Introduction
The genus Rimularia Nyl. is well represented in cold to cool temperate climates of both hemispheres. Introduced by Nylander (Reference Nylander1868) to accommodate the single species R. limborina, it was not until the late 20th century that the genus was brought to the general attention of lichenologists, principally when Hafellner (Reference Hafellner1984) introduced the family Rimulariaceae and Hertel & Rambold (Reference Hertel and Rambold1990) revised the 13 taxa known at that time. Since then, numerous additions to the genus have been made, most frequently from higher latitudes of the Southern Hemisphere (Coppins & Kantvilas Reference Coppins and Kantvilas2001; Fryday Reference Fryday2004; Coppins & Fryday Reference Coppins and Fryday2006; Kantvilas & Elix Reference Kantvilas and Elix2007; Fryday & Øvstedal Reference Fryday and Øvstedal2012) but also from elsewhere, including tropical arid Australia (Kantvilas et al. Reference Kantvilas, McCarthy and Stuckey2008), New Guinea (Aptroot & Sipman Reference Aptroot and Sipman1991), Taiwan (Aptroot & Sparrius Reference Aptroot and Sparrius2003), North America (Rambold & Printzen Reference Rambold and Printzen1992; Timdal Reference Timdal2002; Coppins & Fryday Reference Coppins and Fryday2006) and Great Britain (Coppins & Kantvilas Reference Coppins and Kantvilas2001).
At present, 33 species are accepted in Rimularia (MycoBank database), of which 12 occur in Australasia (Galloway Reference Galloway2007; McCarthy Reference McCarthy2013), with seven of these recorded for Tasmania. The genus is highly variable, with a crustose, effuse, areolate or squamulose thallus that may contain depsidones (norstictic, stictic or lobaric acids), depsides (gyrophoric acid) or dibenzofuranes (porphyrilic acid), and is essentially united by having apothecia with an opaque, cupulate exciple, typically composed of rather parenchymatous hyphae, richly branched and anastomosing paraphyses that are usually moniliform towards the apices, distinctive, eight-spored asci, and simple, non-halonate, hyaline ascospores (Lumbsch Reference Lumbsch1997; Coppins & Kantvilas Reference Coppins and Kantvilas2001; Giavarini & David Reference Giavarini, David, Smith, Aptroot, Coppins, Fletcher, Gilbert, James and Wolseley2009). Currently the genus is placed in the Trapeliaceae (Lumbsch & Huhndorf Reference Lumbsch and Huhndorf2010), together with somewhat similar genera such as Amylora, Lithographa, Placynthiella and Ptychographa.
In the opinion of the author, Rimularia is a challenging and difficult genus. On the one hand, when well developed, the distinctive asci and paraphyses readily separate taxa from the mass of unidentified (and often unidentifiable) crustose lecideoid lichen collections that fill herbaria, especially in Southen Hemisphere countries. On the other hand, many species of Rimularia are exceedingly rare and occur in extreme habitats, often only as poorly developed thalli that have few apothecia, asci or ascospores. This has certainly been the author's experience in Tasmania, where many taxa have been collected repeatedly, but never in adequate quantities for study, and expeditions to their provenance to collect more material, almost inevitably to some remote destination, prove unsuccessful. The fact that Øvstedal et al. (Reference Øvstedal, Tønsberg and Elvebakk2009), in their recent Svalbard flora, include eight species but only five with names, suggests the same situation occurs elsewhere. A further complication is that Rimularia as currently applied is almost certainly heterogeneous (Lumbsch Reference Lumbsch1997; Coppins & Kantvilas Reference Coppins and Kantvilas2001; Coppins & Fryday Reference Coppins and Fryday2006) and will require a general broad-based revision and application of sophisticated methods, such as using DNA sequence data, to elucidate species relationships and define natural groups. For the present, however, the most urgent task in this author's opinion is to bring as many taxa of Rimularia in the broadest sense ‘out of the cupboards’ and, with thorough descriptions and names, to the general attention of lichenologists. This is especially so as several of the species are extremely uncommon, have very restricted distributions, or occur in threatened habitats, such as on the highest mountain pinnacles. In this paper, five such species are described, along with the common and widespread R. psephota with which several of the new taxa are compared. However, this in no way completes the Rimularia story for Tasmania, and the key to the species presented here should be considered as provisional at this stage.
Materials and Methods
The study is based principally on the author's collections, housed in the Tasmanian Herbarium (HO), comparative reference material from other herbaria (mainly the Australian National Herbarium, CANB), published descriptions, and on the author's observations in Tasmania accumulated over the last few decades.
Descriptions are based on hand-cut sections of the thallus and ascomata, mounted in water, 15% KOH, Lugol's iodine, ammoniacal erythrosin and lactophenol cotton blue, and examined with high-power, light microscopy. Dimensions of ascospores are based on at least 60 observations and are presented in the format: 5th percentile–average–95th percentile, with outlying values given in parentheses. Chemical analyses using thin-layer chromatography follow standard methods (Orange et al. Reference Orange, James and White2001); solvent A was the preferred medium. High-performance liquid chromatography (Elix et al. Reference Elix, Giralt and Wardlaw2003) of selected specimens was undertaken by Professor J. A. Elix in Canberra. Nomenclature of pigments follows Meyer & Printzen (Reference Meyer and Printzen2000).
Asci in Rimularia display subtle but consistent differences, as illustrated by Hertel & Rambold (Reference Hertel and Rambold1990). In R. limborina, the type of the genus, the tholus has amyloid flanks and an amyloid cap with vestiges of a narrow, weakly amyloid channel in the upper, central part (also illustrated by Hafellner Reference Hafellner1984); this is the ‘typical’ Rimularia-type ascus. Other species, in particular several that have been described recently from the Southern Hemisphere, have an ascus where the tholus has amyloid flanks and an amyloid cap, but is weakly amyloid to non-amyloid in the central part; this type of ascus has been illustrated by Coppins & Kantvilas (Reference Coppins and Kantvilas2001), where it was referred to as ‘Trapelia-type’. This nomenclature is followed below.
New Species
Rimularia albotessellata Kantvilas sp. nov.
MycoBank No.: MB807726
Thallo albido, areolato-tessellato, pannarinum continenti, apotheciis nigris, gyrosis, 0·1–1·5 mm latis, et ascosporis ellipsoideis vel fusiformibus, 11–19 µm longis, 4–7 µm latis insignis.
Typus: Australia, Tasmania, Mt Rogoona summit, 41°54′S, 146°12′E, 1350 m alt., on alpine dolerite rocks, 27 November 2012, G. Kantvilas 665/12 (HO—holotypus).

Fig. 1. Tasmanian species of Rimularia. A, R. albotessellata (holotype) habit; B, R. albotessellata detail, showing adnate or sunken black apothecia at the margins of, or between, the white thallus areoles; C, R. aspicilioides (holotype) habit; D, R. aspicilioides detail, showing black aspicilioid apothecia surrounded by abraded greyish thalline tissue. Scales: A & C=2 mm; B & D=1 mm. In colour online.
Thallus crustose, areolate, forming irregular, undelimited, rather dispersed patches extending from several centimetres to as much as a metre or more in width. Individual areoles 0·4–2·0 mm wide, 0·15–1·00 mm thick, irregularly roundish or rhomboidal, or ±angular when crowded together, plane to convex or, more rarely, slightly concave and with upturned edges, white to whitish grey in the centre, blackened at the margins, markedly scabrid and/or verruculose, separated by deep cracks; upper cortex poorly defined, c. 40–70 µm thick, composed of very indistinct, entangled hyphae 1–2 µm thick, densely inspersed with minute crystals mostly insoluble in KOH; photobiont a unicellular green alga (?Chlorella) with cells irregularly roundish, ellipsoid or rhomboidal, 7–15 µm wide, encased in a gelatinous sheath 1–2 µm thick; medulla I−, inspersed with minute colourless crystals that fluoresce in polarized light and mostly do not dissolve in KOH. Prothallus black, crustose, subtending the areoles and protruding in the gaps between them, in section reddish brown, K+ intensifying reddish.
Apothecia developed at the margins of the areoles or in the gaps between them, adnate, superficial or rather sunken between the areoles, 0·1–1·5 mm wide, roundish or angular when crowded together, glossy jet black, plane at first and with a thin but distinct proper exciple 30–100 µm thick, soon convex and seemingly immarginate, gyrose-contorted and misshapen, commonly eroded, in section comprising a mass of amorphous, opaque dark brown, K+ reddish brown, sterile excipular tissue with embedded colourless to pale brownish ‘locules’ of hymenial tissue c. 100–200 µm wide and 80–200 µm tall. Hypothecium to 160 µm thick, colourless to pale reddish brown, I−, inspersed with oil droplets, poorly differentiated from the hymenium. Hymenium c. 40 µm thick, colourless to dilutely pale brownish, separating readily in KOH, overlain by an olive-brown, K± greenish grey epithecium 4–10 µm thick. Paraphyses 1·5–2·0 µm thick, sparsely branched and anastomosing; apices rounded and not expanded, or capitate and sometimes moniliform, to 3–4 µm wide, colourless or greenish grey. Asci 8-spored, clavate, very rarely seen (immature asci 30–40×12–15 µm), approximating the Trapelia-type: with a thin, intensely amyloid outer wall, a prominently thickened tholus with intensely amyloid flanks and more weakly amyloid central part, and lacking an ocular chamber. Ascospores simple, hyaline, non-halonate, thin-walled, ellipsoid to fusiform, (11·0–)12·0–15·6–19·0×4·0–5·1–6·0(–7·0) µm.
Conidiomata not found.
Chemistry
Pannarin and deoxypannarin; thallus K+ pale yellow, KC−, C−, P+ pale orange, UV−. These compounds appear high on TLC plates, one as a black spot after charring and the other orange.
Etymology
The specific epithet refers to the thallus which forms an extensive pattern of white areoles, rather like the tiles of a mosaic.
Distribution and ecology
This is a very common species on alpine dolerite rocks in Tasmania, where it forms extensive expanses, often of many square metres in size, on exposed slabs of bedrock and on larger boulders. Dolerite is the predominant rock type in central and eastern Tasmania, with peaks rising to 1500 m elevation (Kantvilas Reference Kantvilas1995). Like many other typically alpine lichens, the new species may also extend to adjacent lower elevations on exposed rocks such as along the banks of rivers and lakes. It is part of a very rich assemblage of lichens in these habitats. Typical associated taxa include species of the blackish, tufted moss, Andreaea, and the lichens Cameronia pertusarioides Kantvilas, Hymenelia sp., Lecanora cf. demersa (Kremp.) Hertel & Rambold, Placopsis spp., Ramboldia petraeoides (Nyl. ex C. Bab. & Mitten) Kantvilas & Elix, Rhizocarpon geographicum (L.) DC., Rimularia circumgrisea Kantvilas, R. psephota (Tuck.) Hertel & Rambold, Trapelia lilacea Kantvilas & Elix and Xanthoparmelia stygiodes (Nyl. ex Cromb.) O. Blanco et al.
Remarks
The combination of a conspicuous thallus of vivid white, mosaic-like areoles with a black margin, the black, contorted apothecia, ellipsoid to fusiform ascospores, and the presence of pannarin, a metabolite previously unrecorded for the genus, characterize this distinctive lichen. A similar morphology is displayed by two other alpine species that may occur in the same habitats: one, Austrolecia antarctica Hertel, is distinguished by containing norstictic acid and having 1-septate ascospores and Catillaria-type asci; the other lacks lichen substances, has Biatora-type asci and 3-septate ascospores, and may represent a species of Toninia.
That this species has been neglected in taxonomic studies until now is due mainly to technical difficulties in observing and interpreting its anatomical details. It is possibly one of Tasmania's most common alpine lichens, yet fertile material is extremely rare. Black, apothecia-like structures are almost invariably present, but these are heavily carbonized, extremely brittle and mostly lack any hymenial tissue. At their youngest stage they are plane, adnate and have a thin, slightly raised proper margin, not unlike those of the lobaric-acid containing species, R. umbratilis Kantvilas & Coppins and its allies (Coppins & Kantvilas Reference Coppins and Kantvilas2001); no fertile asci are present at this stage. As the apothecia develop, the dark excipular tissues become ever more massive; the hymenium becomes very reduced, soon divided by pigmented strands of sterile tissue, then reduced further to single or multiple ‘hymenial locules’ and then finally excluded altogether. Squashes of thin sections of the hymenium revealed abundant paraphyses, occasional ascospores but rarely any well-formed asci. Thus the detailed ascus structure could not be clearly observed or drawn, and was inferred from multiple observations of young or ruptured asci. Even so, I feel that this remarkable species can justifiably be described in Rimularia, pending broader studies of the genus as a whole. Apothecial morphology and anatomy, though extreme, are essentially analogous to that seen in other species that have heavily carbonized, gyrose-contorted, rather lumpy apothecia; for example, R. cerebriformis Kantvilas & McCarthy from mainland Australia and R. gyrizans (Nyl.) Hertel from the temperate Northern Hemisphere.
Specimens examined. Australia: Tasmania: summit of Adamsons Peak, 43°21′S, 146°49′E, 1225 m alt., 2001, G. Kantvilas 14/01 (HO); Mt Mawson plateau, 42°41′S, 146°35′E, 1270 m alt., 2007, G. Kantvilas 382/07 (HO); Lake Judd near Anne River outlet, 43°00′S, 146°26′E, 591 m alt., 2009, G. Kantvilas 45/09 (HO); Skullbone Plains, c. 200 m from road intersection to Nive River, 42°02′S, 146°21′E, 970 m alt., 2012, G. Kantvilas 728/12 (E, HO, MSC); Hartz Mountains, saddle between Hartz Peak and Mt Snowy, 43°15′S, 146°46′E, 1090 m alt., 2012, G. Kantvilas 684/12 (HO); northern summit of Mt Rogoona, 41°53′S, 146°12′E, 1330 m alt., 2012, G. Kantvilas 672/12 (HO); Hartz Peak, 43°15′S, 146°46′E, 1200 m alt., 2012, G. Kantvilas 688/12 (HO); Clarke Falls, 41°55′S, 146°11′E, 930 m alt., 2012, G. Kantvilas 640/12 (HO); Emily Tarn, 43°15′S, 146°46′E, 1060 m alt., 2012, G. Kantvilas 681/12 (HO); Skullbone Plains, 42°02′S, 146°19′E, 1000 m alt., 2012, G. Kantvilas 101/12 (HO); Mountains of Jupiter summit, 41°57′S, 147°11′E, 1320 m alt., 2012, G. Kantvilas 650/12 (HO); Mt Styx, 146°45′S, 42°51′E, 1090 m alt., 2012, G. Kantvilas 743/12 (HO).
Rimularia aspicilioides Kantvilas sp. nov.
MycoBank No.: MB807727
Species insignis, thallo areolato, acidum 2′–O–methylperlatolicum continenti, apotheciis aspicilioideis, 0·2–1·1 mm latis, ascis Rimulariae typo pertinentibus et ascosporis simplicibus, ovatis vel ellipsoideis, 7–12 µm longis, 5·0–7·5 µm latis recognita.
Typus: Australia, Tasmania, Pine Lake, 41°45′S, 146°42′E, 1230 m alt., on alpine dolerite boulders on steep scree slope, 18 February 2009, G. Kantvilas 72/09 (holotypus—HO; isotypus—E).
(Figs 1C & D, 4A)
Thallus crustose, areolate, forming irregular, widely spreading patches to 10 cm wide or more. Individual areoles 0·2–0·3 mm wide, to 0·3–1·0 mm thick, ±neatly roundish, aggregated in tight clusters separated by deep cracks, from the earliest stages with a central, glossy brown apothecial initial, surrounded by a pale to dark grey margin, soon becoming ±entirely excluded by the apothecium; upper cortex composed of a layer c. 10 µm thick of ±vertically orientated, loosely entangled, pale brownish hyphae 3–5 µm thick; photobiont a unicellular green alga with cells globose to ellipsoid, 5–16 µm wide, encased individually or in groups of 2–4 in a gelatinous sheath 1–2 µm thick; medulla I−, inspersed with minute colourless crystals that fluoresce in polarized light and dissolve in KOH. Prothallus black, crustose, ±loosely attached to the substratum, in section reddish brown, K+ intensifying reddish, here and there with a patchy blue-green, N+ crimson pigment.
Apothecia aspicilioid, adnate, sessile or immersed in the areoles, 0·2–1·1 mm wide, roundish, lobate or misshapen, especially when crowded together, very numerous, occurring as individuals or fused together in dense clusters, up to 2–3 mm wide and up to 1·5 mm thick, of multiple, large and small apothecia, loosely attached to the substratum and separated by deep cracks; disc black to brownish black, plane, undulate to slightly convex, not gyrose; thalline margin pale grey to dark grey-black, inrolled, persistent, in section 20–100 µm thick, cupular, at the outer edge typically surrounded by a rim of opaque, reddish black prothalline tissue c. 10–50 mm thick; proper exciple absent. Hypothecium 50–100 µm thick, colourless, I+ blue in the upper part, inspersed with oil droplets, subtended by a layer of photobiont cells. Hymenium 35–50 µm thick, dilutely yellow-brown, K+ olive-brown throughout or only in the uppermost 5–10 µm, separating readily in KOH. Paraphyses 1·5–2·5 µm thick, simple to sparsely branched and anastomosing; apices mostly not capitate, or slightly expanded to 4–5 µm, colourless or brownish. Asci 8-spored, broadly clavate, 28–40×12–22 µm, of the Trapelia-type: with a thin, intensely amyloid outer wall, a prominently thickened tholus with intensely amyloid flanks, weakly amyloid in the central part, and lacking an ocular chamber. Ascospores simple, hyaline, non-halonate, thin-walled, ovate to broadly ellipsoid, rarely ± globose, (7·0–)8·0–9·8–11·0(–12·0)×5·0–5·9–7·0(–7·5) µm.
Conidiomata not found.
Chemistry
2′–O–methylperlatolic acid; all spot tests negative. Traces of an unidentified fatty acid were detected in some extracts but could be the result of contamination by another lichen species.
Etymology
The specific epithet refers to the unusual excipular morphology of the new species, reminiscent of what is seen in some species of the genus Aspicilia.
Distribution and ecology
This species is known only from Tasmania, where it has been collected on alpine dolerite boulders on the Central Plateau, the largest continuous area of alpine environment on the island. It colonizes the broad surfaces of the largest boulders and this, together with its highly brittle, loosely attached habit, makes it difficult to collect. It is part of a very rich assemblage of alpine saxicolous lichens, the dominant species of which include Lecanora farinacea Fée, Placopsis spp., Poeltiaria coromandelica (Zahlbr.) Rambold & Hertel, Pseudephebe pubescens (L.) M.Choisy, Ramboldia petraeoides, Rhizocarpon geographicum, Rimularia circumgrisea, R. psephota, Stereocaulon caespitosum Redinger, Umbilicaria cylindrica (L.) Delise ex Duby, U. umbilicarioides (B. Stein) Krog & Swinscow and Xanthoparmelia stygiodes.
Remarks
This species is very well characterized by the combination of apothecia with a thalline margin and by its unique chemistry. It grows with and, in the field, could be confused with R. circumgrisea in that both species have a superficially dull grey colour and black apothecia occurring singly or in clusters. However, in practice, they will rarely be mistaken, as the different excipular structure (R. circumgrisea has lecideine apothecia) is easily seen with a hand lens, and the norstictic acid of that species is also easily detected.
The structure of the exciple of R. aspicilioides is remarkable and has been a challenge to interpret. Apothecial initials are observed at an early stage as a circular brown area in the centre of each dark grey thallus areole. The initials gradually widen until they develop into a widely exposed disc, broadly adnate or slightly sunken in the surface of the areole, with the surrounding thalline tissue forming an inrolled margin. With further development, black tissue of the prothallus wraps upwards from beneath and between the areoles to form an enclosing opaque cupulate structure. Superficially one observes a seemingly lecanorine apothecium. However, when examined in section, the result is an apothecium with an opaque black-brown cupulate margin, enclosing thalline tissue containing photobiont cells, which in turn encloses the hymenium. In older apothecia, the lateral parts of the thalline ‘margin’ are excluded and only a subhypothecial layer with photobiont cells remains. I have sectioned and stained numerous apothecia at various stages of development and could not detect any sign of a proper excipulum between the thalline ‘margin’ and the hymenium. A further unusual feature of this species is the paraphyses, which are essentially simple or at best sparsely branched. The chemistry of the species is also very unusual, especially for a genus where most species contain norstictic, stictic, lobaric or gyrophoric acids, or similar substances.
The question to be addressed then is, if not in Rimularia (which is heterogeneous as currently defined), where should this remarkable lichen be placed? Its asci are unequivocally of the Trapelia-type: the shape is very broadly clavate, with a rather bulbous apex and very prominent tholus; the apex of the ascoplasm is at first concave, then truncate and finally rounded, and there is no ocular chamber; the tholus is rather more intensely amyloid than is customary, but nevertheless one can interpret more intensely amyloid flanks and a cap, with the central area only weakly amyloid, all consistent with Rimularia as currently applied. The thin-walled ascospores are likewise consistent with those of other Rimularia species.
One possibility that was considered was the genus Amylora (Rambold Reference Rambold1994; Lumbsch Reference Lumbsch1997). In common with R. aspicilioides, this monotypic genus has aspicilioid apothecia with a reduced proper exciple. Yet it has richly branched paraphyses, amyloid, halonate ascospores and an amyloid medulla, none of which are seen in the new species. Clearly R. aspicilioides is an excellent subject for further study, ideally in the context of a broader investigation of other disparities within Rimularia. For the present, Rimularia is seen as the genus of best fit.
Specimens examined. Australia: Tasmania: Long Tarns, 41°47′S, 146°21′E, 1270 m alt., 2012, G. Kantvilas 30/12A (HO); Skullbone Plains, 42°02′S, 146°19′E, 1000 m alt., 2012, G. Kantvilas 91/12 (HO); Mt Rogoona summit, 41°54′S, 146°12′E, 1350 m alt., 2012, G. Kantvilas 677/12 (HO).
Rimularia asteriphila Kantvilas sp. nov.
MycoBank No.: MB807728
Rimulariae caecae valde similis sed thallo epiphloeodico et ascosporis maioribus, praesertim latioribus, 10–15 µm longis, 7–10 µm latis differens.
Typus: Australia, Tasmania, Johnsons Lagoon, 41°59′S, 146°23′E, 1040 m alt., on Ozothamnus hookeri in subalpine heathland, 9 December 2007, G. Kantvilas 401/07 (holotypus—HO).
(Figs 2A & B, 3B, 4B)

Fig. 2. Tasmanian species of Rimularia. A, R. asteriphila (holotype) habit, associated with Ramboldia laeta (large apothecia lower right) in the foreground; B, R. asteriphila detail; C, R. circumgrisea (holotype) habit; D, R. circumgrisea detail, showing clusters of black, lecideine apothecia; E, R. circumgrisea (GK 403/06) detail, showing apothecia-like squamules with a brown centre and pale greyish margin. Scales: A & C=2 mm; B=250 µm; D & E=1 mm. In colour online.
Thallus crustose, inapparent, endophloeodal, forming discontinuous, undelimited patches to c. 1 cm wide and to 80 µm thick, evident only due to the presence of apothecia; upper cortex ± absent, but with the outermost hyphae becoming brownish, cellular, to 5 µm wide; medulla non-inspersed, I−; photobiont a unicellular green alga (Chlorella?) with irregularly roundish to oblong cells, 7–14× 7–12 µm, encased singly or in pairs or tetrads in a gelatinous sheath 1–2 µm thick. Prothallus absent.
Apothecia lecideine, scattered, basally constricted, 0·2–0·4 mm wide, roundish, occasionally ± angular when crowded together; disc black, plane to undulate, not gyrose; proper exciple black, flush with or minutely higher than the disc, persistent, in section cupular, opaque brown, unchanged in KOH, ±parenchymatous, 12–25 µm thick at the margins, to 30–40 µm thick at the base, not extending as a ‘root’ into the thallus. Hypothecium 25–35 µm thick, pale yellowish brown, not inspersed. Hymenium 50–80 µm thick, colourless at the base, in the upper part overlain by a thick layer of olive-grey, sedifolia-grey pigment, K+ violet, C+ violet, that diffuses downwards between the asci. Paraphyses richly branched and anastomosing, 1–2 µm thick, occasionally with oil vacuoles up to 5 µm wide; apices moniliform, unpigmented, 2·0–3·5 µm thick. Asci 8-spored, broadly clavate, 35–55(–70)×15–27 µm, of the Rimularia-type: with a thin, intensely amyloid outer wall, a prominently thickened tholus with intensely amyloid flanks, an amyloid cap and vestiges of a narrow, weakly amyloid channel in the upper, central part, and lacking an ocular chamber. Ascospores simple, hyaline, non-halonate, thin-walled, ovate to broadly ellipsoid, 10·0–12·1–14·0(–15·0)×7·0–7·9–9·0(–10·0) µm.
Conidiomata not found.
Chemistry
No substances detected by TLC or HPLC; all spot tests negative.
Etymology
The specific epithet refers to the host species of the new lichen being a member of the family Asteraceae. Shrubs and trees of this family tend to produce a fibrous bark that can support lichens not found on other phorophytes.
Distribution and ecology
This species is known only from the type locality where it grew on an isolated mature individual of the shrub Ozothamnus hookeri Sonder, in subalpine woodland bordering a shallow lake. Associated species included Hypogymnia lugubris (Pers.) Krog, Lecanora caesiorubella Ach., Mycoblastus campbellianus (Nyl.) Zahlbr., Ochrolechia xanthostoma (Sommerf.) K. Schmitz & Lumbsch, Ramboldia laeta (Stirt.) Kalb et al., Tephromela sorediata Kalb & Elix and Usnea inermis Motyka. In the context of the Tasmanian highlands, this is a relatively dry highland region, in a virtual rain shadow on the eastern side of the Central Plateau, with an annual average rainfall of only c. 950 mm.
Remarks
Rimularia asteriphila is extremely similar to R. caeca (Lowe) Rambold & Printzen, which is common in north-eastern North America (Rambold & Printzen Reference Rambold and Printzen1992). The two species share a corticolous habit, black, sessile apothecia, an epihymenium containing sedifolia-grey pigment, and a thallus that mostly lacks detectable secondary metabolites. The two taxa differ in the size, and more particularly the width, of their ascospores, with those of R. caeca being 7·5–10·5–13·5×4·5–5·5–7·5 µm (Rambold & Printzen Reference Rambold and Printzen1992); or 7·0–9·4–12·0×5·0–5·6–7·0 µm (this study; see also Fig. 3). Rimularia caeca differs further in having an epiphloeodal, verruculose thallus that is occasionally sorediate.
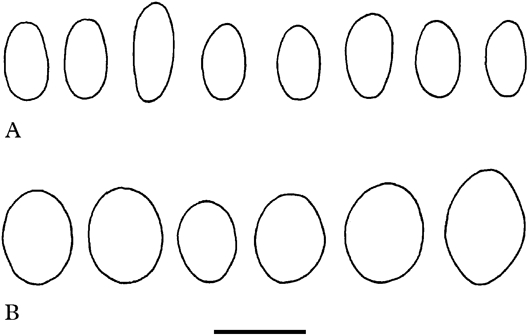
Fig. 3. Comparison of ascospores of A, Rimularia caeca (Wetmore 87339) and B, R. asteriphila (holotype). Scale=10 µm.
Scant though the available material of this new species is, it nevertheless deserves description on account of its clear-cut combination of characters. Furthermore, searches for additional collections have proved unsuccessful. Rimularia is predominantly a saxicolous genus and corticolous species are relatively few. In Tasmania, there is R. applanata Kantvilas & Coppins, which contains lobaric acid and has broad, adnate apothecia and globose ascospores. In addition to R. caeca, the only other corticolous Rimularia species is the Northern Hemisphere's R. fuscosora Muhr & Tønsberg, which is sorediate and contains norstictic acid. Epihymenial sedifolia-grey pigment occurs in several other species (R. psephota, R. subpsephota Fryday and R. ramboldiana Kantvilas & Elix), but all are saxicolous and, with the exception of R. subpsephota, contain secondary metabolites.
In the field, all that is detectable of R. asteriphila are the black, basally constricted, lecideine apothecia with an entire (not gyrose) disc. Hence, there is little indication that it is a Rimularia at all, and it is more likely to be confused with Buellia s. str., several species of which occur in such habitats.
Rimularia circumgrisea Kantvilas sp. nov.
MycoBank No.: MB807729
Rimulariae psephotae fortasse comparanda ob acidum norsticticum continens sed ab illa areolis concavis marginibus griseis, medulla non-amyloidea, apotheciis lecideinibus, planis, non-gyrosis, 0·2–1·2 mm latis, epihymenio pigmentum sedifolia-griseum destituto, et ascosporis parvioribus, 8·5–14·0 µm longis, 5·0–6·5 µm latis differens.
Typus: Australia, Tasmania, Long Tarns, 41°47′S, 146°21′E, 1270 m alt., on alpine dolerite boulders, 17 February 2012, G. Kantvilas 30/12 (holotypus—HO; isotypus—E).

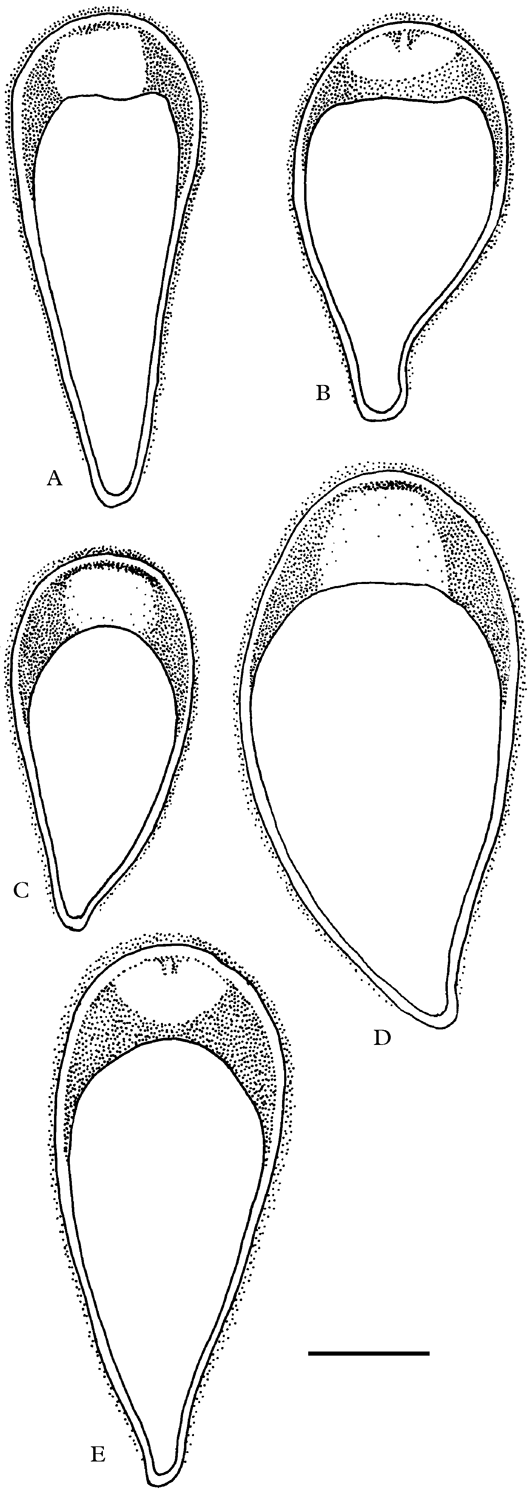
Fig. 4. Asci of Tasmanian species of Rimularia (amyloid parts after pretreatment with KOH stippled). A, R. aspicilioides; B, R. asteriphila; C, R. circumgrisea; D, R. coppinsiana; E, R. psephota. Scale=10 µm.
Thallus crustose, minutely squamulose to areolate, forming irregular, widely spreading patches to 10 cm wide or more. Individual areoles 0·2–0·8(–1·0) mm wide, c. 0·25–0·30 mm thick, at first ± neatly roundish, concave and dispersed, pale fawn brown to dark brown, glossy, typically with a pale bluish grey, inrolled, somewhat thickened, crenulate margin, later irregular in outline, undulate, sometimes deeply cracked in a polygonal pattern, becoming crowded together in irregular clumps up to 2 mm thick and 3 mm wide; upper cortex consisting of ±vertically orientated, loosely entangled, brownish hyphae 2–4 µm thick; photobiont a unicellular green alga with globose cells 10–16 µm diam., encased in a gelatinous sheath 1–2 µm thick; medulla I−, inspersed with minute crystals that fluoresce in polarized light, dissolve in KOH and form red, needle-like crystals (norstictic acid). Prothallus black, crustose, loosely attached to the substratum, in section K+ brownish red, here and there with a patchy blue-green, N+ crimson pigment.
Apothecia lecideine, uncommon, usually occurring in clusters, broadly adnate, sessile or nestled amongst the areoles, 0·2–1·2 mm wide, roundish to rather angular and distorted when crowded together, often fused to form compound apothecia 2–3 mm wide; disc black, plane to slightly convex, sometimes cracked but not gyrose; proper exciple black, glossy, very thin, flush with or minutely higher than the disc, sometimes barely evident, flexuose, in section cupular, opaque dark brown-black to crimson-black, K+ intensifying crimson, at the outer edge also with blue-green, N+ crimson pigment, 20–60 µm thick at the margins, thicker at the base and extending as a ‘root’ into the thallus. Hypothecium 30–50 µm thick, colourless to pale yellowish brown, inspersed with oil droplets, forming red needle-like crystals in KOH. Hymenium 50–60 µm thick, separating easily in KOH, mostly colourless but with the uppermost 5–10 µm yellow-brown, K± olive-brown. Paraphyses 1·5–2·0 µm thick, sparsely branched and anastomosing, sometimes slightly capitate; apices 2·5–4·0 µm thick, with an external brown cap. Asci 8-spored, broadly clavate, 27–44×13–16 µm, approximating the Trapelia-type: with a thin, intensely amyloid outer wall, a prominently thickened tholus with intensely amyloid flanks, weakly amyloid in the central part, and lacking an ocular chamber. Ascospores simple, hyaline, non-halonate, thin-walled, ellipsoid, ovate to rather oblong, (8·5–)9·0–10·8–13·0(–14·0)×5·0–5·4–6·0(–6·5) µm.
Conidiomata not found.
Chemistry
Norstictic and connorstictic acids; although the medulla reacts P+ orange, K+ red, norstictic acid is best detected in thin sections of the thallus and apothecia which develop red, needle-shaped crystals following elution with KOH.
Etymology
The specific epithet refers to the inrolled grey margin of the areoles.
Distribution and ecology
Rimularia circumgrisea is known only from Tasmania, where it occurs at alpine elevations on large dolerite outcrops. It is extremely abundant at the type locality on Tasmania's Central Plateau, where it occurs as part of a very rich association of saxicolous alpine crustose lichens, including Buellia poimenae Elix & Kantvilas, Catillaria contristans (Nyl.) Zahlbr., Lecanora cf. demersa, L. lugubris (C. W. Dodge) D. J. Galloway, Lepraria caesioalba (de Lesd.) J. R. Laundon, Placopsis spp., Rhizocarpon geographicum, Rimularia aspicilioides Kantvilas and R. psephota; associated macrolichens include Usnea torulosa (Müll. Arg.) Zahlbr. and species of Xanthoparmelia and Umbilicaria. It frequently occurs in shallow, concave depressions on the largest boulders (as distinct from growing on rock edges or pinnacles), a microhabitat that perhaps offers slight shelter from abrasive winds and, temporarily, more moisture. This microhabitat also means it is frustratingly difficult to collect, and hence it is poorly represented in herbarium collections.
Remarks
Given its alpine, saxicolous habitat and the presence of norstictic acid, this species could be compared with R. psephota. However, beyond these ecological and chemical features, the two taxa have little in common and should not be confused. The thallus of Tasmanian collections of R. psephota is areolate but usually distinctly bullate, and never with the areoles concave and with grey, inrolled margins; furthermore, its medulla is I+ violet. The apothecia of the two species are also starkly different, with those of R. psephota often being gyrose and rather angular in outline, and with a prominent, persistent, inrolled, proper margin. The epithecium of R. psephota contains sedifolia-grey pigment and reacts K+ violet, C+ violet. Furthermore, the ascospores of the two taxa are markedly different, with those of R. psephota incrementally larger: 12–21×6·5–13·0 µm (this study). Superficially far more similar is R. aspicilioides in so far as both it and R. circumgrisea have a dark grey, loosely adnate, areolate thallus. Indeed both species may occur together and the apothecia with a pale grey, inrolled thalline margin of the former may resemble the sterile areoles of the latter. However, with care, the two species are easily distinguished morphologically at low magnification; they also differ chemically.
Norstictic acid is a frequent metabolite in the genus Rimularia, occurring either alone or together with gyrophoric or stictic acids (Hertel & Rambold Reference Hertel and Rambold1990; Giavarinni & David Reference Giavarini, David, Smith, Aptroot, Coppins, Fletcher, Gilbert, James and Wolseley2009). However, based on comparisons with available herbarium material and published descriptions, no norstictic acid-containing species agrees with R. circumgrisea, especially with regard to its distinctive thallus morphology, plane apothecia with their very thin margin, and relatively small ascospores. Two alpine Tasmanian collections (Kantvilas 275/98, 71/09) containing norstictic acid remain enigmatic and are included under R. circumgrisea with some hesitation. They have apothecia like the new species, but their thallus areoles are minutely lobate, plane and without the typical inrolled grey margins. Yet they have the small ascospores of R. circumgrisea, as well as an I− medulla and yellow-brown epithecium. Both specimens are in rather poor condition and this may be responsible for their unusual morphology.
Specimens examined. Australia: Tasmania: Wylds Craig, 42°28′S, 146°23′E, 1330 m alt., 1998, G. Kantvilas 275/98 (HO); Cathedral Mountain, 41°53′S, 146°06′E, 1380 m alt., 1999, G. Kantvilas 89/99 (HO); Long Tarns, 41°47′S, 146°21′E, 1270 m alt., 2006, G. Kantvilas 403/06 (HO); Pine Lake, 41°45′S, 146°42′E, 1230 m alt., 2009, G. Kantvilas 71/09 (HO); Snowdrift Tarns, 42°55′S, 146°39′E, 1270 m alt., 2012, G. Kantvilas 263/12 (HO); Mountains of Jupiter summit, 41°57′S, 146°11′E, 1320 m alt., 2012, G. Kantvilas 645/12, 652/12 (HO); Skullbone Plains, c. 200 m from road intersection to Nive River, 42°02′S, 146°21′E, 970 m alt., 2012, G. Kantvilas 727/12 (HO); Mt Styx, 146°45′S, 42°51′E, 1090 m alt., 2012, G. Kantvilas 744/12 (HO).
Rimularia coppinsiana Kantvilas sp. nov.
MycoBank No.: MB807730
Rimulariae limborinae similis sed thallo acidum bourgeanicum continenti et ascosporis valde parvioribus, 10–19 µm longis, 7–12 µm latis differt.
Typus: Australia, Tasmania, Falling Mountain, 41°55′S, 146°06′E, 1350 m alt., on soft, weathered, moist dolerite rock face in alpine heathland, 11 March 2011, G. Kantvilas 141/11 (HO—holotypus; E—isotypus).

Fig. 5. Tasmanian species of Rimularia. A, R. coppinsiana (holotype) habit; B, R. coppinsiana detail, showing umbonate and gyrose apothecia; C, R. psephota (GK 110/12) habit; D, R. psephota detail, showing bullate thallus with contorted, entire, gyrose or umbonate apothecia. Scales: A & C=2 mm; B & D=1 mm. In colour online.
Thallus crustose, rimose-areolate, forming spreading, continuous, undelimited patches to 10 cm wide. Individual areoles 0·1–2·0 mm wide, 0·3–0·7 mm thick, irregularly roundish, plane to convex, aggregated together in irregular to rather angular polygonal clusters 0·5–1·5 mm wide, separated by deep cracks, pale brownish to grey-brown, here and there with an orange tinge (possibly due to discoloration from the substratum); upper cortex poorly defined, 10–15 µm thick, composed of entangled, short-celled hyphae 2–4 µm thick; photobiont a unicellular green alga with roundish to ellipsoid cells, 7–20×8–12 µm, encased in a gelatinous sheath 1–2 µm thick; medulla I−, inspersed with minute colourless crystals that fluoresce in polarized light and dissolve in KOH. Prothallus black, crustose, subtending the areoles, in section opaque reddish brown, K+ intensifying reddish, comprising amorphous hyphae interspersed in the upper part with dead algal cells.
Apothecia lecideine, adnate to superficial, 0·4–2·0 mm wide, roundish, irregularly lobate, convoluted or elongate, occurring singly or in fused clusters; disc black, concave to plane, mostly entire, sometimes gyrose or umbonate; proper exciple black, glossy, inrolled, often radially cracked, persistent, in section cupular, opaque dark reddish brown, K± intensifying reddish, 25–60 µm thick at the margins, to 150 µm thick or more at the base and extending as a ‘root’ into the thallus. Hypothecium 40–60 µm thick, pale reddish brown, I−, not inspersed. Hymenium 100–130 µm thick, mostly colourless, dilutely reddish brown in the upper part, separating readily in KOH. Paraphyses 1·5–2·5 µm thick, branched and anastomosing; apices moniliform, to 3·5 µm wide, colourless. Asci 8-spored, clavate, 55–70×22–25 µm, of the Trapelia-type: with a thin, amyloid outer wall, a prominently thickened tholus, faintly amyloid at the flanks and apex, ±non-amyloid in the central part, and lacking an ocular chamber. Ascospores simple, hyaline, non-halonate, thin-walled, subglobose, to ellipsoid to oblong, (10·0–)12·0–14·2–17·0(–19·0)×7·0–9·2–11·0(–12·0) µm.
Conidiomata not found.
Chemistry
Bourgeanic acid; thallus K−, KC−, C−, P−, UV−.
Etymology
The specific epithet honours Brian Coppins, a good friend and mentor, and collaborator on several projects, past and present. It is a particular pleasure to dedicate to Brian a species of Rimularia, a genus to which he has made significant contributions.
Distribution and ecology
This new species is known only from the type locality, a dolerite peak in western Tasmania. Alpine dolerite is a rich habitat for lichens and, in particular, for species of Rimularia; for example, R. albotessellata, R. aspicilioides, R. circumgrisea and R. psephota. However, whereas these species are found in very exposed habitats where the microclimate ranges from very hot to extremely cold with exposure to ice, wind, snow and rain, regardless of the season, R. coppinsiana was found in a sheltered, moist cleft on the relatively soft, eroding surface of a large boulder. No other lichen species were observed in this rather unusual habitat. The habitat is not unlike that of R. umbratilis Kantvilas & Coppins, which occurs in sheltered crevices and underhangs on quartzitic rocks in Tasmania's south-west.
Remarks
This is a very distinctive lichen for the Tasmanian mycota. Even in the field, the gyrose-umbonate apothecia immediately suggest it to be a Rimularia, and the presence of bourgeanic acid is unique for the genus. The morphology of the apothecia is superficially similar to the Northern Hemisphere's R. limborina (H. Magn.) Coppins, which differs by containing gyrophoric acid and by having Rimularia-type asci and significantly larger ascospores [(14–)18–30(–40)×(5–)10–18(–21) µm; Giavarini & David (Reference Giavarini, David, Smith, Aptroot, Coppins, Fletcher, Gilbert, James and Wolseley2009)].
Also Studied
Rimularia psephota (Tuck.) Hertel & Rambold
In Hertel, Mitt. Bot. Staatssamml. München 23: 334 (1987).
(Figs 4E, 5A & B)
Thallus areolate, pale to dark grey, sometimes brownish grey, matt, closely adnate to the substratum; areoles 0·15–0·50 mm wide, convex to bullate; prothallus mostly lacking, or black, effuse, marginal and/or visible between the areoles; medulla I+ violet.
Apothecia to 0·9 mm wide, lecideine, roundish to lobate or elongate, sessile to basally constricted; disc black, plane, usually gyrose; proper exciple prominent, persistent, distinctly elevated above the level of the disc, in section 25–80 mm thick. Epithecium greenish brown, K+ crimson, C+ crimson (sedifolia-grey). Paraphyses richly branched and anastomosing. Asci broadly clavate, 45–65× 20–35 µm, of the Rimularia-type. Ascospores broadly ellipsoid to ovate, very thin-walled and easily misshapen by mutual pressure, hyaline but with occasional spores discoloured brownish, 12·0–16·5–21·0×(6·5–)7·0–9·4–12·0(–13·0) µm.
Conidiomata not seen.
Chemistry
Norstictic and connorstictic acids; these compounds are best detected under the microscope, where squashes of the thallus and apothecia produce red, needle-shaped crystals upon the addition of KOH.
Remarks
The above description is based exclusively on Tasmanian collections and has been compiled de novo. This species is widely distributed in cold environments in the Southern Hemisphere (including Antarctica, Argentina, Chile, Îles Kerguelen, New Zealand), with isolated occurrences in similar environments in the Northern Hemisphere (Iceland, Svalbard). Further descriptive data are provided by Rambold (Reference Rambold1989), Hertel & Rambold (Reference Hertel and Rambold1990) and Øvstedal et al. (Reference Øvstedal, Tønsberg and Elvebakk2009). In Tasmania, it is widespread and locally common at alpine elevations where it occurs on exposed rocks, typically with Cameronia pertusarioides, Hymenelia sp., Trapelia lilacea, Usnea torulosa, Xanthoparmelia stygiodes and several other Rimularia species. Although most collections are from Jurassic dolerite and Tertiary basalt, it has also been recorded, more rarely, on Devonian granite, Precambrian quartzite and Ordovician conglomerate. The combination of a well-developed, saxicolous, norstictic acid-containing thallus, an epithecium with sedifolia-grey pigment, and relatively large ascospores characterize this distinctive species and distinguish it from, for example, R. circumgrisea (norstictic acid, no sedifolia-grey pigment, smaller ascospores) and R. asteriphila (no substances, smaller ascospores). Further comparative observations are provided under the former species, with which it may be associated and with which it shares some features. Although Hertel (Reference Hertel1984) erected the genus Lambiella for this species, Hertel & Rambold (Reference Hertel and Rambold1990) subsequently transferred it to Rimularia; it may well be that Lambiella will be resurrected when the relationships of taxa within the currently broad concept of Rimularia are reviewed.
Selected specimens examined. Australia: Tasmania: Mt Campbell, 41°39′S, 145°59′E, 1972, G. C. Bratt 72/1227a (HO); Lake Kaye, 41°54′S, 146°31′E, 1140 m alt., 2000, G. Kantvilas 90/00 (HO); Sandbanks Tier, 41°51′S, 146°52′E, 1400 m alt., 2000, G. Kantvilas 335/00 (HO); Black Bluff, 41°27′S, 145°57′E, 1330 m alt., 2000, G. Kantvilas 149/00 (HO); Pine Lake, 41°45′S, 146°42′E, 1230 m alt., 2009, G. Kantvilas 73/09 (HO); c. 0·5 km SE of Lake Ada, 41°53′S, 146°29′E, 1150 m alt., 1999, G. Kantvilas 426/99 (HO); c. 1 km N of Lake Ironstone, 41°42′S, 146°28′E, 1190 m alt., 2005, G. Kantvilas 336/05 (HO); Mersey Crag, 41°46′S, 146°20′E, 1420 m alt., 2010, G. Kantvilas 68/10 (HO); c. 3·5 km SE of Mt Meredith, 41°35′S, 145°17′E, 750 m alt., 2011, G. Kantvilas 69/11 (HO); Mt Byron, 42°02′S, 146°04′E, 1370 m alt., 2011, G. Kantvilas 167/11 (HO); Skullbone Plains, 42°02′S, 146°19′E, 1000 m alt., 2012, G. Kantvilas 110/12, 130/12 (HO); northern summit of Mt Rogoona, 41°53′S, 146°12′E, 1330 m alt., 2012, G. Kantvilas 670/12 (HO); Mt Styx, 146°45′S, 42°51′E, 1090 m alt., 2012, G. Kantvilas 745/12 (HO).
Provisional key to the Tasmanian species of Rimularia
-
1 Occurring on bark, wood, humus-rich soil or over bryophytes and other lichens ... 2
Occurring directly on rock ... 5
-
2(1) Thallus glossy pale blue-grey to grey, composed of rather effigurate, scattered or contiguous areoles over a conspicuous black prothallus; apothecia broadly adnate, to 1·7 mm wide, with a markedly lobate to flexuose margin; containing lobaric acid ... 3
Thallus whitish or pale cream and areolate-verruculose, or inapparent; prothallus lacking; apothecia basally constricted, to 0·4 mm wide, single or in botryose clusters; containing porphyrilic acid or lacking secondary metabolites ... 4
-
3(2) Ascospores broadly ellipsoid, 6·0–10·5×4–6 µm; found on shaded, smooth bark in cool temperate rainforest ... R. applanata
Ascospores globose or almost so, 5–8×4·5–7·5 µm; found on soil or overgrowing terricolous lichens and bryophytes in open habitats ... R. globulispora
-
4(2) Thallus whitish or pale cream, areolate-verruculose, containing porphyrilic acid; apothecia typically in botryose clusters of up to 20 or more; epihymenium olive-brown, K−, C−; ascospores 7–13×4–6 µm; common in wet forest, especially at high elevations, growing on soft, moist bark at the base of large trees, or on mounds of humus and peat ... R. hepaticicola
Thallus inapparent, lacking secondary metabolites; apothecia solitary, scattered; epihymenium grey, K+ violet, C+ violet; ascospores 10–15×7–10 µm; very rare in heathland, growing on fibrous bark ... R. asteriphila
-
5(1) Epihymenium greyish, K+ violet, C+ violet ... 6
Epihymenium a shade of olive or brown, never reacting violet in K and/or C ... 7
-
6(5) Thallus olive-brown, containing gyrophoric acid (K−, C+ reddish in squash preparations), typically overgrowing the thallus of Ramboldia blastidiata Kantvilas & Elix in dry sclerophyll woodland; ascospores 9·5–14·0×6–9 µm ... R. ramboldiana
Thallus pale to dark grey or brownish grey, containing norstictic acid (forming red, needle-shaped crystals upon the addition of KOH to squash preparations of the thallus and apothecia, C−), autonomous; found on exposed rocks on the highest peaks; ascospores 12–21×6·5–13·0 µm ... R. psephota
-
7(5) Found in low rainfall areas on exposed, dry rocks; thallus olive-brown, areolate to subsquamulose, becoming plicate and pulvinate, containing confriesiic acid (all reactions negative); ascospores 7–12×6·5–11·0 µm ... R. campestris
Confined to highland areas; thallus areolate to squamulose, never plicate-pulvinate; secondary chemistry variable but never with confriesiic acid ... 8
-
8(7) Thallus vivid white, containing pannarin (P+ pale orange, K+ pale yellow) ... R. albotessellata
Thallus a shade of dull grey or brown; chemical composition and spot tests variable but never including pannarin ... 9
-
9(8) Growing in moist, sheltered, shaded crevices ... 10
Growing on highly exposed rock surfaces ... 11
-
10(9) Thallus very thin (<90 µm), tightly adnate, pale blue-grey to brown-grey and rather dispersed over a conspicuous black prothallus; lobaric acid present; ascospores 9·5–13·0×4–7 µm; occasional on highly siliceous, hard quartzitic rocks ... R. umbratilis
Thallus >300 µm thick, rather loosely attached, continuous and obscuring the prothallus; bourgeanic acid present; ascospores 10–19×7–12 µm; very rare on weathered dolerite ... R. coppinsiana
-
11(9) Apothecia lecideine, with a thin proper exciple; norstictic acid present (squash preparations of the thallus and apothecia yielding red, needle-shaped crystals with the addition of KOH) ... R. circumgrisea
Apothecia aspicilioid, lacking a proper exciple but surrounded by a thin, inrolled thalline margin; 2′-O-methylperlatolic acid present (K−) ... R. aspicilioides
For their companionship in the field in search of Rimularia specimens, I thank Brigitte de Villiers, Ken Felton and Jean Jarman. Assistance with chemical problems from Professor Jack Elix, who identified or confirmed the identity of several critical compounds, is gratefully acknowledged. I especially thank Jean Jarman, who photographed all the species and prepared the illustrations for publication. For helpful comments on aspects of the work, including an earlier version of this paper, I thank Alan Fryday and an anonymous referee. This project was supported by the Australian Biological Resources Study through the award of an Applied Taxonomy Grant.