


1. Introduction
The Tournaisian and the Viséan were formerly considered as series and were divided in Belgium into five stages by Conil et al. (Reference Conil, Groessens and Pirlet1977) from base to top: the Hastarian (lower Tournaisian), the Ivorian (upper Tournaisian), the Moliniacian (lower Viséan), the Livian (middle Viséan) and the Warnantian (upper Viséan). These stages were based mainly on shallow-water deposits, which give thick and relatively complete sedimentary successions rich in fossils including foraminifers, conodonts, corals and brachiopods. The Tournaisian and Viséan were redefined as two of three stages for the Mississippian by the IUGS Subcommission on Carboniferous Stratigraphy (Heckel, Reference Heckel2004) and the former stages are now substages. However, this division into only three stages of different durations seems to be rather impractical. The formal definition of shorter and distinctive time units for more precise global correlation is needed. This need is also supported by common stratigraphic practice in other systems.
According to the 2012 International Chronostratigraphic Chart, the Tournaisian and the Viséan, which correspond to the former Belgian Dinantian, lasted c. 28 Ma (c. 12 and 16 Ma, respectively) and were longer than the Silurian period, which lasted c. 24 Ma. The latter is now divided into 4 series of which 3 are divided into 7 stages of c. 2.8 Ma on average. This clearly advocates for a more detailed formal subdivision of the Mississippian in order to have a more precise global chronostratigraphic scale, comparable to those existing in other periods. The Tournaisian should become the Lower Mississippian series and the Viséan the Middle Mississippian series. These series have to be subdivided into stages. The definition of these stages could be based on the available regional subdivsions. Recent revisions of the Belgian Dinantian litho-, bio- and sequence stratigraphy and of the stages and substages were made by Hance et al. (Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002, Reference Hance, Poty and Devuyst2006a , Reference Hance, Poty and Devuyst b , c), Poty et al. (Reference Poty, Hance, Lees, Hennebert, Bultynck and Dejonghe2002, Reference Poty and Hance2006), Devuyst et al. (Reference Devuyst, Hance and Poty2006), Hance & Poty (Reference Poty and Hance2006), Poty & Hance (Reference Poty and Hance2006a , Reference Poty and Hance2006b ). All the litho- and biostratigraphical units and the substages are now very well zoned and precisely correlated across the Namur–Dinant Basin and beyond. The Belgian substages have been demonstrated to be a powerful tool for correlations with England (Hance et al. Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002), Poland (Poty et al. Reference Poty, Berkowski, Chevalier, Hance and Wrong2003), Turkey (Denayer & Aretz, Reference Denayer and Aretz2012), East European platform and the Urals (Hecker, Reference Hecker2001), South China (Hance et al. Reference Hance, Hou and Vachard2011) and North Africa (Aretz, unpubl. data).
The aim of this paper is to briefly characterize the five Belgian substages, to indicate their potential for global correlation and to initiate the discussion on potential boundaries. Transferring the Belgian outcrops and boundaries into stratotype sections and points is not the aim here; this is an independent process.
2. Hastarian (lower Tournaisian)
2.a. Definition of the substage
The base of the Hastarian was defined in the Hastière section south of Dinant (Figs 1, 2b) by Conil et al. (Reference Conil, Groessens and Pirlet1977), at the base of the shaly limestone bed 311, just above the basal 1.5-m-thick limestone bed (bed 310) of the Hastière Formation. The substage was revised and detailed by Hance & Poty (Reference Poty and Hance2006) and the Anseremme railway bridge section, 2 km south of Dinant (50°14′28″N, 4°54′43″E), was chosen as neostratotype because it is much better exposed and has been studied in more detail (Van Steenwinkel, Reference Van Steenwinkel1984, Reference Van Steenwinkel1990, Reference Van Steenwinkel, Posamentier, Summerhayes, Haq and Allen1993; Casier et al. Reference Casier, Mamet, Préat and Sandberg2004). In the Anseremme section (Fig. 2a, b), the basal bed of the Hastière Formation (no. 159) contains both conodonts (Protognathodus kockeli, Pelekysgnathus inclinatus and Pseudopolygnathus graulichi) and foraminifers, indicating the upper praesulcata Zone, and in its very basal part ‘Devonian’ (pre-Hangenberg) fossils including mainly quasiendothyrid foraminifers, phacopid trilobites and the brachiopod Sphenospira julii (Conil et al. Reference Conil, Dreesen, Lentz, Lys and Plodowski1986; Devuyst & Hance, in Poty et al. Reference Poty, Devuyst and Hance2006). The presence of limestone pebbles and rounded specimen of Campophyllum gosseleti, a rugose coral species of the pre-Hangenberg zone RC0β (Poty et al. Reference Poty, Devuyst and Hance2006), suggests that these fossils are most probably reworked from underlying levels. The Hangenberg event is not marked lithologically, but is located just below bed 159 since bed 157 yields quasyendothyrid foraminifers (DF7 Zone) and Campophyllum gosseleti. The base of the Carboniferous was drawn based on the disappearance of reworked ‘Devonian’ fauna and conodonts of the praesulcata zone, since above bed 159 there was no palaeontological criteria for defining the base of the Hastarian (Conil et al. Reference Conil, Dreesen, Lentz, Lys and Plodowski1986).

Figure 1. Distribution of Tournaisian–Viséan deposits (shaded areas) and the sedimentation areas in southern Belgium and location of the substage stratototypes. 1, Hastarian stratotype; 2, Ivorian stratotype; 3, Moliniacian stratotype; 4, Livian stratotype; 5, Warnantian stratotype; ASA – southern Avesnois sedimentation area; CSA – Condroz sedimentation area; DSA – Dinant sedimentation area; HSA – Hainaut sedimentation area; NSA – Namur sedimentation area.

Figure 2. Hastarian Substage (lower Tournaisian): (a) log of the Hastarian boundary stratotype; (b) view of the Hastarian stratotype at Anseremme south of Dinant; and (c) correlation of the Hastarian in the Namur–Dinant Basin.
In 1980, the IUGS DC Working Group recommended that the base of the Carboniferous be defined by the entry of the conodont Siphonodella sulcata within the evolutionary lineage from S. praesulcata to S. sulcata. That definition was ratified in 1991, and the base of the Hastarian Substage was considered to coincide with that of the Carboniferous. Unfortunately, in the Namur–Dinant Basin the oldest siphonodellids are absent across the Devonian–Carboniferous boundary, and the first representative of the genus (attributed to Siphonodella duplicata) was recorded in bed 174a, c. 7 m above the base of the Hastière Formation at Anseremme (Van Steenwinkel, Reference Van Steenwinkel1984; Webster & Groessens, Reference Webster and Groessens1991). Regardless, the base of the Hastarian must be taken at the GSSP of the base of the Carboniferous. The upper boundary is defined by the base of the overlying Ivorian Substage, which corresponds to the appearance of Polygnathus communis carina (Hance et al. Reference Hance, Poty and Devuyst2006c ).
2.b. Lithostratigraphy and sedimentology
In the Namur–Dinant Basin, the depositional setting during Hastarian time was a south-facing homoclinal ramp (Hance et al. Reference Hance, Poty and Devuyst2001). In the stratotype area situated in the south of the Dinant sedimentation area (Dinant SA; Fig. 1) (Poty, Reference Poty1997; Hance et al. Reference Hance, Poty and Devuyst2001), the Hastarian includes the Hastière Formation except its basal bed, the Pont d'Arcole, Landelies and Maurenne formations (Fig. 2c). The dominant facies are thick-bedded crinoidal packstones and grainstones (middle member of the Hastière and Landelies formations), thin-bedded crinoidal packstones and grainstones interbedded with shales (lower and upper members of the Hastière Formation), shales with minor crinoidal intercalations (Pont d'Arcole and Maurenne formations). The correlations (Fig. 2c) with other lithostratigraphic units in other sedimentary areas of the Namur–Dinant Basin were discussed by Hance & Poty (Reference Poty and Hance2006). The Hastarian covers the upper part of the transgressive systems tract (TST; Fig. 2c), the highstand systems tract (HST) of sequences 1 and 2 and the lowstand systems tract (LST) and a part of the TST of sequence 3 of Hance et al. (Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002).
2.c. Biostratigraphy (Fig. 2c)
2.c.1. Foraminifers
The Hastarian is covered by the foraminifer zones MFZ1 to MFZ3 and to the lower part of MFZ4 of Hance & Devuyst (in Poty et al. Reference Poty, Devuyst and Hance2006). In Belgium, the two lower members of the Hastière Formation are almost devoid of plurilocular foraminifers and correspond to the unilocular Zone of Conil et al. (Reference Conil, Groessens and Pirlet1977) (= MFZ1 – unilocular Zone). From the upper member of the Hastière Formation, plurilocular foraminifers are more common (MFZ2 – Septabrunsiina minuta Zone) but they only become diverse in the upper part of the Landelies Formation (MFZ3 – Palaeospiroplectammina tchernyshinensis Zone). They are even more diverse in the lower part of the Yvoir Formation (north Dinant and Condroz sedimentation areas) where tuberculate foraminifers enter (MFZ4 – Spinochernella brenclei Zone).
2.c.2. Conodonts
The basal bed of the Hastière Formation contains Pelekysgnathus inclinatus, Protognathodus kockeli and Pseudopolygnathus graulichi indicating the upper praesulcata Zone (Ziegler & Sandberg, Reference Ziegler and Sandberg1984; Casier et al. Reference Casier, Mamet, Préat and Sandberg2004). The Hastarian is considered to correspond to the range of siphonodellid conodonts from the entry of S. sulcata upwards (CC1 Zone of Conil et al. Reference Conil, Groessens and Pirlet1977), but there is no evidence that the sulcata Zone is represented in the Hastière Formation.
2.c.3. Rugose corals
The Hastarian comprises the Rugose Coral Zones RC1 (Hastière and Pont d'Arcole formations), RC2 (Landelies Formation) and the base of the RC3 Zone (lower part of the Yvoir Formation in the north Dinant SA) of Poty (Reference Poty1985). The RC1 – Conilophyllum Zone is marked by the arrival of Conilophyllum and Kizilia just above the basal bed of the Hastière Formation, which yields reworked corals of the RC0β Subzone. The upper part of the Zone (Pont d'Arcole Formation) yields Uralinia lobata allowing widespread correlations. The RC2 – Siphonophyllia rivagensis Zone and the base of the RC3 – Caninophyllum patulum Zone remain relatively less diverse.
2.c.4. Palynomorphs
The Hastarian is covered by four of the miospore biozones established for the Tournaisian of southern Ireland (Higgs et al. Reference Higgs, Dreesen, Dusar and Streel1992): the VI: Vallatisporites verrucosus – Retusotriletes incohatus (its base is situated in the upper part of the Strunian Substage (latest Devonian); HD: Kraeuselisporites hibernicus – Umbonatisporites distinctus; BP: Spelaeotriletes balteatus – Rugospora polyptycha; and PC: Spelaeotriletes pretiosus – Raistrickia clavata zones.
2.c.5. Other fossils
Brachiopods are common in the Hastarian, but they need to be revised to be used for the stratigraphy of the substage. No goniatite has been recorded in the Hastière Limestone.
3. Ivorian (upper Tournaisian)
3.a. Definition of the substage
The base of the Ivorian Substage, as revised and emended by Hance et al. (Reference Hance, Poty and Devuyst2006 c) is recognized by the first appearance of the conodont Polygnathus communis carina in bed 140 (Groessens, Reference Groessens1975) in the disused quarry section near the Yvoir railway station (50°19′17″N, 4°52′48″E; Figs 1, 3b). That boundary position is 2.5 m higher than the original placement by Conil et al. (Reference Conil, Groessens and Pirlet1977), at the base of the calcshale bed 134 overlying the cherty limestone bed yielding the last Siphonodella. The boundary is 17.7 m above the base of the Yvoir Formation (Fig. 3a, b). Its top coincides with the base of the Moliniacian Substage, which corresponds to the base of the Viséan.

Figure 3. Ivorian Substage (upper Tournaisian): (a) log of the Ivorian boundary stratotype; (b) view of the Ivorian stratotype at Yvoir north of Dinant; and (c) correlation of the Ivorian in the Namur–Dinant Basin.
3.b. Lithostratigraphy and sedimentology
During Ivorian time, the basin architecture evolved from the homoclinal ramp prevailing during the Hastarian to a rimmed shelf. The inner ramp/shelf facies occur in the Namur, Condroz and the Southern Avesnois sedimentation areas (Fig. 1), whereas outer ramp/shelf facies are restricted to the Dinant sedimentation area (Hance et al. Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002) where the Waulsortian build-ups developed. The Ivorian stratotype is situated in the northern part of the Dinant SA, where it comprises the Yvoir Formation except its lower part including the Hun Member, the Ourthe and Leffe formations and the lowermost part of the Sovet Formation (Fig. 3c). The dominant facies are thick-bedded crinoidal rudstones (Ourthe Formation), thin-bedded cherty crinoidal packstone and rudstone (Yvoir Formation) and cherty poorly fossiliferous wacke- to packstones (Leffe Formation).
The correlations with other lithostratigraphic units in other sedimentary areas of the Namur–Dinant Basin are given in Figure 3c, and discussed by Hance et al. (Reference Hance, Poty and Devuyst2006 c). The Ivorian succession comprises (Fig. 3c) the third-order sequence 3 (except its LST and the beginning of the TST, which belongs to the Hastarian), sequence 4 and the first metres of sequence 5 of Hance et al. (Reference Hance, Poty and Devuyst2001). The HST of sequence 4 was marked by the highest sea level recorded in the Tournaisian (‘Avins event’ of Poty, Reference Poty, Hubmann and Piller2007) and was followed by a very low LST (Hance et al. Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002) causing the end of the Waulsortian build-ups.
3.c. Biostratigraphy
3.c.1. Foraminifers
The Ivorian correlates to the upper part of MFZ4 and to MFZ5 to MFZ8 (Devuyst & Hance, in Poty et al. Reference Poty, Devuyst and Hance2006). The Yvoir and Ourthe formations and their Waulsortian lateral equivalents are almost devoid of plurilocular foraminifers, except in the upper part of the Yvoir Formation where the MFZ5 – Paraendothyra nalivkini Zone is identified. The late Ivorian Longpré Formation (and the top Waulsortian lateral equivalent) is characterized by a progressively richer fauna allowing the recognition of the MFZ6 – Tetrataxis, MFZ7 – Darjella monilis and MFZ8 – Eoparastafella morphotype 1 Zones). The basinal Leffe Formation has an impoverished foraminiferal association with some input of shallow-water elements due to tempestites (MFZ8).
3.c.2. Conodonts
The Ivorian is rich in conodonts and comprises the Polygnathus communis carina and the Scaliognathus anchoralis Zones and the lower part of the CC4 Zone (Mestognathus praebeckmanni Subzone) (Groessens in Conil et al. Reference Conil, Groessens, Laloux, Poty and Tourneur1991; Poty et al. Reference Poty, Devuyst and Hance2006).
3.c.3. Rugose corals
Corals are common and diversified in the relatively shallow-water facies (Yvoir, Ourthe, Martinrive and Longpré formations), but are rare in the deeper-water facies (Leffe Formation and Waulsortian facies). The base of the Ivorian is placed in the lower part of the RC3 – Caninophyllum patulum Zone, above the appearance of Caninophyllum patulum and Sychnoelasma urbanowitschi at the base of the Yvoir Formation and a little below the appearance of the widespread genera Cyathoclisia, Solenodendron, Bifossularia, Keyserlingophyllum and Uralinia cf. gigantea. The rest of the substage is covered by the RC4 – Sychnoelasma hawbankense Zone, except for its latest subzone (RC4β2) which is Moliniacian in age (Poty et al. Reference Poty, Devuyst and Hance2006).
3.c.4. Other fossils
Macrofossils are more abundant and diverse in the Ivorian than in the Hastarian, including mainly crinoids, brachiopods and bryozoan, but they must be revised before they can be used for Ivorian stratigraphy.
4. Moliniacian (lower Viséan)
4.a. Definition of the substage
The Moliniacian is the basal division of the Viséan in Belgium. It was revised and discussed by Devuyst et al. (Reference Devuyst, Hance and Poty2006) and its base now corresponds to the base of the Viséan stage defined by the first occurrence of the foraminifer Eoparastaffella simplex in the lineage E. ovalis – E. simplex (Devuyst & Hance, Reference Devuyst and Hance2003; Devuyst et al. Reference Devuyst, Sevastopoulo, Hance, Hou, Kalvoda and Wu2004; Devuyst, Reference Devuyst2006). In the Moliniacian stratotype of the Salet road section (50°18′40″N, 4°49′49″E; Figs 1, 4b), the first E. simplex appears in bed 124 in the Leffe Formation (Fig. 4a, b). The position of the Ivorian–Moliniacian boundary and its correlation through the Namur–Dinant Basin were problematic and discussed over a long time (Conil et al. Reference Conil, Groessens, Laloux and Poty1989), mainly because the lower part of the substage was largely missing in most of the Namur–Dinant Basin (Hance et al. Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002, Reference Hance, Poty and Devuyst2006b ; Devuyst et al. Reference Devuyst, Hance and Poty2006). Its top is defined by the base of the Livian Substage and coincides with the top of a major bentonite, the ‘Banc d'or de Bachant’.

Figure 4. Moliniacian Substage (lower Viséan): (a) log of the Moliniacian boundary stratotype; (b) view of the Moliniacian stratotype in the Salet road section north of Dinant; and (c) correlation of the Moliniacian in the Namur–Dinant Basin.
4.b. Lithostratigraphy and sedimentology
During the Moliniacian, the Namur–Dinant Basin evolved from the rimmed shelf which had an irregular topography due to the development of Waulsortian build-ups and inherited from the late Tournaisian, to a wide and shallow platform (Hance et al. Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002). The Moliniacian stratotype is situated in the middle part of the Dinant SA (Fig. 1). The substage here comprises the top of the Leffe Formation and the Molignée, Salet and Neffe formations (Fig. 4c). These formations contain only carbonated facies; cherty wackestones in the Leffe Formation; alternation of thin-bedded black pack- to grainstones (‘black marble’) and thick-bedded mudstones to wackestones in the Molignée Formation; and medium- to thick-bedded bioclastic pack- to grainstones, partly dolomitized, in the Salet and Neffe formations. The correlations with other lithostratigraphic units in other sedimentary areas of the Namur–Dinant Basin are given in Figure 4c, and described by Devuyst et al. (Reference Devuyst, Hance and Poty2006). The Moliniacian succession (Fig. 4c) comprises sequence 5 (except its lowermost part, corresponding more or less to its LST) and sequence 6 of Hance et al. (Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002). Sequence 5 is recorded only in the Dinant SA and is missing in other shallower areas, due to the important late Tournaisian sea-level drop and the resulting basin-ward shore-line migration. As a consequence, the TST and the HST never reached the shallower sedimentation areas. Sequence 6 is marked by a major flooding of the platform exposed during the time of sequence 5, and by the smoothing of the topographical irregularities.
4.c. Biostratigraphy
4.c.1. Foraminifers
The Moliniacian correlates with zones MFZ9 to MFZ11 of Devuyst & Hance (in Poty et al. Reference Poty, Devuyst and Hance2006), its base coinciding with the base of the MFZ9 – Eoparastaffella simplex Zone by definition. Foraminifers are abundant and diverse throughout the substage and it is interesting to highlight the appearance of archaediscids, a major Viséan group, marking the base of the MFZ10 – Planoarchaediscus/Ammarchaediscus Zone. In the MFZ11 – Uralodiscus rotundus Zone, archaediscids become more common in most marine facies while their distribution was previously more facies-controlled.
4.c.2. Conodonts
Conodonts are much less diverse and less useful for biostratigraphy than in the Ivorian with the exception of Gnathodus homopunctatus, whose appearance almost coincides with the base of the substage (17 m above the base in the Salet stratotype; Belka & Groessens, Reference Belka and Groessens1986; Devuyst et al. Reference Devuyst, Hance and Poty2006).
4.c.3. Rugose corals
The Moliniacian comprises the rugose coral subzones RC4β2, RC5α and RC5β. Its base correlates approximately with the base of the RC4β2, which is characterized by the appearance of the genera Haplolasma and Axophyllum and the reappearance of Clisiophyllum after its stratigraphic demise with the Hangenberg event (Poty, Reference Poty1999). The base of the RC5 – Siphonodendron Zone is marked by the entry of the genus Siphonodendron (S. ondulosum) in Western Europe (Poty, Reference Poty1984, Reference Poty1985). S. martini, the most common Siphonodendron of the Viséan, appears in the RC5β Zone.
4.c.4. Other fossils
Macrofossils, the subject of numerous but dated publications, are abundant and diverse; they now need to be revised to be used for the Moliniacian stratigraphy. In the stratotype section, the Molignée Formation is a lagerstätte which yielded, among others, very well-preserved but rare fishes, echinoids and graptolites (see Mottequin, Reference Mottequin2004 for a list of references).
5. Livian (middle Viséan)
5.a. Definition of the substage
The substage corresponds to the Cf5 Foraminifera Zone of Conil et al. (Reference Conil, Groessens and Pirlet1977), equivalent to MFZ12 of Devuyst & Hance (in Poty et al. Reference Poty, Devuyst and Hance2006), characterized by the appearance of Koskinotextularia and Pojarkovella nibelis (Paproth et al. Reference Paproth, Conil, Bless, Boonen, Carpentier, Coen, Delcambre, Deprijck, Deuzon, Dreesen, Groessens, Hance, Hennebert, Hibo, Hahn, Hahn, Hislaire, Kasig, Laloux, Lauwers, Lees, Lys, Op De Beek, Overlau, Pirlet, Poty, Ramsbottom, Streel, Swennen, Thorez, Vanguestaine, Van Steenwinckel and Vieslet1983; Conil et al. Reference Conil, Groessens, Laloux, Poty and Tourneur1991). It was revised by Poty & Hance (Reference Poty and Hance2006 a). The stratotype is a cliff situated at Lives (50°28′09″N, 4°55′44″E) on the right bank of the Meuse valley, downstream from the Beez motorway bridge (Figs 1, 5b). The base of the substage is exposed at the foot of the cliff along highway N90 (Hance, Reference Hance1979; Poty & Hance, Reference Poty and Hance2006 b); it is defined at the base of an argillaceous bed of volcanosedimentary origin (‘Banc d'or de Bachant’; bentonite L1 of Delcambre, Reference Delcambre1989), separating the Lives Formation from the underlying Neffe Formation (Fig. 5a). This is a problem because that level marks a gap between the third-order sequences 6 and 7 of Hance et al. (Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002); a new stratotype therefore has to be defined in a more continuous section. The top of the substage corresponds to the entry of the foraminifers of the Cf6 Zone of Conil et al. Reference Conil, Groessens and Pirlet1977 (= MFZ13 of Devuyst & Hance, in Poty et al. Reference Poty, Devuyst and Hance2006), which mark the base of the Warnantian Substage.

Figure 5. Livian Substage (middle Viséan): (a) log of the Livian boundary stratotype; (b) view of the Livian stratotype at Lives east of Namur (is 70 m above the road); and (c) correlation of the Livian in the Namur–Dinant Basin.
5.b. Lithostratigraphy and sedimentology
Livian sedimentation was governed by an aggrading shelf but, unlike previously, the open marine facies developed in the north while restricted facies, including evaporites, developed to the south. This inversion of the former pattern was probably related to the beginning of the Variscan shortening in the southern part of the Namur–Dinant Basin (Hance et al. Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002). The Livian stratotype is situated in the south part of the Namur SA and comprises the Lives and Grands Malades formations (Fig. 5c). The Lives Formation is organized in shallowing-upwards parasequences: in the lower Haut-le-Wastia Member, stromatolites and lime mudstones are dominant in the parasequences; the middle Corphalie Member is a 18-m-thick complex parasequence showing an evolution from open marine limestones to restricted limestones, including locally metric to decametric microbe-bryozoan build-ups (Lauwers, Reference Lauwers1992; Chevalier & Aretz, Reference Chevalier and Aretz2005); and in the upper Awirs Member, bioclastic packstones are dominant and locally cherty. The lower Maizeret Member of the Grands Malades Formation is composed of parasequences of algal boundstones and mudstones–wackestones passing to dolomites and thick beds of calcite (pseudomorphs of anhydrite); it is laterally brecciated by solution collapse giving rise to the ‘Grande Brèche’ unit (Mamet et al. Reference Mamet, Clayes, Herbosh, Préat and Wolfavisz1986). The upper Bay–Bonnet Member is composed of 8 m of stromatolitic limestones locally containing small reefs (Aretz & Chevalier, Reference Aretz, Chevalier, Álvaro, Aretz, Boulvain, Munnecke, Vachard and Vennin2007).
The lithology and the lithostratigraphy of the substage are relatively constant; correlations with other sedimentary areas of the Namur–Dinant Basin are given in Figure 5c and described by Poty & Hance (Reference Poty and Hance2006 a). The Livian succession (Fig. 5c) comprises sequence 7 of Hance et al. (Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002) with successively: the Haut-le-Wastia Member as LST; the Corphalie and Awirs members as TST; the Grands Malades Formation as HST–FSST (falling stage systems tract); and the lower part of the very short sequence 8 (usually 15–20 m total thickness) with the Bay-Bonnet Member as LST (sensu Plint & Nummedal, Reference Plint, Nummedal, Hunt and Gawthorpe2000; considered as TST by Hance et al. Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002).
5.c. Biostratigraphy
5.c.1. Foraminifers
In the Namur–Dinant Basin, the Devuyst & Hance (in Poty et al. Reference Poty, Devuyst and Hance2006) MFZ12 guide Pojarkovella nibelis usually enters at a short distance above the base of the Lives Formation, the lowermost part being almost devoid of plurilocular foraminifers. In the Livian stratotype, the first occurrence data of P. nibelis is 14.3 m above the base of the substage. However, P. nibelis was recorded at the base of the formation in a borehole near Florenne in the Dinant SA (Hance, unpubl. data). Koskinotextularia is another useful auxiliary guide entering at a similar level, low in the zone. Palaeotextularia gr. consobrina, Lituotubella sp., Rhodesinella sp., Omphalotis minima, abundant Eostaffella spp. and Endothyra spp., Nodosarchaediscus spp. and Archaediscus spp. are other typical taxa.
5.c.2. Rugose corals
The Livian is covered by the RC5γ Subzone (appearance of Siphonodendron irregulare) and the RC6 – Lithostrotion araneum Zone (Poty, Reference Poty1985; Conil et al. Reference Conil, Groessens, Laloux, Poty and Tourneur1991; Poty et al. Reference Poty, Devuyst and Hance2006). The RC5γ Subzone is characterized by the appearance of Axophyllum vaughani, Siphonodendron irregulare, Caninophyllum archiaci and Clisiophyllum garwoodi from the first open-marine bioclatic limestones close to the base of the Substage (Haut-le-Wastia Member). Lithostrotion araneum, the oldest representative of the genus Lithostrotion, appears at the base of the Corphalie Member and marks the base of the RC6 Zone. Lithostrotion vorticale, Siphonodendron sociale and Siphonophyllia siblyi appear during the Livian (Zone RC6).
5.c.3. Other fossils
Brachiopods are common, comprising mainly productids which need a revision to be used for statigraphy and the facies-restricted Composita. Goniatites are virtually absent. The facies are unsuitable for conodonts, which belong to the genera Cavusgnathus and Taphrognathus (Webster & Groessens, Reference Webster and Groessens1991).
6. Warnantian (upper Viséan)
6.a. Definition of the substage
The Warnantian Substage was defined by Conil et al. (Reference Conil, Groessens and Pirlet1977) as corresponding to their Cf6 – Neoarchaediscus Zone (= MFZ13–MFZ15 of Devuyst & Hance, in Poty et al. Reference Poty, Devuyst and Hance2006). Its base was placed at the base of the bed of bioclastic limestone marking the base of the Thon–Samson Member of the Bonne River Formation, overlying the top of the stromatolitic limestone of the Bay–Bonnet Member of the Formation (Figs 6a, b). The stratotype is in the ‘Camp de César’ disused quarry (50°28′01″N, 5°00′05″E) at Thon–Samson in the Namur SA (Figs 1, 6b). The substage was revised by Poty & Hance (Reference Poty and Hance2006 b). The Warnantian is easily divided in two units, the base of the upper unit corresponding to the base of the Janischweskina zone, which also corresponds to the base of the RC8 and can be easily traceable. That boundary is very well exposed, rich in micro- and macrofossils, and well documented (Poty et al. Reference Poty, Conil, Groessens, Laloux, Laurent, Laloux, Bouckaert, Conil, Groessens, Laurent, Overlau, Pirlet, Poty, Schiltz and Vandenberghe1988) in the Royseux-I section (Condroz SA, Fig. 1). It corresponds approximately to the base of the British Brigantian, and could be the basis for the definition of an uppermost Viséan Stage. The top of the substage currently corresponds to the base of the Namurian Stage, but it will correspond to the base of the Serpukhovian when the latter is formally defined in a GSSP.
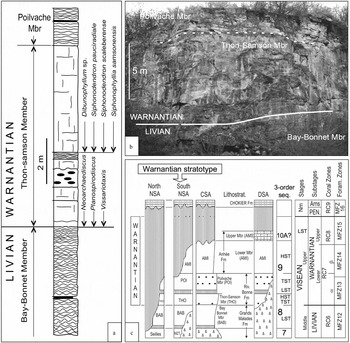
Figure 6. Warnantian Substage (upper Viséan): (a) log of the Warnantian boundary stratotype; (b) view of the Warnantian stratotype at Thon–Samson east of Namur; and (c) correlation of the Warnantian in the Namur–Dinant Basin.
6.b. Lithostratigraphy and sedimentology
As for the Livian, the Warnantian sedimentation was governed by an aggrading shelf where facies were usually more restricted than previously. The stratotype is situated in the south part of the Namur SA (Fig. 1) and only the lowest part of the substage, including the Thon–Samson Member and the lower part of the Poilvache Member (Bonne River Formation), is exposed here (Fig. 6c). The rest is lacking and Namurian siliciclastics overly directly the Poilvache Member. However, the substage is more developed in the Condroz and Dinant sedimentation areas where the Anhée Formation overlies the Bonne River Formation, but its upper part is also lacking. The Thon–Samson Member rests on the stromatolitic limestones of the Livian Bay–Bonnet Member, composed of a shallowing-upwards parasequence dominated by bioclastic limestone overlain by 5–6 m of massive crinoidal rudstone. The Poilvache Member is mainly composed of 20 m of stromatolitic limestones with some interbedded bioclastic levels.
Correlations with other sedimentary areas of the Namur–Dinant Basin are given in Figure 6c and described by Poty & Hance (Reference Poty and Hance2006 b). In the Namur–Dinant Basin, the Warnantian succession (Fig. 6c) comprises the upper part of short sequence 8, sequence 9 and possibly some signs of sequence 10 of Hance et al. (Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002). The upper part of sequence 8 corresponds to the Thon–Samson Member, with the first parasequence as TST and the massive crinoidal limestone as HST–FSST (sensu Plint & Nummedal, Reference Plint, Nummedal, Hunt and Gawthorpe2000, considered as HST in Hance et al. Reference Hance, Poty and Devuyst2001, Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002). Sequence 9 corresponds to the Poilvache Member as LST (+TST?) and the lower member of the Anhée Formation as HST (+TST, +FSST?). Sequence 10 possibly corresponds to the upper member of the Anhée Formation, which deposited in some small residual basins (Poty et al. Reference Poty, Aretz, Denayer, Aretz and Poty2011).
6.c. Biostratigraphy
6.c.1. Foraminifers
The Warnantian correlates with zones MFZ13–15 of Devuyst & Hance (in Poty et al. Reference Poty, Devuyst and Hance2006) (= Cf6 Zone of Conil et al. Reference Conil, Groessens and Pirlet1977, Reference Conil, Groessens, Laloux, Poty and Tourneur1991). Neoarchaediscus, Vissariotaxis, Planospirodiscus and Palaeotextularia with a bilaminar wall, the index taxa of the MFZ13 – Neoarchaediscus Zone, appear in the stratotype in the basal bioclastic bed of the Thon–Samson Member marking the base of the substage. The MFZ14 – Howchinia bradyana Zone covers the uppermost part of the Poilvache Member and the main part of the lower member of the Anhée Formation. The MFZ15 – Janischewskina typica Zone covers the uppermost part of the lower member and the upper member of the Anhée Formation.
6.c.2. Rugose corals
The Warnantian is covered by the RC7–Dibunophyllum and RC8–Lonsdaleia zones (Poty, Reference Poty1985; Conil et al. Reference Conil, Groessens, Laloux, Poty and Tourneur1991; Poty et al. Reference Poty, Devuyst and Hance2006). In the Warnantian stratotype, the base of the RC7 Zone is situated 3 m above the base of the substage and is characterized mainly by the appearance of Dibunophyllum sp., Siphonodendron pauciradiale and S. scaleberense. The later appearances of S. junceum and Lithostrotion maccoyanum (among others) in the Anhée Formation mark the upper part of the RC7β Subzone. Lonsdaleia and Palastraea appear in the uppermost part of the lower member of the Anhée Formation and mark the base of the RC8 Zone.
6.c.3. Other fossils
The Warnantian corresponds to the top of the Gnathodus homopunctatus – Taphrognathus transatlanticus, to the G. bilineatus conodont zones and to the goniatite fauna GF10–GF16 of Ramsbottom (in Paproth et al. Reference Paproth, Conil, Bless, Boonen, Carpentier, Coen, Delcambre, Deprijck, Deuzon, Dreesen, Groessens, Hance, Hennebert, Hibo, Hahn, Hahn, Hislaire, Kasig, Laloux, Lauwers, Lees, Lys, Op De Beek, Overlau, Pirlet, Poty, Ramsbottom, Streel, Swennen, Thorez, Vanguestaine, Van Steenwinckel and Vieslet1983), but conodonts and goniatites are very rare in the Namur–Dinant Basin. The brachiopods are common and diverse, notable for the appearance of gigantoproductids, but they need to be revised to be used for stratigraphy.
7. Geochronology
There are no radiometric data available for the Belgian Tournaisian and Viséan stage boundaries and their substages, and dates from other countries are poorly constrained stratigraphically. According to the compilation of Menning et al. (Reference Menning, Weyer, Drozdzewski and Wendt2001), the Hastarian and the Ivorian each lasted for c. 6 Ma; the Moliniacian 5–7 Ma; the Livian 3–4 Ma; and the Warnantian 8–12 Ma, depending on the timescale used (maximum–minimum ages). An attempt to calibrate the Hastarian using orbitally forced precession sequences and third-order sequences is in progress (Poty, unpubl. data).
8. Correlation of the Tournaisian and Viséan Belgian substages with other chronostratigraphic units
8.a. Correlation with the British substages
George et al. (Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976) divided the British Tournaisian and Viséan series into six stages (now substages; Waters et al. Reference Waters, Somerville, Jones, Cleal, Collinson, Waters, Besly, Dean, Stephenson, Davies, Freshney, Jackson, Mitchell, Powell, Barclay, Browne, Leveridge, Long and McLean2011) named, in ascending order: Courceyan, Chadian, Arundian, Holkerian, Asbian and Brigantian (Fig. 7). A biostratigraphic and sequence stratigraphic correlation between Belgium and SW England was proposed by Hance et al. (Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002), comprising the correlation between the Belgian and British substages. Recently, Waters et al. (Reference Waters, Somerville, Jones, Cleal, Collinson, Waters, Besly, Dean, Stephenson, Davies, Freshney, Jackson, Mitchell, Powell, Barclay, Browne, Leveridge, Long and McLean2011) revised the correlations of the Carboniferous rocks previously realized by George et al. (Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976), and discussed the British chronostratigraphic and biostratigraphic subdivisions. In the Bristol area and south Wales, the base of the Ivorian is easily recognizable in the Black Rock Limestone Subgroup by the appearance of Polygnathus communis carina (Mitchell et al. Reference Mitchell, Reynolds, Laloux and Owens1982; Hance et al. Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002). As in Belgium, this boundary is close to the base of the Caninophyllum patulum Zone of Ramsbottom & Mitchell (Reference Ramsbottom and Mitchell1980). The base of the Chadian was originally defined at the lithological change below the entry of the foraminifer genus Eosparastaffella, i.e. in the Tournaisian, but it was redefined by Waters et al. (Reference Waters, Somerville, Jones, Cleal, Collinson, Waters, Besly, Dean, Stephenson, Davies, Freshney, Jackson, Mitchell, Powell, Barclay, Browne, Leveridge, Long and McLean2011) as corresponding to the new definition of the base of the Viséan ratified in 2008 by the Subcommission on Carboniferous Stratigraphy. The Courceyan therefore corresponds to the Tournaisian. The top of the Chadian corresponds to the appearance of the archaediscid foraminifers marking the base of the Arundian. Chadian and Arundian correspond roughly to the Belgian Moliniacian.

Figure 7. Correlation of the Belgian substages with British, North American and Moscow Basin chronostratigraphic units. Correlation with the latter is mainly based on Lane & Brenckle (Reference Lane, Brenckle and Heckel2001).
The Holkerian is defined at Barker Scar, Cumbria at the lithological change corresponding to the boundary between the Dalton Beds and the Park Limestone, below the entry of the brachiopods Davidsonia carbonaria, Composita ficoides (recorded lower in the stratotype; Ramsbottom, Reference Ramsbottom1981) and Linoprotonia corrugatohemispherica, the rugose corals Axophyllum vaughani and Lithostrotion minus and foraminifers including mainly Koskinotextularia, Pojarkovella nibelis and Dainella holkeriana (Ramsbottom, Reference Ramsbottom1981). P. nibelis is the index taxon of the MFZ12 marking the base of the Livian. L. minus (= L. araneum; Poty, Reference Poty1981) is the index taxon for the RC6 Zone however, indicating that the base of the Holkerian in its stratotype is as old as the base of the RC6 Zone or younger, but does not coincide with the base of the P. nibelis Zone (and therefore the base of the Livian) as assumed. This base of the RC6 Zone can be traced in the Avon Gorge section (Bristol, England), where it occurs at the base of the Lower Clifton Down Limestone (‘Lithostrotion basaltiforme band' of Vaughan, Reference Vaughan1906).
The base of the Asbian was defined at Little Asby Scar, Ravenstonedale, Cumbria on the basis of the entry of Dibunophyllum bourtonense, Siphonodendron pauciradiale and S. junceum (George et al. Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976). It was considered as corresponding to the base of the Warnantian (Paproth et al. Reference Paproth, Conil, Bless, Boonen, Carpentier, Coen, Delcambre, Deprijck, Deuzon, Dreesen, Groessens, Hance, Hennebert, Hibo, Hahn, Hahn, Hislaire, Kasig, Laloux, Lauwers, Lees, Lys, Op De Beek, Overlau, Pirlet, Poty, Ramsbottom, Streel, Swennen, Thorez, Vanguestaine, Van Steenwinckel and Vieslet1983). However, Aretz & Nudds (Reference Aretz and Nudds2005) have shown that the coral fauna in the stratotype, including Siphonodendron martini, Siphonophyllia siblyi and Axophyllum vaughani, was typically Holkerian (coral fauna is known in the upper part of the Livian in Belgium) and not Asbian, the coral index Dibunophyllum bipartitum only present at c. 20 m higher. Lithostrotion maccoyanum and Siphonodendron junceum, which are recorded some metres above the latter, are typical species of the RC7β Zone. This indicates that the overlying Asbian levels are not from the base of the substage as considered by George et al. (Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976) and Ramsbottom (Reference Ramsbottom1981), but are much higher (RC7β Subzone), and that the section is faulted as suggested by White (unpub. Ph.D. thesis, University of Manchester, 1992), putting Livian beds into contact with beds of the lower–upper Warnantian (uppermost Asbian; Aretz & Nudds, Reference Aretz and Nudds2005). The base of the Asbian as it was originally defined by George et al. (Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976) therefore cannot be traced in the Little Asby Scar stratotype.
The Brigantian was defined at Janny Wood, near Kirkby Stephen, Cumbria on the basis of the corals Lonsdaleia floriformis, Palastrea regia and Diphyphyllum lateseptatum, brachiopods including Pugilis pugilis, the foraminifers Janischewskina operculata and Loeblichia paraammonoides (Ramsbottom, Reference Ramsbottom1981) and others recorded by White (unpub. Ph.D. thesis, University of Manchester, 1992). The rugose corals are typical of the RC8 Zone and the foraminifers of the MFZ15, and correlate easily with the Belgian upper Warnantian. However, based mainly on foraminifers (and in particular the first appearances of Howchinia bradyana), Cózar & Somerville (Reference Cózar and Somerville2004) considered that the base of the Brigantian, which is placed at the base of the Peghorn Limestone in the type section of Janny Wood, should be lowered and that the foraminiferal markers J. operculata and L. paraammonoides should be discarded. It was suggested that the new base for the Brigantian corresponds to the base of the MFZ14 and RC7β Zones in Belgium, i.e. much lower than the appearance of the former Brigantian coral index L. floriformis and P. regia (D. lateseptatum appearing earlier in Belgium in the RC7β Zone). The ranges and boundaries of the British Tournaisian and Viséan substages have therefore been shown to be inconsistent and not yet useful in defining proposed new stages for the international scale.
8.b. Correlation with western Russia
Hecker (Reference Hecker2001) discussed and established correlations for the Tournaisian and Viséan between Belgium and the East European platform and Urals, based mainly on the rugose corals zones of Poty (Reference Poty1985) and Conil (et al. Reference Conil, Groessens, Laloux, Poty and Tourneur1991) (Fig. 7). Hecker applied the Belgian substages to the Russian chronostratigraphic scale. However, here we follow the correlations proposed by Lane & Brenckle (Reference Lane, Brenckle and Heckel2001) which are based on microfossils. Polygnathus communis carina, which marks the base of the Ivorian, is recorded in the Kizelovsky horizon (Reitlinger et al. Reference Reitlinger, Vdovenko, Gubareva, Shcherbakov, Einor, Nemirovskaya and Solovieva1996) together with (or after?) Siphonodella (S. crenulata). The bases of the Moliniacian (base of the Viséan), Livian and Warnantian correspond to the bases of the Radaevsky, Tulsky and Aleksinsky horizons, respectively (Lane & Brenckle, Reference Lane, Brenckle and Heckel2001).
8.c. Correlation with North America
Mississippian macrofauna are largely endemic in North America (except Nova Scotia, e.g. rugose corals: Lewis, Reference Lewis1935; Poty, Reference Poty, Hills, Henderson and Bamber2002; brachiopods: Von Bitter & Legrand-Blain, Reference Von Bitter, Legrand-Blain and Wrong2007) with regards to the Eurasiatic, North African and Australian macrofauna (Fig. 7). They therefore do not support correlations which can only be based on foraminifers and conodonts, the former being less diversified than in Eurasia (Lane & Brenckle, Reference Lane, Brenckle and Heckel2001). As for the Hastarian, the base of the Kinderhookian by definition corresponds to the base of the Carboniferous, marked by the appearance of Siphonodella sulcata and Protognathodus kuehni at the base of the Horton Creek Formation (Collinson, Reference Collinson1961). The base of the Osagean was placed by Thomson (Reference Thomson1967) at the appearance of the P. communis carina, which corresponds to the base of the Ivorian, but Lane & Brenckle (Reference Lane, Brenckle and Heckel2001) considered that it must be placed at the base of the Gnathodus punctatus Zone as proposed by Lane (Reference Lane1974). In Belgium, that level can be traced in the lower part of the Yvoir Formation below the last siphonodellids and the appearance of P. communis carina (base of Ivorian), i.e. in the uppermost Hastarian. The Meramecian is defined with the first appearance of brachiopod species, the blastoid Pentremites conoideus and the foraminifer Globoendothyra baileyi (Kammer et al. Reference Kammer, Brenckle, Carter and Ausich1990).
None of these allow us to precisely correlate Belgian or British substages with the Meramecian, the base of which is correlated by Lane & Brenckle (Reference Lane, Brenckle and Heckel2001) with both Holkerian and Livian bases. The base of the Chesterian was defined by Lane & Brenckle (Reference Lane, Brenckle and Heckel2001) at the appearance of Neoarchaediscus and Asteroarchaediscus which approximates the base of the conodont bilineatus Zone of Lane et al. (Reference Lane, Sandberg and Ziegler1980). Neoarchaediscus is the foraminifer index for the base of the Warnantian (base of MFZ13 – Neoarchaediscus Zone), the base of the bilineatus Zone is situated in the MFZ13 Zone (Conil et al. Reference Conil, Groessens, Laloux, Poty and Tourneur1991) and Asteroarchaediscus appears later in the MFZ14 – Howchinia bradyana Zone (Devuyst & Hance in Poty et al. Reference Poty, Devuyst and Hance2006). The base of the Chesterian is therefore situated inside the lower Warnantian, as considered by Lane & Brenckle (Reference Lane, Brenckle and Heckel2001).
8.d. Correlation with South China
Hance et al. (Reference Hance, Hou and Vachard2011) reviewed the foraminifers of the Tournaisian and Viséan of South China (Hunan, Guizhou and Guangxi) and applied the Belgian foraminiferal zonation to the Chinese biostratigraphy. The base of the P. communis carina Zone was recognized in the lower part of the upper member of the Long'an Formation in the Longdianshan section (Guangxi) by Coen et al. (Reference Coen, Groessens and Sevastopulo2004). It is relatively well constrained by the record of Siphonodella obsoleta and S. cooperi in the lower member of the formation and MFZ6 foraminifers above, allowing the Hastarian–Ivorian boundary to be recognized. The stratotype of the Tournaisian–Viséan boundary, and therefore the Ivorian–Moliniacian boundary, was placed in the Pengchong section (Guangxi) in 2008. The boundary was also recognized in the Yajiao, Huaqiao Farm and Huilong sections (Devuyst, Reference Devuyst2006; Jin et al. Reference Jin, Devuyst, Hance, Poty, Aretz, Yin and Hou2007; Hance et al. Reference Hance, Hou and Vachard2011). The entry of Pojarkovella nibelis allows the base of the Livian to be recognized in all four sections and in the Mopanshan section (Devuyst, Reference Devuyst2006; Hance et al. Reference Hance, Hou and Vachard2011). In the latter, primitive specimens of Pojarkovella precede P. nibelis (Hance et al. Reference Hance, Hou and Vachard2011). The MFZ13–Neoarchaediscus Zone, which marks the base of the Warnantian, has not yet been identified in South China, but it is probably time-equivalent to siliciclastic formations yielding no foraminifers (e.g. Ceshui Formation in Hunan consisting of sandstones, siltstones, shales and coal measures; Hance et al. Reference Hance, Hou and Vachard2011; upper part of the Xiangbai Sandstone in Guizhou).
9. Conclusion
Since their definition by Conil et al. in Reference Conil, Groessens and Pirlet1977, the Belgian Tournaisian and Viséan substages were the subject of a very large number of papers and are now very well documented. They are easily recognizable and can be precisely correlated by biostratigraphy and sequence stratigraphy, not only in Europe (Hance et al. Reference Hance, Poty, Devuyst, Hills, Henderson and Bamber2002) but all along Eurasia (e.g. Hance et al. Reference Hance, Hou and Vachard2011 for South China). In contrast, the British substages were not much refined or emended since their definition by George et al. in Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976 (Waters, in Waters et al. Reference Waters, Somerville, Jones, Cleal, Collinson, Waters, Besly, Dean, Stephenson, Davies, Freshney, Jackson, Mitchell, Powell, Barclay, Browne, Leveridge, Long and McLean2011). To be understood, dated and correlated, they now need to be translated into the Belgian biostratigraphy and chronostratigraphy. Since the Belgian substages are very well defined and constrained, they could be the basis for the chronostratigraphic subdivision of the Tournaisian and Viséan. This decision is independent of the rank of the Tournaisian and Viséan which could be recognized as series or, as we prefer, stages.
Acknowledgements
We wish to thank the two anonymous referees for their probing but helpful comments, suggestions and corrections. This research received no specific grant from any funding agency, commercial or not-for-profit sectors.







