INTRODUCTION
Recent molecular biodiversity studies increasingly reveal local genetic differentiation in biphasic marine species with a larval dispersal phase (e.g. Mathews, Reference Mathews2006; Galarza et al., Reference Galarza, Carreras-Carbonell, Macpherson, Pascual, Roques, Turner and Rico2009; Ragionieri et al., Reference Ragionieri, Fratini, Vannini and Schubart2009; Fratini et al., Reference Fratini, Ragionieri, Deli, Harrer, Marino, Cannicci, Zane and Schubart2016). Geographic barriers are the main historical factor determining the biogeography of European marine species, as documented for caridean shrimps (Reuschel et al., Reference Reuschel, Cuesta and Schubart2010; González-Ortegón et al., Reference González-Ortegón, Palero, Lejeusne, Drake and Cuesta2016). The most prominent geographic barrier in European marine waters is the one between the Atlantic Ocean and the Mediterranean Sea. It is located somewhere in between the Alborán Sea and the Gulf of Cádiz, and either the Strait of Gibraltar or the Almería-Oran Front have been postulated to act as the main barrier, as recently revised and discussed (Patarnello et al., Reference Patarnello, Volckaert and Castilho2007; García-Merchán et al., Reference García-Merchán, Robainas-Barcia, Abelló, Macpherson, Palero, García-Rodríguez, Gil de Sola and Pascual2012). In recent geological history, the shallow Strait of Gibraltar isolated the Mediterranean Sea repeatedly from the Atlantic, caused by sea level regressions, sometimes even resulting in major desiccations (Hsü et al., Reference Hsü, Montadert, Bernoulli, Cita, Erickson, Garrison, Kidd, Mèlierés, Müller and Wright1977). These repeated isolations led to a very high level of endemism in the Mediterranean Sea (Hofrichter, Reference Hofrichter2002), and many marine species show a phylogeographic break among Atlantic and Mediterranean populations (Zane et al., Reference Zane, Ostellari, Maccatrozzo, Bargelloni, Cuzin-Roudy, Buchholz and Patarnello2000; Patarnello et al., Reference Patarnello, Volckaert and Castilho2007; Luttikhuizen et al., Reference Luttikhuizen, van Bleijswijk, Peijnenburg and van der Veer2008; Deli et al., Reference Deli, Fratini, Ragionieri, Said, Chatti and Schubart2016). The Messinian Salinity Crisis in the late Miocene (around 5.5 Ma) was probably the most dramatic isolation event (Krijgsman et al., Reference Krijgsman, Hilgen, Raffi, Sierro and Wilson1999) and ended with the Zanclean Flood, i.e. when Atlantic waters re-flooded the Mediterranean Basin, leading to a re-colonization of the Mediterranean Sea with Atlantic species (García-Castellanos et al., Reference García-Castellanos, Estrada, Jiménez-Munt, Gorini, Fernàndez, Vergés and De Vicente2009). Even today, the waters between the Strait of Gibraltar and the Almería-Oran Front seem to act as a barrier to gene flow for many marine species, determining the genetic structure and diversity of many European coastal water species.
The shrimp genus Palaemon Weber, 1795 (Crustacea: Decapoda: Caridea) belongs to the large family Palaemonidae Rafinesque, 1815 and includes many important representatives from coastal waters all over the world. Recently, important systematic and taxonomic changes were postulated for this family, summarized in the review by De Grave & Ashelby (Reference De Grave and Ashelby2013), with the consequence that the genera Palaemonetes Heller, 1869, Exopalaemon Holthuis, 1950 and Coutierella Sollaud, 1914 were synonymized and re-arranged within the genus Palaemon that now contains 83 species (for the most recent phylogeny see Carvalho et al., Reference Carvalho, De Grave and Mantelatto2016). Morphological distinction within this genus is partly difficult, because only small differences serve as identification criteria, as for example the upper antennular flagellum or the denticulation of the rostrum (González-Ortegón & Cuesta, Reference González-Ortegón and Cuesta2006). This often results in wrongly classified species within the genus Palaemon. For example, Palaemon garciacidi Zariquiey Álvarez, 1968 turned out to be a synonym of Palaemon longirostris Milne-Edwards, 1837 (Cuesta et al., Reference Cuesta, Drake, Martínez-Rodríguez, Rodríguez and Schubart2012; Cartaxana, Reference Cartaxana2015). Also the existence of cryptic species is discussed for this genus, since Reuschel et al. (Reference Reuschel, Cuesta and Schubart2010) gave evidence for differentiation along the Atlantic–Mediterranean barrier within Palaemon elegans Rathke, 1837 and the possible existence of a cryptic species within the Mediterranean Sea. Thus, it seems worthwhile to use genetic techniques to study phylogeographic relationships between Atlantic–Mediterranean populations of other coastal species of Palaemon and to help redefine the morphological classification in this taxonomic group. It is of particular interest to focus on species living in both the Atlantic Ocean and the Mediterranean Sea, to determine if similar differentiation patterns may be revealed as in P. elegans. In Europe, six marine and/or estuarine species (including the recently introduced P. macrodactylus Rathbun, 1902 (see González-Ortegón et al., Reference González-Ortegón, Cuesta and Schubart2007; Ashelby et al., Reference Ashelby, De Grave and Johnson2013)) and four fresh to brackish water species of Palaemon can be found (Cuesta et al., Reference Cuesta, Drake, Martínez-Rodríguez, Rodríguez and Schubart2012). The former ones include the widespread and commercially exploited species Palaemon serratus (Pennant, Reference Pennant1777), also known as the common prawn. It could make an interesting study object regarding phylogeographic analyses, as a recent published record indicated that a single Mediterranean specimen from the Turkish coast may be distinct from its Atlantic counterparts (Bilgin et al., Reference Bilgin, Utkan, Kalkan, Karhan and Bekbolet2014).
Palaemon serratus lives in rocky crevices along the European and North African coastlines in shallow waters up to a depth of 40 m. Its distribution area ranges from Scotland and Denmark to Mauritania and it is common in the Atlantic Ocean, with occasional records in the Mediterranean Sea, and the Black Sea (Lagardère, Reference Lagardère1971; d'Udekem d'Acoz, Reference d'Udekem d'Acoz1999). Adult individuals have a total length of 90–110 mm and the cephalothorax and abdomen are transparent with a dark reddish-brown striped pattern which can vary widely between different regions or habitats (Carlisle, Reference Carlisle1955; González-Ortegón & Cuesta, Reference González-Ortegón and Cuesta2006). Larval development is linked to initial larval body mass and female body size and can be modified by environmental conditions experienced by the larvae (Reeve, Reference Reeve1969; González-Ortegón & Giménez, Reference González-Ortegón and Giménez2014).
There is an important commercial fishery on P. serratus, especially around the British Isles, France and northern Spain, and the high commercial value could possibly lead to overfishing, as already expressed by Fahy et al. (Reference Fahy, Forrest, O'Toole, Mortimer and Carroll2006). Even though the latter study only evaluated the fisheries around Ireland, the same problems could also occur elsewhere. Currently, baseline monitoring data are gathered in the UK and Ireland to ascertain the stock status and population trends and to identify possible problems caused by the fishery exploitation (Haig et al., Reference Haig, Ryan, Williams and Kaiser2014). Although this species was proposed as suitable for cultivation (Reeve, Reference Reeve1968; Rodríguez, Reference Rodríguez1981), further studies about its ecology and husbandry conditions are necessary to take this step. For this endeavour, basic knowledge on the natural genetic diversity and the phylogeography of this species will be critically important.
This study thus focused on the genetic differentiation among various populations of P. serratus along European coastlines. DNA sequences for the mitochondrial gene Cox1 were analysed. To confirm the obtained results additional sequences of the more conserved mitochondrial 16S rRNA and the nuclear gene Enolase were included. Cox1 and 16S rRNA have been shown to be suitable marker genes for DNA barcoding in crustaceans (Schubart et al., Reference Schubart, Neigel and Felder2000; Lefébure et al., Reference Lefébure, Douady, Gouy and Gibert2006; Costa et al., Reference Costa, de Waard, Boutillier, Ratnasingham, Dooh, Hajibabaei and Hebert2007) and also within the Palaemonidae (Reuschel et al., Reference Reuschel, Cuesta and Schubart2010; Cartaxana, Reference Cartaxana2015). Special attention is paid to the possible divergence among Atlantic and Mediterranean populations, as previously reported for the closely related prawn P. elegans (see Reuschel et al., Reference Reuschel, Cuesta and Schubart2010).
MATERIALS AND METHODS
Specimens of Palaemon serratus used for the analysis originated from the authors' collections and were stored in 70–95% ethanol. Most individuals were sampled by hand with the aid of a dip net. Sampling localities, coordinates and number of sequences for each available population of P. serratus are shown in Figure 1 and Table 1. For some populations, only the locality was given and the coordinates were estimated. DNA-extractions were performed following either the Puregene Method (from Gentra Systems: Minneapolis, MN55447, USA) or the Realpure – Spin Kit 250 Extract (from Durviz – Gentaur: Brussels, Belgium).

Fig. 1. Sampling localities of Palaemon serratus (Pennant, Reference Pennant1777). The locality numbers correspond to the code numbers in Table 1.
Table 1. Coordinates and number of individuals of the studied populations of Palaemon serratus from west to east.

A, Atlantic Ocean; M, Mediterranean Sea.
Two different mitochondrial DNA gene fragments were amplified by means of PCR reactions: the mitochondrial gene Cox1 encoding subunit 1 of the cytochrome c oxidase gene that is especially suitable for intraspecific comparisons and used as the biological barcoding gene (e.g. Costa et al., Reference Costa, de Waard, Boutillier, Ratnasingham, Dooh, Hajibabaei and Hebert2007) (N = 67 sequences). In addition, one individual of each of five Atlantic populations and four individuals of Mediterranean populations were analysed for variation of the more conserved 16S rRNA gene, transcribed to the structural rRNA of the large subunit 16S of mitochondrial ribosome. The only available 16S-sequence in GenBank (JQ042291) was downloaded and added to the analysis, allowing comparison of a total of 10 sequences. In addition, the nuclear gene Enolase, better suited for interspecific comparisons (e.g. Ip et al., Reference Ip, Schubart, Tsang and Chu2015), was amplified for some of the Mediterranean and Atlantic populations (N = 26 sequences).
For both mitochondrial genes, different primer combinations were used. For Cox1, a few long fragments of 1276 basepairs (bp) were amplified with primers COL6 or COL6a and COH1b. This was not possible for the Mediterranean individuals, so a shorter fragment was amplified using the primers COL1Pe and COH1b (630 bp). For the amplification of the 16S rRNA gene, the forward primer 16L29 and the reverse primer 1472 were used (~580 bp). The nuclear gene Enolase was amplified using the primer combination ENEA1 and ENES1 (409 bp). All primer sequences and the corresponding references are listed in Table 2.
Table 2. DNA primer sequences and corresponding references.

PCR reactions were run in a total volume of 25 µl with different protocols optimized for the respective Taq polymerase (Fermentas Taq, Promega GoTaq or Roche Taq). The temperature profile of the PCR for all three genes consisted of an initial denaturation step at 94°C for 4 min, followed by 35–40 cycles and a final elongation step at 72°C for 5–10 min. The cycle program included denaturation at 95°C for 45 s, annealing at 52°C for the primer combination COL6 or COL6a/COH1b, 48°C for all other mitochondrial gene combinations, and 50°C for Enolase for 45–60 s, and elongation at 72°C for 60 (short fragments) or 90 (long fragments) seconds. In a few difficult cases, the annealing temperature was decreased to 45°C to obtain useable PCR products.
Most of the long fragments of the Cox1 gene were sequenced in both directions, while the short fragments were only sequenced with COH1b. The 16S rRNA fragments were sequenced with 16L29 and the Enolase fragments with ENEA1. Sanger sequencing took place at the Laboratory of Molecular Biology, SAI (Servicios de Apoyo a la Investigación; University of A Coruña, Spain) or was outsourced to Macrogen Europe (the Netherlands). Sequence chromatograms were proofread using Chromas Lite version 2.1.1 (Technelysium Pty Ltd) and edited manually, if necessary. All sequences were aligned with BioEdit version 7.2.0 (Hall, Reference Hall1999) using the ClustalW algorithm (Thompson et al., Reference Thompson, Higgins and Gibson1994) and, if necessary, adjusted manually. Five different alignments were created, three with the Cox1 sequences, depending on the primer combinations and sequence length, and one in each case for the 16S and the Enolase datasets. No ambiguities were encountered during the alignment process. Before building the parsimony network, the alignments were cropped manually to the same length and short sequences were eliminated. All new DNA sequences (all haplotypes in case of Cox1) were submitted to the European Nucleotide Archive (ENA as part of GenBank) and have been assigned accession numbers LT717247 to LT717310. Alignments were converted from FASTA to PHYLIP format with FaBox version 1.41 (URL 1). Finally, five networks were constructed with the software TCS version 1.21 (Clement et al., Reference Clement, Posada and Crandall2000) based on the single alignments. In the three Cox1 networks, the maximum connection steps had to be specified manually (65–130 steps) in order to allow the haplogroups to be connected, while in the Enolase network a 95% and in the 16S network a 94% connection limit between the different genotypes were used. Nucleotide and haplotype diversities were calculated with the software DnaSP version 5.10.01 (Librado & Rozas, Reference Librado and Rozas2009) for six different populations (only populations with more than two individuals were included). A permutation test was performed with 10,000 replicates. Also a mismatch analysis of sequences was carried out with DnaSP version 5.0 for the Cox1 and for the 16S alignments containing Mediterranean individuals. A constant population size was chosen as the model for expected values. Finally, an analysis of molecular variance (AMOVA) was carried out for two of the populations of the Mediterranean (with more than eight individuals) and the population of Guadalquivir using the software DnaSP version 5.0.
RESULTS
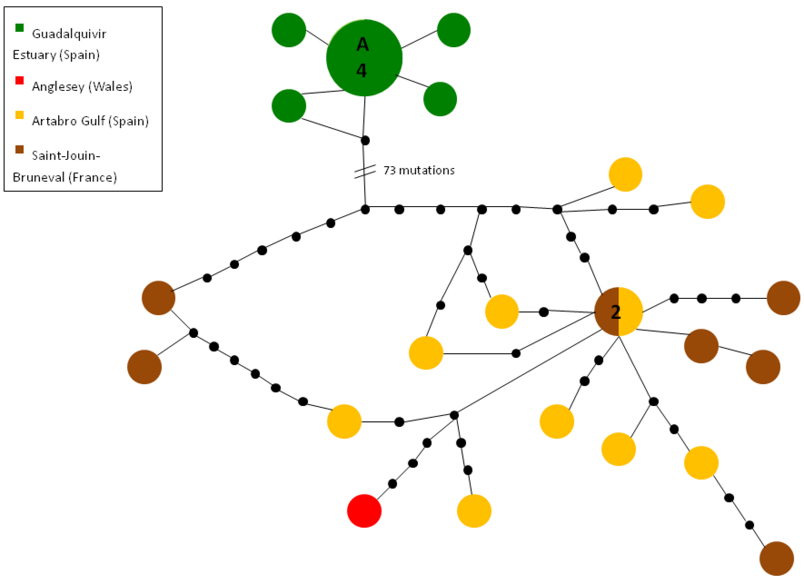
For many of the long fragments of Cox1, clean reads were only obtained for one direction. Therefore, sequences were subdivided into three length groups of which three different alignments and three different parsimony networks were constructed. The first alignment consists of 17 consensus sequences of forward and reverse reads of the long primer combination, resulting in 1199 bp. The second alignment contains only the sequences read with the forward primers COL6 or COL6a (26 sequences, 811 bp) and the third alignment the ones read with the reverse primer COH1b (58 sequences, 611 bp). Since all three parsimony networks show similar distribution patterns, only the one with most individuals, i.e. the one sequenced with the primer COH1b, is shown (Figure 2). The other two can be found in the Supplementary material (Appendices 1–2). For the graphic presentation of the networks, the two localities of Ibiza (Cala Llenya and Cala Nova) were summarized as one population, since they are geographically very close to each other and there were no important differences in their sequences. The same approach was used for the different sites in Greece.

Fig. 2. Maximum parsimony network of Cox1 mtDNA (COH1b alignment) of Palaemon serratus, constructed with TCS version 1.21 and a connection limit of 65 steps, based on an alignment of 58 sequences (611 base pairs). The numbers within the circles correspond to the number of individuals represented by that haplotype (circles without a number represent single individuals). Black spots represent missing haplotypes.
The maximum parsimony network based on the alignment of the COH1b sequences (Figure 2) shows a clearcut dissociation between the Mediterranean populations (including the Guadalquivir Estuary) and the more western Atlantic populations. Sequences from the Guadalquivir Estuary are recovered as four haplotypes, of which haplotype A with 10 individuals holds a central position. Interestingly, the sequences of Guadalquivir Estuary do not share haplotypes with the Mediterranean populations, despite being closely related. Consequently, the Mediterranean populations have their own central haplotype B found in 12 individuals. The Atlantic haplotype closest to the ones of the Guadalquivir Estuary is separated by 62 mutation steps (equivalent to 10.14% of the COH1b alignment length) and belongs to the population of Anglesey. In contrast to the Mediterranean populations and the one of the Guadalquivir Estuary, all the Atlantic populations are mixed and scattered with no recognizable central haplotype. Haplotype and nucleotide diversities were only calculated for the six populations with more than two individuals listed in Table 3. These values can only be compared conditionally, because the number of sequences of each population differs. The population of the Guadalquivir Estuary shows the lowest haplotype diversity with 0.49451, followed by the individuals of Greece, Ibiza and then Anglesey, while Saint-Jouin-Bruneval and Artabro Gulf with 1.0 each have the highest values. The highest nucleotide diversity is also found in the population of Saint-Jouin-Bruneval, followed by Anglesey and the population of Artabro Gulf and then Ibiza and Greece. The lowest nucleotide diversity was found in the population of the Guadalquivir Estuary with 0.0009.
Table 3. Estimates of haplotype and nucleotide diversities of Palaemon serratus, based on the COH1b alignment of the Cox1 gene with 611 base pairs. Calculated with DnaSP version 5.10.01. Only populations with more than two individuals were included. Permutation test with 10,000 replicates.

For the AMOVA, we also used the COH1b-alignment of the Cox1 gene, since it contains most sequences. Eight individuals of the different sites in Greece, 12 individuals of the two populations of Ibiza and 14 individuals of the population of Guadalquivir Estuary were compared. Between Ibiza and Greece, a ΦST value of 0.04562 (P = 0.1) indicates the high level of panmixia between these populations of the Mediterranean Sea. In contrast, the ΦST values of the pairwise differences between Ibiza and the Guadalquivir Estuary (0.62084) and Greece and the Guadalquivir Estuary (0.68065) are highly significant (P < 0.01). They demonstrate that there is restricted gene flow across the Strait of Gibraltar, despite the genetic similarity between the Guadalquivir and Mediterranean populations.
To confirm the results of the Cox1 gene, a small network was constructed with 10 sequences of the 16S mtDNA gene based on an alignment of 499 basepairs (Figure 3). It consists of four different haplotypes. One of these haplotypes represents three Mediterranean individuals, while the fourth Mediterranean individual only differs in one position. Two individuals of the Gulf of Cádiz population directly west of the Strait of Gibraltar (Cádiz and Guadalquivir Estuary) are also separated from the larger Mediterranean haplotype by one mutational step. In contrast, the fourth haplotype is separated from all others by at least 10 mutational steps (equivalent to 2.0%) and is found in four individuals of the other Atlantic populations (one of each location from Artabro Gulf, Calais, Cork and Anglesey). Therefore, the 16S network confirms the structure of the Cox1 network.

Fig. 3. Maximum parsimony network of 16S mtDNA of Palaemon serratus, constructed with TCS version 1.21 and a connection limit of 94%, based on an alignment of 10 sequences (499 base pairs). The number within the circles correspond to the number of individuals represented by that haplotype (circles without a number represent single individuals). Black spots represent missing haplotypes.
The mismatch analyses for both the COH1b sequences of Cox1 mtDNA and 16S rRNA show a bimodal distribution, i.e. two separate peaks, in the frequency of pairwise differences among haplotypes (Figure 4). This reflects the fact that the haplotypes are either very close to each other (within Atlantic and Mediterranean) or more distantly related (between Atlantic and Mediterranean). In Cox1 mtDNA, haplotypes are either separated by not more than nine or between 62–71 mutational steps, whereas in 16S mtDNA there are either up to two or between 10 and 11 mutational steps. In both cases, the observed frequency (dashed line) did not correspond to the expected values for a constant population size (solid line).

Fig. 4. Mismatch distribution of pairwise differences in two mitochondrial genes of Palaemon serratus. Calculated with DnaSP version 5.10.01. The dashed lines show the observed, the solid lines the expected frequencies within a constant population size. (A) Cox1 mtDNA (COH1b alignment with 58 sequences, 611 base pairs). (B) 16S mtDNA (10 sequences, 499 base pairs).
An independent nuclear marker was applied as third evidence and to test for possible ongoing speciation: The nuclear gene Enolase was sequenced from 26 individuals, resulting in an alignment with a length of 409 bp. It includes only two parsimony-informative sites that consistently separate two genotypes. The corresponding maximum parsimony network can be found in Figure 5. Ten individuals originating from Croatia, Greece, Ibiza (Spain) and Corsica (France) and 10 from the population of the Guadalquivir Estuary share one genotype, whereas the second genotype is found in five individuals of the Atlantic coast of France and one specimen from Scotland. Therefore, a clean split is confirmed between the Mediterranean plus the Gulf of Cádiz sequences and the more northern Atlantic populations.

Fig. 5. Maximum parsimony network of Enolase nuDNA of Palaemon serratus, constructed with TCS version 1.21 and a connection limit of 95%, based on an alignment of 26 sequences (409 base pairs). The numbers within the circles correspond to the number of individuals represented by that genotype. Black spot represents missing genotype.
DISCUSSION
Results of three analysed genes (Cox1 mtDNA, 16S rRNA and Enolase nuDNA) show genetic differentiation among the populations of Palaemon serratus in the Mediterranean Sea, the Gulf of Cádiz and the Atlantic Ocean. The individuals of the first two areas are closely related and will henceforth be termed the Mediterranean lineage (ML) which thus includes the populations from the Guadalquivir Estuary and the Mediterranean populations. A sequence from GenBank (JQ042291) corresponding to the 16S mtDNA of an individual from Cádiz confirms that populations from the Gulf of Cádiz are more closely related to Mediterranean populations than to other Atlantic ones. The second lineage, henceforth called the Atlantic lineage (AL), contains all the other Atlantic populations included in this study. The 16S network consists of three different haplotypes in the ML and only one haplotype in the AL, while all three Cox1 networks have a smaller number of haplotypes in the ML than in the AL. This can be due to the fact that only one individual per population was used for the 16S analysis, so that most likely not all occurring haplotypes are represented in the network. In the Enolase alignment, only two different genotypes can be found, corresponding to ML and AL and separating these in the same fashion as the mitochondrial genes.
The Cox1 haplotypes of the populations of the Mediterranean are linked tightly, and while the Greek individuals show a moderate haplotype diversity (h d = 0.60714), the individuals of the populations of Ibiza are quite heterogeneous (h d = 0.80303). This suggests that the Mediterranean stocks represent a healthy and stable system, with relatively high gene flow among the sampled populations. Nevertheless, this should be tested with more individuals and more populations, since only two Mediterranean populations were used for the analysis of haplotype and nucleotide diversities in this study.
The specimens of the Guadalquivir Estuary form a very homogeneous clade, with only a few haplotypes (h d = 0.49451) being very close to each other (n d = 0.0009). This can be best explained by a founder effect, i.e. a new population founded by only a few individuals and therefore with low genetic variety. The proximity of some Cox1 haplotypes from Ibiza to the Guadalquivir haplotype A suggests a relatively recent colonization from the western Mediterranean. Another possible explanation would be reduced gene flow to and from the Guadalquivir population, because of its geographically marginal position, compared with a population that is located in the centre of the range of a genetic lineage. In any case, current gene flow of the population from the Guadalquivir Estuary with the Mediterranean populations seems limited.
The fact that the population of the Guadalquivir Estuary nevertheless belongs to the ML raises the question why it groups together with Mediterranean populations instead of Atlantic ones. Most other studies of species with a genetic separation of the Atlantic and the Mediterranean populations reveal the opposite scenario: They show an extension of the Atlantic genotypes into the Alborán Sea which is the westernmost part of the Mediterranean Sea. This is for example the case in the closely related prawn Palaemon elegans (see Reuschel et al., Reference Reuschel, Cuesta and Schubart2010) as well as in many other marine species, e.g. the scallops Pecten jacobeus and P. maximus (see Ríos et al., Reference Ríos, Sanz, Saavedra and Peña2002) or the euphausiid Meganyctiphanes norvegica (see Zane et al., Reference Zane, Ostellari, Maccatrozzo, Bargelloni, Cuzin-Roudy, Buchholz and Patarnello2000). This extension of Atlantic genotypes into the Mediterranean Sea has been explained by the intrusion of Atlantic waters into the Alborán Sea, where they circulate as two gyres, so that the Almería-Oran Front often represents the real hydrographic boundary of Atlantic and Mediterranean surface waters (Tintoré et al., Reference Tintoré, La Violette, Blade and Cruzado1988), rather than the Strait of Gibraltar itself. However, in a recent study on several crustacean species, the role of the Strait of Gibraltar as a gene flow barrier compared with the one of the Almería-Oran Front has been emphasized (García-Merchán et al., Reference García-Merchán, Robainas-Barcia, Abelló, Macpherson, Palero, García-Rodríguez, Gil de Sola and Pascual2012).
Palaemon serratus is not the only species in which populations of the Guadalquivir Estuary differ genetically from other Atlantic populations, as this was also reported in the mysid Neomysis integer (see Remerie et al., Reference Remerie, Vierstraete, Weekers, Vanfleteren and Vanreusel2009). It remains unknown, when Mediterranean P. serratus may have migrated from the Mediterranean Sea through the Strait of Gibraltar and settled in the Gulf of Cádiz. Furthermore, we do not know if they replaced ‘typical’ Atlantic P. serratus which theoretically should be better adapted to the local conditions, like temperature and tides, in the Gulf of Cádiz. Alternatively, it is conceivable that the Gulf of Cádiz population may have diverged from Atlantic stocks independently and gave rise to the Mediterranean populations with the flooding of the Mediterranean Basin. Hence, more detailed studies about the dispersal and ecology of P. serratus, especially in the area west and east of the Strait of Gibraltar, are necessary to reconstruct how the extension of Mediterranean haplotypes into the Atlantic Ocean took place. A human introduction can be excluded, because the haplotypes from the Guadalquivir Estuary of both mitochondrial genes have so far not been sampled in the Mediterranean Sea and thus appear to be endemic. It remains to be solved, where the contact zone with the other Atlantic populations is, and if there is genetic mixing. One possible explanation for endemic genotypes could be the fact that the population of the Guadalquivir Estuary belongs to the most important nursery area in the Gulf of Cádiz, favouring the reproduction and/or the settlement of local individuals (González-Ortegón et al., Reference González-Ortegón, Baldó, Arias, Cuesta, Fernández-Delgado, Vilas and Drake2015) and probably a more estuarine genotype. Further investigations of different populations along the coastline of Andalusia to Portugal are necessary to define the exact border between AL and the haplotypes of the Gulf of Cádiz, and to determine what induces the biogeographic break within the Atlantic Ocean.
The populations of AL are genetically very diverse with 18 different haplotypes in the COH1b network (Figure 2), and all tested populations show very high haplotype and nucleotide diversities. According to the coalescent theory of Kingman (Reference Kingman1982), high genetic diversity leads to the conclusion that the corresponding species is presumably of a great age (the higher the diversification the longer the required time span to develop this diversity). This provides evidence that the Atlantic stock of Palaemon serratus represents a very stable and healthy system, which has probably existed for a long time. The maximum parsimony networks as well as the haplotype diversities indicate high gene flow among the Atlantic populations, with the exception of the ones from the Gulf of Cádiz. It would be useful to confirm these assumptions with more individuals per population and more populations to cover the whole distribution area of P. serratus and to quantify gene flow statistically, as no populations east of Belgium and south of Spain are represented in this study.
Our results suggest the existence of a potential cryptic species or at least ongoing speciation within P. serratus, i.e. a morphological indistinct lineage separated by genetic differences at species level (Belfiore et al., Reference Belfiore, Hoffmann, Baker and Dewoody2003). The separation of the ML and the AL amounts to 2.0% in the 16S gene and 10.14% in the Cox1 gene and is thus comparable to recognized species from other crustacean genera with an Atlantic–Mediterranean separation, such as in Carcinus (see Geller et al., Reference Geller, Walton, Grosholz and Ruiz1997; Roman & Palumbi, Reference Roman and Palumbi2004; Ragionieri & Schubart, Reference Ragionieri and Schubart2013) or Brachynotus (see Schubart et al., Reference Schubart, Cuesta and Rodríguez2001). Furthermore, the independently evolving nuclear gene Enolase confirms the same differentiation pattern. So far, there are no morphological characters known that differ in Atlantic and Mediterranean specimens of P. serratus, but this needs to be verified in a separate study. It would also be important to know if hybridization of Atlantic and Mediterranean individuals is possible. Considering the fact that in this study only a few populations were represented by more than 10 individuals, statistical possibilities are limited and make further studies with larger sample sizes necessary. No type locality for Palaemon serratus was indicated by Pennant (Reference Pennant1777), but even though a few Mediterranean individuals were included in his first study characterizing the species, most of the individuals studied were from the Atlantic Ocean, i.e. from the English Channel, Belgium and the Netherlands (De Man, Reference De Man1915). For that reason, and because Pennant published his results in British Zoology, it appears logical that the species name Palaemon serratus would remain with the Atlantic individuals, if two separate species were to be recognized in the future.
Overall, this study reveals some interesting phylogeographic aspects, although the results have limited conclusive significance, due to the restricted number of populations and the fact that some analysed populations consisted of very few individuals and therefore could not be used for statistical analyses. Hence, more research is needed regarding the population genetics and taxonomic status of P. serratus, including morphological and morphometric aspects, to understand the remarkable case of an Atlantic population with haplotypes that are more closely related to Mediterranean ones than to the other Atlantic ones, and to decide whether a cryptic species is involved.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/S0025315417000492
ACKNOWLEDGEMENTS
We would like to thank Cédric d'Udekem d'Acoz, Daniel Hamm, Carsten H.G. Müller, Piero Calosi, Henrik M. Schubart, Declan Quigley, Declan McGabhann and Hans De Blauwe for providing us with specimens of Palaemon serratus and for help during collecting. Also thanks to Sammy De Grave for museum specimens and specific literature and to Stefan Buchhauser for help with preparing the figures. One part of the work was carried out using the resources of the Departamento de Biología Celular y Molecular of the Universidade da Coruña, the other one in the Department of Zoology and Evolutionary Biology of the University of Regensburg (thanks to Prof. Jürgen Heinze).
FINANCIAL SUPPORT
This study was financed by the research funds of the Universidade da Coruña AGL2011-23689 and of the Universität Regensburg. EGO was funded by the Juan de la Cierva Research Program (MINECO) and Campus de Excelencia Internacional del Mar (CEIMAR).