly prosogyrate shell (Fig. 5.14). Escutcheon margin straight to slightly convex, at an obtuse angle with the anterior margin (~117–118°). Anterior margin strongly convex, gradually merging with the convex ventral margin. Postero-ventral angle rounded; posterior margin straight to slightly convex. Rounded marginal bend separating flank from area (Fig. 5.15, 5.16). Area occupying 1/4–1/3 of shell surface (Fig. 5.14).
Shell ornamented with commarginal costae, continuous through flank and area (escutcheon not preserved). Intercostal spaces similar in width to costae, sometimes slightly wider; ~6–7 costae/cm at mid-flank. Costae sometimes thinner on the area than on the flank (Fig. 5.15). Area occasionally with bifurcated or extra costellae (Fig. 5.15, 5.16). Costae end abruptly at the escutcheon edge. Costae thinner, more densely packed, and somewhat irregular at late growth stages (Fig. 5.15, 5.16).
Crenulated tooth 3a poorly preserved in one internal mold (Fig. 5.14). Adductor muscle scars very slightly impressed; continuous pallial line (Fig. 5.17).
Materials
Five fragmentary and sometimes strongly corroded shells, one composite mold, one internal mold, and one external mold: MLP 32711, 32786, 32789, 32802, 32803, 32805, 32815, and 32828.
Measurements
MLP 32786: L = 38 mm, H = 31 mm; MLP 32789: L = 39 mm, H = 31 mm; MLP 32805: L = 34 mm, H = 27 mm.
Remarks
The characters observed on the studied specimens show no significant differences from the material described by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) from Chile. The shell seems to be less rectangular in some Argentinian specimens, with the anterior margin slightly more protruding (Fig. 5.14) and narrower area, but such differences are here regarded as intraspecific. Although Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) described thin flank costae separated by much wider intercostal spaces, some of their figured specimens (e.g., pl. 4, figs. 9, 15, pl. 5, fig. 2) show costae as wide as the intercostal spaces. Within the bounds of intraspecific variability accepted by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), the material from west of Quillagua (late Hettangian) is remarkable: shells are orthogyrate to slightly prosogyrate, with a relatively large area (1/3–2/5 of shell surface) and flank costae are densely spaced (8 or more costae/cm). In these shells, area costae are thinner than those from the flank, they are more frequently dichotomous, and the flank costae tend to swell over the marginal bend (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, pl. 4, fig. 3). All these characters closely resemble the genus Frenguelliella, hinting to a possible phylogenetic relationship between both genera, as suggested by Poulton (Reference Poulton1979, p. 18; see also Remarks under Frenguelliella).
Prosogyrotrigonia tenuis has been recorded from the late Hettangian–early Sinemurian of southern South America (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008; this paper). The contemporary species, Prosogyrotrigonia sp. A, is smaller than P. tenuis and more variable in elongation/shell shape (Scholz et al., Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, figs. 10L, 10M), with costae thinner and more densely packed. Shells of P.? cf. P. inouyei in Frebold and Poulton (Reference Frebold and Poulton1977, pl. 2, figs. 5–9) are somewhat larger and coarser, with a somewhat narrower area. The only known specimen of Prosogyrotrigonia sp. 1 of Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, is very similar to P. tenuis, and was found in the same locality where the Hettangian record of that species appeared; the main difference between both taxa is the breaking up of commarginal costae into irregular tubercles in P. sp. 1.
Superfamily Myophorelloidea Kobayashi, Reference Kobayashi1954
Remarks
Cooper (Reference Cooper1991) described the superfamily Myophorelloidea as having posteriorly produced shells with nodate marginal and escutcheon carinae. He characterized the area as broad, with transverse ornamentation and a longitudinal groove. According to his description, flank costae are subcommarginal and entire in primitive forms, but strongly oblique and nodate in more derived forms. The species we describe here share as a distinctive character an area with transverse costellae and a radial median groove (replaced by a row of pustules in certain derived forms, such as Poultoniella new genus).
Family Frenguelliellidae Nakano, Reference Nakano1960
Remarks
Frenguelliellinae has been regarded as a subfamily of the Laevitrigoniidae (Bieler et al., Reference Bieler, Carter and Coan2010; Carter et al., Reference Carter, Altaba, Anderson, Araujo, Biakov, Bogan and Campbell2011). The subfamily Laevitrigoniinae includes mainly genera characterized by weak sculpture, being most likely a polyphyletic group (see Fleming, Reference Fleming1987, p. 37–38 for a discussion on the Laevitrigoniinae). For the time being, we prefer to maintain Frenguelliellidae and Laevitrigoniidae as two different families. The Frenguelliellidae are characterized by shells ornamented with subcommarginal costae (occasionally nodate and oblique), interrupted or attenuated at the antecarinal space (Cooper, Reference Cooper1991).
Genus Frenguelliella Leanza, Reference Leanza1942
Type species
Trigonia inexspectata Jaworski, Reference Jaworski1915, p. 377–380. Pliensbachian, Piedra Pintada (Neuquén, Argentina).
Diagnosis
Shell orthogyrate to clearly opisthogyrate. Well-defined sharp angulations bordering the area, slightly protruding marginal carina in early growth stages. Area with fine commarginal costellae, usually densely spaced. Flank with sub-commarginal costae reaching anterior margin, usually stronger but fewer than in the area. Antecarinal space smooth or delicately ornamented. Escutcheon smooth or with commarginal costellae crossing from the area (though in fewer numbers).
Remarks
Leanza (Reference Leanza1942, p. 164–166) proposed Frenguelliella as a subgenus of Trigonia, and this status was maintained by Cox (Reference Cox1952, p. 54) and Cox in Cox et al. (Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969, p. N478). Tamura (Reference Tamura1959) regarded it as a different genus, and this was followed by most subsequent authors. The original diagnosis by Jaworski (Reference Jaworski1915, Reference Jaworski1925) for Trigonia inexspectata and by Leanza (Reference Leanza1942) for the genus is here amended to encompass the observed range of morphological variability displayed in this genus. Frenguelliella was considered a survivor from the Triassic by Tamura (Reference Tamura1959), who described the subgenus Frenguelliella (Kumatrigonia) Tamura, Reference Tamura1959, based on F. (K.) tanourensis Tamura, Reference Tamura1959, from the Upper Triassic of Japan. He distinguished it from Frenguelliella s.s. by the continuity of flank and area costae in the former. Later, Kobayashi and Tamura (Reference Kobayashi and Tamura1968, p. 107) found shells with more numerous costae on the area than on the flank, and hence considered Frenguelliella and Kumatrigonia as synonyms. Nevertheless, there are some differences between Frenguelliella (Kumatrigonia) tanourensis and the Early Jurassic representatives of Frenguelliella. In Kumatrigonia, there is a prominent marginal carina and an antecarinal sulcus (both as folds of the shell surface, Tamura Reference Tamura1959, pl. 2, figs. 1, 4, discernible even on the internal mold, Tamura Reference Tamura1959, pl. 2, fig. 3). Conversely, Early Jurassic Frenguelliella species bear a prominent carina in the earliest juvenile stages, which is replaced by a sharp marginal angulation and a flat antecarinal space in late juvenile and adult stages. Also, the area in the Triassic species is strongly concave, while in the Jurassic species it is flat. Furthermore, according to Poulton (Reference Poulton1979), there is a transition in the Early Jurassic from Prosogyrotrigonia to Frenguelliella species in North America, expressed as a progressive differentiation of area, flank, marginal angulations, and antecarinal space in Frenguelliella, as well as the development of finer commarginal ornament on the area. According to this view, the distinction of Frenguelliella from the probably ancestral Prosogyrotrigonia is arbitrary and mainly based on the prosogyrate umbos, the simple ovate outline, and the absence of marginal carina or angulation in the types species of Prosogyrotrigonia (Poulton, Reference Poulton1979, p. 18).
On the other hand, the species included in Frenguelliella have an internal radial ridge on the posterior part of the inner surface of the shell. This ridge is not present in Prosogyrotrigonia tenuis, but it seems to occur in Kumatrigonia nemtinovi (Bychkov, Reference Bychkov and Pokhialainen1985, p. 14–15, pl. 4, fig. 8). Given this uncertainty, Kumatrigonia is regarded here as a different genus, containing only Triassic species (K. tanourensis [Tamura, Reference Tamura1959], K. nemtinovi Bychkov, Reference Bychkov and Pokhialainen1985, and probably Frenguelliella? sp. of Newton et al., Reference Newton, Whalen, Thompson, Prins and Delalla1987), and Frenguelliella is considered to have appeared during the Early Jurassic.
Frenguelliella seems to have been restricted to the Pacific coast of the Americas during the Early Jurassic (Feruglio, Reference Feruglio1934; Leanza, Reference Leanza1942; Lambert, Reference Lambert1944; Poulton, Reference Poulton1979; Ishikawa et al., Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983; Leanza, Reference Leanza1993; Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008). The earliest representatives appeared during the Sinemurian and were widely distributed. According to Leanza (Reference Leanza1993, Reference Leanza1996), the genus extended to the early Bajocian in the Andes, but records younger than Toarcian proved to be doubtful. The species Frenguelliella perezreyesi Leanza (Reference Leanza1993, p. 27, pl. 2, figs. 1, 2, 7, 8) was described based on two incomplete specimens from early Bajocian beds at Barda Negra Sur, Neuquén. The morphology of the paratype (Fig. 6.4, 6.5; MOZ-PI 3030/2) and of well-preserved topotypic material personally collected at that locality (Fig. 6.1–6.3; MLP 36314) reveals that this species has prosogyrate umbos, a large smooth lunule, and crenulated margin. This is clearly not a trigoniid, and instead is here referred to the crassatelloidean genus Trigonastarte Bigot, Reference Bigot1895.

Figure 6. Trigonastarte perezreyesi (Leanza) n. comb., early Bajocian, Barda Negra Sur. Scale bar = 10 mm. (1–3) MLP 36314, complete specimen left lateral, dorsal, and anterior views; (4, 5) paratype, MOZ-PI 3030/2, incomplete right valve right lateral and dorsal views (photographs courtesy of B. Boilini).
Frenguelliella eopacifica new species
Figure 7
- Reference Lees1934
Trigonia aff. T. costatula; Lees, p. 42, pl. 4, fig. 6.
- Reference Frebold1964
Trigonia aff. T. costatula; Frebold, p. 14, pl. 5, fig. 6.
- Reference Poulton1979
Frenguelliella sp. B; Poulton, p. 18, pl. 1, fig. 10.
- v Reference Damborenea and Lanés2007
Frenguelliella cf. F. poultoni; Damborenea and Lanés, p. 78, table 3.B, 3.C.
- v Reference Pérez and Reyes2008
Frenguelliella poultoni; Pérez and Reyes in Pérez et al., p. 72, pl. 5, figs. 6–8.
- p ?Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008
Frenguelliella poultoni; Scholz et al., p. 291, fig.10I, 10J.
- v Reference Damborenea and Echevarría2015
Frenguelliella poultoni; Damborenea and Echevarría, appendix 2.
- v Reference Damborenea, Echevarría and Ros-Franch2017
Frenguelliella cf. F. poultoni; Damborenea et al., p. 101, fig. 4, table 2.
Type materials
Holotype: SNGM 541 from Quebrada Pan de Azúcar, 10 km SW of Las Bombas, Atacama Region, Chile; Sinemurian (probably Obtusum Biozone, Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), figured here in Figure 7.5. Paratypes SNGM 561 from Quebrada Pan de Azúcar, 10 km SW of Las Bombas, Atacama Region, Chile, Sinemurian (Fig. 7.1), and MLP 32837 from Las Chilcas, Atuel River region, Mendoza Province, late Sinemurian (Fig. 7.7).
Diagnosis
Small, subtriangular to subrectangular, orthogyrate to slightly opisthogyrate shell. Escutcheon smooth; escutcheon angulation stepped. Area with commarginal costellae; shallow radial groove interrupting them. Prominent marginal carina in early growth stages; sharp angulation in later growth stages. Antecarinal space flat, widening posteriorly. Flank with densely spaced sub-commarginal costae.
Occurrence
Las Chilcas, Arroyo El Pedrero and Codo del Blanco (Atuel River region), Mendoza Province. Sinemurian (≈ Bucklandi–Raricostatum biozones): one specimen (Fig. 7.8) from Coroniceras-Arnioceras Biozone?; all other specimens from Orthechioceras-Paltechioceras Biozone (Fig. 3). The species is also recorded from the early Sinemurian of Yukon, Canada (Poulton, Reference Poulton1979), early Sinemurian of Sierra del Álamo, Mexico (Scholz et al., Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, figs. 10.I, 10.J), and late Sinemurian of Atacama, Chile (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Very small to small, subtriangular (Fig. 7.4) to subrectangular (Fig. 7.3); orthogyrate to slightly opisthogyrate shell. Escutcheon angulation stepped; escutcheon rarely preserved, smooth (Fig. 7.6). Area, ~1/3–1/4 of general shell surface, with fine commarginal costellae, densely spaced (14–19 costae/cm), separated into two sets by a shallow radial groove; costellae dorsal and ventral to the groove may alternate (Fig. 7.4). Prominent marginal carina at early growth stages (Fig. 7.8); a sharp angulation at late growth stages, though highlighted by tubercles corresponding to the costellae on the area (Fig. 7.4, 7.5). Antecarinal space flat, widening posteriorly; somewhat depressed in some shells (Fig. 7.7), most likely a preservational artifact. Antecarinal space barely invaded by area ornamentation (Fig. 7.4), some costellae continuous with the costae of the flank. Flank with densely spaced subcommarginal costae (8–10 or more costae/cm). Flank costae departing from the antecarinal sulcus at right to slightly acute angles (Fig. 7.2).
Etymology
The Greek prefix eo- (from eos = dawn), implying both the eastern horizon and earliness, and -pacifica for the Paleopacific Ocean: “early Frenguelliella from the Eastern Pacific.”
Materials
MLP 18387a, 18389, 32823, 32834, 32843, 32848, 32850, 32851, 32860, 32864, 32875, 32881, 32891: 10 right valves, 7 left valves, and one specimen with both valves, preserved as composite, external, or internal molds, one specimen (Fig. 7.6, 7.9) with shell fragments preserved.

Figure 7. Frenguelliella eopacifica n. sp., Sinemurian of Argentina and Chile. Scale bar = 10 mm. (1) Paratype, SNGM 561, composite mold of left valve, Sinemurian, Quebrada Pan de Azúcar, Chile; (2) MLP 32864a, right valve, late Sinemurian, Las Chilcas; (3) MLP 32875, composite mold of left valve, late Sinemurian, Las Chilcas; (4) MLP 32848, rubber cast of right valve, late Sinemurian, Las Chilcas; (5) holotype, SNGM 541, external mold of left valve, Sinemurian, Quebrada Pan de Azúcar, near Las Bombas, Atacama, Chile; (6, 9) MLP 32864c, fragmented right valve and internal mold, dorsal and right lateral views, late Sinemurian, Las Chilcas; (7) paratype, MLP 32837, right valve, late Sinemurian, Las Chilcas; (8) MLP 32823, rubber cast of right valve, early Sinemurian, Arroyo El Pedrero.
Measurements
Holotype (SNGM 541): L = 11 mm, H = 8 mm. Paratypes: SNGM 561: L = 17 mm, H = 12 mm, and MLP 32837: L = 19 mm, H = 15 mm. MLP 18387a: L = 20 mm, H = 14 mm; MLP 32851: L = 23 mm, H = 17 mm; MLP 32875: L = 13 mm, H = 9 mm; MLP 32881: L = 19 mm, H = 15 mm.
Remarks
Frenguelliella eopacifica n. sp. differs from other Frenguelliella species by its: (1) small size, (2) smooth escutcheon, (3) relatively large area, and (4) orthogyrate to slightly opisthogyrate shell shape. Material assigned to this new species was often referred to as F. poultoni or F. cf. F. poultoni in the literature. Frenguelliella poultoni Leanza, Reference Leanza1993, is regarded here as a junior synonym of Frenguelliella chubutensis (Feruglio, Reference Feruglio1934) (see Remarks under F. chubutensis). The main differences here recognized between F. eopacifica n. sp. and F. chubutensis are the lack of ornamentation on the escutcheon in F. eopacifica n. sp. together with its smaller size. The species is rather homogeneous with regard to these characters throughout the eastern margin of the Paleopacific, from NW Canada to Neuquén. Frenguelliella eopacifica n. sp. differs from F. inexspectata by its much smaller size and proportionally larger area; also, the type species is strongly opisthogyrate.
Although F. eopacifica n. sp. clearly occurs in Argentina, it is more abundant and better preserved in Chile (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008). Specimens from Chile show a smooth escutcheon (Fig. 7.5), a wide area (~1/3 of general shell surface, Fig. 7.1, 7.5), with a shallow radial groove dorsally displaced. Their marginal carina is slightly protruding at the beginning, developing in later growth stages as a sharp angulation, sometimes highlighted by small tubercles arising from the swelling of each area costella (Fig. 7.5). Area costellae and flank costae are closely spaced (16–24 costellae/cm and 9–12 costae/cm, respectively). Area costellae are usually interrupted by the radial groove, and the ventral and dorsal portions of the costellae may alternate (Fig. 7.5). Shell elongation is variable within the species; interestingly, in longer shells the anterior angle between flank costae and antecarinal space tends to be acute, while in short ones it is almost 90°.
Frenguelliella sp. B described by Poulton (Reference Poulton1979) from the early Sinemurian of Yukon (NW Canada) shows no significant differences with the South American specimens. Material referred to Frenguelliella poultoni was also recorded from lower Sinemurian to lower Pliensbachian beds in Sonora (NW Mexico) by Scholz et al. (Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008). Their figured specimens (Scholz et al., Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, fig. 10.I, 10.J, from lower Sinemurian of Sierra del Álamo) agree in shape and size with the material here described. Nevertheless, they considered their specimens as “apparently identical” (Scholz et al., Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, p. 291) to Trigonia cf. T. inexspectata Jaworski, Reference Jaworski1929, which shows significant differences with the material they illustrated (see Remarks under Poultoniella new genus). Since they also included some larger shells up to 45 mm long (Scholz et al., Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, fig. 11), an unusual size for F. eopacifica n. sp., we only refer the two specimens figured by Scholz et al. (Reference Scholz, Aberhan, González-León, Blodgett and Stanley2008, fig. 10.I, 10.J) to F. eopacifica n. sp. The assignment of the remaining specimens will depend on further study.
Frenguelliella chubutensis (Feruglio, Reference Feruglio1934)
Figure 8
- p Reference Jaworski1915
Trigonia inexspectata Jaworski, p. 377 (part; not pl. 5, fig. 2).
- p Reference Jaworski1925
Trigonia inexpectata [sic]; Jaworski, p. 79 (part; not pl. 1, fig. 2).
- Reference Jaworski1926a
Trigonia inexspectata var. densecostata Jaworski, p. 396 (not Trigonia densicostata Röder, Reference Röder1882; nec Trigonia densicostata Marshall, Reference Marshall1919).
- Reference Jaworski1926b
Trigonia inexspectata var. densecostata; Jaworski, p. 180.
- Reference Weaver1931
Trigonia inexspectata var. densecostata; Weaver, p. 233.
- *v Reference Feruglio1934
Trigonia chubutensis Feruglio, p. 34, pl. 4, figs. 9, 11.
- Reference Feruglio1934
Trigonia sp.; Feruglio, p. 36, pl. 4, figs. 10a, b.
- Reference Lambert1944
Trigonia tapiai Lambert, p. 358, pl. 13, fig. 1.
- Reference Pérez and Reyes1977
Trigonia (Frenguelliella) tapiai; Pérez and Reyes, p. 12, pl. 1, fig. 2. [reproduced from Lambert, Reference Lambert1944]
- Reference Volkheimer, Manceñido and Damborenea1978b
Trigonia (Frenguelliella) sp.; Volkheimer et al., p. 212 (table 2).
- v Reference von Hillebrandt and Zeil1980
Frenguelliella tapiai; Hillebrandt, pl. 2, figs. 8a, b.
- v Reference Cuerda, Schauer and Sunesen1982
Frenguelliella tapiai; Cuerda et al., p. 331.
- v Reference Leanza, Garate-Zubillaga and Volkheimer1987
Frenguelliella tapiai; Leanza and Garate-Zubillaga, p. 210, pl. 1, fig. 5.
- v Reference Leanza and Blasco1990
Frenguelliella tapiai; Leanza and Blasco, p. 163.
- v Reference Damborenea and Manceñido1992
Frenguelliella chubutensis; Damborenea et al., pl. 116, fig. 17.
- v Reference Damborenea and Manceñido1992
Frenguelliella tapiai; Damborenea et al., pl. 116, fig. 18.
- Reference Leanza1993
Frenguelliella tapiai; Leanza, p. 26, pl. 1, fig. 8.
- v Reference Leanza1993
Frenguelliella poultoni Leanza, p. 26, pl. 2, figs. 3–6.
- Reference Pérez and Reyes1997
Frenguelliella tapiai; Pérez and Reyes, p. 574.
- v Reference Pérez and Reyes2008
Frenguelliella tapiai; Pérez and Reyes in Pérez et al., p. 70, pl. 5, figs. 1, 3, 4, 9–13.
- v Reference Riccardi, Damborenea, Manceñido, Leanza, Leanza, Arregui, Carbone, Danieli and Vallés2011
Frenguelliella tapiai; Riccardi et al., fig. 7.14.
- v Reference Pagani, Manceñido, Damborenea and Ferrari2012
Frenguelliella sp.; Pagani et al., p. 413, fig. 3c.
Type materials
Lectotype: MLP 3729. Riccardi and Martín (Reference Riccardi and Martín1987, p. 61) listed this specimen as “Holotipo”; yet, since Feruglio (Reference Feruglio1934, p. 34, pl. 4, figs. 9, 11) had established the taxon based on two specimens, their action may be regarded as fixation of lectotype by inference of holotype (ICZN, 1999, Art. 74.6.1.2). The specimen is a composite mold of a right valve, collected by Piatnizky in the Genoa River region (Chubut Province), “lote 20” (most likely the locality now known as Aguada Loca). This is undoubtedly one of Feruglio's syntypes, illustrated by Feruglio (Reference Feruglio1934, pl. 4, fig. 11), by Damborenea et al. (Reference Damborenea, Polubotko, Sey, Paraketsov and Westermann1992, pl. 116, fig. 17), and here (Fig. 8.2). The whereabouts of Feruglio's second specimen (paralectotype) is unknown.
Holotype of Trigonia inexspectata var. densecostata Jaworski
An external mold of a right valve at the Institute of Geosciences, Paleontology Section, Bonn (Germany), IGPB-Jaworski-74, from Cerro Puchenque, Mendoza Province (Figure 8.1).
Type of Trigonia tapiai Lambert
DNGM 43-109, from a bend of arroyo Pichi Picún Leufú, east of Cerro Chachil, Neuquén Province, figured by Lambert (Reference Lambert1944, pl. 13, fig. 1).
Holotype of Frenguelliella poultoni Leanza
MOZ-PI 5315, from Ñireco, east of Cerro Chachil, Neuquén, figured by Leanza (Reference Leanza1993, pl. 2, fig. 3) and here (Fig. 8.3).
Diagnosis
Ovate-subquadrangular to ovate-subrectangular shell, slightly opisthogyrate to orthogyrate. Escutcheon with commarginal costellae crossing from the area. Area wide, with costellae densely arranged, with a submedian radial groove dorsally displaced and sometimes interrupting the costellae. Flank with densely but evenly spaced fine and sharp sub-commarginal costae (up to 20–25 in adult shells), fewer than area costellae (usually about half). Escutcheon angulation stepped; prominent marginal carina at early growth stages, sharp angulation at later ones.
Occurrence
Arroyo La Laguna (Los Patos region) San Juan Province; La Horqueta, Codo del Blanco, Las Chilcas, Quebrada Los Caballos, Puesto Araya, Cerro La Brea (Atuel River region), Arroyo Peuquenes, Arroyo Portezuelo Ancho (Las Leñas/Valle Hermoso region), Arroyo Serrucho, Cerro Puchenque (west of Malargüe region), Mendoza Province; Estación Rajapalo (Cordillera del Viento region), Ñireco, Rahue-Aluminé (Cuerda et al., Reference Cuerda, Schauer and Sunesen1982), Lonqueo (Central Neuquén region), Estancia Santa Isabel (Piedra Pintada region), Neuquén Province; Puesto Peña (central Chubut region), Lomas de Betancourt, Aguada Loca, Nueva Lubecka (Genoa River region), Chubut Province. Early Pliensbachian to late Pliensbachian (M. chilcaense–F. disciforme biozones [≈ Jamesoni–Spinatum biozones]) (Fig. 3), early Toarcian? (Tenuicostatum Biozone).
Description
Small to medium-sized, subquadrangular (Fig. 8.18, 8.19) to subrectangular (Fig. 8.2, 8.15, 8.20) shell. Slightly opisthogyrate to orthogyrate, with umbo anteriorly located. Escutcheon margin slightly convex (Fig. 8.7, 8.19). Area margin long, gently convex or almost straight (Fig. 8.14, 8.19), ventral portion sometimes projected posteriorly (Fig. 8.10, 8.12). Area margin generating right to slightly obtuse angles with escutcheon and ventral margins (Fig. 8.1, 8.2, 8.15, 8.19, 8.20). Ventral margin convex, at a slightly obtuse angle with anterior convex margin. Escutcheon with commarginal costellae crossing from the area (sometimes two area costellae joining into one escutcheon costella; Fig. 8.22); escutcheon margin slightly raised. Escutcheon angulation stepped. Area wide (~1/3–2/5 of general shell surface), with barely impressed radial groove; ventral part slightly wider. Area with fine commarginal costellae, very densely spaced (7–20 costellae/cm; most values 10–16 costellae/cm); sometimes interrupted by the radial groove; dorsal and ventral costellae often alternating (Fig. 8.22); ventral costellae frequently more numerous. Prominent marginal carina at initial growth stages, later as a sharp angulation. Area costellae slightly swollen over the angulations, and most conspicuously tending to bend posteriorly, rendering a pustulose appearance to the marginal angulation (Fig. 8.5, 8.7, 8.9, 8.21, 8.22), which even resembles a prominent carina (Fig. 8.7, 8.10). Antecarinal space widening posteriorly, frequently with flank costae conspicuously weakening; posterior portion usually invaded by area costellae (especially in later growth stages). Costae occasionally deviating when crossing the antecarinal space (Fig. 8.10). Antecarinal space slightly concave in early growth stages, flat in later stages, yet forming a slight angle with the flank (some molds with truly concave sulcus; Fig. 8.10, 8.12, 8.13). Flank with thin, densely spaced (5–15 costae/cm; most values 6–10 costae/cm), sub-commarginal costae (Fig. 8.7, 8.9). Costae sometimes with a rounded posterior bend (Fig. 8.6); most frequently, anterior angle between costa and antecarinal space nearly right (Fig. 8.3, 8.6, 8.12) to slightly acute (Fig. 8.2, 8.5, 8.15, 8.20).

Figure 8. Frenguelliella chubutensis (Feruglio), Pliensbachian to early Toarcian of Argentina. Scale bar = 10 mm. (1) Cast of IGPB-Jaworski-74 (photograph courtesy of G. Heumann), holotype for Trigonia inexspectata var. densecostata Jaworski, an external mold of right valve, Pliensbachian, Cerro Puchenque; (2) MLP 3729, lectotype for Trigonia chubutensis Feruglio, composite mold of right valve, Pliensbachian, Genoa River region; (3) MOZ-PI 5315, holotype for Frenguelliella poultoni Leanza, composite mold of right valve (in association with a left valve), Pliensbachian, Ñireco; (4) rubber cast of MLP 36165, composite mold of right valve with a detail of the hinge, Pliensbachian, Arroyo Serrucho; (5) rubber cast of MLP 36164, external mold of left valve, Pliensbachian, Arroyo Serrucho; (6) MLP 33187c, external mold of left valve, Pliensbachian, Las Chilcas; (7) rubber cast of MLP 36161, external mold of right valve, Pliensbachian, Arroyo Serrucho; (8) MOZ-PI 5316, paratype for Frenguelliella poultoni Leanza, composite mold of right valve, Pliensbachian, Ñireco; (9) rubber cast of MLP 36159a, external mold of right valve, Pliensbachian, Arroyo Serrucho; (10) rubber cast of MLP 28671, external mold of right valve, Pliensbachian, Quebrada Los Caballos; (11) MLP 36179, composite mold of both valves open in butterfly position, Pliensbachian, Lonqueo; (12) MLP 36181, composite mold of right valve, Pliensbachian, Lonqueo; (13) MLP 36172, external mold of right valve, Pliensbachian, Estación Rajapalo; (14) MLP 24468, external mold of right valve, Pliensbachian, Ñireco; (15) MPEF-PI 5861, composite mold of right valve, late Pliensbachian–early Toarcian, Lomas de Betancourt; (16, 17, 20) MPEF-PI 6383, left valve, late Pliensbachian–early Toarcian, Lomas de Betancourt; (16) internal mold; (17) rubber cast of the hinge of 16 (photograph courtesy of A. Pagani); (20) external mold; (18) MPEF-PI 3295, external mold of shell with valves gaping, late Pliensbachian–early Toarcian, Aguada Loca; (19) MPEF-PI 6379, external mold of right valve, late Pliensbachian–early Toarcian, Lomas de Betancourt; (21) MPEF-PI 3390, external mold of juvenile shell open in butterfly position, late Pliensbachian–early Toarcian, Aguada Loca; (22), MPEF-PI 3043, external mold of right valve, late Pliensbachian–early Toarcian, Aguada Loca.
Internal radial ridge in the posterior part of the inner surface, perpendicular to area margin (Fig. 8.12, 8.16). Trigonian-grade hinge (Fig. 8.4, 8.17); densely arranged ridges on both occluding surfaces of 2, 3a, and 3b, on the anterior face of 4b, and on the posterior face of 4a. Muscle scars typical for the group, including the umbonal pedal retractor insertion (Fig. 8.16).
Materials
The type specimens already mentioned above, plus ~220 specimens, MLP: 15321, 15326, 15422, 15428, 15438, 21076, 24467, 24468, 25047, 27821, 27855, 27873, 27877, 27902, 27951, 28671, 28693, 28696, 28698, 28704, 28738, 28985, 28986, 31135, 33185, 33187, 33188, 33191, 33203, 33205, 33267, 36150–36193, 36212, 36310; IANIGLA-PI: 3254–3259, 3264, 3288, 3305–3317, 3319–3343A, 3345–3349, 3352–3354; MPEF-PI: 2911 (part), 2940, 2947, 2957, 3041, 3043, 3048–3051, 3061, 3064, 3293, 3295, 3390, 5787, 5796, 5861, 5875, 6178, 6376–6380, 6382, 6383, 6403–6408, 6410–6416, 6420, 6421; MOZ-PI: 5316, 5317. Specimens from Chile SNGM 501–507.
Measurements
Lectotype (MLP 3729): L = 37 mm, H = 26 mm; largest shell (IANIGLA-PI 3346): L = 61 mm, H = 40 mm; all measurements are plotted in Figure 9.

Figure 9. Scatter plot of height vs. length for Frenguelliella chubutensis (Feruglio).
Remarks
Frenguelliella chubutensis is widely distributed in south-western South America; it seems to be more frequent in the south (Chubut Province), but it is a common component of bivalve faunas from Mendoza and Neuquén provinces as well. A few shells from Chile were also referred to the species (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 70–72, pl. 5, figs. 1, 3, 4, 9–13, as F. tapiai). This taxon is difficult to characterize due to the wide range of morphological variation of the shells. One of the most obvious aspects of intraspecific variability involves elongation: while some shells are subquadrangular (Fig. 8.18, 8.19, 8.21), others are more elongate (Fig. 8.2, 8.5, 8.20). This is evident in H/L ratios, ranging between 0.69–0.92. Allometry analysis of length and height showed an isometric relationship between both variables (slope for log values of height vs. length = 1.03; p(slope=1) = 0.41; Fig. 9), so an ontogenetic variability for the character can be ruled out.
Costae density is also highly variable. For area costellae, the minimum value is 7 costellae/cm, and the maximum is 20 costellae/cm. For the flank, the minimum value is 5 costae/cm (Fig. 8.5, 8.10) and the maximum is 15 costae/cm (Fig. 8.6, 8.8). The main variation shows no clear geographical pattern. On the other hand, much of this variability may actually be a preservational artifact, as illustrated by the material depicted in Figure 8.11: this represents the composite mold of a shell in butterfly position, but due to differential deformation the left valve looks more elongate and with flank costae more densely arranged than the right valve.
The morphology observed in Chilean specimens falls within the variation displayed by the Argentinian material; but there is a single difference: flank costae are interrupted anteriorly by a smooth surface, tapering towards the dorsal and ventral ends, and the costae reach that surface at right angles (although this last feature could be recognized in some Argentinian specimens as well). The material from the Lower Jurassic of Peru described by Ishikawa et al. (Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983, p. 40–41, pl. 1, fig. 7) as Frenguelliella sp. agrees with main shell characters of F. chubutensis (escutcheon “concentric” —commarginal— costellae are mentioned), although its preservation is not good enough to assign it to Feruglio's species with certainty.
Jaworski (Reference Jaworski1926a, p. 396, Reference Jaworski1926b, p. 180) recognized this taxon as new, and named it as a variety of his previously described species, T. inexspectata. He based his detailed description on one external mold of a right valve from Cerro Puchén (= Cerro Puchenque, west of Malargüe region, Mendoza Province, IGPB-Jaworski 74; Fig. 8.1). He also referred two specimens from Arroyo Blanco (Atuel River region) to this variety, which is part of the material he had previously (Jaworski, Reference Jaworski1915) included in T. inexspectata. He did not figure any of the specimens, which has hindered identification of the taxon. Only Weaver (Reference Weaver1931), who had seen Jaworski's specimen at Bonn, used the name T. inexspectata var. densecostata again to refer to specimens from Arroyo Blanco in the Atuel River region, Mendoza, but he did not figure them.
Specimens from central Patagonia (Chubut Province) were described as Trigonia chubutensis by Feruglio (Reference Feruglio1934). Lambert (Reference Lambert1944), in his revision of trigonioids from Neuquén Province, described the new species Trigonia tapiai and distinguished it from T. chubutensis by the higher density of flank costae and lower density of area costellae in the former. Later, Leanza (Reference Leanza1993, p. 2) described the species F. poultoni, based on small shells from central Neuquén, diagnosing it as “finely sculptured and more nearly rectangular in outline than any other known species of Frenguelliella.” He also mentioned, among other characters, the presence of no more than 18 very regular, equally spaced, sub-commarginal costae.
The comprehensive analysis performed here shows that there is a continuous variability of these characters in F. chubutensis (with collections including material from the type localities of all the nominal species involved). Therefore, F. densecostata (Jaworski), F. chubutensis (Feruglio), F. tapiai (Lambert), and F. poultoni Leanza are here regarded as subjective synonyms. Trigonia densecostata Jaworski is a primary junior homonym of Trigonia densicostata Röder, Reference Röder1882 and Trigonia densicostata Marshall, Reference Marshall1919, according to ICZN (1999, Art. 58.12—use of different connecting vowels in compound words). Thus, the next oldest available name (ICZN, 1999, Art. 23.1) for this species is Frenguelliella chubutensis (Feruglio).
As previously discussed (see Remarks under F. eopacifica n. sp.) all the Sinemurian material referred in the literature to F. poultoni (including Frenguelliella sp. B of Poulton, Reference Poulton1979) is here referred to F. eopacifica n. sp.
The diagnosis of Frenguelliella chubutensis is here emended to include the observed variability, which encompasses the type material of the nominal species described by Jaworski (Reference Jaworski1926a), Feruglio (Reference Feruglio1934), Lambert (Reference Lambert1944), and Leanza (Reference Leanza1993). Frenguelliella chubutensis differs from other Frenguelliella species by its: (1) commarginally ornamented escutcheon, (2) relatively large area, and (3) orthogyrate to slightly opisthogyrate shell shape. Frenguelliella inexspectata is more opisthogyrate and has a smooth escutcheon and narrower area. Frenguelliella chubutensis is most likely descended from F. eopacifica n. sp., with which it shares many characters, the main differences being the larger adult size and the presence of escutcheon costellae in F. chubutensis.
Frenguelliella aff. F. chubutensis (Feruglio, Reference Feruglio1934)
Figure 10.1–10.3
Occurrence
Codo del Blanco and Quebrada Los Caballos (Atuel River region), Mendoza Province. Lowermost Pliensbachian, M. chilcaense Biozone (≈ Jamesoni Biozone) (Fig. 3).
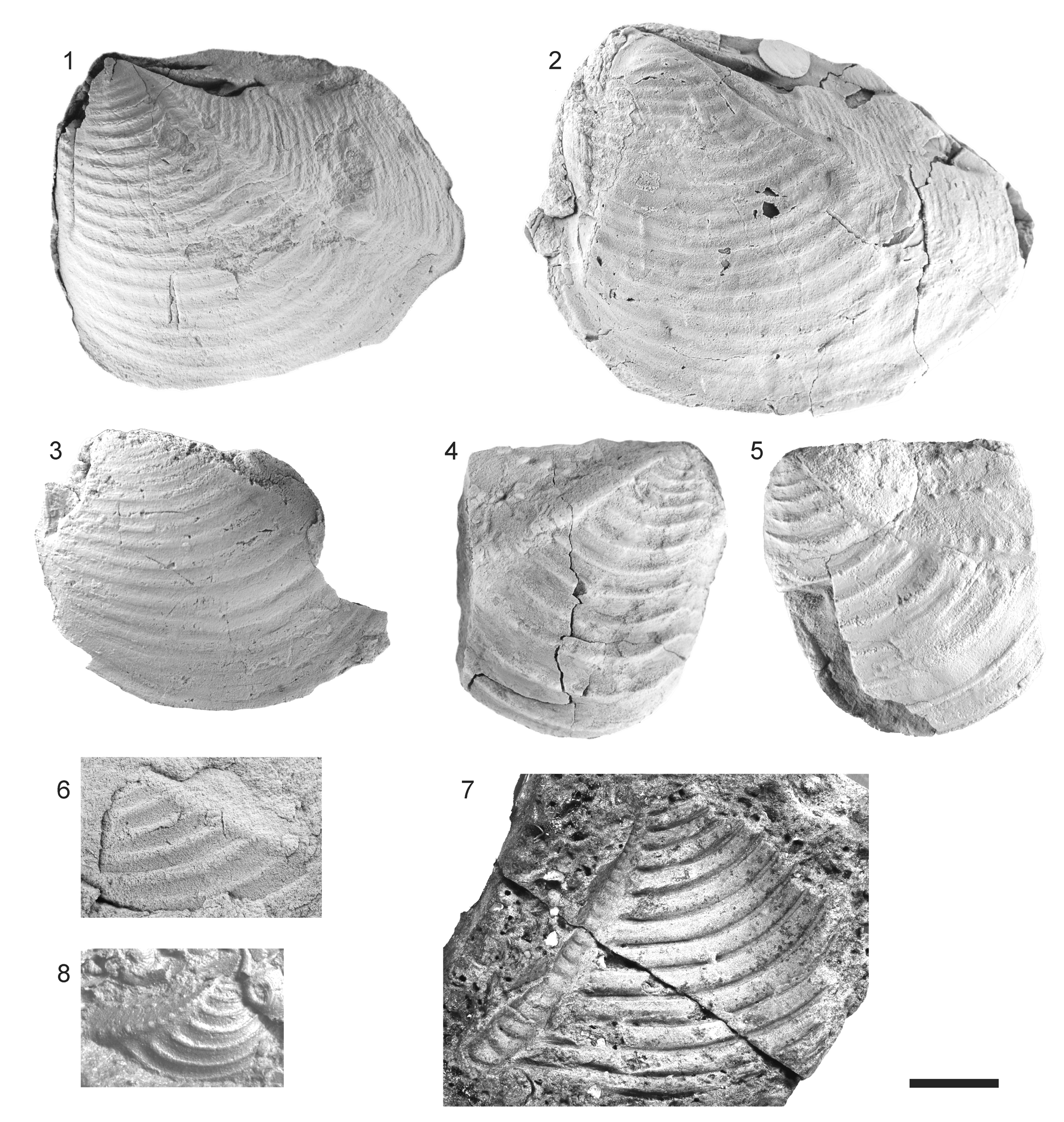
Figure 10. Trigoniida from the Early Jurassic of Argentina and North America. Scale bar = 10 mm. (1–3) Frenguelliella aff. F. chubutensis, Pliensbachian: (1) MLP 28684, composite mold of a left valve, Quebrada Los Caballos; (2) MLP 28994, composite mold of a left valve, Codo del Blanco; (3) MLP 28676, composite mold of a left valve, Quebrada Los Caballos; (4–8) Poultoniella jaworskii n. gen. n. sp.; (4, 5) holotype, MLP 36315, fragmentary shell, right and left lateral views, Toarcian, Arroyo La Laguna; (6) MLP 36317, composite mold of left valve, Toarcian, Arroyo La Laguna; (7) NHMB 28/9/77, n° 527 (photograph courtesy of S. Kühni), external mold of a left valve, Early Jurassic, Sonora, Mexico; (8) SNGM 7281, cast of an external mold of a right valve, late Pliensbachian, Vancouver Island, Canada.
Description
Medium-sized, orthogyrate shell. Escutcheon margin straight or slightly convex (Fig. 10.1). Posterior margin wide, area representing ~1/3–2/5 of shell surface (Fig. 10.1, 10.2). Ventral margin convex; anterior margin gently convex. Umbo anteriorly located. Escutcheon not preserved. Area with commarginal costellae, closely spaced (9–11 costellae/cm), less densely spaced at early growth stages. With marginal angulation. Antecarinal space dominated by attenuated flank costae. Flank costae sub-commarginal, gradually increasing in spacing (Fig. 10.1–10.3) from 6 costae/cm (at early growth stages) to 4 costae/cm (in mature stage). Flank costae slightly converging anteriorly.
Materials
MLP 28676, 28684, 28994 (three left valves, preserved as composite molds).
Measurements
MLP 28994: L = 52 mm, H = 44 mm; MLP 28684: L = 46 mm, H = 37 mm.
Remarks
This taxon might represent an extreme variety of F. chubutensis, but it differs from typical shells of the species by the increase in spacing of flank costae during ontogeny, the larger size, and the orthogyrate shell shape. Frenguelliella inexspectata is similar in size, but strongly differs by its narrower area, its opisthogyrate shell, and its relatively regular costae spacing on the flank.
Trigonia costatula Lycett, Reference Lycett1850, as illustrated by Lycett (Reference Lycett1872–1879, p. 81, pl. 15, figs. 8–10) shows some affinities with Frenguelliella aff. F. chubutensis. The main difference lies in the absence of ornamentation on the area and antecarinal space, and some irregularities on the posterior portion of the last-formed flank costae in the European species.
Frenguelliella inexspectata (Jaworski, Reference Jaworski1915)
Figure 11.1–11.10
- *p Reference Jaworski1915
Trigonia inexspectata Jaworski, p. 377, pl. 5, figs. 2a, b (part; see remarks under F. chubutensis).
- p Reference Jaworski1925
Trigonia inexpectata [sic]; Jaworski, p. 79, pl. 1, fig. 2. (part; see remarks under F. chubutensis).
- Reference Windhausen1931
Trig. inexpectata [sic]; Windhausen, pl. 2, fig. 5. [reproduced from Jaworski, Reference Jaworski1915]
- v Reference Leanza1942
Trigonia (Frenguelliella) inexpectata [sic]; Leanza, p. 165, pl. 7, fig. 1.
- vp Reference Carral-Tolosa1942
Trigonia cf. T. inexpectata [sic]; Carral-Tolosa, p. 57 (non pl. 6, fig. 2).
- Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969
Trigonia (Frenguelliella) inexspectata; Cox in Cox et al., fig. D66.3. [reproduced from Jaworski, Reference Jaworski1915]
- v Reference Damborenea and Manceñido1992
Frenguelliella inexspectata; Damborenea et al., pl. 116, fig. 16.
- Reference Leanza1993
Frenguelliella inexspectata; Leanza, p. 26, pl. 2, fig. 9.
- v Reference Pérez and Reyes2008
Frenguelliella inexspectata; Pérez and Reyes in Pérez et al., p. 74, pl. 6, figs. 1, 3, 4, 7.
- v Reference Camacho, Damborenea, del Río, Camacho and Longobucco2008
Frenguelliella inexspectata; Camacho et al., fig. 14.22.D.
- v Reference Riccardi, Damborenea, Manceñido, Leanza, Leanza, Arregui, Carbone, Danieli and Vallés2011
Frenguelliella inexspectata; Riccardi et al., p. 453, fig. 7.8.
- Reference Ferrari and Bessone2015
Frenguelliella aff. F. inexspectata; Ferrari and Bessone, p. 353, fig. 2.M.

Figure 11. Trigoniida from the Early Jurassic of Argentina. Scale bar = 10 mm. (1–10) Frenguelliella inexspectata (Jaworski) from the Pliensbachian, arrows show the anterior radial ruga: (1, 2) lectotype, IGPB-Jaworski-6 (photograph courtesy of G. Heumann), shell, left and right lateral views, Piedra Pintada region; (3, 6) MLP 13008a, left valve, lateral and dorsal views, Cerro Roth Sur; (4, 5) MLP 36204, left valve, dorsal view and detail of the hinge, Cerro Roth Sur; (7) rubber cast of MLP 36201, external mold of right valve, Subida a Sañicó; (8) MLP 6726, right valve, Cerro Roth; (9) MLP 36198, left valve, Salitral Grande Carrán Curá; (10) MLP 6139, left valve, Cerro Roth; (11–14, 16–21) Jaworskiella burckhardti (Jaworski): (11) holotype, IGPB Jaworski-7 (photograph courtesy of G. Heumann), a fragmentary left valve, Pliensbachian, Piedra Pintada region; (12) BMNH1039-104, holotype for Trigonia catanlilensis Weaver, a weathered shell, left lateral view, Pliensbachian, Catan Lil; (13) MLP 36309 a fragmentary right valve, Pliensbachian, Cerro Roth Sur; (14) MOZ-PI 1064, juvenile left valve retrieved from growth lines on an adult specimen, Pliensbachian, Cerro Roth; (16) MLP 6261, fragmentary right valve, Pliensbachian, Cerro Roth; (17) MLP 36307 shell, dorsal view, Pliensbachian, North of Cerro Roth; (18, 20) MCF-PIPH 503a, fragmentary left valve, dorsal and lateral view, Pliensbachian, Cerro Roth Sur; (19) MPEF-PI 6389, right valve, late Pliensbachian–early Toarcian, Nueva Lubecka; (21) MLP 36306 shell, left lateral view, Pliensbachian, North of Cerro Roth. (15) MLP 36303, probably Jaworskiella burckhardti, right valve, Pliensbachian, Salitral Grande Carrán Curá.
Type material
The lectotype (here designated) is a complete shell with conjoined valves, figured by Jaworski (Reference Jaworski1915, pl. 5, figs. 2a, b), and here (Fig. 11.1, 11.2), from the Pliensbachian of Piedra Pintada region, held at the Institute of Geosciences, Paleontology Section, Bonn, IGPB-Jaworski-6.
Diagnosis
Medium-sized to large, ovate-subtriangular shell; clearly opisthogyrate. Escutcheon smooth, depressed. Area narrow, with closely spaced commarginal costellae. Flank with strong, sharp, sub-commarginal, and widely spaced costae. Anterior radial ruga on the flank separating the commarginal portion of the costae from the oblique one. Antecarinal space bordered by swellings of flank costae, carrying a more subtle ornamentation than the flank.
Occurrence
Salitral Grande Carrán Curá, Estancia Santa Isabel, Subida a Sañicó, Cerro del Vasco, Cerro Roth Sur (Piedra Pintada region), Neuquén Province; Nueva Lubecka (Genoa River region), La Casilda (in Ferrari and Bessone, Reference Ferrari and Bessone2015), Chubut Province. Latest early Pliensbachian to late Pliensbachian (E. meridianus–F. disciforme biozones [≈ Davoei–Spinatum biozones]) (Fig. 3), early Toarcian? (Tenuicostatum Biozone). The species is also recorded from Quebrada Asientos (Atacama, Chile) from the early/late Pliensbachian (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Medium-sized to large, opisthogyrate shells. Straight escutcheon margin, but with the umbo and the posterodorsal angle slightly projecting dorsally (Fig. 11.3). Area margin narrow, area representing ~1/4–1/5 of shell surface. Ventral margin gently convex; anterior margin convex and somewhat projecting anteriorly (umbo located at ~1/4 of shell length). Escutcheon excavated, smooth (some shells with commarginal costellae or striae; Fig. 11.4); escutcheon angulation stepped. Area with sub-median groove, with commarginal costellae (Fig. 11.6), loosely arranged in early growth stages (6 costellae/cm), more closely spaced later on (10–11 costellae/cm); dorsal and ventral portions normally meeting at an angle. Prominent marginal carina in early growth stages, turning into a sharp angulation later. Antecarinal sulcus present at very early growth stages, widening and shallowing posteriorly (antecarinal space), usually dominated by attenuated flank costae; otherwise, posterior half invaded by area costellae (especially at late growth stages). Flank costae extending across the antecarinal space without attenuation at late growth stages. Flank costae strong, widely spaced (3–4 costae/cm), sub-commarginal, reaching the anterior margin nearly at right angles; sometimes, denser costae arrangement at late growth stages (Fig. 11.1, 11.2, 11.8); subtle swelling of flank costae bordering the antecarinal space. Anterior radial ruga on the shell, generating a prominence on the costae (Fig. 11.4–11.6); better developed at early growth stages, fading at later ones.
Trigonian-grade hinge (Fig. 11.5). Nymph short (Fig. 11.4, 11.6).
Materials
About 30 specimens. The lectotype mentioned above, plus MLP 6139, 6726, 13008 (part), 36195–36210, MCF-PIPH 502 (part), 631; MPEF-PI 2911 (part), 6385, 6387, 6400; and DNGM 8851 and 8863. Specimens from Chile SNGM 562, 563.
Measurements
Lectotype: L = 56 mm, H = 46 mm (G. Heumann, personal communication, 2019); MLP 13008a: L = 61 mm, H = 45 mm; MLP 36202: L = 48 mm, H = 37 mm; MLP36201: L = 59 mm, H = 39 mm.
Remarks
In his original description, Jaworski (Reference Jaworski1915) included material from southern Neuquén and also some specimens from southern Mendoza, but the latter were subsequently separated by him as part of his new variety Trigonia inexspectata var. densecostata (see Remarks under F. chubutensis), thus restricting F. inexspectata s.s. to the material from Piedra Pintada region.
The shells from Chile, described by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), show all the main features of the species, but with smaller shell size and more densely spaced flank costae (5–6 costae/cm, or even denser at final growth stages).
This taxon is easily differentiated from other Frenguelliella species from the region by its: (1) larger size, (2) more opisthogyrate shell shape, (3) narrower area, (4) stronger and less densely spaced flank costae, and (5) anterior radial ruga. It has a fairly limited geographic distribution, restricted to certain facies, such as biofacies A in Damborenea et al. (Reference Damborenea, Manceñido and Riccardi1975), which corresponds to shallow-water, well-lit environments.
Genus Jaworskiella Leanza, Reference Leanza1942
Type species
Trigonia burckhardti Jaworski, Reference Jaworski1915, p. 380–382. Pliensbachian, Piedra Pintada (Neuquén Province, Argentina).
Diagnosis
Shell opisthogyrate, with well-defined sharp angulations. Escutcheon smooth and slightly excavated. Area with fine commarginal costellae, usually densely spaced. Antecarinal space developed. Flank ornamentation variable, but always beginning with sub-commarginal sharp costae. Posterior segment of later costae broken up into irregular tubercles (not always developed); anterior segment oblique to growth lines and sharp. Anterior radial ruga variably developed on the flank.
Remarks
The diagnosis is slightly modified from the original one by Leanza (Reference Leanza1942, p. 166–168), when he proposed Jaworskiella as a subgenus of Trigonia. Cox (Reference Cox1952) regarded Jaworskiella as a subgenus of Myophorella, but later considered it as a separate genus (Cox et al., Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969, p. N481), an interpretation that was followed by most subsequent authors.
The type species for the genus, T. burckhardti from the Pliensbachian of Piedra Pintada, seems to be closely related to F. inexspectata (type species of Frenguelliella, described above). Escutcheon and area sculptures are the same in both species, and they both share an anterior radial ruga on the flank. The main differences seem to lie in the development of the anterior flank and of the ornamentation. The anterior margin in J. burckhardti is more extended (compared to the ventral margin) and more convex than in F. inexspectata, resulting in a more opisthogyrate shell; inflation is also stronger in Jaworskiella. Besides, the flank in J. burckhardti is dominated by oblique ornamentation, a pattern already developed on the anteriormost portion of flank costae in F. inexspectata and related to the anterior margin. The tubercles in J. burckhardti (more or less developed) are fairly irregular. Furthermore, there seems to be a gradation in morphologies (Fig. 11.1–11.21) between those typical of F. inexspectata and the extreme forms of J. burckhardti. So, both species are closely related (most likely sister species, or even a single species with a high degree of variation). The two species frequently occur in the same localities and even the same beds. At this time we prefer to maintain the specific and generic distinction, but we regard Jaworskiella as monotypic, other proposed species being referred to different genera.
Taking into account that most of the remaining species that were previously included in Jaworskiella (Pérez and Reyes, Reference Pérez and Reyes1977; Poulton, Reference Poulton1979; Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) show some meaningful morphological differences and/or are Sinemurian in age, we consider unlikely that they could have been genetically related to J. burckhardti. Cox (Reference Cox1952) also referred to this genus the species Trigonia freixialensis Choffat, Reference Choffat1885, and T. kobyi Choffat, Reference Choffat1885, from the Upper Jurassic in Portugal. Even though flank ornamentation in both Portuguese species is reminiscent of that of J. burckhardti, the lack of an antecarinal space and of area ornamentation (except for earliest growth stages) precludes their assignment to the genus.
Jaworskiella burckhardti (Jaworski, Reference Jaworski1915)
Figure 11.11–11.21
- v Reference Burckhardt1902
Trigonia gryphitica Steinm. [sic]; Burckhardt, p. 245, pl. 4, fig. 4.
- * Reference Jaworski1915
Trigonia burckhardti Jaworski, p. 380, pl. 5, fig. 3.
- Reference Jaworski1925
Trigonia burckhardti; Jaworski, p. 81, pl. 1, fig. 3.
- v Reference Weaver1931
Trigonia catanlilensis Weaver, p. 233, pl. 20, fig. 98.
- Reference Windhausen1931
Trigonia Burckhardti; Windhausen, pl. 20, fig. 4. [reproduced from Jaworski, Reference Jaworski1915]
- Reference Windhausen1931
Trigonia gryphytica Stein. [sic]; Windhausen, pl. 22, fig. 7. [reproduced from Burckhardt, Reference Burckhardt1902]
- v Reference Leanza1942
Trigonia (Jaworskiella) burckhardti; Leanza, p. 166, pl. 4, figs. 2, 3.
- v Reference Carral-Tolosa1942
Trigonia aff. T. Burckhardti; Carral-Tolosa, p. 58, pl. 6, fig. 2 (as Trigonia cf. T. inexpectata [sic] in plate caption).
- Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969
Jaworskiella burckhardti; Cox in Cox et al., fig. D68.6. [reproduced from Jaworski, Reference Jaworski1915]
- v Reference Pérez and Reyes1977
Jaworskiella burckhardti; Pérez and Reyes, p. 14, pl. 2, figs. 1, 3, 6. [figs. 1, 3 reproduced from Leanza, Reference Leanza1942; fig. 6 reproduced from Jaworski, Reference Jaworski1915]
- v Reference von Hillebrandt and Zeil1980
Jaworskiella burckhardti; Hillebrandt, pl. 2, figs. 5a, b.
- v Reference Leanza, Garate-Zubillaga and Volkheimer1987
Jaworskiella burckhardti; Leanza and Garate-Zubillaga, p. 210, pl. 1, figs. 1, 2.
- v Reference Leanza1993
Jaworskiella burckhardti; Leanza, p. 28, pl. 1, figs. 1, 6, 7, 10.
- Reference Pérez and Reyes1997
Jaworskiella burckhardti; Pérez and Reyes, p. 574.
- v Reference Pérez and Reyes2008
Jaworskiella burckhardti; Pérez and Reyes in Pérez et al., p. 75, pl. 6, figs. 2, 8, 9, pl. 7, figs. 1–3, 6, 7, pl. 8, fig. 3.
Type materials
Holotype: IGPB Jaworski-7, an incomplete left valve from the Pliensbachian of Piedra Pintada (Neuquén Province), wrongly attributed originally to the Upper Jurassic. Jaworski's (Reference Jaworski1915) original description was based on this single left valve (illustrated in his pl. 5, fig. 3; Fig. 11.11).
Holotype of Trigonia catanlilensis Weaver
A weathered juvenile shell at the Burke Museum of Natural History, Seattle (USA), BMNH1039-104 (a cast at Museo de La Plata under collection number MLP 22356), from the late Early Jurassic of Catan Lil (Piedra Pintada region), Neuquén Province (Fig. 11.12).
Diagnosis
Large to very large opisthogyrate shell, ovate-subtriangular to semicircular, anteriorly inflated. Escutcheon smooth, depressed. Area narrow, with closely disposed commarginal costellae. Flank ornamentation beginning with sub-commarginal sharp costae; later costae with posterior segment broken up into irregular tubercles (not always developed), and anterior segment oblique and sharp. Anterior radial ruga on the flank. Antecarinal space more subtly ornamented than the flank.
Occurrence
Salitral Grande Carrán Curá, Cerro del Vasco, Cerro Roth Sur (Piedra Pintada region), Neuquén Province; Nueva Lubecka (Genoa River region), Chubut Province; doubtful specimen from La Bajada (Las Leñas/Valle Hermoso region), Mendoza Province. Latest early Pliensbachian (A. behrendseni Biozone) to early Toarcian (Tenuicostatum Biozone?; Fig. 3). The species is also recorded from the late Pliensbachian and Toarcian in Quebrada Asientos (Atacama) and Punilla Oriental (Coquimbo) in Chile (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Very large opisthogyrate shell, strongly inflated anteriorly. Straight and short escutcheon margin, with the umbo and the posterodorsal angle projecting dorsally (Fig. 11.15, 11.19, 11.21). Posterior margin not preserved in the known specimens; short and slightly convex according to growth lines. Convex ventral margin, passing gradually to the strongly convex anterior margin (Fig. 11.19, 11.21). Umbos prominent, anteriorly situated. Escutcheon narrow, slightly excavated, smooth (Fig. 11.17, 11.18) or with strong growth lines. Escutcheon angulation stepped. Area narrow, ~1/4 –1/5 (or less) of general shell surface, though hard to estimate due to fragmentation; somewhat wider on the shells closest to F. inexspectata (Fig. 11.15). Submedian radial groove developed; ventral portion of the area wider than dorsal portion. Area with commarginal, somewhat irregular costellae, less densely spaced in late ontogenetic stages, widening when crossing both angulations; ventral portion with some extra costellae interrupted at the groove. Marginal carina slightly protruding in early growth stages, changing gradually to a sharp angulation (sometimes with tubercles enhancing it). Antecarinal space flat, widening posteriorly, smooth, or, more frequently, with thin costellae (weakened continuation of flank costae) perpendicular to marginal angulation (especially in early growth stages) or commarginal (mostly in late growth stages). Flank ornamentation highly variable; strong and sharp commarginal costae developed in early growth stages, with a bend on the posterior portion, at right angles with marginal carina/angulation. Flank costae oblique in late growth stages, with the posterior segment weakened, giving rise to the antecarinal space. Some shells with flank costae similar to those of Frenguelliella inexspectata, although always at right angles with the antecarinal space (Fig. 11.13, 11.15). Maximum costae separation at the point of maximum shell inflation (Fig. 11.11, 11.16, 11.21), resulting in costae with a diverging posterior segment and a converging anterior one (recognized by Jaworski, Reference Jaworski1925). Costae posterior segment usually fragmented into irregular tubercles (Fig. 11.11, 11.14, 11.16, 11.20, 11.21), but not always (e.g., Fig. 11.13). Costae anterior segment continuous, frequently with irregularities and bifurcations (Fig. 11.16, 11.21). Anterior radial ruga developed in early stages, fading later on.
Trigonian-grade hinge (Fig. 11.18). Nymph short and wide (Fig. 11.17).
Materials
More than 30 specimens were examined: MLP 6257, 6261, 36213, 36302, 36304–36309, and probably 17145, 36303; MOZ-PI 1064, 3043, 3044; DNGM 8684 (cast = MLP 19080); MPEF-PI 2910, 2997, 6388, 6389, 6391, 6401, and probably 6398; MCF-PIPH 502 (part), 503. The specimen from Piedra Pintada illustrated by Burckhardt (Reference Burckhardt1902, pl. 4, fig. 4) as Trigonia gryphitica was examined at Basel (Switzerland): NHMB-G 16636. Specimens from Chile SNGM 568–572.
Measurements
Holotype (IGPB Jaworski-7): L = 110 mm, H = 65 mm (G. Heumann, personal communication, 2019), though the ventral margin is broken; few shells are complete enough to be measured: MLP 36306: L = 82 mm, H = 60 mm, W = 49 mm; MLP 36307: L = 79 mm, H = 59 mm, W = 46 mm; MPEF-PI 6389: L = 73 mm, H = 50 mm, V = 20 mm.
Remarks
The strong resemblance of this species to F. inexspectata, from which it differs mostly in shell size and anterior inflation, strongly suggests a sister-species relationship. Some differences in ornamentation seem to be the result of alterations of the shell. For example, in some shells of J. burckhardti (Fig. 11.21), it is clear that posterior portions of costae (at least in latest growth stages) are still commarginal, and their anterior portions are oblique, as is actually the case in F. inexspectata. It is also strongly suggestive that both species frequently occur together. These similarities sometimes make the distinction of both taxa difficult. The specimen on Figure 11.15, for example, shows many characters typical of F. inexspectata, such as the continuous and sharp flank costae, the relatively large area and the less-inflated anterior flank; in contrast, the strongly opisthogyrate shell shape, the slightly irregular costellae on the area, and the anterior irregularities on flank costae are characteristic of J. burckhardti.
As correctly suggested by Poulton (Reference Poulton1979, p. 19), the type material shows tubercles on the posterior portion of flank costae (Fig. 11.11), unlike Jaworski's drawing (Reference Jaworski1915, pl. 5, fig. 3), which shows smooth flank costae. The wide but continuous variability in shell ornamentation (Fig. 11.11–11.21) allows for the inclusion of specimen BMNH1039-104 (Fig. 11.12) within the species. This heavily weathered material is the holotype for Trigonia catanlilensis Weaver, Reference Weaver1931, and hence this last species is considered as a junior synonym of Jaworskiella burckhardti. This specimen is a small juvenile, comparable to other juvenile shells included within the species (e.g., Fig. 11.14).
Genus Poultoniella new genus
Type species
Poultoniella jaworskii new species, by original designation herein. Late Pliensbachian–early Toarcian of the Pacific coast of the Americas.
Other species
Trigonia malladae Choffat, Reference Choffat1885, from the late Toarcian of Portugal, may be tentatively referred to Poultoniella n. gen.
Diagnosis
Orthogyrate to opisthogyrate. Prominent marginal carina with transversely elongate tubercles on top. Area smooth except for a median radial row of well-spaced, round, small pustules. Very narrow antecarinal space. Flank with sharp sub-commarginal costae.
Occurrence
Late Pliensbachian and Toarcian from the Pacific coast of the Americas and maybe Western Europe.
Etymology
The genus is dedicated to Terence P. Poulton, Canadian paleontologist and specialist on fossil Trigoniida.
Remarks
Although the material of the type species was referred to Frenguelliella or related to species now included in it (Jaworski, Reference Jaworski1929; Poulton, Reference Poulton1979), area ornamentation and marginal carina morphology prevent its assignment to that genus, as here understood. Furthermore, previous characterizations of Frenguelliella emphasized the absence of any trace of radial ornament on the area (Cox in Cox et al., Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969, p. N478; Poulton, Reference Poulton1979, p. 18). Yet, we consider it unlikely that the single radial row of pustules along the area midline would relate this genus to Trigonia, which has well-developed radial costae over the entire area.
Instead, Poultoniella n. gen. is here interpreted as a descendant of Frenguelliella, with the rows of elongate tubercles on the marginal carina and escutcheon angulation, and the central row of pustules representing thickened remains of former transverse costellae across the area. The prominent marginal carina would be another innovation of this taxon. The abrupt appearance of these features hinders relating it to any particular species of Frenguelliella.
The description and figure of Trigonia malladae Choffat (Reference Choffat1885, pl. 9, fig. 23, Reference Choffat1888, p. 37) suggest the presence of Poultoniella n. gen. in the late Toarcian of Portugal.
Poultoniella jaworskii new species
Figure 10.4–10.8
- v Reference Jaworski1929
Trigonia cf. T. inexspectata Jaw.; Jaworski, p. 7, pl. 1, fig. 4.
- p Reference Poulton1979
Frenguelliella sp. A; Poulton, p. 18, pl. 1, figs. 4, 5, 22, non figs. 6–8.
Diagnosis
Orthogyrate and trigonally elongate shell. Escutcheon carina as a row of spaced, very small, and transversely elongate tubercles. Area ~1/3 of shell surface, smooth except for a median radial row of well-spaced, round, small pustules. Prominent marginal carina with regular transversely elongate tubercles; very narrow antecarinal space. Flank costae sub-commarginal, gradually increasing in spacing, sharp, with a sudden downward bend anteriorly.
Occurrence
Arroyo La Laguna (Los Patos region), San Juan Province. Early Toarcian (somewhere between late D. hoelderi Biozone and early C. chilensis Biozone [≈ Serpentinum Biozone or, more likely, Bifrons Biozone]) (Fig. 3). Material assignable to this species was recorded from the late Pliensbachian of Vancouver Island, British Columbia (Canada) by Poulton (Reference Poulton1979) and the Early Jurassic of Sonora (Mexico) by Jaworski (Reference Jaworski1929).
Description
Medium-sized, orthogyrate, and apparently trigonally elongate shell (all materials fragmentary). Anterior and ventral margins slightly convex, meeting at nearly right angles (Fig. 10.4, 10.5). Escutcheon poorly preserved, with some tuberculate costellae. Escutcheon carina as a row of very small, spaced, transversely elongate tubercles. Area ~1/3 of shell surface, smooth; without median groove, but with a radial row of small, well-spaced, and round pustules. Prominent marginal carina with regular transverse ridge-like tubercles. Antecarinal space present as an interruption of flank costae, very narrow but widening posteriorly. Flank with sharp sub-commarginal costae showing gradually increasing spacing (Fig. 10.4, 10.5), from 7 costae/cm on the umbonal region to 2–3 costae/cm in final growth stages. Flank costae with a sudden downward bend near the anterior margin, becoming somewhat irregular (Fig. 10.4) and meeting the margin at high angles.
Etymology
The species is dedicated to the memory of Erich Jaworski, German paleontologist, who first described material referable to this taxon.
Materials
The holotype, plus eight isolated valves and some fragments, MLP 36312b, 36316–36320, mostly recrystallized shells and composite molds. The specimen from Mexico described by Jaworski (Reference Jaworski1929) is housed in the Naturhistorisches Museum Basel (Switzerland): NHMB 28/9/77, n° 527 (Fig. 10.7). Casts of the material described by Poulton (Reference Poulton1979) as Frenguelliella sp. A are housed at the SERNAGEOMIN in Santiago (Chile) under catalog numbers SNGM 7280–7283 (one of them illustrated in Fig. 10.8).
Measurements
Holotype, MLP 36315 (broken posteriorly): L = 30 mm (the whole shell was longer), H = 33 mm. MLP 36317: L = 28 mm, H = 19 mm. Jaworski's (Reference Jaworski1929) specimen from Mexico: L = 45 mm, H = 36 mm (though shell margins are not well preserved).
Remarks
The material from Argentina is strikingly similar to that found in North America. Both the description and the figure by Jaworski (Reference Jaworski1929, p. 7, pl. 1, fig. 4; Fig. 10.7) agree with P. jaworskii n. gen. n. sp. Likewise, Frenguelliella sp. A (Fig. 10.8) described by Poulton (Reference Poulton1979, p. 18, pl. 1, figs. 5, 22 and potentially fig. 4, from Geological Survey of Canada, GSC locality 93587, but not figs. 6–8, from the GSC locality 89090) shows the same characters as the Argentinian material, although the antecarinal space is broader in the specimens from Vancouver Island. This material was initially regarded as of controversial age (specimens either Sinemurian or Toarcian, according to Poulton, Reference Poulton1979); subsequently, Aberhan (Reference Aberhan1998, appendix I) listed the locality involved as late Pliensbachian in age (Kunae Biozone [≈ Margaritatus Biozone]).
As highlighted by Poulton (Reference Poulton1979), the most noticeable character of this species is the lack of commarginal costellae on the area. Jaworski (Reference Jaworski1929) described some faint transverse costellae in initial growth stages that later fade, although he considered that their absence might be also due to a preservational bias.
Flank costae somewhat resemble those of F. inexspectata. Although this last species usually has a sharp marginal angulation, on some shells there seems to be an incipient carina similar to that of Poultoniella jaworskii n. gen. n. sp. In contrast, the progressive spacing of flank costae in Poultoniella jaworskii n. gen. n. sp. is similar to the pattern found in Frenguelliella aff. F. chubutensis, but the Toarcian species is more elongate, with a narrower area and a better-defined nodose carina.
Poultoniella? malladae (Choffat, Reference Choffat1885) n. comb., from the late Toarcian of Portugal, differs from Poultoniella jaworskii n. gen. n. sp. in being higher and shorter and with closer spacing of flank costae.
Genus Quadratojaworskiella Reyes and Pérez, Reference Reyes and Pérez1980
Type species
Jaworskiella (Quadratojaworskiella) pustulata Reyes and Pérez, Reference Reyes and Pérez1980, p. 87–93. Early Pliensbachian, northern Chile.
Diagnosis
Very large subquadrate shell, slightly opisthogyrate. Area wide, divided by a shallow median groove; ornamented by commarginal costae. Marginal carina blunt, slightly overhanging a narrow antecarinal sulcus. Flank ornamented with sharp sub-commarginal costae in early growth stages, changing gradually during development to rows of tubercles.
Remarks
Quadratojaworskiella was proposed by Reyes and Pérez (Reference Reyes and Pérez1980) as a subgenus of Jaworskiella; the diagnosis here presented is modified from their original one. Subsequently, Pérez and Reyes (Reference Pérez and Reyes1991) and Pérez et al. (Reference Pérez, von Hillebrandt, Reyes and Aberhan2006, Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) regarded Quadratojaworskiella as a distinct genus. Its early development suggests a direct derivation from Frenguelliella rather than a close relationship to Jaworskiella or Moerickella n. gen. (see below). Quadratojaworskiella is endemic of southern South America (Chile and Argentina).
Quadratojaworskiella acarinata Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, from the Hettangian and late Sinemurian of Chile, was diagnosed as having a “shell without marginal carina and antecarinal sulcus…” (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 80). These authors also described the Sinemurian species Prosogyrotrigonia sp. 1 and Prosogyrotrigonia sp. 2, both characterized by commarginal flank costae, which in late ontogeny break up into rows of tubercles. Because Prosogyrotrigonia lacks a true carina (the genus is characterized by a marginal bend) and already shows an early tendency to generate tubercles on the flank, we refer Q. acarinata Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, to Prosogyrotrigonia. Quadratojaworskiella is thus regarded as monotypic.
Quadratojaworskiella cf. Q. pustulata (Reyes and Pérez, Reference Reyes and Pérez1980)
Figure 12
- cf. Reference Pérez and Reyes1977
Jaworskiella (Quadratojaworskiella) pustulata Pérez and Reyes, p. 15, pl. 2, figs. 2, 4, 5 (nom. nud.)
- *v cf. Reference Reyes and Pérez1980
Jaworskiella (Quadratojaworskiella) pustulata Reyes and Pérez, p. 89, pl. 1, 2.
- cf. Reference von Hillebrandt and Zeil1980
Quadratojaworskiella pustulata; Hillebrandt, pl. 2, fig. 9.
- cf. Reference Pérez1982
Jaworskiella (Quadratojaworskiella) pustulata; Pérez, pl. 15, figs. 7–10, appendix 1.
- cf. Reference Pérez and Reyes1997
Quadratojaworskiella pustulata; Pérez and Reyes, p. 574.
- v cf. Reference Pérez, Aberhan, Reyes and von Hillebrandt2008
Quadratojaworskiella pustulata; Pérez et al., p. 76, pl. 11, figs. 2–4, 6, 7.
- v cf. Reference Pérez and Reyes2008
Quadratojaworskiella pustulata; Pérez and Reyes, p. 28.

Figure 12. Quadratojaworskiella cf. Q. pustulata (Reyes and Pérez), Pliensbachian, Arroyo Portezuelo Ancho. Scale bar = 10 mm. (1, 2) IANIGLA-PI 3263, external mold of the shell; (1) right and (2) left valves.
Occurrence
Arroyo Portezuelo Ancho (Las Leñas-Valle Hermoso region), Mendoza Province. Latest early Pliensbachian–late Pliensbachian (Fanninoceras s.l. Biozone; Fig. 3). The species Q. pustulata is found in Quebrada Asientos (Atacama, Chile) in the early/late Pliensbachian (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Fragmentary single specimen preserved as external mold of the ventroposterior portion of a left valve and a portion of a right flank. Very large shell (probably >100 mm long). Area and flank surfaces gently sloping away, separated by a straight marginal angulation or marginal bend. Area wide, bearing commarginal strong and irregular rugae (Fig. 12.2), sometimes fragmented into tubercles. Flank with commarginally aligned strong tubercles, somewhat elongate (parallel to growth lines) in late growth stages (Fig. 12).
Material
External mold of a shell (in butterfly position), IANIGLA-PI 3263.
Remarks
The few observable characters agree well with those described for Q. pustulata. Although marginal carina and antecarinal sulcus were described and illustrated by Reyes and Pérez (Reference Reyes and Pérez1980), these features lose prominence as growth proceeds (e.g., Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, pl. 8, fig. 6, pl. 9, fig. 1, pl. 10, fig. 4). Their absence in our material might well be a preservational bias (it is an incomplete fragment preserved as external mold). Because the umbonal part of the shell is missing, the specific assignment remains uncertain. This is the first record of the genus for Argentina.
Family Myophorellidae Kobayashi, Reference Kobayashi1954
Remarks
The family Myophorellidae was characterized as having conspicuously nodate flank costae, sub-commarginal near the umbo, but becoming strongly oblique in maturity (Cooper, Reference Cooper1991). Because those characters may develop in some Frenguelliellidae (such as Jaworskiella), and taking into account the inferred evolutionary relationships of the species discussed here, we only include in Myophorellidae those species that are likely descendants of Moerickella n. gen.
Genus Moerickella new genus
Type species
Trigonia infraclavellata Möricke, Reference Möricke1894, p. 46–48, pl. 2, fig. 3, from the Early Jurassic of Las Amolanas, Chile.
Other species
Jaworskiella siemonmulleri Poulton, Reference Poulton1979; Jaworskiella supleiensis Poulton, Reference Poulton1979.
Diagnosis
Shell opisthogyrate with slightly protruding carinae, usually rounded angulations in late growth stages. Escutcheon slightly depressed. Area with fine commarginal costellae, usually densely spaced. Antecarinal space variably developed. Flank with oblique costae, subparallel, anteroventrally sloping, and orthogonal to the antecarinal space. Flank costae sometimes sharp and continuous on the anteriormost segment; otherwise formed by small tubercles, frequently elongate along growth lines.
Occurrence
Sinemurian and Pliensbachian from the Pacific coast of the Americas.
Etymology
The genus is dedicated to Wilhelm Möricke, German paleontologist who described the type species.
Remarks
Moerickella n. gen. is established based on Trigonia infraclavellata Möricke, Reference Möricke1894; the species Trigonia gryphitica Möricke, Reference Möricke1894 is here regarded as synonym of T. infraclavellata. The characters shared with J. siemonmulleri Poulton, Reference Poulton1979, from the late Sinemurian of Nevada (USA) and Early Jurassic of British Columbia (Canada), and with J. supleiensis Poulton, Reference Poulton1979, probably from the late Pliensbachian of Oregon (USA), allow for assignment of the North American species to Moerickella n. gen. Previously discussed characters relating J. burckhardti to F. inexspectata preclude assigning the species here included in Moerickella n. gen. to Jaworskiella.
When discussing the genus Jaworskiella, Poulton (Reference Poulton1979, p. 19–20) provided an emended diagnosis that was partly considered here for the diagnosis of Moerickella n. gen. According to his interpretation, Jaworskiella and the species now included under Moerickella n. gen. show a gradual rotation of the costae over a major part of the flank, while in Myophorella Bayle, Reference Bayle1878 (including Promyophorella) only the posteriormost segments of costae were rotated. This results in subparallel smoothly curved costae departing from the antecarinal space at right angles in Moerickella n. gen., and diverging costae, usually increasing curvature during development, and departing from the antecarinal space at obtuse anterior angles in Promyophorella.
Moerickella n. gen. differs from Jaworskiella by having a much more regular ornamentation pattern (also recognized by Poulton, Reference Poulton1979, p. 20, for M. siemonmulleri n. comb.), with subparallel flank costae bearing well-developed and elongate tubercles on their posterior segments. The presence of an anterior radial ruga in Jaworskiella is the main distinctive character separating it from Moerickella n. gen. and other morphologically convergent forms.
Moerickella infraclavellata (Möricke, Reference Möricke1894) new combination
Figure 13
- * Reference Möricke1894
Trigonia infraclavellata Möricke, p. 46, pl. 2, fig. 3.
- Reference Möricke1894
Trigonia gryphitica Möricke, p. 45, pl. 3, fig. 5.
- non v Reference Burckhardt1902
Trigonia gryphitica; Burckhardt, p. 245, pl. 4, fig. 4.
- non Reference Windhausen1931
Trigonia gryphytica; Windhausen, pl. 22, fig. 7. [reproduced from Burckhardt, Reference Burckhardt1902]
- Reference Pérez and Reyes1977
Jaworskiella infraclavellata; Pérez and Reyes, p. 15, pl. 3, fig. 5. [reproduced from Möricke, Reference Möricke1894]
- Reference Pérez and Reyes1977
Jaworskiella gryphitica; Pérez and Reyes, p. 15, pl. 3, fig. 4. [reproduced from Möricke, Reference Möricke1894]
- Reference Pérez and Reyes1997
Jaworskiella infraclavellata; Pérez and Reyes, p. 574.
- Reference Pérez and Reyes1997
Jaworskiella gryphitica; Pérez and Reyes, p. 574.
- v Reference Pérez, Aberhan, Reyes and von Hillebrandt2008
Jaworskiella gryphitica; Pérez and Reyes in Pérez et al., p. 74, pl. 6, figs. 5, 6, 10, 11.

Figure 13. Moerickella infraclavellata (Möricke) n. comb. from the Early Jurassic of Argentina and Chile. Scale bar = 10 mm. (1) SNGM 779, shell in butterfly position, Sinemurian, Sierra Muñiz, Chile; (2) MLP 18387b, fragmentary left valve, Sinemurian, Codo del Blanco; (3) MLP 32846, fragmentary right valve, Sinemurian, Las Chilcas; (4) MLP 31153, fragmentary left valve, Sinemurian, Las Chilcas; (5) SNGM 778, right valve, Sinemurian, Sierra Muñiz, Chile; (6) MLP 31119a, external mold of a left valve, earliest Pliensbachian, La Horqueta.
Type materials
Möricke (Reference Möricke1894) based his description on two specimens collected by G. Steinmann from beds referred to the Early Jurassic at Las Amolanas, Chile; he only figured an incomplete right valve (Möricke, Reference Möricke1894, pl. 2, fig. 3). We could not trace the whereabouts of his syntypes.
Diagnosis
Small opisthogyrate shell. Escutcheon smooth, with a stepped escutcheon angulation. Large area with commarginal, densely spaced costellae; with submedian groove. Marginal carina tuberculate, slightly protruding at early growth stages, later as a rounded angulation. Antecarinal space smooth and well developed, widening posteriorly. Flank semicircular, with oblique, subparallel costae fragmented into very small and elongate tubercles. Delicate sculpture throughout the shell.
Occurrence
La Horqueta, Codo del Blanco, Las Chilcas, Puesto Araya (Atuel River region), Mendoza Province. Late Sinemurian (Orthechioceras-Paltechioceras Biozone [≈ Raricostatum Biozone]) to early Pliensbachian (M. externum Biozone [≈ Ibex Biozone]; Fig. 3). In Chile, this species occurs in late Sinemurian beds at Río Manflas, Quebrada del Pobre, Sierra de Muñiz, Quebrada de Las Vizcachas, Sierra de la Ternera (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Small, slightly opisthogyrate shell. Escutcheon not preserved in Argentinian specimens, smooth in Chilean material; with slightly protruding escutcheon carina to stepped angulation. Area ~1/3 of general shell surface in Chilean samples (Fig. 13.1, 13.5). Area with commarginal costellae, densely spaced (~12 costellae/cm), sometimes interrupted by a faint groove; costellae on the ventral and dorsal sides of the groove sometimes alternate (Fig. 13.6). Chilean specimens with a submedian groove on the area; dorsal and ventral surfaces of the area converging at the groove. Marginal carina slightly protruding at early growth stages, a rounded angulation later; with well-developed tubercles, usually related to an area costella (only half of the costellae with tubercles; Fig. 13.1). Antecarinal space smooth and well developed, widening posteriorly. Flank semicircular, with oblique, subparallel costae fragmented into very small and elongate tubercles.
Materials
Fifteen specimens; from Argentina: MLP 13354, 18387b, 31118, 31119, 31153, 32846, 32885, 33235; from Chile: SNGM 564–566, 778, 779.
Measurements
MLP 31153: L = 23 mm, H = 21 mm. Some Chilean specimens are more complete; SNGM 778: L = 35, H = 26 mm; SNGM 779: L = 39 mm, H = 29 mm.
Remarks
Moerickella infraclavellata n. comb. differs from other species of Moerickella n. gen. by its: (1) smooth and widening antecarinal space, (2) delicate sculpture, and (3) small size. We regard T. gryphitica Möricke as a subjective synonym of T. infraclavellata Möricke. Because relative page position is not a mandatory criterion when choosing between simultaneously published names, in accordance with ICZN 1999 (Art, 24.2.2 and Rec. 24A), precedence over T. gryphitica is given to T. infraclavellata. Such action best serves nomenclatural stability because the material originally referred to the latter species by Möricke (Reference Möricke1894) is described more accurately (it was probably better preserved, too), providing the diagnostic features recognized here. Pérez and Reyes (Reference Pérez and Reyes1977) and Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) considered both species as distinct and valid; they distinguished them by the presence of more widely spaced flank costae in T. gryphitica (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 75). When describing both species, Möricke (Reference Möricke1894) related T. gryphitica to the old “section Undulatae,” which is characterized by the presence of a bend in the flank costae, although this character in T. gryphitica was not as sharp as in the typical “Undulatae” (Möricke, Reference Möricke1894, p. 45). According to our analysis of the Argentinian and Chilean material, such differences are here regarded as part of the intraspecific variation of a single species.
Moerickella siemonmulleri (Poulton, Reference Poulton1979) n. comb. and M. supleiensis (Poulton, Reference Poulton1979) n. comb. from North America have larger shells with stronger sculpture than M. infraclavellata n. comb., both characters more developed in M. supleiensis n. comb. Moerickella siemonmulleri n. comb. also has an ornamented antecarinal space. In both North American species, flank costae have continuous anterior segments.
Möricke (Reference Möricke1894, p. 46) already recognized M. infraclavellata n. comb. as the oldest species of the “section Clavellatae,” characterized by flanks with nodate curved costae. The transitional character of this species is quite remarkable. The Sinemurian samples seem close to F. eopacifica n. sp., bearing tubercles mostly on the posterior segment of flank costae; the anterior segment being sharp and continuous, like in Frenguelliella (Fig. 13.3). Part of the material illustrated by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, pl. 6, figs. 5, 10, 11) closely resembles these samples. On the other hand, Pliensbachian samples (Fig. 13.6), together with Chilean material under collection numbers SNGM 778, 779 (Fig. 13.1, 13.5), resemble Promyophorella basoaltorum n. sp., with tubercles well developed along the whole costa, but they bear subparallel flank costae orthogonal to the antecarinal space, while P. basoaltorum n. sp. shells have diverging flank costae departing from antecarinal space at markedly obtuse angles anterior to the costae.
Genus Pseudovaugonia new genus
Type species
Vaugonia hectorleanzai Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 91–92, from the late Sinemurian–early Toarcian of Antofagasta and Atacama Regions, Chile.
Other species
Vaugonia vancouverensis Poulton, Reference Poulton1976; Vaugonia oregonensis Poulton, Reference Poulton1979.
Diagnosis
Elongate, opisthogyrate shell; with slightly protruding carinae at early growth stages, rounded angulations at late growth stages. Escutcheon slightly depressed, smooth. Area with fine commarginal costellae, usually densely spaced; costellae wavy or irregular at late growth stages. Sub-median groove developed on the area. Antecarinal space barely developed or absent. Flank with oblique costae, in three sets: posterior one subparallel, anteroventrally sloping and orthogonal to marginal angulation; medium set sub-horizontal to slightly anterodorsally directed; anterior set short and orthogonal to anterior margin. Posterior and median sets of costae meeting towards mid-flank; median and anterior sets of costae meeting along a radial anterior line. Anterior and median sets of costae replaced by thin commarginal costae at late growth stages. Flank costae (especially the posterior set) with small, irregular tubercles, sometimes barely developed or fading away.
Occurrence
Sinemurian to Toarcian from the Pacific coast of the Americas.
Etymology
The Greek prefix pseudo- meaning false, and -vaugonia for the morphologically similar genus: “False Vaugonia.”
Remarks
Pseudovaugonia hectorleanzai n. comb. shares with other Myophorelloidea the area ornamentation pattern, with commarginal costellae and a sub-median radial groove. The posterior set of flank costae, orthogonal to the marginal angulation, relates it to M. infraclavellata n. comb. Vaugonia oregonensis, from late Pliensbachian beds of Oregon (Poulton Reference Poulton1979, p. 23, figs. 10.14–10.19), shows strong affinities with P. hectorleanzai n. comb., as already recognized by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), and hence it is included in the new genus. Vaugonia vancouverensis, from the Pliensbachian? of Vancouver Island, British Columbia (Poulton Reference Poulton1976, p. 48, pl. 8, figs. 13–20), shows most of the characters of Pseudovaugonia n. gen. (area with submedian groove, posterior set of costae coarser, with tubercles and almost perpendicular to the antecarinal space). There is no clear indication of an anterior set of costae at early growth stages, but Poulton (Reference Poulton1979, p. 23) stated that juveniles of V. oregonensis and V. vancouverensis may be indistinguishable.
The species included in Pseudovaugonia n. gen. were originally referred to the genus Vaugonia (Poulton, Reference Poulton1976, Reference Poulton1979; Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) due to the V-shape of flank costae. We regard this shared character as convergent and consider Pseudovaugonia n. gen. as closely related to Moerickella n. gen., and thus a member of the Myophorellidae.
The genus Vaugonia Crickmay, Reference Crickmay1930 was assigned to the old “section Undulatae,” which includes most species with a V or L pattern of flank costae (e.g., Kobayashi and Mori, Reference Kobayashi and Mori1955, p. 77). The holotype of the type species of Vaugonia, V. veronica Crickmay, Reference Crickmay1930, from the Middle Jurassic of British Columbia (GSC27715, a cast of which, under number SNGM 7530, was studied at the SERNAGEOMIN, Chile; Fig. 14.1), has a commarginally ornamented area divided in two, although instead of a groove, the division is formed by a small step, with the dorsal part at a lower level than the ventral part. This step is what Crickmay (Reference Crickmay1930), in his original diagnosis for Vaugonia, characterized as a median carina with a median furrow along the dorsal side. No tubercles are found on flank costae in V. veronica; umbonal costae are sub-commarginal, later developing a bend, which finally results in a subdivision of costae into two sets: a sub-vertical posterior one and a sub-horizontal to sub-commarginal anterior one. These two sets meet at right angles on the central to posterior flank, generating an L pattern rather than a V pattern (Fig. 14.1).

Figure 14. Vaugonia species from Japan and North America. Scale bar = 10 mm. (1) Vaugonia veronica Crickmay, cast of the holotype (SNGM 7530), Middle Jurassic, British Columbia, Canada; (2) Vaugonia yokoyamai var. gracilis Kobayashi and Mori, external cast of a left valve (SNGM 7535), Hettangian, Shizukawa area, Japan; (3) Vaugonia niranohamensis Kobayashi and Mori, external cast of the holotype, a left valve (SNGM 7523), Hettangian, North coast of Bentenzaki, Japan.
The oldest known species assigned to Vaugonia are V. niranohamensis Kobayashi and Mori, Reference Kobayashi and Mori1955 (Fig. 14.3), V. yokoyamai Kobayashi and Mori, Reference Kobayashi and Mori1955 (including its variety gracilis; Fig. 14.2), V. namigashira Kobayashi and Mori, Reference Kobayashi and Mori1955, and V. kojiwa Kobayashi and Mori, Reference Kobayashi and Mori1955, from the Hettangian of Japan. Vaugonia niranohamensis bears a smooth area (Fig. 14.3), while the remaining three species bear commarginal costellae, like in V. veronica (Fig. 14.1, 14.2). All of them bear a radial sub-median step in the area, like the one found in V. veronica, but most of them (with the exception of V. kojiwa) also bear small rounded tubercles (Fig. 14.2, 14.3).
The shared characters of V. veronica with the Hettangian Japanese species suggest a true phylogenetic relationship among them. Consequently, we consider as diagnostic features of the genus Vaugonia (1) a V or L flank costae pattern together with (2) a radial sub-median step in the area, with the dorsal part depressed relative to the ventral part. Its area can be smooth or commarginally ornamented. A thorough revision of the genus, far beyond the scope of this paper, would be necessary to determine if such variations represent two evolutionary lineages or else iterative features. The presence of tubercles and a widening antecarinal space seem to be primitive characters within Vaugonia, both subsequently lost in the type species.
Considering this characterization, the genus Vaugonia seems to be absent from the Lower Jurassic of South America. Specimens attributed to V. niranohamensis and V. yokoyamai were mentioned and described from the Batá Formation from Colombia (Geyer, Reference Geyer1973, p. 77–79), originally thought to be Jurassic in age, but were later assigned to the genus Syrotrigonia Cox, Reference Cox1952 (Etayo-Serna et al., Reference Etayo-Serna, Solé De Porta, De Porta and Gaona2003; Echevarría et al., Reference Echevarría, Damborenea and Manceñido2015), and their age was established as Valanginian–Hauterivian (Etayo-Serna et al., Reference Etayo-Serna, Solé De Porta, De Porta and Gaona2003). Vaugonia sp. 1 and Vaugonia sp. 2 described by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), from the Toarcian (sp. 1) and Pliensbachian (sp. 2) of Atacama, have a flat area, suggesting they are not genuine representatives of Vaugonia sensu stricto. Some of the species described by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) within Vaugonia may be better placed in Promyophorella (see Remarks under Promyophorella). Kobayashi (in Kobayashi and Mori, Reference Kobayashi and Mori1955, p. 74), when revising the Vaugoniinae, considered Vaugonia and Myophorella as belonging to two independent lineages. Such a claim is strongly supported by the diverse Hettangian record of Vaugonia in Japan (Kobayashi and Mori, Reference Kobayashi and Mori1955) on one hand, and, on the other, by the Sinemurian–Pliensbachian phylogenetic series presented here, comprising Frenguelliella eopacifica n. sp., Moerickella infraclavellata (Möricke, Reference Möricke1894) n. comb., and Promyophorella basoaltorum n. sp.
Pseudovaugonia n. gen. and Moerickella n. gen. share many characters (compare Fig. 15.7 with Fig. 13.1, 13.5): the escutcheon is smooth; the area bears commarginal costellae and a median groove (instead of a step); the posterior set of costae departs from the antecarinal space almost at right angles (whereas this angle is obtuse in Vaugonia, Fig. 14.1–14.3); flank costae carry similar irregular elongate tubercles, suggesting that the former evolved from the latter rather than being closely related to Vaugonia. The main morphological changes from Moerickella n. gen. to Pseudovaugonia n. gen. are: (1) development of the anterior and median sets of flank costae, resulting in the V pattern; (2) the reduction of the antecarinal space; and (3) the wavy or irregular costellae on the posterior part of the area (Fig. 15.7).

Figure 15. Pseudovaugonia hectorleanzai (Pérez and Reyes in Pérez et al.) n. comb. from the Early Jurassic of Argentina and Chile. Scale bar = 10 mm. (1, 4) MLP 36322, right valve preserved as internal mold and external mold with shell remains, Toarcian, Arroyo Lapa; (2, 3, 5) MLP 36321a–c, Toarcian, Arroyo Lapa; (2) fragmentary composite mold of left valve; (3) fragmentary external mold of right valve; (5) fragmentary composite mold of left valve; (6) MLP 36324, external mold of a left valve, Toarcian, Las Overas; (7) SNGM 644, right valve, Sinemurian, Sierra Limón Verde, Chile; (8) holotype, SNGM 631, shell, left lateral view, Toarcian, Quebrada Chanchoquín, Chile.
Pseudovaugonia hectorleanzai (Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008)
Figure 15
- v Reference von Hillebrandt and Zeil1980
Vaugonia n. sp.; Hillebrandt, pl. 2, fig. 1.
- *v Reference Pérez, Aberhan, Reyes and von Hillebrandt2008
Vaugonia hectorleanzai Pérez and Reyes in Pérez et al., p. 91, pl. 13, fig. 9, pl. 15, figs. 3, 5–11, pl. 16, figs. 1, 3, 8, 9, 11, 12, 14, 15.
- v Reference Pérez and Reyes2008
Vaugonia hectorleanzai; Pérez and Reyes, p. 28.
Type material
Holotype: SNGM 631, from the early Toarcian of Quebrada Chanchoquín/Paitepén, Atacama, Chile, figured by Hillebrandt (Reference von Hillebrandt and Zeil1980, pl. 2, fig. 1), Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, pl. 15, fig. 7), and Figure 15.8.
Diagnosis
Medium-sized, opisthogyrate, elongate shell. Area narrow with commarginal costellae, irregular or geniculate towards the posterior end; submedian groove coincident with the inflection of posterior costellae. Antecarinal space barely developed. Anterior radial ruga developed on flank. Flank costae added from the antecarinal space (posterior set of strong, tuberculate costae) and the anterior radial ruga (anterior and median sets of smooth, narrow costae). Posterior and median sets meeting at mid-flank in a V pattern at early stages (more pronounced at late juvenile stages), or in an L pattern (adult stages).
Occurrence
Las Overas (West of Malargüe region), Mendoza Province; Arroyo Lapa, central Neuquén Province; early Toarcian (D. hoelderi Biozone [≈ Serpentinum Biozone]) (Fig. 3). Also probably at Arroyo La Laguna (Los Patos region), San Juan Province; Pliensbachian or Toarcian. The species is also found in beds referred to the late Sinemurian and early Toarcian of Antofagasta and Atacama in Chile (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008).
Description
Medium-sized, opisthogyrate, elongate shell. Escutcheon not preserved. Area narrow, with transverse commarginal costellae, somewhat irregular or geniculated towards the posterior end (Fig. 15.5); with a submedian groove coincident with the inflection of posterior costellae. Marginal carina slightly protruding at early growth stages (Fig. 15.2, 15.6), an angulation at late stages (Fig. 15.5, 15.6), with small tubercles at the intersection with area costellae (Fig. 15.3). Flank with narrow, barely developed antecarinal space (Fig. 15.3, 15.6). Flank with oblique costae showing a V-shaped pattern. New flank costae added from two radial elements: the antecarinal space and the anterior radial ruga (Fig. 15.2, 15.6, 15.7). Two branches added on the anterior radial ruga during shell growth: one anteroventrally directed (anterior set), the other posteroventrally (median set). Ventrally directed branch (posterior set) added adjacent to antecarinal space. Costae of posterior and median sets meeting at mid-flank. Anterior and median sets sub-commarginal and continuous in late growth stages.
Materials
One right valve preserved as internal and external molds with shell fragments: MLP 36322; fragmented composite and external molds MLP 36321, 36323, 36324. MLP 36398, a poorly preserved shell, most likely belonging to this species. Specimens from Chile SNGM 630–644, 673.
Measurements
MLP 36322: L = 51 mm, H = 32 mm; holotype (SNGM 631; slightly broken posteriorly): L = 40 mm, H = 28 mm.
Remarks
The main characteristics of the Argentinian material agree with those described and illustrated by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) as Vaugonia hectorleanzai from the late Sinemurian–early Toarcian of Chile. Pseudovaugonia hectorleanzai n. comb. can be distinguished from other Pseudovaugonia n. gen. species by: (1) the presence of a radial anterior ruga, and (2) the presence of tubercles on the posterior set of flank costae. As pointed out by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), the species Pseudovaugonia oregonensis (Poulton, Reference Poulton1979) n. comb. from the late Pliensbachian of Oregon (USA) is very similar to P. hectorleanzai n. comb., but differs by lacking tubercles on the posterior set of flank costae. Pseudovaugonia vancouverensis (Poulton, Reference Poulton1976) n. comb., from the (probable) Pliensbachian of Vancouver Island (Canada), is only incipiently tuberculate and has a more delicate sculpture. No anterior radial ruga is mentioned for any of the North American species.
Trigonia substriata Giebel in Burmeister and Giebel, Reference Burmeister and Giebel1861, from the Early–Middle Jurassic of Juntas, Chile, shares with Pseudovaugonia hectorleanzai n. comb. an early development of V-shaped flank costae. Nevertheless, according to the figure provided in Burmeister and Giebel (Reference Burmeister and Giebel1861, pl. 2, fig. 4), the posterior set of flank costae departs from the antecarinal space at obtuse anterior angles (like in Promyophorella), and according to his descriptions, the area in T. substriata is smooth or with very fine transverse wrinkles, unlike the costellae observed in P. hectorleanzai n. comb.
Genus Promyophorella Kobayashi and Tamura, Reference Kobayashi and Mori1955
Type species
Myophorella (Promyophorella) sigmoidalis Kobayashi and Tamura, Reference Kobayashi and Tamura1955, pl. 5, figs. 1–3; Hayami, Reference Hayami1975, pl. 5, fig. 9. Bajocian, Miyagi Prefecture (Japan).
Diagnosis
Slightly to clearly opisthogyrate shell; typically crescentic. Well-defined prominent tuberculate carinae at early growth stages, sharp tuberculate angulations at late growth stages. Area surface variably developed, with thin commarginal costellae, sometimes fading at late growth stages; with median groove. Flank with narrow oblique and diverging costae carrying small numerous regular tubercles aligned along their tops; costae generating a clearly obtuse anterior angle with the antecarinal space. Antecarinal space smooth. Escutcheon smooth or with transverse pustulose costellae.
Remarks
When analyzing the variability within the genus Myophorella Bayle, Reference Bayle1878 (pl. 120), Kobayashi and Tamura (Reference Kobayashi and Tamura1955) defined the subgenus Promyophorella for those species with narrow slender costae bearing numerous small tubercles on top. Myophorella s.s. differs from Promyophorella by its subtriangular, slightly opisthogyrate to orthogyrate, large shell, and its stout flank costae with large button-like tubercles. Kobayashi and Tamura (Reference Kobayashi and Tamura1955) considered Promyophorella as ancestral to Myophorella s.s., but they did not regard them as different genera because they recognized numerous intermediate forms. This transition might have occurred iteratively by heterochrony, with the general morphology characteristic of Myophorella s.s. representing a paedomorphic state (Echevarría, Reference Echevarría2016). The diagnosis provided here is a modification from that of Kobayashi and Tamura (Reference Kobayashi and Tamura1955), stressing the main differences from Myophorella s.s., but also from Moerickella n. gen., which differs from Promyophorella by its subparallel smoothly curved flank costae orthogonal to the antecarinal space.
Fleming (Reference Fleming1987) followed Kobayashi and Tamura (Reference Kobayashi and Tamura1955), who listed Scaphogonia argo Crickmay, Reference Crickmay1930, type species of Scaphogonia, as a species of Myophorella (Promyophorella), and considered Promyophorella as a junior synonym of Scaphogonia.
We regard Scaphogonia as a separate genus, related to Promyophorella, but differing from it by its flank ornamentation pattern formed by two sets of ornamentation: a main set formed by nodate costae departing from the area at obtuse anterior angles and roughly orthogonal to the ventral margin; and an anterior set of nodate costae orthogonal to the anterior margin that do not meet the costae of the posterior set.
When characterizing the genus Scaphogonia, Crickmay (Reference Crickmay1930, p. 51) also defined the genus Haidaia Crickmay, with Trigonia dawsoni Whiteaves, Reference Whiteaves1878, as type species. Kobayashi and Tamura (Reference Kobayashi and Tamura1955) considered Haidaia as a third subgenus in their conception of Myophorella, being characterized by narrow ridges extending ventrally from tubercles on flank costae (Haidaia-like ornamentation in the sense of Leanza, Reference Leanza1993, p. 60). Although we regard this particular kind of ornamentation as a significant character within the Myophorellidae, we consider it unlikely to characterize a particular phylogenetic lineage within the family because in some of the early species here described it seems to be already outlined. Poulton (Reference Poulton1979, p. 33–34), when describing Trigonia dawsoni based on new, abundant, and well-preserved material, found that the discrepant anterior ornamentation may also be present in many shells of this species, so we regard Haidaia as junior synonym of Scaphogonia.
As remarked under Pseudovaugonia hectorleanzai n. comb., the L or V patterns of flank costae might have appeared within Promyophorella convergently with that observed in Vaugonia. We consider that the material assigned by Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008 to Vaugonia cf. V. gottschei (Möricke, Reference Möricke1894) from Atacama (Pliensbachian?) would be better placed under Promyophorella, close to P. exotica. Vaugonia cf. V. substriata (Burmeister and Giebel, Reference Burmeister and Giebel1861) Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), from Atacama (middle Toarcian), is also assigned to Promyophorella, showing a similar ornamentation pattern to that of P. reginae (Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008). In both cases, the flank ornamentation pattern is relatively simple, and the area shows characters typical of early species of Promyophorella (commarginal area ornamentation with a submedian groove, tuberculate marginal angulation and a widening antecarinal space). Most of the Jurassic species described as Vaugonia (Vaugonia) from Antarctica (V. [V.] orvillensis, V. [V.] sp.; Kelly, Reference Kelly1995) and New Zealand (V. [V.] spedeni, V. [V.] kawhiana, V. [V.] sp. A, V. [V.] sp. B; Fleming, Reference Fleming1987) are actually better placed within the Promyophorella lineage, revealing the strong southern diversification of the group during the Middle and Late Jurassic. The exception is Vaugonia kahuika Fleming, Reference Fleming1987, from the lower Temaikan (≈ Aalenian) from New Zealand, which is actually referable to the Vaugoniidae.
The genus Orthotrigonia Cox, Reference Cox1952, originally proposed as a subgenus of Myophorella and later (Cox in Cox et al., Reference Cox, Newell, Boyd, Branson, Casey, Chavan and Coogan1969) considered a subgenus of Vaugonia, is close to Promyophorella, sharing with it the area with commarginal ornamentation and a submedian groove, and tuberculate marginal carina/angulation. Orthotrigonia differs from Promyophorella by its flank ornamentation pattern, with V-shaped or L-shaped costae at early growth stages and steep, straight costae bifurcating anteriorly at late growth stages.
The genus Scaphorella Leanza, Pérez, and Reyes, Reference Leanza, Pérez and Reyes1987, also shows affinities with Promyophorella, both sharing an area with thin commarginal costellae and a median groove, and flank costae with small tubercles on top. The species S. susanae Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, from the latest Toarcian and early Aalenian of Atacama, Chile, even has a smooth antecarinal space, widening posteriorly, although this character is absent in the type species, S. leanzai (Lambert, Reference Lambert1944). Scaphorella differs from Promyophorella by its two sets of flank costae: a posterior one of almost straight costae, orthogonal to the ventral margin, and an anterior one of arcuate costae, reaching the anterior margin at right angles; both sets meet at the mid-anterior flank, frequently with two anterior costae meeting a posterior one (bifurcating flank costae). This ornamentation pattern is very similar to that found in Scaphogonia, being the only difference the continuity of anterior and posterior costae in Scaphorella. A revision of the Middle Jurassic species involved is beyond the scope of this paper; for the time being, we tentatively consider them as separated taxa.
Promyophorella basoaltorum new species
Figure 16.1–16.24, 16.27, 16.28
- ? Reference Behrendsen1891
Trigonia substriata Giebel; Behrendsen, p. 387.
- ? Reference Behrendsen1922
Trigonia substriata Giebel; Behrendsen, p. 172.
- v Reference Carral-Tolosa1942
Trigonia exotica Steinmann [sic]; Carral Tolosa, p. 56, pl. 6, figs. 1a–c.
- v Reference Volkheimer, Manceñido and Damborenea1978a
Myophorella cf. M. araucana (Leanza); Volkheimer et al., p. 304.
- v Reference Volkheimer, Manceñido and Damborenea1978b
Myophorella cf. M. araucana (Leanza); Volkheimer et al., p. 212 (table 2).
- v Reference Volkheimer, Manceñido and Damborenea1978b
Myophorella? sp.; Volkheimer et al., p. 212 (table 2).
- v Reference von Hillebrandt and Zeil1980
Myophorella catenifera (Hupé); Hillebrandt, pl. 2, fig. 4.
- v Reference von Hillebrandt and Schmidt-Effing1981
Myophorella catenifera (Hupé); Hillebrandt and Schmidt-Effing, p. 10.
- Reference Pérez1982
Myophorella (Myophorella) araucana (Leanza); Pérez, pl. 14, fig. 9.
- v Reference Cuerda, Schauer and Sunesen1982
Myophorella araucana (Leanza); Cuerda et al., p. 331.
- ? Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983
Myophorella (Promyophorella) sp. indet.; Ishikawa et al., p. 41, pl. 1, fig. 8.
- vp Reference Leanza, Garate-Zubillaga and Volkheimer1987
Myophorella (Myophorella) araucana (Leanza); Leanza and Garate-Zubillaga, p. 211, pl. 1, figs. 6, 7, but not fig. 8.
- v Reference Leanza1993
Myophorella (Myophorella) catenifera (Hupé); Leanza, p. 29, pl. 2, figs. 10, 11.
- Reference Leanza1993
Myophorella (Myophorella) cf. M. tuberculata (Agassiz); Leanza, p. 29, pl. 1, fig. 9.
- v Reference Pérez and Reyes2008
Myophorella (Myophorella) araucana (Leanza); Pérez and Reyes in Pérez et al., p. 82, pl. 11, figs. 1, 4, 5, 8, pl. 12, figs. 2–7, 9, 11, 12, pl. 13, figs. 1, 7.
- v Reference Echevarría2016
Myophorella araucana? (Leanza); Echevarría, p. 179, figs. 1d, e, m, 3a, 6d, 10.

Figure 16. Promyophorella species from the Early Jurassic of Argentina and Chile. Scale bar = 10 mm. (1–24, 27, 28) Promyophorella basoaltorum n. sp. (1) SNGM 609, right valve, Pliensbachian, Quebrada Asientos, Chile; (2) SNGM 600, right valve, Pliensbachian, Quebrada Asientos, Chile; (3) SNGM 604, shell, left lateral view, Pliensbachian, Quebrada Asientos, Chile; (4) SNGM 605, left valve, Pliensbachian, Quebrada Asientos, Chile; (5) MLP 36370, composite mold of right valve, Pliensbachian, Arroyo La Laguna; (6) MLP 28761a, composite mold of right valve, Pliensbachian, Arroyo Serrucho; (7) IANIGLA-PI 3273, external mold of right valve, Pliensbachian, Arroyo Serrucho; (8) IANIGLA-PI 3276, composite mold of shell in butterfly position, Pliensbachian, Arroyo Serrucho; (9) IANIGLA-PI 3250, composite mold of right valve, Pliensbachian, Arroyo Serrucho; (10) IANIGLA-PI 3235, external mold of left valve, Pliensbachian, Arroyo Serrucho; (11) holotype, IANIGLA-PI 3230, external mold of a right valve, Pliensbachian, Arroyo Serrucho; (12) MLP 28761b, composite mold of left valve, Pliensbachian, Arroyo Serrucho; (13) MLP 36383, composite mold of left valve, Pliensbachian, Cerro Puchenque; (14) MLP 36388a, right valve, Pliensbachian, Cerro Tricolor; (15) MLP 36388b, left valve, Pliensbachian, Cerro Tricolor; (16) MLP 15322, external mold of left valve, Pliensbachian, South from Rahue-Aluminé; (17) MLP 36389, composite mold of right valve, Pliensbachian, Arroyo Lonqueo; (18) MLP 36394, composite mold of right valve, Subida a Sañicó; (19–21) paratype, MLP 6725, complete shell, right lateral, anterior and dorsal views, Pliensbachian, Cerro Roth; (22, 23) MPEF-PI 6386a, b, fragmentary right valves, late Pliensbachian–early Toarcian, Nueva Lubecka; (24) MPEF-PI 6399, fragmentary left valve, late Pliensbachian–early Toarcian, Nueva Lubecka; (27) MPEF-PI 6392, left valve, late Pliensbachian–early Toarcian, Nueva Lubecka; (28) MPEF-PI 6390, anterior part of left valve, late Pliensbachian–early Toarcian, Nueva Lubecka; (25, 26, 29) Promyophorella aff. P. basoaltorum n. sp., CPBA 19657, external molds preserved on a single slab, early Pliensbachian, Cerro Cuche: (25) CPBA 19657c, detail of an external mold of right valve; (26) rock surface showing several external molds; (29) CPBA 19657a, detail of an external mold of left valve.
Type materials
Holotype: IANIGLA-PI 3230, right valve external mold from the Pliensbachian of Arroyo Serrucho (Fig. 16.11); paratype MLP 6725, a well-preserved shell with both valves from the Pliensbachian of Cerro Roth (Fig. 16.19–16.21).
Diagnosis
Opisthogyrate, crescentic, slightly rostrate shell, with semicircular anteroventral margin. Escutcheon with transverse costellae (oblique to growth lines) variably developed. Area wide with commarginal costellae. Both carinae prominent at early growth stages, sharp angulations at late stages. Flank with arcuate diverging costae with small tubercles along their tops.
Occurrence
Arroyo La Laguna (Los Patos region), San Juan Province; Puesto Araya (Atuel River region), Arroyo Peuquenes, Arroyo Portezuelo Ancho, Portezuelo Ancho, Santa Elena, La Bajada (Las Leñas/Valle Hermoso region), Arroyo Serrucho, Cerro Puchenque (west of Malargüe region), Cerro Tricolor (Grande River region), Mendoza Province; Ñiraico, Estación Rajapalo (Cordillera del Viento region), 13.5 km south of Rahue-Aluminé, Lonqueo (Central Neuquén region), Estancia Santa Isabel, Subida a Sañicó, Cerro Roth Sur (Piedra Pintada region), Neuquén Province; Nueva Lubecka (Genoa River region), Chubut Province. Early Pliensbachian (M. externum or E. meridianus Biozone [≈ Jamesoni/Ibex Biozone]) to early Toarcian (Tenuicostatum Biozone; dubious specimens in D. hoelderi Biozone [≈ Serpentinum Biozone]) (Fig. 3). In Chile, the species was found in Quebrada Asientos, Atacama (Pérez and Reyes, Reference Pérez and Reyes1977; Pérez, Reference Pérez1982; Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008 under Myophorella [Myophorella] araucana Leanza, Reference Leanza1942) in the late Pliensbachian. A very small shell from the Early Jurassic of southern Peru (Ishikawa et al., Reference Ishikawa, Maeda, Kawabe and Rangel Zavala1983) agrees in most general characters with this species.
Description
Small, clearly opisthogyrate shell; crescentic, sometimes slightly rostrate. Semicircular antero-ventral margin; straight and wide posterior margin, with area occupying ~1/3 of shell surface. Escutcheon margin slightly concave, with the umbo and the posterodorsal angle slightly projecting dorsally. Escutcheon with transverse costellae (oblique to growth lines) (Fig. 16.11, 16.21) continuous with those on the area. Escutcheon carina prominent at early growth stages, later becoming a sharp angulation, with small pustules at its intersection with costellae (Fig. 16.7, 16.10, 16.11, 16.21). Area with commarginal costellae, evenly arranged (8–14 costellae/cm) in early growth stages, later replaced by irregular and fine ridgelets (Fig. 16.13, 16.18, 16.19). Sub-median groove (slightly displaced dorsally) in the area (Fig. 16.11, 16.21), sometimes barely impressed. Prominent marginal carina at early growth stages, sharp angulation at late stages; with small pustules at its intersection with costellae (swellings of the costellae at late growth stages) (Fig. 16.4, 16.10–16.12, 16.16). Antecarinal space smooth, widening posteriorly (Fig. 16.1–16.13, 16.16), crossed by flank costae at very early growth stages. First two flank costae sub-commarginal, later on, oblique to growth lines, evenly curving anteriorwards; in some specimens, initial segment of the costae slightly curved, increasing curvature during ontogeny (Fig. 16.5–16.7). Flank costae bearing small tubercles along top. Flank costae departing from the antecarinal space at obtuse anterior angles and diverging during growth.
Internal radial ridge in the posterior part of the inner surface, perpendicular to area margin (Fig. 16.17). Posterior adductor muscle scar rounded, dorsally located towards mid-length, half underneath the area, and half underneath the escutcheon. Nymph short (Fig. 16.21).
Etymology
Dedicated to the Basoalto family from Arroyo Serrucho (type locality of the species), for their warm hospitality and invaluable help to geologists and paleontologists through many years.
Materials
The types plus ~125 specimens: IANIGLA-PI 3216–3253, 3262, 3265–3271, 3272A, 3273–3285, 3286A, 3287, 3289–3291, 3292A, 3293–3296, 3303, 3304, 3318, 3350, 3351; MPEF-PI 2907, 2909, 2965, 3158, 3375, 6386, 6390, 6392, 6394, 6396, 6399; MLP 6253 (part), 15322, 15437, 15535, 15554, 17148, 17503, 28761, 33221, 36368–36394; MOZ-PI 1487, 2682. Specimens from Chile: SNGM 596–616.
Measurements
Holotype (IANIGLA-PI 3230): L = 25 mm, H = 21 mm; paratype (MLP 6725): L = 29 mm, H = 23 mm, W = 16 mm; MLP 36370: L = 25 mm, H = 20 mm; MLP 15322: L = 42 mm, H = more than 27 mm; MLP 36394: L = 34 mm, H = 27 mm; MLP 28761: L = 28 mm, H = 20 mm; the remaining measurements are plotted on Figure 17.

Figure 17. Scatter plot of height vs. length for Promyophorella basoaltorum n. sp.
Remarks
Promyophorella basoaltorum n. sp. is the earliest species of Promyophorella and is most likely the ancestral stock of all younger species, but the genus is in need of a revision in order to understand its later diversification. Some of the variability within this species relates to flank ornamentation. Most specimens bear evenly curved costae (Fig. 16.5, 16.11, 16.19), while in others, the costae have an almost straight initial portion and a sudden increase in curvature (J pattern) (Fig. 16.2, 16.6). Usually costae are widely spaced (Fig. 16.11, 16.19, 16.28), whereas some shells show closely spaced costae (Fig. 16.6). Shells are typically small, on occasion, medium-sized (Fig. 16.1, 16.2, 16.16). Elongation is also variable within the species. Although the more typical form has a semicircular antero-ventral margin (Fig. 16.3, 16.4, 16.12, 16.13, 16.19), some shells are more elongate (Fig. 16.6–16.8, 16.17), with the ventral margin longer and slightly less convex than the anterior margin.
The specimens referred by Pérez and Reyes (in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) to Myophorella araucana are included here in P. basoaltorum n. sp. That material is also highly variable; some shells are semicircular in their antero-ventral margin, while others are more elongate and somewhat rostrate. Flank costae may show a variable pattern (of homogeneous curvature or more J-shaped), but they always diverge clearly. Escutcheon usually has transverse costellae variably developed: from small thorn-like projections developing from the escutcheon carina pustules, to well-developed costellae through entire escutcheon surface.
This taxon is one of the most common trigonioids from the Lower Jurassic of southern South America, having been frequently assigned to closely related but significantly different species. Behrendsen (Reference Behrendsen1891, Reference Behrendsen1922) referred (but did not figure) some Early Jurassic material from Portezuelo Ancho to T. substriata, which may belong here based on specimens collected at the same locality.
Material referable to P. basoaltorum n. sp. was referred by Carral-Tolosa (Reference Carral-Tolosa1942) to Trigonia exotica Möricke, Reference Möricke1894 (she erroneously cited Steinmann as author), but T. exotica differs from P. basoaltorum n. sp. by having strongly bent flank costae, usually with anteriorly elongate tubercles on the ventral portion. Many specimens referred in the literature to “Myophorella araucana (Leanza, Reference Leanza1942)” (e.g., Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008), belong in P. basoaltorum n. sp. It should be noted that specimen MLP 6725 (now paratype of P. basoaltorum n. sp.) was labelled as Trigonia exotica in the MLP collection, and though it was available to Leanza, it was excluded from the type series of P. araucana. Early Jurassic material from many localities in Chile and Argentina (Hillebrandt, Reference von Hillebrandt and Zeil1980; Hillebrandt and Schmidt-Effing, Reference von Hillebrandt and Schmidt-Effing1981; Leanza, Reference Leanza1993) referred to Trigonia catenifera Hupé, Reference Hupé and Gay1854, is here included in P. basoaltorum n. sp.
Promyophorella basoaltorum n. sp. differs from other Promyophorella species by its: (1) clearly opisthogyrate shell shape, (2) relatively large area surface, and (3) simple (without bifurcations or disruptions) flank costae, evenly curving anteriorwards or with increasing curvature, but never bending.
Promyophorella? catenifera (Hupé, Reference Hupé and Gay1854) was first described from Coquimbo, Chile, from beds then referred to the Middle Jurassic (Hupé, Reference Hupé and Gay1854, p. 328), but later assigned to the Early Jurassic (Pérez and Reyes, Reference Pérez and Reyes1977, annex I, p. 45). The material described and figured by Hupé is deposited in the Museum National d'Histoire Naturelle in Paris: MNHN-GG2005-16393 (Fig. 18.1–18.4). No holotype was fixed by Hupé (Reference Hupé and Gay1854), so specimen GG2005-16393b (Fig. 18.1) is here designated as lectotype. The species differs from P. basoaltorum n. sp. by having bifurcated costae towards the anteroventral margin (Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 86, under remarks for Myophorella reginae; Fig. 18.3), sometimes even bifurcating twice (e.g., Fig. 18.3). This character was not mentioned in Hupé's description of the species and is poorly rendered in his figure (Hupé, Reference Hupé and Gay1854, p. 328, pl. 5, fig. 8), and may also relate it to the genus Scaphorella (see Remarks under Promyophorella? sanjuanina n. sp.). Pérez and Reyes (Reference Pérez and Reyes1977, p. 12–13, table 4) and Reyes and Pérez (Reference Reyes and Pérez1979, p. 18, table 2) considered that P.? catenifera was present in the Pliensbachian–Toarcian of Atacama and Coquimbo regions of Chile. Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) maintained this temporal and geographical distribution (and they assigned the material from the type locality, Doña Ana, to the Toarcian), but they neither discussed nor illustrated the species.

Figure 18. Promyophorella? catenifera (Hupé), MNHN-GG2005-16393 (photographs courtesy of C. Noyes), Lower Jurassic, Coquimbo, Chile. Scale bar = 10 mm. (1) Lectotype, MNHN-GG2005-16393b, left valve (antero-ventrally broken); (2) MNHN-GG2005-16393a, left valve; (3, 4) MNHN-GG2005-16393c, right valve, antero-ventral and lateral views.
Trigonia signata Agassiz var. keideli Weaver, Reference Weaver1931, from Early Jurassic deposits of Catan Lil, in Neuquén, seems most likely related to, if not synonymous with, P. basoaltorum n. sp. The area/flank surface ratio, the diverging flank costae, the broad, smooth antecarinal space, are all well described by Weaver (Reference Weaver1931, p. 235–236), being the main difference the apparently less opisthogyrate shell shape (subquadratic shell with almost straight escutcheon margin, according to Weaver), together with the larger size (Weaver's specimen is 53 mm long and 34 mm high). Unfortunately, Weaver (Reference Weaver1931) did not illustrate it, and the type specimen for the variety could not be found (E. Nesbitt, personal communication, 2019). Moreover, in the same work, Weaver also established and figured Trigonia literata Young and Bird var. keideli Weaver (Reference Weaver1931, p. 239–240, pl. 20, fig. 102), which is a primary homonym (ICZN 1999, Art, 57.2), and comes from the Middle Jurassic of central Neuquén. Since both species-group names “keideli” were published exactly on the same date, Art. 24.2.2 applies to solve this previously unnoticed homonym conflict. As first revisers, we consider that precedence should be given to Trigonia literata var. keideli for the sake of nomenclatural stability. Notice that this Middle Jurassic taxon was redescribed by Leanza and Garate-Zubillaga (Reference Leanza and Garate-Zubillaga1983) as Anditrigonia keideli (Weaver, Reference Weaver1931), and was subsequently designated as type species of the subgenus Eoanditrigonia Leanza (Reference Leanza1993, p. 46). Therefore, the species-group name keideli Weaver (Reference Weaver1931, p. 235) becomes permanently invalid and should not be used for this Early Jurassic myophorelloid species.
Further, the description provided by Weaver (Reference Weaver1931, p. 238–239) for other material from the lower Middle Jurassic of southern Mendoza assigned to Trigonia signata, suggests a taxon closely related to P. basoaltorum n. sp.; unfortunately, no figure was provided.
Promyophorella araucana (Leanza, Reference Leanza1942, p. 162–164, pl. 6, figs. 4–6), from the late Pliensbachian of Neuquén and Chubut provinces, differs from this species by its less opisthogyrate shell shape and relatively smaller area. Promyophorella exotica (Möricke, Reference Möricke1894, p. 49, pl. 1, fig. 9, pl. 6, fig. 9), from the Aalenian and Bajocian of Atacama and Valparaíso, Chile (Pérez and Reyes, Reference Pérez and Reyes1977) differs from P. basoaltorum n. sp. by the presence of an inflection in flank costae at late growth stages, resulting in an anterior portion of the costae being commarginal and with weak and elongate tubercles. Promyophorella reginae (Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, p. 86–88, pl. 12, fig. 8, pl. 14, figs. 1, 3–6, 8), from the middle and late Toarcian of Atacama, Chile, differs from P. basoaltorum n. sp. by its less opisthogyrate shell shape and an anterior bending developed on flank costae at late growth stages.
The type species, P. sigmoidalis from the late Aalenian–early Bajocian of Japan, differs from P. basoaltorum n. sp. in having a denser ornamentation on the flank, and flank costae bending downwards close to the anterior margin (a sigmoidal pattern). Promyophorella orientalis Kobayashi and Tamura, Reference Kobayashi and Tamura1955, from the Callovian–Berriasian of Japan (Hayami, Reference Hayami1975) and the Callovian of Alaska (Poulton, Reference Poulton1979), also has denser flank ornamentation, with the anterior part of the flank costae more strongly curved.
The European late Toarcian P. spinulosa (Young and Bird, Reference Young and Bird1828) differs from P. basoaltorum n. sp. by having more elongate (sometimes subhorizontal) distal portions of flank costae, and the presence of a plicated median carina on the area (Francis, Reference Francis2000). According to the figures by Lycett (Reference Lycett1872–1879, pl. 3, figs. 4–6), area costellae seem to be more densely spaced in P. spinulosa.
Promyophorella hokonuiensis (Fleming, Reference Fleming1987), from the lower Temaikan (≈ Aalenian) of New Zealand, differs from P. basoaltorum n. sp. by its more elongate and less opisthogyrate shell, and a narrower antecarinal space. A similar elongate morphology is also found in Myophorella sp. B of Poulton, Reference Poulton1979, from the “early Bajocian” (now Aalenian) or late Toarcian of Yukon, Canada, although this last species also differs from P. basoaltorum n. sp. by the presence of incipient reticulate ornamentation on the escutcheon.
Promyophorella aff. P. basoaltorum new species
Figure 16.25, 16.26, 16.29
- Reference Pezzuchi and Takigawa1984
Myophorella sp.; Pezzuchi and Takigawa, p. 490.
- v Reference Massaferro2001
Myophorella (Myophorella) cf. M. araucana Leanza; Massaferro, p. 244, figs. 3d, f.
Occurrence
Cerro Cuche, Chubut Province, early Pliensbachian (E. meridianus? Biozone [≈ Ibex/Davoei Biozone]) (Fig. 3) according to Massaferro, Reference Massaferro2001, p. 246, based on material referred to Polymorphites? sp.
Description
Very small to small, opisthogyrate, rostrate shell. Slightly convex anterior margin, gradually passing to the convex ventral margin; posterior margin relatively wide (area ~1/3 of shell surface). Escutcheon excavated, with small transverse costellae variably developed. Escutcheon angulation stepped, a prominent carina at early growth stages. Area with commarginal costellae, increasing in spacing; sometimes posterior half with closely arranged irregular ridgelets. Antecarinal space smooth and widening posteriorly. Prominent marginal carina at early growth stages, sharp marginal angulation at late stages; area costellae swelling on the angulation. Flank costae oblique to growth lines, anteriorly curved, and diverging, sometimes bearing small, feeble, and closely spaced pustules along top.
Materials
CPBA 19657 (= cast MLP 20712), a slab with 20 isolated valves.
Measurements
MLP 20712a: L = 19 mm, H = 15 mm; MLP 20712b: L = 22 mm, H = 16 mm.
Remarks
This taxon differs from P. basoaltorum n. sp. by its smaller shell with finer and more densely arranged ornamentation. It also has a slightly rostrate shell shape. Otherwise, it is similar to P. basoaltorum n. sp., with which it is probably closely related if not conspecific.
Promyophorella araucana (Leanza, Reference Leanza1942)
Figure 19
- v Reference Burckhardt1902
Trigonia aff. T. angulata Sow.; Burckhardt, p. 246, pl. 4, figs. 5, 6.
- v Reference Windhausen1931
Trigonia angulata Sow.; Windhausen, pl. 2, fig. 5. [reproduced from Burckhardt, Reference Burckhardt1902]
- v* Reference Leanza1942
Trigonia (Clavitrigonia) araucana Leanza, p. 162, pl. 6, figs. 4–6.
- Reference Pérez and Reyes1977
Myophorella (Myophorella) araucana; Pérez and Reyes, p. 13, pl. 3, figs. 1–3. [reproduced from Leanza, Reference Leanza1942]
- v Reference Damborenea and Manceñido1992
Myophorella (Myophorella) araucana; Damborenea et al., pl. 116, fig. 12.

Figure 19. Promyophorella araucana (Leanza) from the Pliensbachian of Argentina. Scale bar = 10 mm. (1, 2) Lectotype, MLP 6724, shell, right lateral and right dorsal views, Piedra Pintada; (3, 4) paralectotype, MLP 3904, fragmentary right valve, lateral and dorsal views, Piedra Pintada.
Type materials
Lectotype (here designated): MLP 6724 (Fig. 19.1, 19.2); paralectotypes: some of the other specimens from Leanza's (Reference Leanza1942) extant syntypes, MLP 3904 (Fig. 19.3, 19.4), 6253 (part), and 6735. From the type series, we exclude the smallest specimen in MLP 6253.
Diagnosis
Opisthogyrate, elongate shell, with subtriangular flank. Escutcheon slightly impressed, smooth. Area narrow, with commarginal costellae. Escutcheon bounded by stepped angulation; marginal angulation sharp (stepped in early growth stages). Antecarinal space smooth, widening posteriorly. Flank with oblique, slightly diverging costae bearing small tubercles along top; costae typically with an initial almost straight segment and a sudden anteriorwards inflection towards the anteroventral margin (J pattern).
Occurrence
Estancia Santa Isabel, Salitral Grande Carrán Curá, Cerro del Vasco, Cerro Roth Sur (Piedra Pintada region), Neuquén Province; possibly Nueva Lubecka (Genoa River region), Chubut Province. Late early Pliensbachian (A. behrendseni Biozone [≈ Davoei Biozone]) to late Pliensbachian (F. fannini Biozone [≈ Margaritatus Biozone]) (Fig. 3).
Description
Medium-sized, opisthogyrate, somewhat elongate shell. Subtriangular flank in the most typical forms (Fig. 19.1, 19.3); anterior and ventral margins slightly convex, with a strongly curved transition between them. Area margin narrow; area ~1/4 of shell surface. Dorso-posterior angle obtuse and slightly projecting dorsally (Fig. 19.1). Escutcheon slightly impressed, with escutcheon margin slightly raised; escutcheon smooth, sometimes with short irregular costellae crossing from the area (Fig. 19.2, 19.4). Both angulations stepped, at least in early growth stages, with pustules at their intersection with area costellae (Fig. 19.3, 19.4). Area with commarginal costellae, evenly arranged (~8 costellae/cm) and regular anteriorly; in late growth stages irregular and less closely spaced (4 costellae/cm) or fading (Fig. 19.1, 19.2). Mid-area radial groove barely developed and slightly displaced dorsally (Fig. 19.1). Antecarinal space smooth, and widening posteriorly. Flank with oblique costae bearing small tubercles on top; in some shells, small narrow ridges extending posteroventrally from tubercles (Haidaia-like ornamentation). Flank costae slightly diverging, with an initial almost straight segment and a sudden anteriorwards inflection towards the anteroventral margin (J pattern; Fig. 19.1, 19.3).
Materials
The types plus 15 specimens: MLP 13008 (part), 31011, 31573?, 36334–36337; MPEF-PI 6393?, 6395?.
Measurements
Lectotype (MLP 6724): L = 45 mm, H = 33 mm.
Remarks
Promyophorella araucana can be distinguished from other Promyophorella species by its: (1) larger size, (2) less opisthogyrate shell, (3) (almost) smooth escutcheon, (4) narrower area, (5) subtriangular flank, and (6) J pattern in flank costae. Although P. basoaltorum n. sp. specimens may show some of the features described above, this combination of characters is only found in shells from southern Neuquén Province (and maybe northern Chubut), suggesting an endemic taxon derived from P. basoaltorum n. sp. Many of the shells from elsewhere referred in the literature to P. araucana actually belong to P. basoaltorum n. sp.
The materials from Cerrito Roth (= Cerro Roth Sur) described and figured by Leanza and Garate-Zubillaga (Reference Leanza, Garate-Zubillaga and Volkheimer1987, pl. 1, fig. 8) and from La Amarga (Neuquén) described and figured by Leanza (Reference Leanza1993, pl. 1, figs. 4, 5), in both cases as M. araucana, most likely belong to a different species, because they differ from both P. araucana and P. basoaltorum n. sp. in their: (1) larger size, (2) relatively large area, (3) narrow antecarinal space that does not expand posteriorly, and (4) strong flank costae bifurcating towards their distal end. These large, subtriangular, slightly opisthogyrate shells, with large areas, resemble juveniles of P. basoaltorum n. sp., and may even suggest a first step along the paedomorphic trends within the group (see Echevarría, Reference Echevarría2016). The bifurcating costae resemble those of Promyophorella? sanjuanina n. sp. and P.? catenifera.
Promyophorella cf. P. exotica (Möricke, Reference Möricke1894)
Figure 20
- *cf. Reference Möricke1894
Trigonia exotica Möricke, p. 49, pl. 1, fig. 9, pl. 6, fig. 9.
- cf. Reference Pérez and Reyes1977
Vaugonia (Vaugonia) exotica; Pérez and Reyes, p. 16, pl. 3, fig. 13. [reproduced from Möricke, Reference Möricke1894]
- v Reference Volkheimer, Manceñido and Damborenea1978b
Vaugonia sp.; Volkheimer et al., p. 212 (table 2).

Figure 20. Promyophorella cf. P. exotica (Möricke) from the Toarcian of Argentina. Scale bar = 10 mm. (1) MLP 36338, composite mold of left valve, Arroyo La Laguna; (2) MLP 36339, composite mold of right valve, Arroyo La Laguna; (3) MLP 36340, composite mold of right valve, Arroyo La Laguna; (4) MLP 36346, external mold of left valve, Arroyo Serrucho; (5, 6) MLP 36354a, composite mold of a left valve over external and internal mold surfaces, Arroyo Serrucho; (7) MLP 36347a, composite mold over the external mold surface of right valve, Arroyo Serrucho; (8) MLP 36348a, composite mold over the external mold surface of right valve, Arroyo Serrucho; (9) MLP 36349, composite mold of left valve, Arroyo Serrucho; (10) MLP 36350, composite mold on the external mold surface of left valve, Arroyo Serrucho; (11) MLP 36399a, composite mold of right valve, Arroyo Serrucho; (12) MLP 36351a, composite mold of left valve, Arroyo Serrucho; (13) MLP 36352a, composite mold of right valve, Arroyo Serrucho; (14) MLP 36400a, external mold of right valve, Arroyo Serrucho; (15) MLP 36355a, composite mold of right valve, Arroyo Serrucho; (16) MLP 36364a, composite mold of right valve, Cerro Puchenque; (17) MLP 36365a, composite mold of right valve, Cerro Puchenque; (18) MLP 36366a, composite mold, right lateral view, Cerro Puchenque; (19, 20) MLP 15547a, shell, dorsal and right lateral views, Estación Rajapalo.
Occurrence
Arroyo La Laguna (Los Patos region), San Juan Province; Arroyo Serrucho, Las Overas, Cerro Puchenque (West of Malargüe region), Mendoza Province; Ñiraico (Cordillera del Viento region), Neuquén Province. Early Toarcian (D. hoelderi Biozone [≈ Serpentinum Biozone] to P. pacificum Biozone [≈ Bifrons Biozone]) (Fig. 3).
Description
Small, opisthogyrate, sometimes elongate shell. Semicircular to subtriangular flank; area margin wide (area ~1/3 of shell surface). Escutcheon smooth (Fig. 20.19). Escutcheon angulation stepped, with pustules, each one linked to several area costellae. Area with fine commarginal costellae (Fig. 20.5–20.11, 20.14); umbonal 2 mm with costellae evenly spaced, later very densely spaced ridgelets (up to 30 ridgelets/cm). Mid-area radial groove barely developed, slightly displaced dorsally. Sharp marginal angulation (maybe as prominent carina in initial growth stages), with pustules on every other area costellae (one costella with pustule, one without; Fig. 20.4, 20.5, 20.14, 20.20). Narrow and smooth antecarinal space, widening posteriorly. Flank sculpture variable, with costae oblique to growth lines, bearing small, well-defined tubercles on top. Early costae anteriorly arcuate; late costae with distal extremes projecting anteriorly (usually with tubercles elongate antero-posteriorly; Fig. 20.4–20.7, 20.10, 20.13, 20.14, 20.20). Some shells with a third type of costa, with an almost straight initial segment and a subhorizontal to subcommarginal distal segment (‘L’ shaped; Fig. 20.1–20.3, 20.8, 20.9, 20.12, 20.15, 20.16). Nymph short (Fig. 20.19). Internal radial ridge in the posterior part of the inner surface, perpendicular to area margin (Fig. 20.3, 20.8, 20.15).
Materials
Nearly a hundred specimens, MLP 15547, 15548?, 36338–36367, 36399, 36400.
Measurements
MLP 36340: L = 30 mm, H = 22 mm; MLP 36341: L = 25 mm, H = 20 mm; MLP 36344: L = 24 mm, H = 17 mm; MLP 36366: L = 25 mm, H = 19 mm; MLP 36359: L = 21 mm, H = 16 mm; MLP 15547: L = 16 mm, H = 13 mm.
Remarks
Promyophorella cf. P. exotica shows a high degree of variability, mainly in the flank costae pattern. While in some shells the costae show a curved inflection (Fig. 20.4, 20.13, 20.14, 20.20), in others they develop an L-shaped pattern (Fig. 20.1–20.3, 20.7, 20.11, 20.15), and others even show both patterns (Fig. 20.9, 20.10). Costae tubercles can be small and numerous (Fig. 20.4, 20.8, 20.14), or relatively large and stout (Fig. 20.1, 20.3, 20.15). The inflection on flank costae can be developed towards the mid-flank (Fig. 20.7–20.12, 20.16) or somewhat closer to the anteroventral margin (Fig. 20.4–20.6, 20.20). Spacing and degree of development of area costellae can also show some variability (compare Fig. 20.14 with Fig. 20.15).
The species Trigonia exotica was described by Möricke (Reference Möricke1894) based on shells from the “Humphriesianus-Schichten” from Chile, implying a Bajocian age. Pérez and Reyes (Reference Pérez and Reyes1977) assigned an Aalenian–Bajocian range for the species in Chile. The species was later identified by Tornquist (Reference Tornquist1898) in Bajocian outcrops from Paso del Espinacito (San Juan Province, Argentina), although flank costae show an anterior disruption according to the figured shell. The material studied by Möricke could not be located; nevertheless, he described an anterior bend on flank costae (scarcely indicated in the more umbonal ribs, but very distinct in the anteroventral ones), from which the ribs run in the direction of the growth lines, and tubercles are weakly expressed. This description agrees well with the main pattern of flank ornamentation in our material. Nonetheless, the Toarcian shells described here show more closely spaced flank costae and thinner, more numerous area costellae (at least compared to Möricke's figure), so the taxon is treated as Promyophorella cf. P. exotica.
Due to its flank costae pattern, P. exotica was assigned to the genus Vaugonia by previous authors (e.g., Kobayashi and Mori, Reference Kobayashi and Mori1955; Pérez and Reyes, Reference Pérez and Reyes1977). Nevertheless, the shared characters of the material here described with P. basoaltorum n. sp. suggest a closer relationship with it than with any Vaugonia species (see Remarks under Pseudovaugonia n. gen.). Promyophorella cf. P. exotica differs from other Promyophorella species by its: (1) simple (without bifurcation) strongly bent and/or L-shaped flank costae, (2) densely spaced area costellae, (3) proportionally large area, and (4) smooth antecarinal space that widens posteriorly.
Shell shape in P. cf. P. exotica is very similar to that of P. basoaltorum n. sp., although in the first species it may be less opisthogyrate and sometimes may have a slightly larger area. Promyophorella cf. P. exotica also differs from P. basoaltorum n. sp. by its L-shaped flank costae and more densely spaced area costellae.
The shells identified by Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) as Vaugonia cf. V. gottschei from the late Toarcian and Pliensbachian? share with P. cf. P. exotica most characters, with the costae always showing an L pattern (even at early growth stages); they differ mainly by having a narrower area with finer and more densely spaced costellae. The Middle Jurassic species Trigonia chunumayensis Jaworski, Reference Jaworski1915, and Trigonia argentinica Jaworski, Reference Jaworski1926a, usually assigned to the genus Vaugonia, may have evolved from this stock by elongation of the shell.
Vaugonia (Vaugonia) spedeni Fleming, Reference Fleming1987, from the lower Temaikan (≈ Aalenian) of New Zealand is remarkably similar to P. cf. P. exotica and is here considered as a Promyophorella species. The main differences from P. cf. P. exotica are the anterior bifurcation of flank costae in some specimens of V. spedeni and a slight ventral displacement (i.e., at later growth stages) of the costae main locus of angulation in others, but none of those characters appears in every specimen of the species.
Some of the shells described here (Fig. 20.19, 20.20) are remarkably similar to P. spinulosa (Young and Bird, Reference Young and Bird1828), which, according to Francis and Hallam (Reference Francis and Hallam2003), is the earliest Promyophorella species in central-western Europe (late Toarcian to late Bajocian). The main difference is the presence of a finely tuberculate carina posteriorly bordering the mid-area groove in the European species (Lycett, Reference Lycett1872–1879; Francis, Reference Francis2000). According to Francis’ (Reference Francis2000) description, P. spinulosa shows a degree of intraspecific variation similar to that observed in P. cf. P. exotica described here. Promyophorella formosa (Lycett, Reference Lycett1872), figured by Fürsich et al. (Reference Fürsich, Berndt, Scheuer and Gahr2001, fig. 5F) from the Toarcian of Spain, is also very similar to some of the shells from Argentina (e.g., Fig. 20.13), although initial flank costae show more regular curvature and, once again, the European species seems to have a finely tuberculate carina bordering the mid-area groove.
Promyophorella sugayensis Kobayashi and Tamura, Reference Kobayashi and Tamura1955, from the Sugaya Formation (= Yamagami Formation, of probable Callovian age; Masatani and Tamura, Reference Tamura1959; Sato, Reference Sato and Westermann1992) from Japan is very similar to P. cf. P. exotica, even showing the same variability, with P. sugayensis var. geniculata having an angular geniculation on flank costae. It differs from P. cf. P. exotica by having the costae bending closer to the carina, and the antecarinal space seems to be narrower.
Some of the younger shells included within P. cf. P. exotica (most likely from P. pacificum Biozone) show the late costae fragmented into tubercles aligned both vertically and horizontally, sometimes rearranged in an en échelon pattern. This kind of ornamentation is reminiscent of some early species included in the genus Orthotrigonia, suggesting a close relationship of this last genus to Promyophorella. Orthotrigonia waipahiensis Fleming, Reference Fleming1987 (from the lower Temaikan of New Zealand) has a similar costation pattern. The New Zealand species differs by its narrower area and more elongate shell. Such an ornamentation pattern is also found in Orthotrigonia? sohli Poulton, Reference Poulton1979, from the Gypsum Spring Formation (Aalenian to early Bajocian in age) of Wyoming (USA), though the North American species is smaller. Although some specimens of P. cf. P. exotica can have L-shaped costae at early growth stages (Fig. 20.1), and some others can have some extra anterior flank costae (Fig. 20.16), the species does not show the combination of characters diagnosing the genus Orthotrigonia (see Remarks under Promyophorella).
Promyophorella agniaensis (Levy, Reference Levy1966)
Figure 21.1–21.3, 21.6–21.8
- v* Reference Levy1966
Myophorella (Myophorella) agniaensis; Levy, p. 238, fig. 1a, b.

Figure 21. Trigoniida from the Early Jurassic of Argentina. Scale bar = 10 mm. (1–3, 6–8) Promyophorella agniaensis (Levy): (1–3) SIRAME-SEGEMAR 7346, late Pliensbachian–early Toarcian, Pampa de Agnia; (1) SIRAME-SEGEMAR 7346a, holotype, left valve; (2) SIRAME-SEGEMAR 7346b, cast of an external mold of right valve; (3) SIRAME-SEGEMAR 7346c, juvenile left valve; (6) MLP 36325, composite mold of right valve, Toarcian, Cerro Tricolor; (7, 8) MLP 36326a, composite mold of right valve, internal and external mold surfaces, Toarcian, Cerro Tricolor. (4, 5, 9–11) Promyophorella? sanjuanina n. sp. from the late Pliensbachian to early Toarcian of Argentina: (4) paratype, MLP 36330, left valve, Toarcian, Arroyo La Laguna; (5) MLP 36331, fragmentary right valve, Toarcian, Arroyo La Laguna; (9) holotype, MLP 36329, right valve, Toarcian, Arroyo La Laguna; (10) paratype, MLP 36328, right valve, Toarcian, Arroyo La Laguna; (11) CPBA 17442, left valve, Arroyo de Las Vegas, late Pliensbachian–early Toarcian; (12–14) Promyophorella species indet. from the Toarcian of Cerro Tricolor, MLP 36327: (12) MLP 36327c, right valve; (13) MLP 36327d, fragmentary left valve; (14) MLP 36327a, fragmentary right valve. (15–19) Myophorelloidea genus and species indet. from the Toarcian of Cañada Colorada: (15–18) MLP 36395; (15, 18) MLP 36395a, right valve, lateral and dorsal views; (16) MLP 36395b, left valve; (17) MLP 36395c, fragmentary left valve; (19) MLP 36396, external mold of left valve.
Type material
Holotype: DNGM 641a (= SIRAME-SEGEMAR 7346a) from the late Early Jurassic (probably early Toarcian) of Sierra de Pampa de Agnia, Chubut Province, Argentina. Adult shell figured by Levy (Reference Levy1966, p. 239, fig. 1b) and Figure 21.1.
Diagnosis
Opisthogyrate shell. Flank with tuberculate costae, subcommarginal next to the umbo, then oblique to growth lines and curved, almost perpendicular to the ventral margin at late growth stages. Area with fine commarginal costellae; median groove well defined. Antecarinal space wide and smooth.
Occurrence
Cerro Tricolor (Grande River region), Mendoza Province; Pampa de Agnia (central Chubut region). Early Toarcian (Tenuicostatum Biozone–D. hoelderi? Biozone [≈ Serpentinum Biozone]) (Fig. 3).
Description
Medium-sized, elongate, opisthogyrate shell. Escutcheon partly observable in the holotype; smooth except for small thorn-like projections from the tubercles on the escutcheon angulation. Area with fine costellae at early growth stages, strong growth lines at late growth stages; sometimes with irregular costellae posteriorly (Fig. 21.1). Marginal angulation rounded, slightly overhanging the antecarinal sulcus, with rounded prominent tubercles. Very shallow antecarinal sulcus, smooth and posteriorly widening (Fig. 21.1, 21.2). Posterior flank costae almost straight, perpendicular to ventral margin, wide and with stout, somewhat elongate tubercles along top (Fig. 21.3, 21.7, 21.8); intercostal spaces barely wider than costae (Fig. 21.1–21.3, 21.6–21.8). Flank costae anteriorly fading, being recognized only by the aligned tubercles (Fig. 21.1, 21.6); anteriormost portion of the flank smooth. Umbonal flank costae oblique to growth lines, curved, surrounding the umbo (Fig. 21.3).
Materials
Besides the holotype, SIRAME-SEGEMAR 7346 includes an external mold of a right valve (Fig. 21.2) and a left juvenile valve (illustrated by Levy, Reference Levy1966, p. 239, fig.1a; Fig. 21.3). Three specimens from Mendoza (MLP 36325, 36326) were also studied.
Measurements
Holotype (SIRAME-SEGEMAR 7346): L = 53 mm, H = 32 mm; juvenile illustrated by Levy: L = 32 mm, H = 20 mm; third specimen from SEGEMAR: L = 37 mm, H = 30 mm; MLP 36326: L = ~40 mm, H = ~24 mm.
Remarks
Promyophorella agniaensis can be distinguished from other Promyophorella species by: (1) the stout flank costae perpendicular to the ventral margin and almost as wide as the intercostal spaces, (2) the wide antecarinal sulcus, and (3) the fading of anterior flank costae at late growth stages. The material from Mendoza (Fig. 21.6–21.8) can be assigned with confidence to this species. This new record from Cerro Tricolor reveals a wide geographical distribution for the species within the basin.
Levy (Reference Levy1966) assigned the species to the subgenus Myophorella (Myophorella) due to the well-developed tubercles, separated by smooth spaces. Nevertheless, we regard the ornamentation pattern in P. agniaensis as closer to that of P. sigmoidalis than to that of M. nodulosa. Tubercles on flank costae in Myophorella are much larger and more prominent than the tubercles in P. agniaensis.
The species most likely evolved from P. basoaltorum n. sp. by developing a stronger ornamentation pattern, with flank costae less anteriorly curved at initial growth stages. Although the straight flank costae on adult shells are comparable to those of the genus Scaphorella and to those of some of the species here described (e.g., P. araucana), none of these taxa shows a smooth anteriormost flank.
Promyophorella species indet.
Figure 21.12–21.14
Occurrence
Cerro Tricolor (Grande River region), Mendoza Province. Early Toarcian (Tenuicostatum? Biozone) (Fig. 3).
Description
Small, orthogyrate to slightly opisthogyrate shell. Large area (Fig. 21.12, 21.14; ~2/5 of shell surface or more); with commarginal costellae at early growth stages, replaced later by strong growth lines (Fig. 21.13). Sharp marginal angulation, with tubercles on top. Antecarinal space smooth, slightly widening posteriorly (Fig. 21.12–21.14). Straight, slightly diverging flank costae and orthogonal to ventral margin at early growth stages, curving anteriorly towards the anteroventral margin (Fig. 21.13). Flank costae with tubercles on top.
Materials
Fragmentary shells: MLP 36327; probably also part of MLP 17186.
Measurements
MLP 36327a: L = 23 mm, H = 17 mm; MLP 36327b: L = 23 mm, H = 20 mm.
Remarks
The material cannot be assigned to any of the Promyophorella species already recognized in the basin. Slightly diverging J-shaped flank costae is a character typical of P. araucana, but in this last species the costae curve anteriorwards from the beginning. Flank costae on Promyophorella agniaensis start their development with a straight portion, but Promyophorella species indet. differs from this species (and from P. araucana) by having a larger area and orthogyrate shell shape. The straight portion of the flank costae in Promyophorella species indet. is like that found on the genus Scaphorella, although the former lacks the anterior bifurcating ornamentation observed in the latter.
Promyophorella basoaltorum n. sp. most likely gave rise to this species by a change in flank costae pattern, a widening of the area margin, and development of a less-opisthogyrate shell.
Promyophorella? sanjuanina new species
Figure 21.4, 21.5, 21.9–21.11
- pv Reference Volkheimer, Manceñido and Damborenea1978b
Vaugonia sp.; Volkheimer et al., p. 212 (table 2).
- v Reference Álvarez, Ramos, Aguirre-Urreta, Álvarez, Cegarra, Cristallini, Kay, Lo Forte, Pereyra and Pérez1996
Myophorella cf. M. araucana; Álvarez, p. 92, fig. 23h.
- v Reference Álvarez1997
Myophorella cf. M. araucana (Leanza); Álvarez, p. 231, pl. 1, fig. h.
- v Reference Pérez and Reyes2008
Myophorella (Myophorella) sp. 2; Pérez and Reyes in Pérez et al., p. 88, pl. 14, fig. 9.
Type materials
Holotype: MLP 36329, paratypes: MLP 36328, 36330.
Diagnosis
Small to medium-sized, slightly opisthogyrate, elongate, trapezoidal shell. Escutcheon smooth; escutcheon angulation stepped, with small tubercles on top. Wide area with submedian groove dorsally displaced. Area with commarginal costellae, less dense and slightly stronger at late growth stages. Marginal angulation with tubercles. Antecarinal space feebly developed. Two sets of flank costae, both with well-developed regular tubercles: a sub-vertical regular dorso-posterior set, and a sub-horizontal to sub-commarginal irregular and frequently bifurcated anterior set (sometimes with costae broken up into tubercles).
Occurrence
Arroyo La Laguna (Los Patos region), San Juan Province; Las Overas (west of Malargüe region), Mendoza Province. Early Toarcian (D. hoelderi–P. largaense or maybe P. pacificum biozones [≈ Serpentinum–Bifrons biozones]) (Fig. 3). The material illustrated by Álvarez (Reference Álvarez, Ramos, Aguirre-Urreta, Álvarez, Cegarra, Cristallini, Kay, Lo Forte, Pereyra and Pérez1996) is from late Pliensbachian or early Toarcian (Tenuicostatum Biozone) from Arroyo de Las Vegas (~7 km north from Arroyo La Laguna), and she mentioned its presence in Arroyo La Laguna, both localities in San Juan Province. The material from Chile (Quebrada Asientos, Atacama Region) is from the late Pliensbachian F. fannini Biozone (≈ Margaritatus Biozone).
Description
Small to medium-sized, slightly opisthogyrate, elongate, trapezoidal shell. Escutcheon excavated, smooth (Fig. 21.9). Escutcheon angulation stepped, with small tubercles on top (projecting slightly towards the escutcheon). Area wide (~1/3 of shell surface), with submedian groove dorsally displaced. With closely spaced commarginal costellae; dorsal and ventral portions normally meeting at an angle towards the groove. Area costellae densely arranged at early growth stages, increasing spacing and becoming stronger during ontogeny (Fig. 21.4, 21.10). Sharp to rounded marginal angulation; with tubercles, though not as strong as those on the escutcheon angulation. Antecarinal space barely developed, but interrupting flank costae (Fig. 21.4, 21.9, 21.10). Flank costae strong, tuberculate, oblique to growth lines; with a sub-vertical dorso-posterior portion (slightly incurved anteriorwards) and an anterior, irregular, and frequently bifurcated sub-horizontal to sub-commarginal portion; anteroventral costae sometimes fragmented into tubercles, resulting in a reticulate appearance (Fig. 21.4, 21.5). Anteriormost portion of flank costae sharp and continuous, without tubercles in some specimens.
Etymology
In reference to San Juan Province, from where the type material was collected.
Materials
Eight specimens, the types plus MLP 36312c, 36331–36333, CPBA 17442 and SNGM 665.
Measurements
MLP 36329 (holotype): L = 35 mm, H = 23 mm; MLP 36330 (paratype): L = 40 mm, H = 24 mm; CPBA 17442: L = 38 mm, H = 26 mm.
Remarks
General morphology and ornamentation (including the bifurcation on the anterior portion of flank costae) match those of Myophorella sp. 2 of Pérez et al. (Reference Pérez, Aberhan, Reyes and von Hillebrandt2008) from the late Pliensbachian of Chile, and thus is considered here as conspecific, although the Chilean material is relatively shorter and higher. Promyophorella? sanjuanina n. sp. differs from other species within the Neuquén and Chubut basins by its: (1) bifurcating flank costae, sometimes with a reticulate pattern; (2) reduced antecarinal space, and (3) progressively stronger and more spaced area ornamentation. Promyophorella? catenifera (Fig. 18.1–18.4), from the Pliensbachian–Toarcian of Chile, shows similar flank ornamentation, although in this last case costae do not bend anteriorwards as markedly as in Promyophorella? sanjuanina n. sp. Besides, P.? catenifera bears a well-developed antecarinal space and an almost smooth area. One of the specimens referred to Myophorella araucana by Leanza and Garate-Zubillaga (Reference Leanza, Garate-Zubillaga and Volkheimer1987, pl. 1, fig. 8, MOZ PI-0937/1) from the early Pliensbachian at Cerrito Roth (Neuquén Province) and the one referred to the same species by Leanza (Reference Leanza1993) from the Pliensbachian at La Amarga (Neuquén Province) also share with Promyophorella? sanjuanina n. sp. the bifurcating flank costae and the reduction of the antecarinal space; the material from Neuquén, nonetheless, is larger and with stronger ornamentation.
Vaugonia (?) yukonensis Poulton, Reference Poulton1979 (p. 25–26, pl. 8, figs. 6–11), probably from the “middle Bajocian” (now early Bajocian) of Yukon, shows a similar somewhat irregular anteriorwards costation pattern to that of Promyophorella? sanjuanina n. sp., even with the reticulate ornamentation. Shell sculpture is much coarser and sometimes more densely spaced in the North American species. Poulton (Reference Poulton1979) recognized some affinity of Vaugonia (?) yukonensis to Myophorella (including Promyophorella).
The anterior bifurcation and disruption in the flank costae of all these species, including P.? sanjuanina n. sp., also suggest a relationship to Scaphorella. The type species, Scaphorella leanzai (Lambert, Reference Lambert1944) bears a posterior set of sub-vertical, tuberculate, nearly straight, narrow costae, with the anteriormost ones strongly curving anteriorly (and some of them bifurcating) to generate the anterior set of sub-horizontal costae. This pattern differs from the one seen in Promyophorella? sanjuanina n. sp., where the anterior set of costae is much more irregular (especially at late growth stages). Scaphorella susanae Pérez and Reyes in Pérez et al., Reference Pérez, Aberhan, Reyes and von Hillebrandt2008, is the earliest species assigned to the genus (late Toarcian and early Aalenian from Atacama, Chile); the anterior disruption of the ornamentation pattern in this species is similar to that found on P.? sanjuanina n. sp., but in S. susanae there is a wide and well-developed, smooth antecarinal space, and area costellae are thinner than on the new species. Besides, the reticulate pattern on some specimens of P.? sanjuanina n. sp. does not develop in the known species assigned to Scaphorella.
Considering the well-defined regular tubercles on oblique diverging, gently curved, costae departing from antecarinal space at obtuse anterior angles, we include P.? sanjuanina n. sp. in the genus Promyophorella, but using open nomenclature.
Myophorelloidea genus and species indet.
Figure 21.15–21.19
Occurrence
Cañada Colorada (west of Malargüe region), Mendoza Province. Late Toarcian (Fig. 3).
Description
Small, slightly opisthogyrate, elongate shell. Escutcheon smooth (Fig. 21.18). Area smooth or bearing very thin ridglets (almost imperceptible); with a median row of tubercles (Fig. 21.17–21.19). Escutcheon and marginal angulations with transverse tubercles. Antecarinal space very narrow (Fig. 21.15–21.17). Flank with oblique, L-shaped costae; posterior branches strong, subradial, and diverging; anterior branches slightly concave ventrally and thinner than posterior ones (Fig. 21.15–21.17). Flank costae with small, round tubercles.
Materials
MLP 36395–36397.
Measurements
MLP 36395d: L = 35 mm, H = ~26 mm.
Remarks
This taxon differs from other Myophorelloidea species by its: (1) smooth area, except for the median radial row of small tubercles; (2) reduced antecarinal space; and (3) oblique, L-shaped flank costae with the anterior branches slightly concave ventrally and with tubercles on top. Area ornamentation is remarkably similar to that of Poultoniella n. gen., but the flank costae pattern strongly differs. The taxon here described has a similar ornamentation pattern to that of Promyophorella? sanjuanina n. sp., as well as a reduced antecarinal space, but, in contrast, the latter has a commarginally ornamented area. Although frenguelliellids may develop oblique costae on the flank, even with tubercles, the regularity in the ornamentation pattern and the flank costae generating a clearly obtuse anterior angle with the antecarinal space suggest a closer affinity with the Myophorellidae.
Trigonia lycetti Gottsche, Reference Gottsche and Stelzner1878, from the Bajocian of Paso del Espinacito, San Juan Province (type material housed at the SEGEMAR under collection numbers SIRAME-SEGEMAR 7877, 7981, and 7982), also shares a similar flank ornamentation pattern with this taxon, and it even has a median “carina” on the area with small tubercles. It differs, nonetheless, by having well-developed transverse costellae on the area (finely preserved on SIRAME-SEGEMAR 7877), a well-developed antecarinal space, and a middle step on the area (Trigonia lycetti may actually be a species of Vaugonia). The material from Cañada Colorada does not show any sign of a middle step on the area, precluding the assignment of the taxon to Vaugonia.
Vaugonia sp. cf. V. (?) yukonensis described by Poulton (Reference Poulton1979, p. 26, pl. 8, fig. 12), probably from the Bajocian of southern Yukon Territory, Canada, resembles the material here described, but differs by having a sharper umbo and a median groove in the area. In addition, the Canadian material has flank costae more densely disposed, and posterior and anterior branches are similarly developed and continuous, whereas in the Argentinian material the posterior branches tend to be stronger, and the anterior ones may be more numerous (Fig. 21.15–21.17).
Acknowledgments
We want to express our most grateful acknowledgement to A. Riccardi for invaluable help with the biostratigraphic control of the material here described. C. Gulisano, S. Lanés, and A. Pagani are thanked for collecting many of the specimens. J. Posik, technician at the Invertebrate Paleontology Division at MLP, aided with the preparation of part of the material. A. Matsuoka provided some relevant bibliographic items. The curators of all the visited and consulted collections are deeply thanked, too: A. Riccardi (MLP), A. Pagani (MPEF), B. Boilini and A. Garrido (MOZ), M. Tanuz (CPBA), S.M. Devincenzi (IANIGLA), M. Serrano (SIRAME-SEGEMAR), E. Pérez D'Angelo and A. Rubilar (SNGM), W. Etter, S. Kühni, and F. Wiedenmayer (NHMB), G. Heumann (IGPB), D. Merle and C. Noyes (MNHN), and E. Nesbitt (BMNH). We are likewise indebted to M. Griffin for his thorough idiomatic revision of the text. The text greatly benefited from the comments and suggestions of both reviewers, T. Poulton and S. Schneider, as well as C. Maples and editorial staff; we thank them all for their painstaking work. Necessary permits, issued by the respective authorities of Mendoza, Neuquén, and Chubut provinces, enabled collecting fossil specimens during the last 17 years. CONICET funded the investigations of the three authors during most of their careers (most recently, through PIP 112-200801-01567 and PIP 112-201101-01088), as well as UNLP (most recently, through 11/N873).
Appendix 1: List of specimens from Argentina
Specimens of each Trigoniida species listed by locality from North to South. Locality map, repository abbreviations, and local biozonation in text. N = Number of specimens






















