


INTRODUCTION
Paleolithic sites located in karstic environments frequently show an occupational alternation between carnivores and hominids, which generate overlapped accumulations that are difficult to interpret archaeologically (Blumenschine Reference Blumenschine1988; Cruz-Uribe Reference Cruz-Uribe1991; Marean and Spencer Reference Marean and Spencer1991; Lam Reference Lam1992; Marean et al. Reference Marean, Spencer, Blumenschine and Capaldo1992). In fact, carnivores could have affected the remains left by the hominids, modifying, destroying, and/or mixing them, and thus erasing significant archaeological information (Binford Reference Binford1981; Camarós et al. Reference Camarós, Cueto, Teira, Tapia, Cubas, Blasco, Rosell and Rivals2013). Therefore, the activities carried out by the carnivores can pose serious problems in attesting the human presence as well as the final interpretation of the site. Their possible coexistence, the use of different spaces for different purposes in the cave, and the chronological attribution of the site strictly assigned to the human presence are still the most challenging questions in this kind of context. Some of these points are already discussed in previous papers (Higham et al. Reference Higham, Basell, Jacobi, Wood, Bronk Ramsey and Conard2012; Hublin et al. Reference Hublin, Talamo, Julien, David, Connet, Bodu, Vandermeersch and Richards2012; Talamo et al. Reference Talamo, Peresani, Romandini, Duches, Jéquier, Nannini, Pastoors, Picin, Vaquero, Weniger and Hublin2014; Wood et al. Reference Wood, Arrizabalaga, Camps, Fallon, Iriarte-Chiapusso, Jones, Maroto, de la Rasilla, Santamaría, Soler, Soler, Villaluenga and Higham2014); therefore, this article presents a hyena den site and the way to deal specifically with this case.
Teixoneres Cave, located in the village of Moià (Barcelona, Spain), is one of the cavities belonging to the karstic system called the Toll Caves (Figure 1). Partial excavation during the 1950s and 1970s revealed an important Holocene sequence and a late Pleistocene paleontological record (Serra-Ràfols et al. Reference Serra-Ràfols JdC, Thomas and Fuste1957; Crusafont Reference Crusafont1960; De Lumley Reference De Lumley1971; Hopf Reference Hopf1971; Guilaine et al. Reference Guilaine, Barbaza, Geddes and Vernet1982). Teixoneres is a U-shaped cave, 30 m long, with three differentiated chambers called X, Y, and Z. The cave has two entrances: the main one is Chamber X and the second one is Chamber Z, which is smaller and was probably opened more recently than the main chamber. Archaeological work in the cave started in the 1950s by a local speleological group. Three deep test drillings in the main chamber (Chamber X) were made at that time, in which some lithic remains and an important Pleistocene faunal assemblage were recovered. Later, in 1973, another small intervention was carried out and focused on the paleontological record (Castellví Reference Castellví1974). These studies affected mainly units I and II, and to a lesser extent the lower units. The cave was then closed until 2003, when new excavations and research projects were opened, and have remained active until today. The current excavations have focused mainly on unit III, since only partial portions of the upper sedimentary units were preserved at the site, especially in the outermost area. These works are applying a methodology based on the excavation in extension. According to Carbonell et al. (Reference Carbonell, Cebrià, Allué, Cáceres, Castro, Díaz, Esteban, Ollé, Pastó, Rodríguez, Rosell, Sala, Vallverdú, Vaquero and Vergés1996), this system comprises the simultaneous excavation of the entire surface of the cave, trying to recover the natural topographic relief of the paleosurfaces and their respective landforms. The main aim of this methodology is to recover the objects in their full context, providing useful information about the associations between different items and their respective taphonomic history. In the case of Teixoneres, this system has been applied successfully following mainly the disposition of the large limestone blocks, which mark the bottom (and the real slope) of the flowstones. In the same way, general changes in the macrocomposition, texture, and color of the sediments are also considered.

Figure 1 Top: location of Teixoneres Cave (Moià, Barcelona) in the northeastern Iberian Peninsula (top); Bottom: general view of Teixoneres Cave during the 2011 excavation season.
The preliminary results of the new excavation, which focused on Chamber X, were published in Rosell et al. (Reference Rosell, Blasco, Rivals, Chacón, Menéndez, Morales, Rodríguez-Hidalgo, Cebrià, Carbonell and Serrat2010), in which Neanderthal activity at the bottom of the sequence (unit III) was identified and carnivore dens were recovered inside the cave. Calcite flowstones were found at the top of the sequence of Chamber X (unit I) and at the medium sequence (unit IV). A series of U-Th dates on the stalagmite of unit IV have placed this unit confidently in the MIS 5c with an average date of 100.3±6.1 ka BP (Tissoux et al. Reference Tissoux, Falguères, Bahain, Rosell, Ardevol, Cebria, Carbonnel and Serrat2006). Less certain are the data for the stalagmite of unit I, which probably corresponds to the MIS 2 between ~14–16 ka BP (Tissoux et al. Reference Tissoux, Falguères, Bahain, Rosell, Ardevol, Cebria, Carbonnel and Serrat2006). The presence of Hystrix sp. and Iberomys cabrerae at the bottom of unit III (subunit IIIb) of Teixoneres Cave allowed for the establishment of a relative chronology of this unit between ~90 to 60 ka BP (López-García et al. Reference López-García, Blain, Burjachs, Ballesteros, Allué, Cuevas-Ruiz, Rivals, Blasco, Morales, Hidalgo, Carbonell, Serrat and Rosell2012).
Complementing the relative microfaunal chronology and the U-Th dates, we decided to attempt radiocarbon dating of subunits II a+b and III a+b. The 14C results surprisingly reveal a robust chronology of the site and all but one of the dates for the two lower units (II and III) are within the 14C time range. Unfortunately, the 14C resolution at the limit of the method is not high enough to provide the answer of who occupied the cave first, the carnivores or the humans, and we cannot establish the temporal contributions of each predator, but we can certainly determine, even if we are dealing with a carnivore den site, a well-defined chronological sequence, through which we can clearly attest the human passage.
THE PRESENT-DAY SITE SITUATION
Stratigraphy
Chamber X of Teixoneres Cave is filled more than 800 cm high by well-stratified sediments. A pit excavated during the 1950s (current squares N-O/14-15) was used by Serra-Ràfols et al. (Reference Serra-Ràfols JdC, Thomas and Fuste1957) and by Serrat and Albert (Reference Serrat and Albert1973) to describe 15 lithostratigraphic units using petrographic and paleontological criteria. During the recent fieldworks, the entire upper part of the cave was excavated in extension, and the stratigraphic profile of reference has been changed to the squares J-K/13-16, where a more general overview of the cave deposition could be observed (Figure 2).
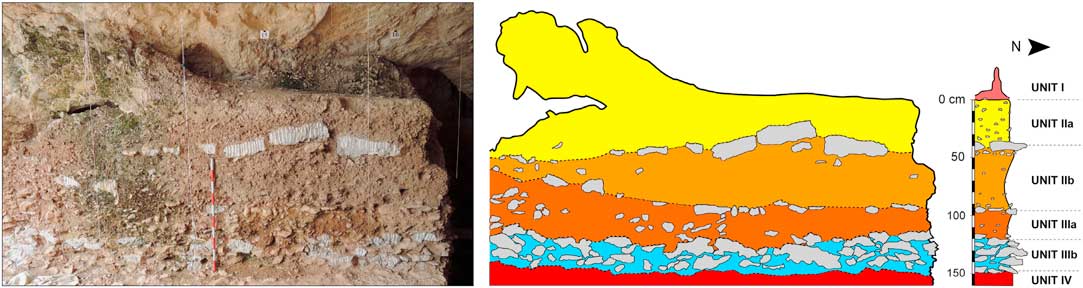
Figure 2 Photograph and stratigraphic profile of the excavation area in Chamber X of Teixoneres Cave.
The stratigraphic sequence of the upper part excavated so far is 160 cm thick and is divided into four lithostratigraphic units separated by the most significant stratigraphic boundaries (I–IV from top to bottom). Units II and III have been subdivided into two subunits following the disposition of the larger limestone blocks and general changes in the color of the sediments. Thus, the upper part of the stratigraphy can be defined as an alternation of autochthonous sediments, decimeter-scale collapsed limestone blocks from the cave roof and flowstones, and allochthonous clay and silt with centimeter-sized limestone clasts derived from the erosion of slope soils. From bottom to top, the main features of the differentiated stratigraphic units are as follows:
-
- Unit IV consists of a flowstone, up to 30 cm thick, which is best developed along the east wall of the cave and its thickness decreases towards the central area. Its surface is undulant with decimeter-wide stalagmites. It has a laminated structure consisting mainly in columnar calcite crystal fabrics. Unit IV also includes discontinuous beds of variable-sized limestone clasts. The main water inflow area is located in the east side of the cave.
-
- Unit IIIb is a homometric monomictic orthobreccia of limestone cobbles and boulders. It is 30 cm thick and is composed of tabular limestone clasts up to 30 cm long and 6 cm thick. The limestone clasts are arranged horizontally, parallel to the cave floor, forming a kind of natural flooring. The pores between limestone blocks are filled by a brown silty clay matrix with a lumpy texture and abundant porosity that includes small (<2 cm) limestone clasts.
-
- Unit IIIa is 27 cm thick and is composed of a homometric monomictic limestone orthobreccia. The clasts are approximately 4 cm in diameter and show rounded edges. The unit shows a coarsening-upward sequence. Towards the top of the sequence, the limestone clasts increase in size and the clayey matrix changes from crumbly to more of a massive texture. The upper contact of the unit is sharp and somewhat irregular.
-
- Unit IIb is 40 to 50 cm thick. It is made up of 2- to 4-cm-long limestone clast homometric monomictic orthobreccia. It shows a fining-upward sequence in the lower half, increasing the amount of silt-clay matrix, and a coarsening-upward sequence in the upper half. A discontinuous horizontal layer of decimeter-scale limestone cobbles and boulders, similar to the clasts from Unit IIIb, defines the top.
-
- Unit IIa is 40 cm thick and is composed of a homometric monomictic limestone clast orthobreccia. The limestone clasts are 2 to 4 cm long and are included in a sparse silty matrix. The top is sharp and is sometimes cemented with spelothemic calcite derived from the formation of Unit I speleothems.
-
- Unit I is made up of many speleothems (flowstone, columns, and stalagmites) developed from different water inflow points, and partially covers the top of the Unit IIa.
Although a distinction among different subunits is described and observed at the stratigraphic profile (Figure 2), this distinction was not always visible in the entire excavated surface. In order to cover several areas of the cave, accelerator mass spectrometry (AMS) 14C dating samples were selected according to their main stratigraphic unit (Units II and III) and their Z value (depth below datum) (Figure 3).

Figure 3 Topography and spatial distribution of archaeological items and 14C samples from Chamber X (Units IIa and IIb) and Chamber Y (Unit 1) of Teixoneres Cave.
Chamber Y is still under excavation; at the moment only Unit 1 is recognized with the presence of one hearth and a few burnt and unburnt bones. The change in the unit numbering (from Roman to Arabic numbers) is due to the fact that at the beginning of the excavation of Chamber Y, in 2012, it was difficult to determine the exact stratigraphic position of its sediments in relation to the sediments coming from Chamber X. It is important to note that this specific area corresponds to a small and residual zone of the inner part of the cave that is almost completely isolated from the rest of the cavity. Unit 1 (and, in general, all the sediments coming from Chamber Y) seems to come from a different sedimentary cone situated at the back of the cave, showing a sedimentary filling process different from the one described in Chamber X. At this stage of the study, we could tentatively suggest that Unit 1 from Chamber Y could be located in a later chronostratigraphic position than Units II and III from Chamber X.
Technological Analysis
The total amount of the lithic assemblage of Teixoneres Cave is 2167 pieces (Unit II = 45 and Unit III=2123). The raw material mostly used is quartz, whereas sedimentary and metamorphic rocks are present in lesser percentages (Tables 1 and 2). The nearby stream of Mal includes cobbles in secondary position of sandstone, quartzite, and quartz. In this latter raw material, the occurrence of fissures made the nodules mediocre for the flake production. Within this local source, more primary and secondary outcrops are located eastwards at about 10–15 km from the site (Mangado and Nadal Reference Mangado and Nadal2001). In this area have been identified different sources of chert, limestone, hornfels, and quartzite. Other outcrops in secondary depositions are found about 8 km south of Teixoneres Cave at San Quirze (quartz and chert) and about 15 km northwest at Can Tripeta (chert). This distribution of the raw material sources point out two main patterns of stone procurement ranging from local to semi-local (Geneste Reference Geneste1988; Turq Reference Turq2000).
Table 1 Total amount and percentages of the lithic assemblage of Unit IIIb (* preliminary amount).

Table 2 Total amount and percentages of the lithic assemblage of Unit IIIa (*preliminary amount).

The analyzed lithic assemblage of Unit III is composed of 20 cores, 269 flakes, 268 fragments, and 44 retouched tools (Tables 1–2; Figure 4). The counting of the quartz assemblage is preliminary because it is still under study, whereas for the other raw materials the whole amount has been analyzed. The technological reading of the lithic collection shows the use of different reduction strategies on the basis of the types of rocks utilized. Quartz nodules and plaquette were mostly exploited opportunistically using the tranche de saucisson method (Turq Reference Turq1989; Bourguignon Reference Bourguignon1996; Hiscock et al. Reference Hiscock, Turq, Faivre and Bourguignon2009; Faivre Reference Faivre2011). The resulting byproducts were thick and short cortical flakes. These blanks are characterized by an abrupt edge that were occasionally retouched and transformed in scrapers, denticulates, and notched tools. The other raw materials were instead transported to the site as isolate blanks or configured cores that were used for short reduction sequences. Limestone, slate, and ludite cores were exploited opportunistically using unidirectional or centripetal methods. The use of Levallois and discoid technology is conversely attested in the chert, quartzite, and hornfels raw materials. Although the operative chains are highly fragmented, with the production of few flakes by cores, in the assemblage are recorded five Levallois recurrent centripetal (four in Subunit IIIb and one in Subunit IIIa) and seven Levallois recurrent unidirectional flakes (five in Subunit IIIb and two in Subunit IIIa), and 12 discoid byproducts (eight in Subunit IIIb and four in Subunit IIIa). Moreover, the analysis documented 43 blanks including pseudo-Levallois points, centripetal and core-edge flakes that technologically might have been produced from both Levallois recurrent centripetal and discoid reduction. In the core assemblage, only one Levallois recurrent centripetal in Unit IIIb and one Levallois preferential artifact in Unit IIIa were recognized showing the general characteristics documented by Boëda (Reference Boëda1993, Reference Boëda1994, Reference Boëda2013). The analysis detected also the recurrent exploitation of core-on-flakes even if the starting blanks are small with limited potential of flake production. Retouched tools comprise mostly scrapers (some of them clear convergent tools) and few denticulates and notched tools. The general interpretation of the lithic assemblage of Unit III confirms the attribution to Neanderthal technical behaviors. Moreover, the technological characteristic of the assemblage and the high fragmentation of the knapping activities (Turq et al. Reference Turq, Roebroeks, Bourguignon and Faivre2013; Moncel et al. Reference Moncel, Chacón, La Porta, Fernandes, Hardy and Gallotti2014) corroborate the short-term occupation of the site. The spatial distribution of the lithic finds show that the knapping activities were mostly carried out in the front and in the area near the drip line of the cave. Further studies on lithic refitting and the spatial pattern analysis (currently under study) will disclose the location of the different activity areas and their relation with the hearths and the subsistence strategies developed at the site.

Figure 4 Examples of lithic artifacts from Unit III: (A) chert flakes, (B) cores on quartzite and chert, (C) retouched tools on chert and quartzite, (D) cores on quartz, (E) flakes and retouched tools on quartz.
The analysis of the lithic assemblage of Unit II has been biased by the loss of the finds previously recovered during the excavations carried out between 1950 and 1970. The current collection has been uncovered from the few portions of Unit II left as testimony in the western part of cave and only 44 lithic items and 1 core were discovered (Table 3). The technological analysis documented a large amount of small fragments and knapping chips but few complete flakes (Table 3). The chert collection includes one pseudo-Levallois point and one natural core-edge flake, probably detached from the same nodule, one centripetal flake, and blanks of preparation of the flaking surface and the striking platform. Conversely, the quartz assemblage includes one cortical and two ordinary flakes. The single crystal blank was broken by a siret knapping accident. Within the limestone artifacts are recorded one core with the opportunistic detachment of a refitted small flake. Although the chert assemblage might be interpreted as a result of discoid exploitation and the variety of the raw materials used resemble the patterns of transport evidenced in the Mousterian Unit III, the scant technological information available and the lack of diagnostic pieces impedes the secure attribution of the lithic production to Neanderthals or Anatomically modern human (AMH). In this scenario, the lithic assemblage of Unit II may attest only the human presence without any further interpretation.
Table 3 Total amounts of the lithic assemblage of Unit II.

Faunal Remains
The faunal record from Teixoneres Cave shows a high diversity of taxa (Table 4). This diversity is concordant with the paleoecological reconstruction obtained from the combination of microvertebrate and paleoflora data (López-García et al. Reference López-García, Blain, Burjachs, Ballesteros, Allué, Cuevas-Ruiz, Rivals, Blasco, Morales, Hidalgo, Carbonell, Serrat and Rosell2012), which presents a mosaic landscape dominated by a wet forest and meadows. Leporids (especially Oryctolagus cuniculus), horses (Equus ferus), red deer (Cervus elaphus), and large bovines (Bos/Bison) are the most represented specimens in terms of number of identified specimens (NISP), followed by roe deer (Capreolus capreolus) and wild boar (Sus scrofa). Carnivores are also significant in the cave, specifically cave bears (Ursus spelaeus) and spotted hyenas (Crocuta crocuta). Other carnivores present are wolves (Canis lupus), foxes (Vulpes vulpes), lynx (Lynx sp.), and badgers (Meles meles).
Table 4 NISP of macromammals by units from Teixoneres Cave (MNI = minimum number of individuals; NISP = number of identified specimens).

From a taphonomic point of view, a significant proportion of the ungulate bones show tooth marks and fractures generated by these carnivores (Table 5). Occasionally, these modifications are associated with coprolites (n = 48). On the other hand, the faunal record associated with these human occupations can be identified by the presence of cut-marks, anthropogenic fractures, and burning damage (Figure 5).

Figure 5 Examples of cut-marked bones from Teixoneres Cave: (A) flat bone of a large-sized ungulate; (B) tibia of cervid.
Table 5 Modifications generated on ungulate bones by hominins and carnivores from Units II and III.

The taphonomical studies suggest that Teixoneres Cave was a carnivore den, which could have been used during the winter by bears for hibernation and by other carnivores during the respective breeding season. In this respect, tooth-marked and fractured bones correspond to the portions of hunted/scavenged prey around the cave. However, this natural dynamic was altered occasionally by some human groups that visited the cave during their movements around the territory (Rosell et al. Reference Rosell, Blasco, Rivals, Chacón, Menéndez, Morales, Rodríguez-Hidalgo, Cebrià, Carbonell and Serrat2010; Sánchez-Hernández et al. Reference Sánchez-Hernández, Rivals, Blasco and Rosell2014). This dichotomy in the occupation of the cave can be solved archaeologically by the spatial distribution of the record. While the main carnivore activities seem to have occurred in the inner sectors of the cave, the human occupations are clustered in the main entrance of Chamber X, at least in the case of Unit III (Rosell et al. Reference Rosell, Blasco, Rivals, Chacón, Menéndez, Morales, Rodríguez-Hidalgo, Cebrià, Carbonell and Serrat2010).
RADIOCARBON APPROACH AT TEIXONERES
Over the past several years, there has been substantial progress in the 14C methodology in dating Paleolithic bones. It has been convincingly shown that bone collagen extraction generally requires ultrafiltration (Brown et al. Reference Brown, Nelson, Vogel and Southon1988; Brock et al. Reference Brock, Bronk Ramsey and Higham2007; Talamo and Richards Reference Talamo and Richards2011) and careful cleaning of the filter (Bronk Ramsey et al. Reference Bronk Ramsey, Higham, Bowles and Hedges2004). It has also been demonstrated that the standard pretreatment procedures for charcoal (acid-base-acid; ABA) may not remove young contamination in old charcoal sufficiently, and a subsequent step of oxidation and stepped combustion (ABOX-SC) results in older ages compared to ABA (Wood et al. Reference Wood, Douka, Boscato, Haesaerts, Sinitsyn and Higham2012). On the other hand, some scholars find no significant difference between the ABA and the new ABOX-SC procedures (Cuzange et al. Reference Cuzange, Delque-Kolic, Slar, Grootes, Higham, Kaltnecker, Nadeau, Erlin, Paterne, van der Plicht, Bron, Valladas, Tes and Geneste2007; Haesaerts et al. Reference Haesaerts, Damblon, Nigst and Hublin2013).
To obtain reliable chronologies attesting human occupation in a carnivore den site, there could be other problems, which are not simply related to the pretreatment procedures. For this reason, the sampling strategy is crucial and must be planned in detail. This involves an accurate analysis of the site in question and most importantly an intense collaboration between the archaeologists involved in the study of the site and the 14C specialists.
The first aspect to consider in the process of selecting samples for 14C dating is to prefer bones that display a clear association with human activities. This association can be inferred from the presence of cut-marks and anthropogenic fractures on bones (human-modified bones) and by the spatial location of charcoals in the hearths. However, this type of selection process, for bone samples, can lead to some biases related to the preservation of collagen (Hublin et al. Reference Hublin, Talamo, Julien, David, Connet, Bodu, Vandermeersch and Richards2012). For this reason, the number of samples taken for 14C dating will be the second most important factor because it will help to constrain the chronological events at the site, which is a decisive aspect in a carnivore den site.
The spatial distribution of the bone remains is another fundamental aspect to be contemplated, in order to investigate the contribution of each predator in the site. Once the living floor of the two groups has been attested (for Teixoneres, carnivores occupied the inner part and humans the front space), samples can be selected for 14C dating in the two different areas.
At Teixoneres, 16 samples of animal bones (7 human modified, 9 unmodified) and one charcoal were selected for 14C dating from two archaeological units (14 samples from Units IIa+b and IIIa+b) in Chamber X and three samples from Unit 1 in Chamber Y (Figures 3 and 5). Knowing the carnivores’ role at the site, the second step is to select samples that are internal (Chamber Y) versus external (Chamber X) to the cave, since the human occupation found in Unit III is most pronounced in the entrance (Rosell et al. Reference Rosell, Blasco, Rivals, Chacón, Menéndez, Morales, Rodríguez-Hidalgo, Cebrià, Carbonell and Serrat2010). In order to attest the carnivore activities in the inner sectors of the cave, we selected three samples from Unit 1 (one charcoal from a hearth, one ungulate bone, and one hyena bone) in Chamber Y. The reason for selecting the hyena bone is that we want to confirm if there are any differences in terms of ages between the human occupations of the site, using the hearth as reference, and the hyena time period. Seven samples from Unit II were selected from the center of the cave and seven samples from Unit III, most of them from the entrance of the cave.
Bone samples were pretreated at the Department of Human Evolution at the Max Planck Institute for Evolutionary Anthropology (MPI-EVA), Leipzig, Germany, using the method described in Talamo and Richards (Reference Talamo and Richards2011): the outer surface of the bone samples is first cleaned by a shot blaster and then 500 mg of bone is taken. The samples are then decalcified in 0.5M HCl at room temperature until no CO2 effervescence is observed, usually for about 4 hr. Then, 0.1M NaOH is added for 30 min to remove humics. The NaOH step is followed by a final 0.5M HCl step for 15 min. The resulting solid is gelatinized following Longin (Reference Longin1971) at pH 3 in a heater block at 75°C for 20 hr. The gelatin is filtered in an Eeze-Filter™ (Elkay Laboratory Products, UK) to remove small (<80 μm) particles. The gelatin is then ultrafiltered (Brown et al. Reference Brown, Nelson, Vogel and Southon1988) with Sartorius “VivaspinTurbo” 30kDa ultrafilters. Prior to use, the filter is cleaned to remove carbon-containing humectants (Brock et al. Reference Brock, Bronk Ramsey and Higham2007). The samples are lyophilized for 48 hr. The charcoal sample was sent directly to the Klaus-Tschira-AMS facility of the Curt-Engelhorn Centre in Mannheim, Germany, where it was pretreated with the ABOX method (ABA followed by immersion in K2Cr2O7 in H2SO4 at 60°C). All dates were corrected for a residual preparation background estimated from pretreated 14C-free bone samples, kindly provided by the Mannheim laboratory and pretreated in the same way as the archaeological samples.
To identify the preservation of the collagen C:N ratios, %C, %N, collagen yield, and δ13C and δ15N values must be evaluated. The C:N ratio should be between 2.9 and 3.6 and the collagen yield no less than 1% of the weight (Ambrose Reference Ambrose1990; van Klinken Reference van Klinken1999). Stable isotopic analysis is evaluated at MPI-EVA, Leipzig (lab code S-EVA), using a ThermoFinnigan Flash EA coupled to a Delta V isotope ratio mass spectrometer. Once these fundamental criteria are estimated, the collagen extract, between 3 and 5 mg, is weighed into precleaned tin capsules and sent to the Mannheim AMS laboratory (lab code MAMS), where they were graphitized and dated (Kromer et al. Reference Kromer, Lindauer, Synal and Wacker2013).
RESULTS
For Teixoneres, the isotopic results and the C:N ratios fully satisfy the acceptable range. Only one of the 16 samples displayed a slightly under 1% collagen yield; however, it displayed a normal C:N ratio. No differences are observed between the human-modified versus non-human-modified bones in terms of collagen preservation (Table 6).
Table 6 Isotopic values, C:N ratios, and amount of collagen extracted (%coll) refer to the >30kDa fraction. The results of AMS 14C dating of 17 samples from Teixoneres Cave from Unit I to Unit III. δ13C values are reported relative to the VPDB standard and δ15N values are reported relative to the AIR standard. The human-modified bones are indicated by asterisks (*) following the MPI lab code.

14 C Results
The 14C results of Chamber X (Units II and III) surprisingly (considering the carnivore presence) are all in agreement with the stratigraphic order attesting the human presence at the site (Table 6). The uncalibrated 14C dates of Unit III range from >51,000 to 40,610 14C BP. The seven dates from Unit II range from 40,800 to 30,780 14C BP.
On the other hand, the 14C results of Chamber Y (Unit 1) displayed the already suspected role of the carnivores, showing three different situations: one date at 43,100±400 14C BP (MAMS-17595) which is the hyena bone, one charcoal from a hearth at 28,390±80 14C BP (MAMS-19078), and the youngest date on aunmodified bone is at 10,343±29 14C BP (MAMS-17597).
DISCUSSION
The Teixoneres Cave 14C dates were calibrated using OxCal v 4.2 (Bronk Ramsey Reference Bronk Ramsey2009; Bronk Ramsey and Lee Reference Bronk Ramsey and Lee2013) and IntCal13 (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013) (Figure 6, Table 7).

Figure 6 Bayesian model of dates from Chamber X at Teixoneres. Radiocarbon dates are calibrated in IntCal13 (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013); the model and boundaries were calculated using OxCal v 4.2 (Bronk Ramsey and Lee Reference Bronk Ramsey and Lee2013) including the performance of the General t-type Outlier Model (Bronk Ramsey Reference Bronk Ramsey2009). Human-modified bones are shown in red while non-human-modified bones are given in black. An Osmond-type isochron for the U-Th results was created based on the data published in Tissoux et al. (Reference Tissoux, Falguères, Bahain, Rosell, Ardevol, Cebria, Carbonnel and Serrat2006). The uncertainty of 230Th/232Th ratio should be assumed. Uncertainties may be different when using original data. The results are linked with the (NGRIP) δ18O climate record.
Table 7 Calibrated boundaries at Teixoneres Cave, provided by OxCal v 4.2 (Bronk Ramsey Reference Bronk Ramsey2009; Bronk Ramsey and Lee Reference Bronk Ramsey and Lee2013) using the IntCal13 calibration curve (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013).

The combination of the absolute dating evidence and the relative stratigraphic sequence from the archeological site in Bayesian models provide the basis to build reliable chronologies. The Bayesian model was performed only for Chamber X. This is a sequence of two contiguous phases (Units III and II), considering that the units are in direct contact with each other and no break or discontinuities between units are observed. An Osmond-type isochron for the U-Th dates published in Tissoux et al. (Reference Tissoux, Falguères, Bahain, Rosell, Ardevol, Cebria, Carbonnel and Serrat2006) was created and added to the model in order to show the upper Unit I, which is at the top of the sequence.
For Chamber Y, we limited the discussion on the calibrated ranges without modeling them. The reason to create the Bayesian model only for Chamber X and not for Y is primarily due to the fact that a different sedimentary filling process in the two chambers was determined and secondly because only in Chamber X is the human presence fully attested.
In the Bayesian model, bone samples are colored to distinguish the human-modified samples in red and those without human modifications in black (Figure 6). The t-type outlier analysis, performed to detect problematic samples with prior probabilities set at 5%, confirms the integrity of the chronology detecting only 1 outlier in 13 samples. The sample MAMS-18671 (47,200 ± 670 14C BP) was not computed in the overall analysis because it is outside the range of IntCal13. In this case, the lower calibrated range for this sample is shown in the graph, but it is not modeled and the date is followed by a question mark in Figure 6. The overall agreement index is 85.6%, well above the minimal acceptable level of 60%. A calibrated start boundary for the lower part of the sequence (Unit III) at Teixoneres cannot be defined because the bottom of unit III is older than 51,000 14C BP.
A general observation is that, even in a carnivore den site, a chronology can be well defined. Good agreement between the 14C dates and the stratigraphic layer can be achieved if the sampling strategy is well planned. Moreover, it is shown that all the human-modified bone samples are in the same age range compared to the unmodified samples, confirming once more the importance of the coordination between the taphonomic studies and the sample selection for 14C dating. However, given the 14C resolution for this time period, it is impossible to resolve the temporal sequence between the presence of the two main actors, humans and carnivores, at the site.
Focusing on the lower Unit III in Chamber X, Neanderthals, which are the hominids that occupied the site based on the lithic evidence (Mousterian), inhabited the cave until 44,210 cal BP (68.2%). For the overlying Unit II, no diagnostic lithic pieces firmly assigned to Neanderthals or AMH have been found; therefore, we cannot attribute this unit to a hominid species, but based on human-modified samples we can confirm that humans were present at Teixoneres between 44,210 to 33,060 cal BP (68.2%). The youngest date (MAMS-18670) ranging from 34,870 to 34,580 cal BP (68.2%) in the model is flagged as an outlier from the Bayesian model, with 8% posterior probability. This sample, found at the top of Unit II, could belong to a final phase of the stratigraphic sequence, chronologically very close to Unit 1 in Chamber Y, due most likely to the rapid sedimentation. The explicit carnivore’s work is documented in Chamber Y, where three different ages (non-overlapping at 2σ) are determined in one unit (Unit 1). The hyena bone from this unit ranges between 46,690 and 45,830 cal BP (68.2%). This result suggests that the hyena occupied the Chamber Y (inner part of the cave) while Neanderthals occupied Unit III in Chamber X (external part of the cave). Given the 14C resolution at this time period, it is impossible to attest the interaction between them. In the same unit, we also have evidence of human activity due to the hearth found inside the cave between 32,580 to 32,050 cal BP (68.2%). The last sample elected from Unit 1 produced the youngest date (MAMS-17597), ranging from 12,380 to 12,060 cal BP (68.2%), which overlaps with the U-Th result on the stalagmite crust in Chamber X (14,000+4000–3900 yr BP Osmond-type isochron). All of the results in Chamber Y point towards the sedimentological attribution as proposed, wherein Unit 1 seems to correspond to the absent sediment under the stalagmite crust (Unit I) in Chamber X, due to the chronological range we obtained. This range confirms the U-Th date on the stalagmite crust and the fact that it is more recent than Unit II when correlated with Chamber X.
CONCLUSION
Sites occupied by both humans and carnivores should be treated with extreme caution when constructing a chronology. Just a single passage of carnivores in the site can disrupt the chronology by moving objects within the units, resulting in 14C ages incompatible with the stratigraphy (e.g. in Chamber Y).
With this study, we demonstrate that it is possible to overcome these problems using a careful strategy. An accurate sample selection based on taphonomic studies, a meticulous introspective of the site, the spatial distribution of the archaeological remains, as well as the close collaboration and communication between the archaeologists and the 14C specialist strongly contributed to build a reliable chronology for the site of Teixoneres.
Teixoneres, which is seen as a stop along the way for the hominids during the Late Middle Paleolithic (OIS 3), located in the northeastern part of the Iberian Peninsula and also a carnivore den site (Rosell et al. Reference Rosell, Blasco, Rivals, Chacón, Menéndez, Morales, Rodríguez-Hidalgo, Cebrià, Carbonell and Serrat2010), finally reveals a minimum of 12,000 yr (68.2% confidence) of human presence. The absence of diagnostic lithic industries in Unit II does not allow us to distinguish between Neanderthals or AMH but the human-modified bones dated attest the human occupation at this unit from 44,210 to 33,060 cal BP at 68.2%.
The 14C results of the samples of Unit III place the Neanderthal occupation of Teixoneres Cave in the chronological range of the late Middle Paleolithic (Higham et al. Reference Higham, Douka, Wood, Bronk Ramsey, Brock, Basell, Camps, Arrizabalaga, Baena, Barroso-Ruiz, Bergman, Boitard, Boscato, Caparros, Conard, Draily, Froment, Galvan, Gambassini, Garcia-Moreno, Grimaldi, Haesaerts, Holt, Iriarte-Chiapusso, Jelinek, Jorda Pardo, Maillo-Fernandez, Marom, Maroto, Menendez, Metz, Morin, Moroni, Negrino, Panagopoulou, Peresani, Pirson, de la Rasilla, Riel-Salvatore, Ronchitelli, Santamaria, Semal, Slimak, Soler, Soler, Villaluenga, Pinhasi and Jacobi2014). The spatial distribution of the carnivore bones in the cave shows that they tended to occupy the interior part of the cave (Rosell et al. Reference Rosell, Blasco, Rivals, Chacón, Menéndez, Morales, Rodríguez-Hidalgo, Cebrià, Carbonell and Serrat2010); this situation is confirmed by 14C results in Unit 1 from Chamber Y. Moreover, the 14C dates produced at Teixoneres are well defined within the stratigraphic context, allowing to definitely attest the human presence from beyond 51,000 14C BP to 32,050 cal BP (68.2%). These results may thus be included in the extensive 14C database in the northeastern Iberian Peninsula, and they could be used for a broad comparison with other Mousterian sites, in a geographic area that could have been a way of communication among the main valleys of the central Catalonia in the northeast of the Iberia Peninsula.
ACKNOWLEDGMENTS
This project was funded by the Max Planck Society. The archaeological research was supported by funding from the Spanish Ministry of Science and Innovation-FEDER, project nos. CGL2012-38434-C03-03, CGL2012-38358, HAR2010-19957, HAR2013-48784-C3-1-P, and CGL-BOS-2012-34717, from Generalitat de Catalunya 2014/100573 and 2014 SGR 900, and the SéNeCa Foundation project 19434/PI/14. R Blasco and J M López-García are Beatriu de Pinós post-doctoral research fellowship recipients (Generalitat de Catalunya and COFUND Marie Curie Actions, EU-FP7). Anna Rufà is the beneficiary of a predoctoral research fellowship FPU from the Ministry of Education, Culture and Sports, Spain. E Camarós is the beneficiary of FI Grant from Generalitat de Catalunya and financed by the European Social Fund. Carlos Sánchez-Hernández is the beneficiary of a predoctoral research fellowship FPI from the Ministry of Economy and Competitiveness, Spain. A Picin is the beneficiary of a Humboldt Postdoctoral Research Fellowship from the Alexander von Humboldt Foundation. We thank Dirk Hoffmann for advice on U-Th dating and the Osmond-type isochron calculation. We are indebted to Lysann Rädisch and Sven Steinbrenner of the Department of Human Evolution at the Max Planck Institute for Evolutionary Anthropology for technical assistance. Thanks are also due to the Editor and to two anonymous referees for their comments and suggestions that have contributed to improve this research paper.













