INTRODUCTION
Extinctions have played key roles in shaping island biotas and ecosystems. Reduced land area, dispersal barriers and biota with insular adaptations and low population sizes make island biotas sensitive to environmental, climatic and anthropogenic pressures and therefore particularly susceptible to extinction (e.g. Manne et al. Reference Manne, Brooks and Pimm1999; Karels et al. Reference Karels, Dobson, Trevino and Skibiel2008). For example, >90% of historic bird extinctions were on islands (Johnson & Stattersfield Reference Johnson and Stattersfield1990). Whilst the Quaternary fossil record has revealed several ‘natural’ extinction events on islands (e.g. Louys et al. Reference Louys, Curnoe and Tong2007), these are largely on islands connected to the mainland during glacials. Most island extinctions have arisen via anthropogenic impacts. Extinctions and introductions of novel species have transformed island biotic and ecological systems, as the traits of introduced species may not match those of the biota they replace (e.g. Lee et al. Reference Lee, Wood and Rogers2010). Projects are now underway on islands around the world to restore ecological processes lost through extinction.
Here, we review the timing and main historic drivers of extinctions on islands. In order to distil such a complex and detailed topic, we identify common patterns in extinction chronologies between islands and present a generalized model for anthropogenic extinctions. We then discuss the model through reviewing in greater depth the rich fossil, archaeological and historical records of New Zealand and the Mascarenes. These two well-documented archipelagos, with contrasting environments, climates and Holocene settlement histories, present ideal case studies for testing the general applicability of the proposed model. Finally, we discuss how species introductions have altered the functional composition of island biotas, review some recent efforts where palaeoecological records have helped to better inform and guide restoration of island ecosystems and discuss the continuing threats to, and restoration of, island ecosystems.
PLEISTOCENE ENVIRONMENTAL AND CLIMATIC DRIVERS OF EXTINCTIONS ON ISLANDS
Changing sea levels during the Pleistocene created land bridges to islands or reduced island areas, facilitating migrations and extinction. For example, during the late Middle Pleistocene, the formation of a land bridge between the continental island of Sicily and the European mainland led to the replacement of the highly endemic Sicilian fauna by mainland fauna (Bonfiglio et al. Reference Bonfiglio, Mangano, Marra, Masini, Pavia and Petruso2002). Details of such events are difficult to resolve because the fossil record becomes more incomplete and dating less precise with age. Moreover, the Pleistocene arrival of hominins on islands is poorly constrained (small populations may be difficult to detect in archaeological records) and likely coincided with significant environmental and climatic change. In such instances, disentangling the contributions of environmental and anthropogenic drivers of island extinctions can prove as complex as it has been for continents (e.g. Barnosky et al. Reference Barnosky, Koch, Feranec, Wing and Shabel2004).
Island South East Asia provides examples of prehistoric extinctions caused by environmental change. During glacials, most current Sundanese islands were connected by savannah corridors that facilitated dispersal of terrestrial biota (Heaney Reference Heaney1991; Bird et al. Reference Bird, Taylor and Hunt2005; Louys & Meijaard Reference Louys and Meijaard2010). While several faunal turnover events were associated with glacial connections, the interglacial loss of savannahs is thought to be responsible for most Pleistocene large mammal extinctions in this region (Louys et al. Reference Louys, Curnoe and Tong2007; Louys Reference Louys and Elewa2008, Reference Louys2014). The earliest Homo on Java, Indonesia, date to the Early Pleistocene (Swisher et al. Reference Swisher, Curtis, van den Bergh, Suprijo and Widiasmoro1994), but there is no evidence suggesting that they contributed to local extinctions (Louys et al. Reference Louys, Curnoe and Tong2007). Moreover, the appearance of hominins on islands in the region at c. >200 ka (van den Bergh et al. Reference Van Den Bergh, Li, Brumm, Grün, Yurnaldi, Moore, Kurniawan, Setiawan, Aziz, Roberts and Storey2016) does not coincide with extinctions, even though they hunted local faunas (Morwood et al. Reference Morwood, Sutikna, Saptomo, Westaway, Due and Moore2008; Piper et al. Reference Piper, Rabett and Kurui2008; van den Bergh et al. Reference Van Den Bergh, Li, Brumm, Grün, Yurnaldi, Moore, Kurniawan, Setiawan, Aziz, Roberts and Storey2016). On some islands of Nusa Tenggara, Indonesia, extinctions either pre-dated or significantly post-dated the arrival of humans, implicating environmental or natural demographic processes for the former and technological innovations for the latter (e.g. Glover Reference Glover1986; Louys et al. Reference Louys, Price and O'Connor2016).
On Bermuda, rising sea levels at c. 400 ka reduced land area and caused the extinction of four endemic bird species (Olson & Wingate Reference Olson and Wingate2000; Hearty et al. Reference Hearty, Olson, Kaufman, Edwards and Cheng2004). However, on most remote oceanic archipelagos, biotic communities were resilient to major climatic and environmental change throughout the Pleistocene, and background rates of extinction were exceedingly low. For example, in New Zealand, there are no known extinctions during the 50 ka preceding human settlement, and evidence suggests that many species tracked changes in habitat as the climate warmed after the last glacial maximum (Rawlence et al. Reference Rawlence, Metcalf, Wood, Worthy, Austin and Cooper2012).
HOLOCENE ANTHROPOGENIC DRIVERS OF EXTINCTIONS ON ISLANDS
Extinction rates on islands increased dramatically during the Holocene. Even on islands settled during the Pleistocene, extinction rates increased after human populations became more technologically advanced and less nomadic. For example, in island South East Asia, the development of complex hunting tools and the development of swidden agriculture and large-scale deforestation at the beginning of the Holocene (e.g. Maloney Reference Maloney1980, Reference Maloney1985; Rabett & Piper Reference Rabett and Piper2012; Hunt & Rabett Reference Hunt and Rabett2014) contributed more to extinctions and extirpations than initial Pleistocene colonization (Sodhi et al. Reference Sodhi, Koh, Brook and Ng2004; Corlett Reference Corlett2007; Louys Reference Louys and Elewa2008; Morwood et al. Reference Morwood, Sutikna, Saptomo, Westaway, Due and Moore2008). Apart from some inter-island variability, there is overall a remarkable consistency to the progression of anthropogenic extinctions on islands first visited or settled during the Holocene. We summarize this through a generalized model for the process presented below (Fig. 1).

Figure 1 Hypothetical curves describing general patterns in the number of predator introductions and faunal extinctions (cumulative) and the rate of forest loss associated with human visitation and Holocene settlement of islands.
Holocene extinction phase 1: pre-settlement visitation
Many islands were visited by people prior to settlement, and others were visited but never settled, which often resulted in extinctions due to the introduction of alien predators. For example, the house mouse (Mus musculus) was introduced to Madeira no later than c. 903–1036 CE, some 400 years prior to settlement. Its impact may have been severe, especially on seabirds and small flightless birds (Rando et al. Reference Rando, Pieper and Alcover2014). The pre-settlement introduction of mammals in the Mascarenes also caused several bird extinctions (Cheke Reference Cheke2013). The introduction of the black rat (Rattus rattus) to these islands by Arab traders during the 14th century (Cheke Reference Cheke2013; Hume Reference Hume, Göhlich and Kroh2013) caused the extinction of a large skink (Leiolopisma mauritiana), an endemic starling (Cryptopsar ischyrhynchus; Hume Reference Hume2014a) and at least three undescribed passerine species (Hume Reference Hume, Göhlich and Kroh2013). On sub-Antarctic Macquarie Island, which has never been permanently settled, an endemic parakeet became extinct following the introduction of rabbits, which increased feral cat populations, causing increased predation on the parakeets (Taylor Reference Taylor1979).
Holocene extinction phase 2: initial or prehistoric settlement events
During the Holocene, many islands were first settled by a founding group, and then much later by Europeans. Others had multiple prehistoric settlement events. Almost always these coincided with an increase in extinctions, though the severity and rate of extinction varied between islands (e.g. Turvey et al. Reference Turvey, Oliver, Narganes and Rye2007; Perry et al. Reference Perry, Wheeler, Wood and Wilmshurst2014) due to factors such as composition of the native fauna and which commensal species were introduced. The three major drivers of extinction associated with initial or prehistoric human settlement of islands were the introduction of predators (and any associated pathogens or diseases), hunting and vegetation burning.
Alien species
While introduced cultigens appear to have had relatively little impact on native plant communities (other than indirectly through forest clearance), introduced fauna drove extinctions in biota that are susceptible to the specific traits of the introduced species. For example, the introduction of the Pacific rat (Rattus exulans) caused extinctions among small, flightless and ground-nesting birds, terrestrial beetles and numerous other invertebrates on east Polynesian islands (Worthy & Holdaway Reference Worthy and Holdaway2002; Roff & Roff Reference Roff and Roff2003; Steadman Reference Steadman2006; Hunt Reference Hunt2007; Liebherr & Porch Reference Liebherr and Porch2015).
Other taxa intentionally introduced included fauna for protein resources (e.g. goats (Capra hircus) and pigs (Sus scrofa)) and hunting aids such as dogs (Canis familiaris; Schüle Reference Schüle1993; Prebble Reference Prebble2008; Anderson Reference Anderson2009). On some remote oceanic islands (e.g. New Zealand), following the initial establishment phase, there is little or no evidence of any subsequent introductions during the prehistoric settlement period, suggesting that travel back to homelands was limited after a few centuries (Fig. 1). Further introductions of alien taxa only occurred later in subsequent settlement phases.
Hunting
Insular megafauna (the largest animals within an island ecosystem irrespective of actual body mass; sensu Hansen & Galetti Reference Hansen and Galetti2009) typically disappeared rapidly after human arrival during the Holocene (e.g. Perry et al. Reference Perry, Wheeler, Wood and Wilmshurst2014). Factors implicated in their loss include predation by commensal species (Fleming Reference Fleming1969), environmental change (Clarke et al. Reference Clarke, Miller, Fogel, Chivas and Murray-Wallace2006), loss of prey and habitat (Worthy & Holdaway Reference Worthy and Holdaway2002: 335) and introduced pathogens (Bover & Alcover Reference Bover and Alcover2003). However, hunting was the key driver. Extensive archaeological bone middens and historic records show that this was the case with birds c. >4 kg in New Zealand (Duncan et al. Reference Duncan, Blackburn and Worthy2002; Worthy & Holdaway Reference Worthy and Holdaway2002; Roff & Roff Reference Roff and Roff2003; Perry et al. Reference Perry, Wheeler, Wood and Wilmshurst2014), and similarly with large birds and reptiles of Indian (Cheke & Bour Reference Cheke, Bour and Gerlach2014) and Pacific Ocean islands (e.g. Steadman et al. Reference Steadman, Pregill and Burley2002). A possible exception is the extinction of many Mediterranean island megafauna species, because although their extinctions closely coincide with human arrival, there is no definitive evidence of human involvement (Bover & Alcover Reference Bover and Alcover2003; Vigne et al. Reference Vigne, Zazzo, Saliège, Poplin, Guilaine and Simmons2009; Zazzo et al. Reference Zazzo, Lebon, Quiles, Reiche and Vigne2015; Bover et al. Reference Bover, Valenzuela, Torres, Cooper, Pons and Alcover2016). The case for hunting driving Holocene extinctions of megafauna is nevertheless well established on islands compared to continents, where the roles of humans and climate change remain contested.
Fire
Prehistoric settlement of islands during the Holocene was often accompanied by rapid loss of natural vegetation (particularly forest) through fire, the rate of which declined as dry, easily burnt areas were lost (Fig. 1). In dry, fire-prone regions with low rates of natural ignition, modelling shows prehistoric deforestation required intentional burning rather than random ignitions (Perry et al. Reference Perry, Wilmshurst, McGlone, McWethy and Whitlock2012). This may have been to clear forest for swidden agriculture (e.g. Weisler Reference Weisler1995; Bowman et al. Reference Bowman, Balch, Artaxo, Bond, Cochrane and D'Antonia2011; Wilmshurst et al. Reference Wilmshurst, Moar, Wood, Bellingham, Findlater, Robinson and Stone2014), improving access to resources (McGlone et al. Reference McGlone, Wilmshurst and Leach2005) or even for hunting. Initial rates of vegetation loss may have been accelerated by positive feedback that promoted early-successional flammable plant species (Perry et al. Reference Perry, Wilmshurst, McGlone, McWethy and Whitlock2012). On larger, climatically diverse islands, such as New Zealand, forest was cleared from dry, lowland regions within decades (McWethy et al. Reference McWethy, Wilmshurst, Whitlock, Wood and McGlone2014), but remained at higher elevations and in wetter areas. Conversely, many smaller islands suffered near-total loss of forests (<2% remaining in Mauritius; Page & D'Argent Reference Page and D'Argent1997; Cheke & Hume Reference Cheke and Hume2008) or complete clearance (e.g. Rapa Nui; Flenley et al. Reference Flenley, King, Jackson, Chew, Teller and Prentice1991). Deforestation not only destroyed faunal habitat, but also drove plant extinctions (e.g. Hunt Reference Hunt2007; Prebble & Dowe Reference Prebble and Dowe2008; Athens Reference Athens2009; Connor et al. Reference Connor, van Leeuwen, Rittenour, van der Knaap, Ammann and Björck2012).
Holocene extinction phase 3: European settlement
Many islands initially settled during prehistory were subsequently colonized by Europeans (Grove Reference Grove1996). Others were first settled during recent times. The same drivers of extinction associated with prehistoric or initial settlement are often replicated in recent – or European – settlement phases, but with slight variations (Fig. 1).
New suites of alien species, introduced accidentally or deliberately by acclimatization societies (sensu Dunlap Reference Dunlap1997), or for biological control or economic reasons, increased the functional diversity of predator communities, thereby putting a wider range of native species at risk and resulting in further extinctions (Fig. 1). For example, the introduction of the weasel (Mustela nivalis) to Mallorca and Menorca c. 2 ka after initial settlement extirpated the endemic lizard Podarcis lilfordi from both islands (Valenzuela & Alcover Reference Valenzuela and Alcover2013). The lizard is now restricted to weasel-free islets. Another clear demonstration of this pattern is the positive relationship between the number of bird extinctions and introduced predator species following European arrival on oceanic islands (Blackburn et al. Reference Blackburn, Cassey, Duncan, Evans and Gaston2004). Herbivore introductions have led to declines and extinctions of plant species (de Lange et al. Reference de Lange, Norton, Heenan, Courtney, Molloy and Ogle2004) and increased soil erosion, degrading habitat quality and leading to extinctions. For example, erosion and a decline in palm forest following the rabbit and goat introduction to Round Island (Mauritius) in 1840 may have contributed to the extinction of the burrowing boa (Bolyeria multocarinata; Bullock & North Reference Bullock and North1984). Invasive plants have also contributed to the displacement and decline of native island plant species (e.g. Meyer & Florence Reference Meyer and Florence1996; Loope et al. Reference Loope, Hughes, Meyer, Foxcroft, Pyšek, Richardson and Genovesi2013). Although there are far fewer examples of island plant extinctions than animal extinctions, the rate of plant species naturalizations continues steadily, and there may be an extinction debt yet to be realized (Sax & Gaines Reference Sax and Gaines2008).
Hunting or harvesting of island fauna continued during the historic settlement phase, but, as many of the large, slow-reproducing species that are most susceptible to hunting had already gone extinct following initial settlement, these activities resulted in fewer extinctions. However, over-harvesting during the third phase has been responsible for continuing loss of a wide range of island species, both terrestrial and marine (Thaman Reference Thaman2002).
Increased forest clearance (Fig. 1) for timber and agriculture accompanied historical settlement, causing significant reductions of some landsnail and beetle faunas (e.g. Goodfriend et al. Reference Goodfriend, Cameron and Cook1994; Griffiths & Florens Reference Griffiths and Florens2006; Terzopoulou et al. Reference Terzopoulou, Rigal, Whittaker, Borges and Triantis2015). In Mauritius, most plant extinctions (70 species, or 10.4% of the Mauritian flora; Atkinson & Sevathian Reference Atkinson and Sevathian2007) resulted from forest clearance and over-grazing (Page & D'Argent Reference Page and D'Argent1997; Atkinson & Sevathian Reference Atkinson and Sevathian2007; Cheke & Hume Reference Cheke and Hume2008). Over-grazing has also been implicated in plant losses on the Galápagos (Bush et al. Reference Bush, Restrepo and Collins2014) and in New Zealand (de Lange et al. Reference de Lange, Norton, Heenan, Courtney, Molloy and Ogle2004). On some islands, rates of forest loss have declined (e.g. Ewers et al. Reference Ewers, Kliskey, Walker, Rutledge, Harding and Didham2006), but on others they continue to rise (e.g. Margono et al. Reference Margono, Potapov, Turubanova, Stolle and Hansen2014).
In addition to these three pervasive factors, several new drivers are associated with more recent extinctions. Draining of wetlands for agriculture and disease prevention (affecting up to 87% of global wetlands since 1700 CE; Davidson Reference Davidson2014) has contributed to island extinctions (Steadman Reference Steadman, Reaka-Kudla, Wilson and Wilson1996). Novel pathogens and diseases have caused the decline of many island species (Wikelski et al. Reference Wikelski, Foufopoulos, Vargas and Snell2004), a recent example being Hawaiian bird species declines due to mosquito-borne avian malaria (Warner Reference Warner1968). The potential role of pathogens and diseases in older island extinctions has been suggested (e.g. Bover & Alcover Reference Bover and Alcover2003) and may have been more significant than appreciated. A convincing demonstration is the Christmas Island rat (Rattus macleari), for which DNA of a novel pathogenic trypanosome has been detected in museum specimens collected after the introduction of black rats (Wyatt et al. Reference Wyatt, Campos, Gilbert, Kolokotronis, Hynes and DeSalle2008). Specimen collection for scientific purposes is also a relatively recent pressure, but its role in causing extinctions has been debated (Minteer et al. Reference Minteer, Collins, Love and Puschendorf2014; Rocha et al. Reference Rocha, Aleixo, Allen, Almeda, Baldwin and Barclay2014) and is perhaps overstated. Collection may not independently drive extinction, but collecting may have at least hastened the extinctions of some island species that were already in decline (e.g. huia, Heteralocha acutirostris).
General model of Holocene anthropogenic extinctions on islands: case studies
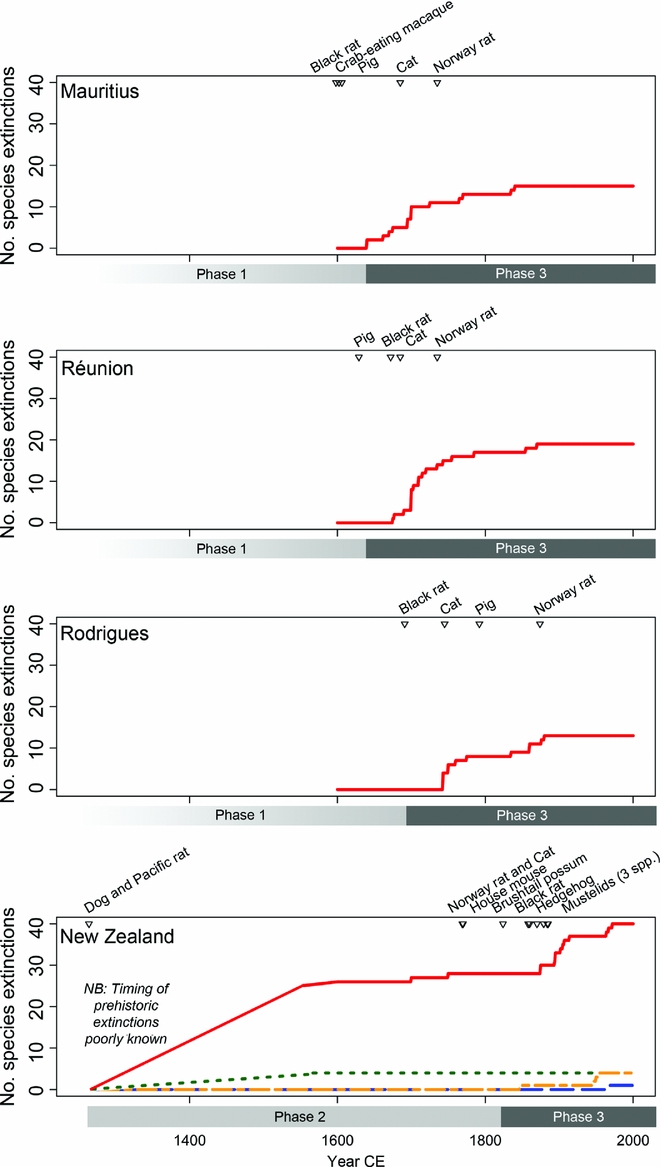
Resolving the sequence and timing of anthropogenic impacts and associated extinctions depends on robust fossil, archaeological and historical records and large numbers of accurately dated specimens. Very few islands meet all of these criteria. However, the Mascarenes and New Zealand offer rich, long-term records of species extinctions and introductions (Fig. 2). The Mascarenes were visited prior to settlement (phase 1), during which time the black rat, crab-eating macaque (Macaca fascicularis) and pig were introduced (Fig. 2), causing faunal extinctions (Cheke Reference Cheke2013). The Rodrigues blue pigeon (Alectroenas payandeei) and a Mauritius population of Réunion harrier (Circus maillardi) disappeared, while much of the native fauna declined (Hume Reference Hume, Göhlich and Kroh2013). However, most endemic species survived until historic settlement (phase 3). Most predatory mammal introductions occurred before 1900 (Fig. 2), reflecting a declining rate of introductions (as in Fig. 1). Increased bird extinction rates on all three main islands (Mauritius, Rodrigues and Réunion) coincided with the local timing of predatory mammal introductions (Fig. 2). On Mauritius and Réunion, this occurred in 1638–1695, causing the loss of all flightless birds, and increased hunting pressure and habitat destruction resulted in a second wave of extinctions in 1710–1840. Rodrigues, the most isolated of the islands, remained unsettled until the late 18th century, but lost almost all of its native fauna in 1726–1761 (Hume Reference Hume, Göhlich and Kroh2013). The extinction of large tortoises on Mauritius (c. 1715), Rodrigues (c. 1795) and Réunion (c. 1850) appears to have been related to harvesting rather than introduced predators or habitat loss (Cheke Reference Cheke2013).

Figure 2 Holocene extinction phases, timing of introductions of predatory mammal species and cumulative bird extinction curves (red solid line) for the Mascarenes and New Zealand. For New Zealand, cumulative extinction curves for reptiles/amphibians (green short-dashed line), mammals (blue long-dashed line) and plants (yellow dot-dashed line) are also shown. Data for Mascarenes are from Cheke (Reference Cheke2013) and for New Zealand are from Wood (Reference Wood and MacLeod2013) and King (Reference King1990). For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.
There is no evidence for pre-settlement visitation (phase 1) to New Zealand, which was first settled by Polynesians during the early 13th century CE (phase 2; Wilmshurst et al. Reference Wilmshurst, Anderson, Higham and Worthy2008), when two predatory mammal species were introduced (the Pacific rat and dog; Fig. 2). After this, there is no evidence for further species introductions until the European settlement phase, beginning in the late 18th century (as in Fig. 1). A period of rapid deforestation (McWethy et al. Reference McWethy, Wilmshurst, Whitlock, Wood and McGlone2014) and extinction followed initial settlement. The New Zealand moa (Aves: Dinornithiformes) constitutes perhaps the best resolved prehistoric island extinction, with hundreds of radiocarbon dates revealing their extinction within 200 years (Perry et al. Reference Perry, Wheeler, Wood and Wilmshurst2014). Many other large bird species disappeared at the same time (e.g. geese (Cnemiornis spp.) and adzebills (Aptornis spp.)), suggesting that these first two centuries were a time of rapid exploitation and extermination (as in Fig. 1). There is also evidence for rapid phase 2 extinctions on larger islands in Near Oceania (Steadman et al. Reference Steadman, Pregill and Burley2002; Steadman Reference Steadman2006) and high prehistoric extinction rates of birds on other Pacific islands (Duncan et al. Reference Duncan, Boyer and Blackburn2013). While it has been suggested that humans coexisted with extinct species on some Pacific islands for much longer periods (Steadman Reference Steadman2006), uncertainty in the exact timing of human settlement on some island groups may partly account for this. Recent work has shown that settlement occurred more recently than previously thought on many of these islands (Wilmshurst et al. Reference Wilmshurst, Moar, Wood, Bellingham, Findlater, Robinson and Stone2014), supporting the idea of rapid extinctions following human arrival. With few or no subsequent species introductions and a reduction in vegetation clearance, the rate of extinction likely declined with time in phase 2 (as in Fig. 1). After the arrival of James Cook's expedition in 1769 and the likely contemporaneous introduction of cats (Felis domesticus) and Norway rats (Rattus norvegicus), the second period of New Zealand settlement began in the early 19th century (i.e. phase 3). The rate of predatory mammal introductions declined (as in Fig. 1): seven species became established during the 19th century, but none have done so since (Fig. 2). The combined impacts of these mammals resulted in an increased rate of faunal extinctions in the latter part of the 19th century (Fig. 2). With no more predatory mammal introductions, the extinction rate declined by the early 20th century, with the exception of three extinctions on Big South Cape Island following the arrival of rats there in the 1960s (Fig. 2) (Bell et al. Reference Bell, Bell and Merton2016).
ECOLOGICAL CONSEQUENCES OF ISLAND EXTINCTIONS
There is a wide range of literature exploring how extinctions can sever ecological interactions, affecting ecosystem processes and functions on islands (e.g. Meehan et al. Reference Meehan, McConkey and Drake2002; Johnson Reference Johnson2009; Hansen Reference Hansen2010; Lee et al. Reference Lee, Wood and Rogers2010). The effects can cascade across ecological networks and take centuries to be fully realized (Brodie et al. Reference Brodie, Aslan, Rogers, Redford and Maron2014). For species with specialized interactions, the extinction of one species may cause the loss of others (i.e. co-extinction). On islands, co-extinction is a risk for specialized insect-pollinated (Dunn et al. Reference Dunn, Harris, Colwell, Koh and Sodhi2009) or faunal-dispersed plants (e.g. Meehan et al. Reference Meehan, McConkey and Drake2002) and has caused the loss of many host-specific parasites (e.g. Mey Reference Mey1990; Wood et al. Reference Wood, Wilmshurst, Rawlence, Bonner, Worthy, Kinsella and Cooper2013).
Extinctions are biased towards taxa with susceptible traits (Roff & Roff Reference Roff and Roff2003; Duncan & Blackburn Reference Duncan and Blackburn2004), causing heavy losses in certain trophic groups (Hansen & Galetti Reference Hansen and Galetti2009; Johnson Reference Johnson2009). For example, large vertebrate herbivores are prone to extinction, causing ‘trophic skew’ (Duffy Reference Duffy2003), altering plant communities and ecosystem function. In New Zealand, the loss of all c. >4-kg avian herbivore species may have altered dynamics in avian-induced vegetation communities (Lee et al. Reference Lee, Wood and Rogers2010) and had minor effects on the abundances of some forest plants (Wood & Wilmshurst Reference Wood and Wilmshurst2017). Loss of ecosystem engineer species can also have profound effects. Seabirds, for example, transport large amounts of nutrients into terrestrial ecosystems and burrow into soils, influencing vegetation (Smith et al. Reference Smith, Mulder, Ellis, Mulder, Anderson, Towns and Bellingham2011), but, since human settlement, seabird communities have been severely impacted on islands around the world (e.g. Steadman Reference Steadman1995; Worthy & Holdaway Reference Worthy and Holdaway2002; Rando & Alcover Reference Rando and Alcover2007).
While some exotic species may perform ecological roles approximating those of extinct species (Hansen Reference Hansen2010), there are many specialized ecological roles performed by extinct island species that no exotic species could entirely replace. Moreover, exotic species seldom offer complete replacement for extinct biota at the community level. In birds, for example, even if the number of species introduced to an island approximates the number of those that have gone extinct, the ecological traits and phylogenetic diversity are rarely comparable, leading to significantly different functional assemblages (Sobral et al. Reference Sobral, Lees and Cianciaruso2016). This may partly be because exotic species favour non-native habitats (Lee et al. Reference Lee, Wood and Rogers2010). A comparison of broad trophic groups of extinct and extant bird species from New Zealand and the Mascarenes (Fig. 3) shows that invertebrate and seed consumers are better represented currently due to the post-settlement expansion of grasslands (Lee et al. Reference Lee, Wood and Rogers2010). However, patterns for other groups are inconsistent; New Zealand is now depauperate in terrestrial folivores, whereas this guild is enriched in the Mascarenes, and the opposite is true for aquatic plant-feeders, frugivores and vertebrate predators (Fig. 3).

Figure 3 A comparison of broad avifaunal dietary guild changes on Mascarenes and New Zealand.
Removal of exotic species is a key aspect of island conservation (Keitt et al. Reference Keitt, Campbell, Saunders, Clout, Wang, Heinz, Veitch, Clout and Towns2011), but restoring lost ecosystem processes and functions is a further step, requiring knowledge about pre-human states. Fossil records are the key to such information, which can help guide restoration efforts (Barnosky et al. Reference Barnosky, Hadly, Gonzalez, Head, Polly and Lawing2017).
USING THE PAST AS A GUIDE FOR RESTORATION
Fossil deposits record the composition of, and interactions between, species in pre-disturbance ecosystems and can form baselines in order to guide ‘rewilding’ (e.g. Van Leeuwen 2008; Wilmshurst et al. Reference Wilmshurst, Moar, Wood, Bellingham, Findlater, Robinson and Stone2014; Barnosky et al. Reference Barnosky, Hadly, Gonzalez, Head, Polly and Lawing2017). While ‘rewilding’ has been widely discussed for continents, the relatively recent extinctions and smaller scale of islands (Hansen & Galetti Reference Hansen and Galetti2009) make them better candidates (Hansen Reference Hansen2010). Yet, while many islands have had past biotic communities reconstructed from fossil deposits, this understanding has had little influence on management and restoration. The potential for palaeoecology to help us better understand past island ecosystems, and some examples of practical uses, are presented below.
Restoration of island vegetation communities
Palaeoecology shows how island habitats have changed following settlement, identifies plant extinctions and sheds light on their implications. Moreover, palaeoecological records can reveal the suitability of a site for different plant species, thereby increasing the range considered for replanting (Burney & Burney 2015). Limited taxonomic resolution of some palaeoecological proxies means that determining exact species may be problematic (e.g. Chapin et al. Reference Chapin, Wood, Perlman and Maunder2004), but prehistoric baselines are useful at a broad scale. For example, the natural vegetation of Tawhiti Rahi (off northern New Zealand) was thought to be low-statured pohutakawa (Metrosideros excelsa) forest. However, pollen and DNA from a soil core showed that, before settlement, the island was in tall podocarp forest with a palm-rich understorey (Wilmshurst et al. Reference Wilmshurst, Moar, Wood, Bellingham, Findlater, Robinson and Stone2014). On San Cristobal Island (Galápagos), pollen records showed that replanting strategies were based on a heavily modified system rather than the natural ‘pre-grazing’ shrubland (Bush et al. Reference Bush, Restrepo and Collins2014). On the Galápagos, pollen and macrofossils have shown that plants considered to be ‘doubtful natives’ were present before settlement, and their management has been adjusted (Coffey et al. Reference Coffey, Froyd and Willis2011).
An exceptional example is at the Makauwahi Cave Reserve (Kauai), where replanting of 81 plant species (22 absent from the area) has been guided by their presence in late Holocene deposits, using a reference point in the early post-settlement period (Burney & Burney Reference Burney and Burney2016).
Reintroduction of extirpated animal species to islands
A wide range of taxa, including birds, mammals, invertebrates (e.g. Watts et al. Reference Watts, Stringer, Sherley, Gibbs and Green2008), amphibians and reptiles (Dodd and Seigel Reference Dodd and Seigel1991), have been translocated back to islands from where they had been extirpated. For example, since 1863, there have been >1100 attempts to translocate 55 bird species (41 successfully) in New Zealand (Miskelly & Powlesland Reference Miskelly and Powlesland2013). While six taxa have been translocated beyond their ‘natural’ ranges, the remainder represent reintroductions (Miskelly & Powlesland Reference Miskelly and Powlesland2013). Although these translocations were primarily for the conservation of birds (i.e. moving to predator-free sanctuaries), they also restored ecological processes.
There may be a specific case for translocating certain species in order to restore ecosystem processes (e.g. seabirds; Miskelly et al. Reference Miskelly, Taylor, Gummer and Williams2009) or interactions (Seddon et al. Reference Seddon, Griffiths, Soorae and Armstrong2014). For example, analysis of a Holocene coprolite of the New Zealand kakapo (Strigops habroptilus) revealed abundant pollen of the threatened plant Dactylanthus taylorii, suggesting a lost pollination relationship (Wood et al. Reference Wood, Wilmshurst, Holzapfel, Worthy and Cooper2012). This could only have been detected using palaeoecology, as kakapo and Dactylanthus populations no longer overlap naturally. Dactylanthus was assumed to have only been pollinated by a bat, and the rarity of the bat was regarded as a limitation to translocation of the plant (Ecroyd Reference Ecroyd1995).
Restoring ecological function to islands using analogues for extinct species
Translocation of faunal species has been used to ‘rewild’ islands. In the simplest scenario, extant close phylogenetic relatives of the extinct species are used as surrogates. Giant tortoises on Indian Ocean islands performed many important functions, and experiments on Mauritius and Rodrigues using Aldabra giant (Aldabrachelys gigantea) and radiated (Astrochelys radiata) tortoises as surrogates for the extinct endemic species have shown that native seeds passing through tortoises have extremely high germination rates (Griffiths Reference Griffiths and Gerlach2014). Tortoises are also ‘de-weeders’, and their faeces and urine provide vital soil nutrients, making tortoises ideal candidates for rewilding projects on Mauritius and Rodrigues (Griffiths et al. Reference Griffiths, Hansen, Jones, Zuēl and Harris2011; Griffiths Reference Griffiths and Gerlach2014; Hume Reference Hume and Gerlach2014b), Madagascar (Pedrono et al. Reference Pedrono, Griffiths, Clausen, Smith, Griffiths, Wilmé and Burney2013), Galápagos (Gibbs et al. Reference Gibbs, Marquez and Sterling2008) and in the Caribbean (Hansen et al. Reference Hansen, Donlan, Griffiths and Campbell2010). In New Zealand, the extant North Island kokako (Callaeas wilsoni) has been released on Secretary Island in order to restore the ecological functions of the extinct South Island kokako (Callaeas cinerea; Seddon et al. Reference Seddon, Strauss, Innes, Ewen, Armstrong, Parker and Seddon2012). Use of phylogenetic relatives in order to restore extinct plants and birds has also been advocated for Lord Howe Island (Hutton et al. Reference Hutton, Parkes and Sinclair2007).
In more radical scenarios, extant species that are more distantly related to the extinct species might be considered as surrogates. On Kauai, African spurred tortoises (Centrochelys sulcata) benefit native vegetation communities by suppressing undergrowth (presently dominated by invasive species), increasing soil nutrients and aiding seed germination; these are functions once provided by a now-extinct community of large grazing waterfowls (Burney & Burney Reference Burney and Burney2016).
Rewilding may provide improvements to island ecosystems, but it has inherent difficulties and potentially unforeseen consequences (Nogués-Bravo et al. Reference Nogués-Bravo, Simberloff, Rahbek and Sanders2016). Given the risks of introducing species to islands, island rewilding projects should be underpinned by consideration of issues such as the similarity of ecological roles with extinct taxa, potential threats to survival, available stock and rigorous risk/benefit analysis (Louys et al. Reference Louys, Corlett, Price, Hawkins and Piper2014; Seddon et al. Reference Seddon, Griffiths, Soorae and Armstrong2014).
CONCLUSIONS
Islands have a long history of natural extinction from climatic and environmental changes, but extinction rates have increased dramatically through anthropogenic impacts during the Holocene. We have identified some general patterns in terms of the interactions between human settlement phases, exotic species introductions, vegetation clearance and extinction rates (Fig. 1), yet further research (particularly on islands with multiple phases of prehistoric settlement) may help refine the model.
Island extinctions have not yet ceased. The extinction debt (e.g. Triantis et al. Reference Triantis, Borges, Ladle, Hortal, Cardoso and Gaspar2010) and threats to island species (e.g. via continued loss of native vegetation and new invasive species) are ongoing (Manne et al. Reference Manne, Brooks and Pimm1999; Sax & Gaines Reference Sax and Gaines2008). Moreover, anthropogenic climate change may also represent a new driver of island extinctions (Benning et al. Reference Benning, LaPointe, Atkinson and Vitousek2002).
Palaeoecological records provide insights into intact pre-human island ecosystems that are crucial to restoration goals. However, returning island ecosystems to prehistoric states may not always be possible, or even desirable. Humans are now integral parts of island ecosystems, and restoration projects now often combine traditional knowledge and human desires with palaeoecological evidence in order to accommodate cultural, social and economic outcomes, in addition to ecological ones (Lyver et al. Reference Lyver, Wilmshurst, Wood, Jones, Fromont and Bellingham2015; Russell et al. Reference Russel, Meyer, Holmes and Pagadin press). The challenge for the numerous restoration projects that are underway will be to offset biodiversity loss through continued spread of invasive species and climate change (with associated sea-level rise, reduced land area and habitat loss). Developments such as novel pest control systems and de-extinction (Shapiro Reference Shapiro2015; Wood et al. Reference Wood, Perry and Wilmshurst2017), as well as visionary targets such as removing exotic predators from entire archipelagos (Russell et al. Reference Russell, Innes, Brown and Byrom2015), may offer new avenues for slowing extinction rates and restoring island ecosystems.
ACKNOWLEDGEMENTS
We thank M. McGlone and two anonymous referees for comments that greatly improved this manuscript.