


Introduction
Host quality is a critical factor in determining developmental rate and success of parasitoids (Liu et al., Reference Liu, Li and Xu2013). For the immature parasitoid developing within the host, the host represents the sole source of nutrients. As a result, evaluation of host quality by the parental female parasitoid is vital to her reproductive success and offspring fitness, and a host selection trade-off results due to variation in host quality and the developmental requirements of the offspring (Harvey & Strand, Reference Harvey and Strand2002; Beckage & Gelman, Reference Beckage and Gelman2004). The life stage of the host is an important factor in determining host quality and, as such, plays a key role in host selection (Godfray, Reference Godfray1994; Colinet et al., Reference Colinet, Salin, Boivin and Hance2005; Kishani Farahani & Goldansaz, Reference Kishani Farahani and Goldansaz2013). Different host stages may represent qualities and quantities of various resources due to variation in size, physiological, behavioral, and immunological status (Chong & Oetting, Reference Chong and Oetting2006). Many studies suggest that host quality preference by parasitoids affects adult size and reproductive performance of progeny (Lampson et al., Reference Lampson, Morse and Luck1996; Harvey, Reference Harvey2005), female egg load at emergence (Liu, Reference Liu1985; Mills & Kuhlmann, Reference Mills and Kuhlmann2000), as well as sex allocation, percent parasitism and immature developmental time of parasitoids (Godfray, Reference Godfray1994; Schmidt, Reference Schmidt, Wajnberg and Hassan1994; Kishani Farahani & Goldansaz, Reference Kishani Farahani and Goldansaz2013).
The major nutritive components involved in development are triglycerides, carbohydrates, and proteins. Essential amino acids are necessary for viability, thus imbalances in dietary amino acids can lead to significant effects upon development and fitness of both immatures and adults (Dadd, Reference Dadd1985) leading to dietary restrictions on lifespan (Grandison et al., Reference Grandison, Piper and Partridge2009). Carbohydrates provide the required energy for development and also represent the mechanism by which energy is stored for future use (Dadd, Reference Dadd1985). Lipids, primarily triglyceride, are storage lipids in insects and have several roles in energetic biological demands such as flight and reproduction, both of which are imperative in the efficiency of parasitoids (Bauerfeind & Fischer, Reference Bauerfeind and Fischer2005; Fischbein et al., Reference Fischbein, Bernstein and Corley2013). Visser & Ellers (Reference Visser and Ellers2012) believed that the addition of a lipid source improved or maintained nutrient availability for parasitoids and increased their effectiveness as biological control agents within agro-ecosystems. Thus, studying the content of these resources in adults may provide an index to correlate trade-offs in decision making during the host selection process by mothers and the obtained benefits by offspring.
Numerous environmental factors including humidity, photoperiod and temperature (Pizzol et al., Reference Pizzol, Desneux, Wajnberg and Thiéry2012), in addition to biotic factors such as host age or size (Berrigan, Reference Berrigan1991; Martel et al., Reference Martel, Darrouzet and Boivin2011) are known to influence effective parasitism by Trichogramma parasitoids. To date, limited studies have documented the potential effects of host egg age on Trichogramma wasp fitness (Pak, Reference Pak1986; Moreno et al., Reference Moreno, Perez-Moreno and Marco2009). However, the effect of host nutritional quality on adult wasp fitness across multiple life history traits, has not been well studied. This study represents the first study to investigate the impact of host nutritional quality on multiple aspects of wasp fitness within a single study. Assessing multiple life history traits within a single study will provide valuable, comparative information on how and which traits are impacted by host nutritional quality, enabling us to elucidate the optimal host age to maximize wasp fitness.
The study species of the current research is Trichogramma brassicae Westwood (Hym.: Trichogrammatidae). Species belonging to the Trichogramma genus are endoparasitoids of lepidoperan eggs, although some have the potential to attack eggs of other insect taxa such as Diptera and Coleoptera (Mansfield & Mills, Reference Mansfield and Mills2002). T. brassicae is a biological control agent, which has been used against various pests (Ebrahimi et al., Reference Ebrahimi, Pintureau and Shojai1998; Van Lenteren, Reference Van Lenteren, Wratten and Gurr2000; Van Lenteren & Bueno, Reference Van Lenteren and Bueno2003; Bigler et al., Reference Bigler, Babendreier, Van Lenteren, Parra, Consoli and Zucchi2010; Parra et al., Reference Parra, Consoli and Zucchi2010; Poorjavad et al., Reference Poorjavad, Goldansaz, Machtelinckx, Tirry, Stouthamer and van Leeuwen2012) and is thus of great importance within agro-ecosystems. The current study aims to investigate the effect of host quality on adult fitness using T. brassicae as a study organism. By understanding how and which traits are impacted by host nutritional quality, we may determine the optimal host age for maximum wasp fitness, with such knowledge feeding into the mass rearing of wasps for biological control purposes. More specifically, the study aims to test the following hypotheses: (1) hosts of different ages vary in nutritional quality, (2) parasitoids reared on hosts of different ages will be provided with different amounts of protein, triglyceride and glycogen during immature development and this in turn will affect multiple aspects of their life history, including body size, longevity. and fecundity.
Materials and methods
Parasitoids and their host
Parasitoids were obtained from cultures maintained at the Biological Control Research Department of the Iranian Research Institute of Plant Protection. The original source of the cultures were parasitoids obtained from parasitized eggs of Ostrinia nubilalis Hübner (Lep.: Pyralidae), collected from northern Iran (Baboulsar Region, South of the Caspian Sea) in 2014. Parasitoids were reared at 25 ± 1°C, 50 ± 5% RH, and 16:8 L: D on eggs of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). Eggs were obtained from a culture, reared at 25 ± 1°C on wheat flour and yeast (5%), maintained at the Insectary and Quarantine Facility of University of Tehran. Approximately 20 mated female moths were kept in glass containers (500 ml) to provide eggs for experiments.
To produce adult wasps for experiments, 101–day-old eggs (high-quality eggs), 25-day-old (intermediate quality eggs) and 45-day-old eggs (low-quality eggs) were exposed to 1-day-old females for 24 h to rear wasps on different host qualities. After 24 h, the eggs were removed, and kept under controlled conditions at 25 ± 1°C, 16L: 8 D, and 50 ± 5% RH in a growth chamber and checked until emergence of adult wasps. The 25-day-old host treatment was performed separately to show the intermediate host age effects on adult wasp fitness.
Determination of glycogen, triglyceride, and protein concentration
To determine the resources obtained from high-, intermediate-, and low-quality hosts by adult wasps, 50 newly emerged wasps were exposed separately to 1-, 25-, and 45-day-old hosts for 24 h, maintained in tubes (10 × 1 cm) and prepared with 100 host eggs glued on cardboard. To avoid superparasitism by adults, only one female was introduced to each tube. Females were fed with a 10% honey solution, and maintained under controlled conditions of 25 ± 1°C, 70 ± 10 RH and 16:8 (L: D). Wasps reared on each host quality were used for the extraction of macromolecules utilizing the methods detailed below.
Glycogen determination
Fat bodies of 30 adults per treatment were removed and immersed in 1 ml of 30% KOH w/Na2SO4. Tubes containing the samples were covered with foil to avoid evaporation and boiled for 20–30 min. Tubes were subsequently shaken and cooled in ice. Two ml of 95% EtOH was added to precipitate glycogen from the digested solution. Samples were again shaken and incubated on ice for 30 min. Following the incubation on ice, tubes were centrifuged at 13,000 rpm for 30 min. Supernatant was removed and pellets (glycogen) were re-dissolved in 1 ml of distilled water and shaken. Standard Glycogen (0, 25, 50, 75, and 100 mg ml−1) was prepared before adding phenol 5%. Incubation was performed on an ice bath for 30 min. Standards and samples were read at 492 nm (Microplate reader, Awareness Co., USA) and distilled water was used as a blank (Chun & Yin, Reference Chun and Yin1998).
Triglyceride determination
A diagnostic kit from PARS-AZMOON® Co. was used to measure the amount of triglyceride in the adult parasitoids. A total of 100 wasps from each treatment group were used for triglyceride measurements. Reagent solution contained phosphate buffer (50 mM, pH 7.2), 4-chlorophenol (4 mM), Adnosine Triphosphate (2 mM), Mg2 + (15 mM), glycerokinase 0.4 kU l−1), peroxidase (2 kU l−1), lipoprotein lipase (2 kU l−1), 4-aminoantipyrine (0.5 mM) and glycerol-3-phosphate-oxidase (0.5 kU l−1). Samples (10 µl) were incubated with 10 µl distilled water and 70 µl of reagent for 20 min at 25°C (Fossati & Prencipe, Reference Fossati and Prencipe1982). The optic density (ODs) of samples and reagent as standard was read at 546 nm. The following equation was used to calculate the amount of triglyceride:
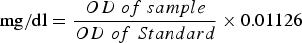 $${\rm mg/dl} = \displaystyle{{OD\; of\; sample} \over {OD\; of\; Standard}} \times 0.01126$$
$${\rm mg/dl} = \displaystyle{{OD\; of\; sample} \over {OD\; of\; Standard}} \times 0.01126$$
Protein determination
Protein concentrations were assayed according to the method described by Lowry et al. (Reference Lowry, Rosebrough, Farr and Randall1951). The method recruits reaction of Cu2+, produced by the oxidation of peptide bonds with Folin–Ciocalteu reagent. In the assay, 20 µl of the sample was added to 100 µl of reagent, and incubated for 30 min prior to reading the absorbance at 545 nm (Recommended by Ziest Chem. Co., Tehran-Iran). In total 100 adult wasps from each treatment were used in this experiment.
Morphometric measurements
Body size
To correlate body size with fitness parameters, the length of the left hind tibia of each individual was measured using a binocular microscope (0.5 × 6.3, Olympus SZ-CTV) connected to a video camera (JVC KY-F). Tibia length is a commonly used indicator of body size in parasitoid wasps and correlates strongly to other measures such as dry mass (Godfray, Reference Godfray1994). From photographed images, tibia length was determined using Image J software.
The wing loading value was obtained by calculating the ratio between the body mass and the wing area. Wing loading of females establishes a good index of their flight capacity. Lower wing loadings are considered to represent better dispersal capacities for individuals (Gilchrist & Huey Reference Gilchrist and Huey2004; Vuarin et al., Reference Vuarin, Allemand, Moiroux, Van Baaren and Gibert2012). Using weight as an index of size, for each treatment reared on high, intermediate and low-quality hosts, a minimum of 40 females were selected randomly and frozen in liquid nitrogen on emergence to be weighed on a microbalance to ± 0.1 µg (Mettler Toledo XP2U) (Ismail et al., Reference Ismail, Vernon, Hance, Pierre and van Baaren2012). At least 40 females for each host quality treatment were photographed under a binocular microscope (0.5 × 6.3, Olympus SZ-CTV) connected to a video camera (JVC KY-F). The Image J software was used to determine the area of the left wing.
Longevity
Following wasp emergence, adult longevity without food (but with access to water) was measured to estimate longevity with only capital resources available (n = 40 females reared on high, intermediate or low-quality hosts, i.e., a total of 120 females). This represents the amount of energy reserves within the body after development. Individual adults were placed in small tubes (1.5 cm in diameter and 10 cm long) and were monitored hourly until death after the first 12 h of life.
Fecundity
To compare parasitoid fecundity among treatments, 120 randomly selected newly emerged wasps (40 per host quality) were maintained in tubes (10 × 1 cm2) prepared with 100 host eggs glued on cardboards. The females were fed with a 10% honey solution. Egg cards were replaced every 12 h (until the wasp died) and maintained under controlled conditions of 25 ± 1°C, 70 ± 10 RH, and 16:8 (L: D). The preliminary test showed that adults oviposited the majority of eggs in the first 6 h of life. Subsequently, 40 newly emerged wasps from each treatment group (a total of 120 females) were selected and exposed individually to 100 host eggs for 1 h before removing the egg cards. This was repeated for the first 6 h of an individual wasp's life. Lifetime fecundity was determined by counting the number of parasitized (blackened) eggs. Parasitoids were sexed according to antennae morphological differences (Pinto, Reference Pinto1998), providing sex ratios associated with different types of hosts.
Statistical analysis
Numerical data were analyzed by Generalized Linear Models (GLM) based on a Poisson distribution and log-link function. Likelihood ratio tests were used to assess the significance of the ‘host age’ factor. The rate of produced females was analyzed by GLM based on a Binomial Logit distribution (Crawley, Reference Crawley1993; Le Lann et al., Reference Le Lann, Lodi and Ellers2014). All the recorded times were compared with Cox Proportional Hazards models. When a significant effect of the treatment was found, the tests were followed by Bonferroni's post hoc multiple comparison tests, and the two-by-two comparisons were evaluated at the Bonferroni-corrected significance level of P = 0.05 k−1, where k is the number of comparisons. Data are presented as means ±SE. All statistical analyses were performed using SAS software.
Results
Host eggs
Host age significantly affected the protein content of the host (χ2 = 94.79, P < 0.0001), with the results showing that protein amount dropped significantly in response to egg aging. One-day-old eggs contained significantly more protein than 25- and 45-day-old eggs, respectively (χ2 1 vs. 25 = 396.8, P < 0.0001, χ2 1 vs. 45 = 327.9, P < 0.0001), and 45 significantly more than 25-day-old eggs (χ2 = 9.42, P = 0.009). The amount of triglyceride in hosts was also significantly affected by host age (χ2 = 28.27, P < 0.0001). One- and 25-day-old eggs showed no significant difference in the amount of triglyceride (χ2 = 1.36, P = 0.51), while 1- and 45-day-old eggs were significantly different (χ2 = 7.47, P = 0.02), as were 25- and 45-day-old eggs (χ2 = 11.98, P = 0.0025). Finally, the glycogen content of the host was also significantly affected by host age (χ2 = 12.62, P = 0.0004). One- and 25-day-old eggs (χ2 = 15.19, P < 0.0001) and 1- and 45-day-old eggs (χ2 = 12.57, P = 0.0004) significantly differed as regards to glycogen content. However, no significant difference was revealed between 25- and 45-day-old eggs (χ2 = 0.13, P = 0.72) (fig. 1).

Fig. 1. Total concentration (Mean ± SE) of protein, triglyceride, and glycogen in 1-, 25-, and 45-day-old eggs of Ephestia kuehenliea and the adult wasps reared on these hosts.
Adult parasitoids
Host age significantly affected the protein content of the emerging wasps (χ2 = 121.53, P < 0.0001) (fig. 1). Wasps reared on 1-day-old eggs contained significantly more protein than 25- and 45-day-old, respectively (χ2 1 vs. 25 = 35.6, P < 0.0001, χ2 1 vs. 45 = 30.4, P < 0.0001), while no significant differences were observed between 25- and 45-day-old eggs (χ2 = 0.14, P = 0. 93). Host age significantly affected triglyceride amount in wasps reared on different host ages (χ2 = 36, P < 0.0001). One- and 25-day-old eggs showed significant differences in triglyceride (χ2 = 8.29, P = 0.015) as did 1- and 45-day-old eggs (χ2 = 15.6, P = 0.0004). In addition, triglyceride content differed between 25- and 45-day-old eggs (χ2 = 7.61, P = 0.022). According to our findings, glycogen amount in the emerging wasps was not affected significantly by host age (χ2 = 0.37, P = 0.544). The glycogen content of wasps reared on 1- and 25-day-old (χ2 = 1.15, P = 0.56), 1- and 45-day-old (χ2 = 1.46, P = 0.48) and 25- and 45-day-old eggs (χ2 = 0.06, P = 0.96) did not show significant differences (fig. 1).
Host age showed significant effects on wasp fecundity (χ2 = 5.67, P = 0.01). Adult wasps reared on 1-day-old hosts produced the same offspring number when compared with wasps reared on 25-day-old hosts (fig. 2). However, adult wasps laid more female eggs in 1-day-old hosts with a sex ratio of 1:3 (M: F), whereas the wasps laid more male eggs in 25- and 45-day-old hosts with a sex ratio of 2:1 and (M: F). Adult wasp longevity was significantly affected by host age (χ2 = 19.47, P < 0.0001), with wasps reared on high-quality hosts living longer than those reared on 25- and 45-day-old eggs, respectively (fig. 2). Survival curves of wasps reared on different host qualities are shown in fig. 3.

Fig. 2. Longevity (H) and fecundity (Mean ± SE) of 50 wasps, T. brassicae, reared on 1-, 25-, and 45-day-old eggs of Ephestia kuehenliea. Different letters indicate significant differences between the treatments after Bonferroni correction (P = 0.0166).

Fig. 3. Survival curves of 50 wasps, T. brassicae, reared on 1-, 25-, and 45-day-old eggs of Ephestia kuehenliea.
Tibia length (χ2 = 61.83, P < 0.0001) and weight (χ2 = 6.58, P = 0.01) were significantly affected by host age. Wasps reared on 1-day-old eggs showed higher tibia length (χ2 = 7.75, P = 0.0054) and weight (χ2 = 61.83, P < 0.0001) than wasps reared on 25-day-old eggs (fig. 4).

Fig. 4. Mean (±SE) weight (μg), tibia length (mm) of 50 wasps, T. brassicae, reared on 1-, 25-, and 45-day-old eggs of Ephestia kuehneilla. Different letters indicate significant differences between the treatments after Bonferroni correction (P = 0.0166).
Wing area was significantly affected by host age (χ2 = 53.94, P < 0.0001), with this parameter decreasing with host age (1–25-days-old) (fig. 5). Furthermore, wing loading index was significantly affected by host age (χ2 = 7.03, P = 0.009) (fig. 5).

Fig. 5. Mean (±SE) wing area (mm2) and wing loading index (mg m−2) of 50 wasps, T. brassicae, reared on 1-, 25-, and 45-day-old eggs of Ephestia kuehneilla. Different letters indicate significant differences between the treatments after Bonferroni correction (P = 0.0166).
Discussion
The current study provides the first study to investigate the effect of host quality across multiple fitness parameters within a single study. The study thus provides comparative information, enabling us to elucidate how host quality affects multiple life history traits (body size and wing loading, longevity, fecundity, and adult energy reserves) of parasitoid wasps, and ultimately wasp fitness. From a biological control perspective, this knowledge can inform the commercial mass rearing of parasitoid wasps, informing which age of host should be utilized to maximize both the proportion of female offspring and the fitness of the emerging parasitoids, and ultimately their efficacy as biological control agents.
Host eggs of different ages were shown to provide differing nutritional resources for the developing immature, thus supporting our first hypothesis. Results showed that host age, acting as a proxy for host quality, significantly affected life history traits and the nutritional reserves of T. brassicae adults. Wasps reared on high-quality hosts were bigger, with greater fecundity and longevity, and produced more female offspring compared with those reared on intermediate and low-quality hosts. Furthermore, wasps reared on high-quality hosts showed lower wing loading index compared with wasps reared on low-quality hosts. Wasps reared on high-quality hosts also contained greater energy reserves, as determined by the body content of triglyceride, glycogen and protein.
For many endoparasitic Hymenopterans such as Trichogramma spp., their eggs possess no yolk and, as such, the parasitoids lay their eggs inside the body of a host, which subsequently provides all nutrients for both embryonic and larval development (Chapman, Reference Chapman, Simpson and Douglas2012). In the body of insects, glycogen, triglyceride and protein represent the three main storage macromolecules responsible for several energetic demand processes. Phosphorylation of glycogen and triglyceride, as well as transamination of protein molecules, provides intermediate components for the electron transport system providing energy, oxygen, and water (Nation, Reference Nation2008; Arrese & Soulages, Reference Arrese and Soulages2010). The presence of these components, as obtained from the egg host, is thus essential for embryo development. In particular, it is the fatty acids stored as triglyceride, and fat reserves that are the most important reserve, used by insects to provide energy for the developing embryo (Athenstaedt & Daum, Reference Athenstaedt and Daum2006; Ziegler & Van Antwerpen, Reference Ziegler and Van Antwerpen2006). Reserves are subsequently carried through to adulthood and are depleted during periods of starvation or reproduction. In larval stages, glycogen is stored in fat bodies followed by active feeding by wasps larvae. In addition, glycogen represents the primary source of energy fuel for biological activity of larvae (Klowden, Reference Klowden2007; Chapman, Reference Chapman, Simpson and Douglas2012). Due to the precise processes behind the utilization of storage macromolecules, changes in the amounts of triglyceride, protein, and glycogen may alter the suitability of the host for the development of parasitoid offspring, and host acceptance by the parental parasitoid. This is supported by a previous study by Barrett & Schmidt (Reference Barrett and Schmidt1991), which investigated discrepancies in the amino acid content of the egg hosts of Trichogramma minutum. Whilst variation in amino acid content was evident, variation was greater in the egg hosts than in the emerging parasitoids, suggesting that metabolic compensation is occurring, although at a detriment to development. Furthermore, ovipositing females are believed to allocate eggs in accordance with the nutritional quality of the host, allocating proportionately fewer eggs to low-quality hosts (Barrett & Schmidt, Reference Barrett and Schmidt1991).
The nutritional content of host eggs is known to vary with age, as the chemical composition of the insect eggs changes rapidly from a more fluid medium to complex tissues as the egg develops. Our results showed that the total amount of protein and triglyceride in 45-day-old eggs (low-quality eggs) significantly decreased as a result of egg aging. Such changes to egg composition can further exert a negative effect on parasitism via pre-imaginal mortality, most likely the result of poorer resource availability (Brodeur & Boivin, Reference Brodeur and Boivin2004; Da Rocha et al., Reference Da Rocha, Kolberg, Mendonça and Redaelli2006). According to Benoit & Voegelé (Reference Benoit and Voegelé1979) Trichogramma parasitoids do not oviposit in old host eggs, with modification to the host tissues offering an explanation as to why Tricogramma wasps do not accept older hosts within which to oviposit.
The present study revealed that host quality significantly affected life history traits of the emerging parasitoids. Adults of T. brassicae reared on high-quality hosts (1-day-old eggs) displayed higher longevity than those reared on low-quality hosts (45-day-old eggs). Several studies have reported a relationship between host quality and parasitoid survival (Lauzière et al., Reference Lauzière, Brodeur and Pérez-Lachaud2001; Sagarra et al., Reference Sagarra, Vincent and Stewart2001; Li & Sun, Reference Li and Sun2011; Kishani Farahani & Goldansaz, Reference Kishani Farahani and Goldansaz2013). In parasitoids, like other insects, large adult body size is often related to an increase resource carry-over from the larval stage, and is manifested in higher energy reserves (López et al., Reference López, Hénaut, Cancino, Lambin, Cruz-López and Rojas2009, Kant et al., Reference Kant, Minor, Trewick and Sandanayaka2012). Our results support this, indicating that host age at oviposition affects adult survival because larger hosts provide more resources for the larval stages of the parasitoid. López et al. (Reference López, Hénaut, Cancino, Lambin, Cruz-López and Rojas2009) stated that host quality influenced the life expectancy of Diachasmimorpha longicaudata (Hym.: Braconidae) as starved females and males emerging from high-quality hosts lived significantly longer than wasps emerging from lower quality hosts.
In addition to longevity, host quality was also shown to affect gross and net fecundity of the parasitoid, with females emerging from high-quality hosts being the most fecund. According to our results, female fecundity was affected by host age, with the most fecund wasps emerging from high-quality hosts (1-day-old eggs) than low-quality hosts (45-days-old eggs). Host egg age is known to affect the fecundity and parasitism rate of Trichogramma parasitoids (Brand et al., Reference Brand, Van Dijken, Kole and Van Lenteren1984; Calvin et al., Reference Calvin, Losey, Knapp and Poston1997; Pizzol, Reference Pizzol2004; Moreno et al., Reference Moreno, Perez-Moreno and Marco2009; Pizzol et al., Reference Pizzol, Desneux, Wajnberg and Thiéry2012). In female parasitoids, fecundity is often correlated with the adult body size and quality of the food resources available to the parasitoid during development (Jervis et al., Reference Jervis, Ellers and Harvey2008; Saeki & Crowley, Reference Saeki and Crowley2013). According to obtained results, low-quality hosts contained less protein. Large amounts of proteins, such as storage proteins are used as an amino acid reservoir for morphogenesis, lipophorins responsible for the lipid transport in circulation, or vitellogenins for egg maturation (Fortes et al., Reference Fortes, Salvador and Cônsoli2011; Guo et al., Reference Guo, Dong, Ye, Li, Zhu, Fang and Hu2011). Total amount of available protein during adulthood strongly affects reproduction (vitellogenins) (Fortes et al., Reference Fortes, Salvador and Cônsoli2011). Cônsoli & Parra (Reference Cônsoli and Parra2000) showed that rearing Trichogramma galloi Zucchi and T. pretiosum Riley on artificial diets containing high amounts of protein led to an increased number of produced eggs. It seems that lower fecundity of low quality reared wasps may be due to less protein available during embryo growth and adulthood. Our results therefore show that there is a direct relation between the protein content of host eggs and the resultant number of eggs produced by adult wasps. As a consequence, rearing wasps on hosts with greater protein content, which can provide enhanced protein resources carried over into adulthood, may result in more fecund wasps. This finding has implications for biological control programs, since more fecund wasps would result in greater rates of parasitization, thus enhancing the efficacy of natural biological control.
Most parasitoid wasps, including T. brassicae, have a haplo-diploid sex determination system (Quicke, Reference Quicke1997; Beukeboom & Van de Zande, Reference Beukeboom and Van de Zande2010). This system allows the ovipositing female to control the sex of her offspring by controlling sperm access to eggs. In fact, the adult females of many parasitoid species respond to a number of environmental variables by changing offspring sex ratio. Among the variables, host type (e.g. host size, age, and species) is one of the most important factors influencing the offspring sex ratio of parasitoid wasps (Kraft & Van Nouhuys, Reference Kraft and Van Nouhuys2013; Kishani Farahani et al., Reference Kishani Farahani, Ashouri, Goldansaz, Farrokhi, Ainouche and van Baaren2015; Ueno, Reference Ueno2015). The relationships between offspring sex ratio and host quality have been investigated in many parasitoid wasps (Godin & Boivin, Reference Godin and Boivin2000; Ode & Heinz, Reference Ode and Heinze2002; Kishani Farahani & Goldansaz, Reference Kishani Farahani and Goldansaz2013; Ueno, Reference Ueno2015). Host age or quality is considered as a major factor affecting offspring sex ratio (King, Reference King, Wrensch and Ebbert1993; Ueno, Reference Ueno2015). A correlation between host quality and offspring sex ratio has commonly been demonstrated for solitary parasitoids (King, Reference King, Wrensch and Ebbert1993; Van Baaren et al., Reference Van Baaren, Landry and Boivin1999; Ode & Heinz, Reference Ode and Heinze2002), a higher proportion of female offspring tend to emerge from higher quality hosts compared with low-quality hosts. Accordingly, we showed that increased host quality results in a bias towards female production in T. brassicae. In the mass rearing of biological control agents, the number of produced females is a key factor in the success of mass release programs (Ode & Heinz, Reference Ode and Heinze2002). As such, utilization of higher quality eggs in the mass rearing of biological control agents such as T.brassicae would result in the production of a higher proportion of females, thus increasing the efficiency of biological control programs.
Previous work has suggested that wing size and shape may increase parasitoid fitness and dispersal ability in the field (Kölliker-Ott et al., Reference Kölliker-Ott, Blows and Hoffmann2003, Reference Kölliker-Ott, Bigler and Hoffmann2004) and as such, could act as a predictor of field performance of mass reared parasitoids. In the current study, we investigated the effect of host quality on parasitoid wing loading and the potential implications for parasitoid mass rearing. Results revealed that the wing loading index of T. brassicae reared on high-quality hosts were reduced when compared with wasps reared on low-quality hosts. Wing loading corresponds to the pressure exerted by the wings on the surrounding air (Gilchrist & Huey, Reference Gilchrist and Huey2004). Thus, the cost of transport is influenced in an important way by the wing surface area, which supports the body mass (Starmer & Wolf, Reference Starmer and Wolf1989; Duthie et al., Reference Duthie, Abbott and Nason2015). The lower the wing loading, the less costly the act of flight is to the individual. This reduced wing loading may facilitate flight (Gilchrist & Huey, Reference Gilchrist and Huey2004; Duthie et al., Reference Duthie, Abbott and Nason2015) in an environment where females have to move over large distances to find hosts that are patchy in distribution. Flying over large distances to find hosts is an energy demanding activity (Ruohomaki, Reference Ruohomaki1992; Ellers et al., Reference Ellers, Van Alphen and Sevenster1998). A study by Kalcounis & Brigham (Reference Kalcounis and Brigham1995) investigated the relationship between wing loading and habitat usage in bats. Results showed that bats with a higher wing loading index foraged in less cluttered areas. In the current study, the wing index suggests a higher maneuverability of wasps when reared on high-quality hosts, which will enable them to forage in environments further afield to exploit new patches, whilst utilizing less energy resources. From a biological control perspective, an enhanced dispersal activity may allow wasps to cover a greater area for foraging and searching. This in turn could increase the efficiency of mass reared wasps by increasing the potential to parasitize more hosts.
In conclusion, our results show how host nutritional quality impacts adult wasp fitness by affecting wasp life history traits. Wasps reared on high-quality hosts are provided with higher food resources (protein, glucose and triglyceride) during immature development, resulting in enhanced adult resource reserves. Higher amounts of protein and triglyceride will enhance the production of offspring, while higher glycogen amount will enhance energy reservoirs. This in turn has implications for adult fitness, resulting in larger body sizes, increased longevity, greater fecundity, and lower wing loading index. A reduced wind loading has the potential to increase adult maneuverability, aiding dispersal ability and thus access to patchy resources. Such individuals could be at an evolutionary advantage, providing their offspring with increased energy and structural resources during development. According to our results, the optimum host age for the mass rearing of this parasitoid is 1-day-old eggs of E. kuehniella, which offer greater nutritional resources, enhancing wasp fitness and, in turn, their efficiency in biological control programs.
Acknowledgements
We would like to thank Mr. Hossein Parsa for the technical support provided and Mr. Amir Javdani for assistance with insect rearing. This study was financially supported by University of Tehran, but the sponsor had no involvement in the study design, the collection, analysis, or interpretation of data, the writing of the report or in the decision to submit the paper for publication.





