




Introduction
During the Iron Age the lives of thousands of people came to their end in the wetlands of North-west Europe (van der Sanden Reference van der Sanden1996). Bodies deposited in fens were reduced to skeletons, whereas bodies placed in acidic peat bogs were transformed into naturally mummified ‘bog bodies’, with preserved skin, hair, nails and sometimes even intestines. Some of these individuals may have fallen accidentally into the bogs, while others may have been intentionally put there, the subjects of punishment, murder or even ritual sacrifice (e.g. van der Sanden Reference van der Sanden1996). During the Danish Early Iron Age (500 BC–AD 200), bogs were used not only for practical purposes, such as cutting peat, but also for the ritual deposition of objects (e.g. Mortensen et al. Reference Mortensen, Christensen, Johannesen, Stidsing, Fiedel and Olsen2020). Understood in this context, some of the bog bodies probably represent human sacrifices.
The research potential of bog bodies
Well-preserved bog bodies represent a rare type of evidence that can offer unique insights into aspects of daily life in prehistory, such as the appearance of individuals, their dress, health, diet, final meal and manner of death. Furthermore, investigations of bog bodies add to the general understanding of prehistoric societies, especially when patterns emerge through the study of several individuals. Analyses of gut contents, for example, can offer insights into the preparation and composition of meals and on general hygiene and health.
Last meals
Food is often connected to rituals (e.g. Mintz & du Bois Reference Mintz and du Bois2003). Consequently, the last meals consumed by individuals preserved in bogs have been discussed in relation to human sacrifice (e.g. Glob Reference Glob1956: 109; Scaife Reference Scaife, Stead, Burke and Brothwell1986). A meal does not have to include special ingredients to form part of a ritual: the ritual element may be connected, for example, to how it was eaten. As the latter is impossible to demonstrate archaeologically, the investigation of a ritual aspect has focused on the search for ‘special’ ingredients, or a composition that deviates from the expected content of an ‘ordinary’ Iron Age meal. But such comparison is hampered by the fact that actual meals from this period can only be studied from bog bodies. Elsewhere, carbonised grains, animal bones and isotope analyses merely indicate that the Iron Age diet was primarily based on grain and animal products from pigs, cows and sheep.
The last meals of 12 Northern European Iron Age bog bodies have previously been analysed (e.g. van der Sanden Reference van der Sanden1996; Behre Reference Behre2008). These meals are generally interpreted as porridge or bread, occasionally accompanied by meat or berries. There is no sound evidence for ingredients with special properties having been added. Mistletoe, which had medicinal properties and had religious significance, was found in Lindow Man's gut, but only four pollen grains were recovered—too few to conclude that his last meal or drink had been particularly ‘special’ (Scaife Reference Scaife, Stead, Burke and Brothwell1986). The suggestion that ergot was given as a hallucinogen before execution (Hillman Reference Hillman, Stead, Burke and Brothwell1986: 103) has been disproved by the reinvestigation of Grauballe Man, as there was insufficient ergot in his gut to cause such a reaction (Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007). The considerable quantity of seeds from uncultivated plants in the gut contents of several bog bodies is, however, notable (Brandt Reference Brandt1951; Holden Reference Holden1999; Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007; Behre Reference Behre2008). Although seeds from wild plants are known from several Iron Age sites (Helbæk Reference Helbæk1954; Behre Reference Behre2008), grains from cultivated crops always dominate at sites where grain stores have been found (Henriksen & Robinson Reference Henriksen and Robinson1996). The plant component of an ordinary Iron Age meal would therefore be expected to consist primarily of grain. The abundance of seeds from wild plants in bog bodies may suggest that these seeds were used on special occasions.
When evaluating published studies of gut contents, the preservation conditions and the degree of pre- and post-excavation contamination is a challenge because they all vary greatly between bog bodies. Furthermore, most studies were undertaken at a time when plant macrofossil analysis was less well developed (van der Sanden Reference van der Sanden1996). Comparisons of gut contents are therefore challenging and can only be made cautiously, based on the presence or absence of ingredients.
A recent analysis of Grauballe Man (Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007) highlights the potential of re-analysing gut contents, as not only were the various ingredients of his last meal quantified, but the analysis also revealed that the meal contained a surprisingly large quantity of threshing waste. This analysis has prompted a renewed interest in the gut contents of Tollund Man.
Tollund Man
Tollund Man's astonishingly well-preserved head makes him one of the most famous bog bodies (Figure 1c). He was found during peat cutting in 1950 in Bjældskovdal, some 10km west of Silkeborg, Denmark (Figure 1a; Fischer Reference Fischer2012). He was 30–40 years old when he died c. 405–380 BC, at the beginning of the Danish Early Iron Age (Nielsen et al. Reference Nielsen, Philippsen, Kanstrup and Olsen2018). He had been hanged and subsequently placed in a sleeping position in a peat-cutting pit. Widely accepted as the victim of a sacrifice (Fischer Reference Fischer2012), Tollund Man's extraordinary treatment is underlined when considered in relation to the contemporaneous burial practices of cremation and burial on dry land (Jensen Reference Jensen2003: 56–63).

Figure 1. a) Map showing the location of Tollund Man's discovery in Bjældskovdal (figure by N.H. Nielsen, Museum Silkeborg; contains data from the Danish Geodata Agency); b) Tollund Man's large intestine (photograph courtesy of the Danish National Museum); c) the well-preserved head of Tollund Man (photograph by A. Mikkelsen); d) the original honey glass jar in which the contents of the large intestine were kept until 2018; e) the remaining 140ml of the large intestine content (d–e photographs by N.H. Nielsen, Museum Silkeborg).
The stomach and intestinal tract of Tollund Man were removed during forensic examinations in 1950 (Figure 1b). Subsequently, the contents of the different parts of his gut were separately extracted (Helbæk Reference Helbæk1951). The stomach contained 0.5ml of material, the small intestine 10ml, and the large intestine 260ml, together comprising primarily the remains of a meal eaten 12–24 hours before death. Helbæk (Reference Helbæk1951) analysed all the stomach and small intestine samples for plant macrofossils, as well as 50ml of the large intestine material.
Helbæk (Reference Helbæk1951) identified cereal grains, as well as seeds from wild plants, encountering both whole seeds and large fragments. Other than three glume fragments from oats (Avena sp.), all grain residues were identified as six-rowed barley (Hordeum vulgare), both hulled and naked. Helbæk concluded that Tollund Man ate a porridge consisting mainly of barley, flax (Linum usitatissimum), and seeds of the wild plants gold-of-pleasure (Camelina sativa) and pale persicaria (Persicaria lapathifolia s.l.). He also found the remains of a further 16 plant species, but did not analyse the finer material or quantify the meal content due to the degradation of the plant residues. We have re-investigated the gut contents of Tollund Man with the aim of quantifying the ingredients in his last meal and gaining a better understanding of what such an Iron Age meal may have contained. Additionally, we searched for unusual ingredients that could relate to rituals, and explored whether the presence of weed seeds could represent a food shortage. A further aim was to gain insights into Iron Age cooking practices by seeking evidence of crop and seed processing and meal preparation. Moreover, we have sought to investigate Iron Age health and sanitation through the analysis of intestinal parasite eggs.
New analysis of Tollund Man's gut content
Nearly all the material from Tollund Man's gut was obtained from the large intestine. The food residues were thus expected to be better preserved than they would be in coprolites (Shillito et al. Reference Shillito, Blong, Green and van Asperen2020), but more degraded than those found in the small intestine, such as in Grauballe Man (Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007). To gain the most information from the gut content, we decided to analyse the plant macrofossils, pollen, non-pollen palynomorphs (NPPs), lipids/steroid markers and proteins. Pollen and NPP analysis can reveal microscopic evidence of consumed food and drink. While the gut contents of a few bog bodies have been analysed for pollen (e.g. Scaife Reference Scaife, Stead, Burke and Brothwell1986, Reference Scaife, Turner and Scaife1995), our work constitutes the first application of a full NPP analysis of a bog body's gut content. In addition, the analysis of parasite eggs can illuminate individual health conditions; they have been observed in several cases (van der Sanden Reference van der Sanden1996), including Tollund Man, where whipworm (Trichuris) eggs were recorded (Helbæk Reference Helbæk1959). Analyses of steroids and proteins can reveal chemical traces of consumed food and drink (e.g. from animal products) (Shillito et al. Reference Shillito, Blong, Green and van Asperen2020), which so far have not been documented in Tollund Man. To our knowledge, this is the first protein and steroidal analysis of bog body gut contents.
Materials and methods
Sampling
The 140ml of precipitated material from the large intestine (Figure 1e) comprised just over half the original sample (260ml). The 50ml that Helbæk analysed was never returned to the main sample. In 2015, 15ml of precipitated sample material was taken by Martin Jensen Søe for (unsuccessful) DNA analysis and is now kept separately. Furthermore, we extracted a 0.5ml sample for protein analysis in 2016. Although approximately 55ml of the original sample is still unaccounted for, we consider the material that is still available to be representative; this situation underlines the importance of using as little as possible of an irreplaceable material in new investigations.
The content of the large intestine, weighing 283.2g (including ethanol and formalin), was sub-sampled by dividing it in a riffle-box. One half of the last subdivision (14.9g, including the liquid) was used for plant macrofossil analysis; from the other half, 10 × 1ml were used for pollen and NPP analysis and 2ml for GC-MS analysis. The 14.9g corresponds to 5.3 per cent of the existing sample. Although small, it is considered sufficient, as statistically reliable results regarding the plant macrofossils were obtained from only 0.6 per cent of the material from Grauballe Man's gut (Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007).
Plant macrofossils
In the time allocated to our study, a complete analysis of the entire macrofossil assemblage, which includes thousands of unidentifiable fragments (Figure 2), was unfeasible. As the main purpose of this analysis was to quantify the meal's ingredients, we focused on the eight most frequently encountered taxa, although the presence of other identifiable species was also recorded. Seed degradation varied between species, and the method of calculating the number of seeds was adjusted appropriately. As bran and glume fragments of barley were too degraded to estimate the original number of grains, the better preserved hilum—an elongated scar from the attachment of the grain to the placenta—was used. Hila appeared both as 1–3mm-long fragments and as intact hila on the complete pericarps identified by Helbæk (Figure 3a). All hilum fragments in the analysed sub-sample were measured under the microscope. As the hilum on an intact barley testa in Helbæk's (Reference Helbæk1951) material measured 7mm, the total minimum number of barley grains in our sub-sample could be estimated by dividing the total length of all hilum fragments by 7mm.

Figure 2. Photomicrograph of Tollund Man's gut content (photograph by P.S. Henriksen, the Danish National Museum).

Figure 3. Photographs of: a) barley (Hordeum vulgare) pericarp with a dark hilum line; b) sand; c) food crust; d) pointed ends of flax (Linum usitatissimum) seeds from Tollund Man's gut content. Each square in a, b and d measures 1 × 1mm (photographs by P.S. Henriksen, the Danish National Museum).
Flax seeds were also very fragmented and separated into the outer and inner layers of the seed coat. A minimum number of flax seeds could, however, be estimated by counting their pointed ends—the most resistant part of the seed (Figure 3d). Pale persicaria was quantified by sorting and counting whole seeds and fragments larger than 1mm. All smaller fragments were also counted, and a representative portion was selected, counted and weighed in order to calculate the total weight of the fragments. The weight of one seed was calculated on the average weight of all whole seeds in the sample to obtain an estimate of the original content of pale persicaria. The content of black-bindweed (Fallopia convolvulus) was estimated in the same way.
Quantification of gold-of-pleasure, corn spurrey (Spergula arvensis), fat hen (Chenopodium album), wild turnip (Brassica rapa), hemp-nettle (Galeopsis sp.) and field pansy (Viola arvensis) was achieved by identifying all the whole seeds and fragments and estimating the equivalent number of whole seeds of each species. Sand grains were also counted and weighed.
Pollen and NPPs
Analyses of pollen and NPPs were undertaken on 10 sub-samples to minimise any effects of unequal distribution of palynomorphs in the main sample. In order to preserve as many NPPs as possible, a mild treatment (Enevold Reference Enevold2018) with cold HCl and KOH (10 per cent solutions) was applied, and the residues were mounted in silicone oil and sealed. A minimum of 100 terrestrial pollen grains were counted per sub-sample, in addition to NPPs and charred fragments. A few slides of non-prepared gut content were screened to check that the samples were not biased by the preparation, but no obvious differences were observed. All terrestrial pollen was included in the pollen sum. Beug (Reference Beug2004) was consulted for the pollen identification and van Geel (Reference van Geel, Smol, Birks and Last2001) for the NPPs (among others).
Steroidal biomarkers
The chemical compounds cholesterol, sitosterol, coprostanol and 5β-stigmastanol, epi-coprostanol, 5α-cholestanol, 5α-stigmastanol and epi-5β-stigmastanol were analysed by gas chromatography-mass spectrometry (GC-MS) on samples of gut content and skin from the buttock region of Tollund Man (for details, see OSM1 in the online supplementary material). Cholesterol is a zoolipid (originating from animals), while sitosterol is a phytolipid (originating from plants). Coprostanol and 5β-stigmastanol are gut-conversion products of cholesterol and sitosterol, respectively. Hence, the ratio between coprostanol and 5β-stigmastanol GC-MS peak areas represent the ratio of digested animal fat to digested plant fat. Different ratios found in herbivore, omnivore and carnivore coprolites have been utilised to describe past diets (e.g. Sistiaga et al. Reference Sistiaga, Berna, Laursen and Goldberg2014a). Here, the method was applied to estimate the relative zoolipid:phytolipid ratio in Tollund Man's last meal. Stanol ratios yield information about lipids that have been digested and converted—in the case of Tollund Man, before or shortly after death. Analysis of skin from the buttock region made it possible to compare body fat to ingested fat. Epi-coprostanol, 5α-cholestanol, 5α-stigmastanol and epi-5β-stigmastanol are products from the diagenesis of coprostanol, cholesterol, sitosterol and 5β-stigmastanol, respectively (Sistiaga et al. Reference Sistiaga, Mallol, Galván and Summons2014b). Coprostanol, cholesterol, sitosterol and 5β-stigmastanol were identified in the chromatograms using pure reference samples; the other identifications were based on analysis of mass spectra and the use of the NIST database, as described in OSM1.
Proteins
Proteins in a sub-sample of the large intestine were identified by mass spectrometry (see OSM2). The analysis provides information on peptide sequences, which can be searched and matched to specific proteins and, if the sequence is unique, can also identify the species from which the protein originated. The data were searched against all animal and plant protein sequences in the uniprot database (www.uniprot.org). All identified peptides were inspected to see whether they could be assigned to a human protein from Tollund Man himself.
Results
Plant macrofossils
Table S1 details the results from the plant macrofossil analysis. The number of seeds was generally determined by counting seeds and fragments, but the barley grains were calculated from measuring over 1000 hilum fragments. The total length of hilum fragments measured 2214mm, corresponding to 316 grains of barley in the sub-sample. Helbæk's (Reference Helbæk1951) analysis showed that most of the epidermal remains came from barley, while some may have come from other grass species, as these can be very similar to barley.
Barley was the dominant taxon, accounting for 85 per cent of the sample by weight, followed by pale persicaria (9 per cent) and flax (5 per cent). All other species accounted for less than 1 per cent by weight. We therefore estimate that the original 260ml sample from the large intestine contained approximately 11 150 barley grains (335g), 9600 pale persicaria seeds (29g), 3250 flax seeds (16g) and negligible amounts of other seeds. Although the sample originally included more than 1100 seeds of corn spurrey and 670 seeds of gold-of-pleasure, we estimate that their small size meant that they contributed only 0.6g and 0.8g, respectively. We also recorded less common taxa (not quantified), identifying three new wetland plant species: marsh willowherb (Epilobium palustre), compact/soft rush (Juncus conglomeratus/effusus) and marsh violet (Viola palustris).
Small fragments of charcoal, food crust and numerous grains of sand were also identified in the sub-sample (Figure 3). Forty-two coarse sand particles (>0.5mm) weighing a total of 13.7mg, and 575 smaller sand grains weighing 34.1mg, were recorded. The total amount of sand in the original sample is estimated to have been 1.6g.
Pollen and NPPs
Figure 4 gives an overview of the results from the pollen and NPP analysis (for complete tables (Tables S3–4), see OSM4). In total, 1266 pollen grains were counted. Clusters of barley pollen (Hordeum-type) were frequently seen (Figure 5a), and were counted as one pollen grain, regardless of how many grains the cluster contained. Barley dominated the pollen assemblage (>90 per cent), followed by grasses (Poaceae; 4.6 per cent) and knotweed (Polygonum aviculare-type; 1 per cent). The remaining pollen taxa, each comprising less than 1 per cent of the total, were dominated by open dryland taxa. Many taxa probably represent the same species identified in the macrofossil analysis (e.g. Persicaria, Spergularia and Brassicaceae).

Figure 4. Proportions of pollen (top) and non-pollen palynomorphs (bottom) in Tollund Man's gut content (figure by R. Enevold, Moesgaard Museum).

Figure 5. Photomicrographs of frequent palynomorphs in Tollund Man's gut contents: a) cluster of barley (Hordeum vulgare) pollen; b) epidermis cells from flax (Linum usitatissimum); c) epidermis cells from barley; d) egg from whipworm (Trichuris trichiura); e) egg from mawworm (Ascaris sp.); f) egg from tapeworm (Taenia sp.) (photographs by R. Enevold, Moesgaard Museum).
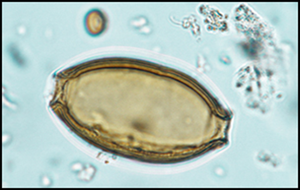
A total of 801 NPPs were recorded, covering 34 types (see Table S4). Microscopic plant fragments comprise 63 per cent of the total NPP count. Approximately half of these represent two of the main components of the meal: barley and flax (Figure 5b–c). The other half includes plant fragments that could not be further identified, and were therefore only assigned a type number. Parasite eggs, including intestinal whipworm (Trichuris sp.; 22 per cent of the total NPP count), tapeworm (Taenia sp.; 11 per cent) and mawworm (Ascaris sp.; 1 per cent), were recorded (Figure 5d–f). The size range of 178 Trichuris eggs (Figure 6) corresponds to that of a species that prefers humans or pigs as hosts (Trichuris trichiura) (Brinkkemper & van Haaster Reference Brinkkemper and van Haaster2012). A small proportion of the remaining non-pollen palynomorphs originated from freshwater organisms, such as algae (1 per cent).

Figure 6. Left) photomicrograph of Trichuris trichiura egg and measurement criteria (m1 and m2); right) size distribution of measured Trichuris eggs (blue dots) and size range for each Trichuris type (coloured squares; Brinkkemper & van Haaster Reference Brinkkemper and van Haaster2012). The blue dots are layered each time an equal measure was encountered (figure by R. Enevold, Moesgaard Museum).
Steroids
Figure 7 shows the steroid region of chromatograms recorded on a sub-sample of the gut contents (Figure 7 top) and the buttock skin sample (Figure 7 bottom). Analysis revealed the presence of cholesterol, coprostanol, sitosterol, 5β-stigmastanol (Figure 7: a–d), epi-coprostanol, 5α-cholestanol, 5α-stigmastanol and epi-5β-stigmastanol (Figure 7: 1–4) in the gut. The coprostanol/5β-stigmastanol peak-area ratio was calculated to be 3:1. The buttock skin sample predominantly contained cholesterol and 5α-cholestanol. The peaks labelled 1–4 on Figure 7 correspond to epi- and 5α-compounds that are diagenesis products of the other four compounds labelled a–d.

Figure 7. Chromatograms (TIC) showing the steroid region in GC-MS analyses (MCps = megacounts per second) of a sub-sample of Tollund Man's large intestine contents (top) and a skin sample from the buttock region (bottom). Peaks were identified as: a) cholesterol; b) coprostanol; c) sitosterol; d) 5β-stigmastanol; 1) epi-coprostanol; 2) 5α-cholestanol; 3) 5α-stigmastanol; 4) epi-5β-stigmastanol (figure by M.N. Mortensen, the Danish National Museum).
Proteins
Protein analysis identified a large quantity of collagen peptides (see OSM6). While these peptides could originate from sheep, pig or cow, all but one peptide can also be assigned to a homolog human protein. Five peptides unique to bony fish, however, were also identified, indicating the presence of fish in the sample. In addition, peptides originating from tapeworm (Taeniidae), mawworm (Ascaris), an unspecified nematode (Strongylida), barley and the cabbage family (Brassicaceae) were identified. The peptides were all significant at a threshold of p<0.01 and with an expected value of each peptide below 0.005.
Discussion
Plant ingredients in Tollund Man's last meal
The results of the present plant macrofossil analysis generally concur with Helbæk's (Reference Helbæk1951) analyses, with only a few species not recorded in both investigations (see OSM3). It is, however, evident that Helbæk's (Reference Helbæk1951) analysis of primarily whole and half seeds led to biased estimations of the quantities; some taxa (e.g. corn spurrey) were underestimated, while others (e.g. gold-of-pleasure) were overestimated.
Figure 8 shows the main ingredients of Tollund Man's last meal in proportions reflected in the plant macrofossils. The dominance of barley, pale persicaria and flax is clear. This is also partly shown by the pollen and NPP results, with barley comprising 90 per cent of the total pollen sum, and barley and flax being the most common microscopic plant fragments among the NPPs. It is nevertheless notable that pale persicaria is less well represented in the pollen assemblage than in the macrofossil assemblage. This seems surprising, as persicaria pollen is both robust and easily recognisable; possibly the plant was deflowered when harvested.

Figure 8. Reconstruction of the ingredients in Tollund Man's last meal, shown in quantities relative to the extant 140ml of intestinal contents: A) barley (Hordeum vulgare); B) pale persicaria (Persicaria lapathifolia s.l.); C) barley rachis segments; D) flax (Linum usitatissimum); E) black-bindweed (Fallopia convolvulus); F) fat hen (Chenopodium album); G) sand; H) hemp-nettles (Galeopsis sp.); I) gold-of-pleasure (Camelina sativa); J) corn spurrey (Spergula arvensis); K) field pansy (Viola arvensis) (photograph by P.S. Henriksen, the Danish National Museum).
The plant-based component in the gut contents is also reflected by the presence of sitosterol (a phytosterol) and protein from plants (from barley and the cabbage family). Additionally, the presence of 5β-stigmastanol—the product of the action of gut bacteria on sitosterol (Bull et al. Reference Bull, Lockheart, Elhmmali, Roberts and Evershed2002; Sistiaga Reference Sistiaga, Mallol, Galván and Summons2014b)—indicates that the ingested phytosterol was subjected to gut bacteria for some time before Tollund Man's death.
The last meal of Tollund Man contained different components that were probably mixed together. Barley and flax are not cultivated together, as their seasons of growth and maturation are very different (Madsen-Mygdal Reference Madsen-Mygdal1937: 177ff & 633ff). The seeds of the weed pale persicaria were presumably harvested along with the barley crop. Finds of unthreshed grain dated to the last century BC in Alrum in western Jutland and Overbygård in northern Jutland (Henriksen & Robinson Reference Henriksen and Robinson1994, Reference Henriksen and Robinson1996) show that harvested but unthreshed crops could contain more than ten per cent of wild plant seeds by number. As the number of wild plant seeds in Tollund Man's gut is more than equal to the number of barley grains, it is unlikely that threshed but uncleaned grain was used in the meal. Rather, the barley and the seeds must have been separated before they were added to the meal as individual ingredients.
Archaeobotanical evidence from Overbygård shows that threshing and subsequent cleaning separates nearly all wild plant seeds from the grain (Henriksen & Robinson Reference Henriksen and Robinson1996). The final step in grain cleaning was sifting, which removes small weed seeds, sand and other impurities picked up from the threshing floor (Viklund Reference Viklund1998). The presence of numerous pale persicaria seeds, along with other small weed seeds, sand grains and charcoal fragments in Tollund Man's gut suggests that it was this sifted fraction that was added to the barley grains and flax seeds when cooking the meal. Such use of threshing waste as a cooking ingredient was also inferred from the analysis of Grauballe Man (Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007).
Evidence of animal products
Helbæk (Reference Helbæk1951: 322) argued that meat had not been part of Tollund Man's last meal, as there was no visible evidence of bones, sinews or muscle tissue. Animal meat and fat, along with egg or dairy, however, would leave no physical traces. As both cholesterol and coprostanol were detected in the gut content, cholesterol conversion must have taken place in the gut before death. The cholesterol could, however, have come from both ingested cholesterol (e.g. from animal fat, egg or dairy) and Tollund Man's own tissues (e.g. intestinal epithelium); coprostanol would form from both of these sources. The coprostanol:5β-stigmastanol ratio is lower than one in faeces from herbivores, which only ingest phytosterols, and higher than one for omnivores, which ingest both phytosterols and zoosterols (Bull et al. Reference Bull, Lockheart, Elhmmali, Roberts and Evershed2002; Sistiaga et al. Reference Sistiaga, Berna, Laursen and Goldberg2014a, Reference Sistiaga, Mallol, Galván and Summons2014b). The coprostanol:5β-stigmastanol peak-area ratio of 3:1 for Tollund Man represents a zoosterol level higher than in herbivores, indicating that Tollund Man's last meal included both cholesterol-containing and plant-based foods.
The skin sample from the buttock region was dominated by cholesterol from body fat and 5α-cholestanol, which both form via cholesterol diagenesis (Bull et al. Reference Bull, Lockheart, Elhmmali, Roberts and Evershed2002; Sistiaga et al. Reference Sistiaga, Berna, Laursen and Goldberg2014a, Reference Sistiaga, Mallol, Galván and Summons2014b). The gut-conversion product coprostanol was detected in only minor quantities (although cholesterol was present in the buttock sample), confirming that conversion takes place in the gut rather than in the skin. The absence of significant quantities of sitosterol, 5β-stigmastanol, 5α-stigmastanol and epi-5β-stigmastanol in the buttock sample further supports the interpretation of the 5β-stigmastanol and coprostanol in the gut being the result of ingested or converted food rather than of contamination.
Five peptides uniquely assigned to bony fish provide clear evidence of the consumption of fish as part of the last meal. A fatty fish, such as eel, may account for the lipids found in the gut content. The lack of identified fish bones does not contradict this interpretation, as these often do not survive digestion (Shillito et al. Reference Shillito, Blong, Green and van Asperen2020). Although previous isotope analyses of Tollund Man show that fish did not constitute a significant part of his diet (Nielsen et al. Reference Nielsen, Philippsen, Kanstrup and Olsen2018), the proximity of Bjældskovdal to a lake and watercourses suggests that fish was probably consumed, at least occasionally. Lipids from other animals may also have been included in the meal, but the identified peptides that could possibly come from sheep, pig or cow could also be assigned to human proteins. For now, the ingestion of fatty fish appears to be the most likely explanation for the lipids.
Preparation, consumption and character of the last meal
Our investigation provides insights into how the last meal was prepared. The fragmentation of the cereals and other seeds suggests that these had been ground before being cooked. Experiments with grinding vs chewing of persicaria seeds in connection with the Grauballe project showed that the seed fragmentation in the stomach and gut contents of both Tollund and Grauballe Man can only be achieved by thoroughly grinding the seeds (unpublished data), indicating that not only grains but also sometimes wild plant seeds were ground before consumption.
The presence of a charred food crust in Tollund Man's gut content suggests that the meal was a porridge cooked in a clay vessel. Such carbonised food crusts, fragments of which were also found in Grauballe Man's stomach contents (Harild et al. Reference Harild, Robinson, Hudlebusch, Asingh and Lynnerup2007), are frequently found in prehistoric pottery. We do not know, however, whether the fish was cooked separately or with the porridge. The many sphagnum leaves, seeds from wetland plants and algal remains indicate that the porridge was made with water from a lake, pond or bog. Alternatively, Tollund Man may have drunk the water.
The porridge contained no ingredients with known intoxicating or other ‘magical’ properties, but, as in several other bog bodies, weed seeds (primarily pale persicaria) formed part of the meal. It is striking that the hitherto most detailed studies of bog body gut contents (Tollund Man and Grauballe Man) indicate that threshing waste was included in the meals. Future studies should, we hope, determine whether this was a common feature in the last meals of bog bodies. The high proportion of wild seeds to grains in several bog bodies compared with those of burnt grain stores (e.g. Henriksen & Robinson Reference Henriksen and Robinson1996) could indicate that wild seeds were an ingredient used for special occasions, including human sacrifice. They could, however, have been added to improve the nutritional value or flavour of the meal.
Nutritionally, the meal—including the threshing waste—was far from poor. The estimated nutritive content of the three dominant ingredients (barley, pale persicaria and flax) is 5380kJ (Table 1). This constitutes nearly half of the recommended daily intake of energy for a person with limited physical activity, as recommended by the Danish health authorities (Saxholt et al. Reference Saxholt, Fagt, Mathiessen and Christensen2010). The estimated proportions of the nutrients from plant ingredients in the meal are 13 per cent protein, 77 per cent carbohydrate and 10 per cent lipid. With the addition of an unknown amount of animal fat/fatty fish, the meal probably differed little from the present-day recommended intake of 10–20 per cent protein, 55–60 per cent carbohydrates and 25–30 per cent lipids (Saxholt et al. Reference Saxholt, Fagt, Mathiessen and Christensen2010). Thus, Tollund Man's final meal shows no evidence of a severe food shortage.
Table 1. The nutritive content in Tollund Man's last meal (percentage composition of nutrients in barley and flax after Madsen-Mygdal (Reference Madsen-Mygdal1937); persicaria after Hameed et al. (Reference Hameed, Dastagir and Hussain2008)).

Parasitic infections
The NPP and protein analyses show that Tollund Man was infected with three different species of parasites: tapeworm, whipworm and mawworm. While whipworm and mawworm have been found in several bog bodies (van der Sanden Reference van der Sanden1996), it is the first time that tapeworm has been reported. The most likely cause of tapeworm infection was that Tollund Man had previously consumed raw or undercooked meat infected with tapeworm cysts (Roberts et al. Reference Roberts, Janovy and Nadler2013: 332). The infestation with whipworm and mawworm reflects poor hygiene, as these are spread by contaminated food and water (Roberts et al. Reference Roberts, Janovy and Nadler2013: 378). This indicates that Iron Age lifeways (e.g. food being cooked on fires, people living closely alongside animals, and potentially limited access to clean water) affected their health.
Conclusion
Our study shows that it can be beneficial to re-analyse bog body gut contents stored in museum collections, and that combining pollen, NPP, macrofossil, steroid and protein analyses can yield further useful data. Our quantification and identification of the different ingredients in Tollund Man's last meal at a new level of detail can be used for comparison in future projects. We conclude that the plant ingredients (by weight) comprised approximately 85 per cent barley, 9 per cent pale persicaria and 5 per cent flax, with the other 20 plant taxa representing only 1 per cent of the total. Moreover, we show that early estimations of the importance of different species can be misleading. The identification of threshing waste and animal fat—probably from fish—in the meal are important new results. Ongoing analyses involving the search for aquatic lipid biomarkers will, we hope, support our conclusion regarding the intake of fish. Our study also provides new information about the handling of harvested crops and wild seeds, the preparation and cooking of Tollund Man's final meal, and his parasitic infections. Future improved residue analyses will undoubtedly add further detail concerning the gut contents, diet and, perhaps, manner of death of Northern European bog bodies, and hence contribute to our understanding of life in the Danish Early Iron Age.
Acknowledgements
We thank Catherine Jessen for improving the text of the article and two anonymous reviewers for their valuable comments.
Funding statement
This study was supported by the Research Fund of the Danish Ministry of Culture (FORM2018-0023) and the Novo Nordic Foundation (NNF18OC0032724).
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2021.98











