Introduction
Superparasitism is common in parasitoids, long considered as a mistake made by imperfect parasitoids but now generally recognized as adaptive in a number of situations (van Alphen & Visser, Reference van Alphen and Visser1990). For solitary parasitoids, superparasitism is easily determined by dissection of hosts to examine the number of eggs deposited in a host, since only one offspring can survive the larval stage. For gregarious parasitoids, however, it is not as easily determined by host dissection, since more than one offspring can survive the larval stage. Therefore, the study and the understanding of superparasitism in gregarious parasitoids present a particular challenge (Dorn & Beckage, Reference Dorn and Beckage2007).
A gregarious parasitoid often has two successive decisions to make on oviposition in response to a parasitized host. First, the parasitoid should decide if the host is acceptable for oviposition. This decision depends on its ability to distinguish between unparasitized and previously parasitized hosts, an ability called ‘host discrimination’ (Godfray, Reference Godfray1994). Most parasitoids studied have shown this ability (Ueno & Tanaka, Reference Ueno and Tanaka1994; Gauthier et al., Reference Gauthier, Monge and Huignard1996), but there are exceptions. For example, a few studies failed to find evidence for host discrimination by Cotesia glomerata (Hymenoptera: Braconidae), a gregarious endoparasitoid of Pieris spp. (Lepidoptera: Pieridae) (Masurier, Reference Masurier1991; Gu et al., Reference Gu, Wang and Dorn2003). Yet, host discrimination can be influenced by a variety of factors, such as the previous experience of oviposition, the time interval between two attacks on the host (van Alphen & Visser, Reference van Alphen and Visser1990; Godfray, Reference Godfray1994; Yazdani et al., Reference Yazdani, Glatz and Keller2015), and the kind of superparasitism (self vs. conspecific superparasitism) (Dijken et al., Reference Dijken, van Stratum and van Alphen1992; King, Reference King1992; Visser, Reference Visser1993). For example, numerous studies indicate that naive and oviposition-experienced parasitoids have different responses in the acceptance of parasitized hosts for oviposition (Dijken et al., Reference Dijken, van Stratum and van Alphen1992; King, Reference King1992; Godfray, Reference Godfray1994; Ito & Yamada, Reference Ito and Yamada2014). Second, once accepting a previously parasitized host for oviposition, a gregarious parasitoid should decide on clutch size and sex allocation. According to the local mate competition theory (Hamilton, Reference Hamilton1967; Suzuki & Iwasa, Reference Suzuki and Iwasa1980; Parker & Courtney, Reference Parker and Courtney1984), when encountering a previously parasitized host, a gregarious parasitoid should decrease her clutch size but increase the male proportion of her progeny. This theoretical model has gained broad support from empirical studies of a wide variety of organisms (Wylie, Reference Wylie1965; Werren, Reference Werren1980, Reference Werren1984; Ikawa & Suzuki, Reference Ikawa and Suzuki1982; Vet et al., Reference Vet, Datema, Janssen and Snellen1994; Shuker & West, Reference Shuker and West2004; West, Reference West2009). There are, however, a few exceptions in gregarious parasitoids. For example, similar clutches in size and sex ratio were laid by Trichogramma evanescens in parasitized and unparasitized host eggs (Suzuki et al., Reference Suzuki, Tsuji and Sasakawa1984; Dijken & Waage, Reference Dijken and Waage1987); clutches even contrary to the theoretical predictions were laid by Nasonia vitripennis and Bracon hebetor in superparasitism (King, Reference King, Wrensch and Ebert1993). In addition, other factors may also influence clutch decisions made by superparasitizing parasitoids. For example, a parasitoid may adjust her clutch decision in response to a host that was previously attacked by either herself or a conspecific female, resulting in self- or conspecific superparasitism. These two kinds of superparasitism can have different adaptive consequences for the ovipositing solitary parasitoid, by inducing a competition among either sibling or non-sibling offspring parasitoids in the host (van Alphen & Visser, Reference van Alphen and Visser1990). Therefore, to understand superparasitism as an adaptive oviposition strategy in gregarious parasitoids, more studies are needed.
Oomyzus sokolowskii (Kurdjumov) (Hymenoptera: Eulophidae) is a larval–pupal endoparasitoid, exclusively attacking the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae) (Talekar & Hu, Reference Talekar and Hu1996; Sarfraz et al., Reference Sarfraz, Keddie and Dosdall2005), which is one of the major herbivorous pests on cruciferous crops (Talekar & Shelton, Reference Talekar and Shelton1993; Furlong & Zalucki, Reference Furlong and Zalucki2007; Zalucki et al., Reference Zalucki, Shabbir, Silva, Adamson, Liu and Furlong2012; Furlong et al., Reference Furlong, Wright and Dosdall2013). The parasitoid prefers host larvae of later instars to the earlier (Mushtaque, Reference Mushtaque1990; Talekar & Hu, Reference Talekar and Hu1996; Nakamura & Noda, Reference Nakamura and Noda2001; Sow et al., Reference Sow, Arvanitakis, Niassy, Diarra and Bordat2013), laying a clutch of 10–20 eggs with often an female-biased sex ratio (Wang et al., Reference Wang, Liu, Guo and Lin1999; Nakamura & Noda, Reference Nakamura and Noda2002; Li et al., Reference Li, Zhu, Meng and Li2017). Superparasitism occurs in the field (Mushtaque, Reference Mushtaque1990), and yields more offspring parasitoids with smaller body sizes yet greater longevity under the provision of honey food, but offspring sex ratio does not change with times of parasitization (Silva-Torres et al., Reference Silva-Torres, Filho, Torres and Barros2009). It is not yet known whether O. sokolowskii females are able to discriminate against parasitized hosts and how they adjust their progeny allocation to them due to some circumstances.
In this study, we made a series of single-factor-designed experiments using O. sokolowskii – P. xylostella as a gregarious parasitoid–host system to address two questions. First, do female parasitoids discriminate between unparasitized and parasitized host larvae? To answer this question, we first examined their preference in the willingness to attack between the two types of hosts and their progeny allocation to them by using irradiation-sterilized parasitoids. Second, do parasitoids adjust their progeny allocation in superparasitism in response to the time interval between two bouts of oviposition on the host, to their own experience of oviposition on healthy hosts, or to whoever made the first attack? To answer this question, we compared brood size and sex ratio of progeny emerged from superparasitized hosts receiving different treatments. The answer to these questions will deepen our understanding of superparasitism as an adaptive reproductive strategy for gregarious parasitoids.
Materials and methods
Insect preparation
The host diamondback moth P. xylostella was collected in the summer of 2013 from a pakchoi (Brassica rapa subsp. chinensis L.) field, Nanjing city, Jiangsu province, eastern China and thereafter maintained on potted pakchoi seedlings in an insectary at 25 ± 1°C, 60 ± 5% RH, and a photoperiod of 16:8 h (L:D). The parasitoid O. sokolowskii was obtained in the year of 2013 from the Key Laboratory of Northwest Loess Plateau Crop Pest Management of Ministry of Agriculture of China, Northwest A&F University, Yangling, Shaanxi, China, and then maintained with P. xylostella as the host in an insectary. Parasitism was made in a 4 ml vial, where four final instar larval hosts were exposed to parasitism by a female parasitoid for 24 h. Then the host larvae were individually reared in vials where fresh pakchoi leaf discs (0.5 cm diameter) were provided as food ad libitum and replaced daily until pupation. The pupae were collected in groups in vials for the emergence of offspring parasitoids, which were allowed to mate. Adult female parasitoids of 4–6 days old were used in all experiments.
To obtain sterilized parasitoids (producing infertile eggs), mated females were irradiated using 40 Gy 60Co at a dose rate of 2 Gy min−1 (processed at the Atomic Energy Department of Jiangsu Academy of Agriculture Sciences, Nanjing). Our exploratory trial showed that such treatment can make O. sokolowskii females (n = 30) oviposit infertile eggs on P. xylostella fourth-instar larvae. The irradiated parasitoids were used in the experiment within 2 days after the treatment.
Host discrimination
Two experiments examined host discrimination of female parasitoids between unparasitized and parasitized hosts, measuring their attacking preference in a dual-choice trial and comparing their progeny allocation to the two types of hosts in a non-choice trial. In the dual-choice trial, a pair of unparasitized and just parasitized fourth-instar larval hosts were exposed to an attack by a female parasitoid. The two larval hosts for the choice were similar in body size yet distinguishable from each other by one of them (at random) being marked on the dorsal thorax by a dark ink spot; the marking had negligible impact on the behaviors of both hosts and parasitoids as shown in our exploratory trial. A naive female parasitoid (without the experience of oviposition) was introduced into a petri-dish (5 cm diameter × 1 cm height) containing the host pair and she was observed continuously until making an attack on one of them. An attack was defined as a bout of oviposition lasting more than 120 s, by which the host had 97.6% probability (n = 30) to be oviposited as shown in our exploratory observation. The trial was replicated for 80 female parasitoids.
The second experiment compared progeny allocation of parasitoids attacking an unparasitized host and one previously parasitized by a sterilized parasitoid, which had been irradiated with 60Co to produce infertile eggs in attacking the host. A fourth-instar larval host was introduced into a vial (1.5 cm diameter × 5 cm height) and an irradiated parasitoid was then released. Once having made an attack, the parasitoid was removed and another healthy parasitoid was immediately released to make the next attack on the host. This trial was replicated for 36 pairs of parasitoids. As the control, a fourth-instar larval host was exposed in a vial to a single attack by a healthy parasitoid; the trial was replicated for 30 parasitoids. The host larva obtained above was then removed and reared in a petri-dish on food plant discs (0.05 cm diameter) supplied ad libitum and replaced daily until the larva pupated. The pupa was placed in a vial for the emergence of parasitoid offspring. The adult parasitoids emerged from a single host were counted by sex to obtain brood size and sex ratio; sex was determined by the antennae: male with long hairs on each segments and female without the hairs.
Comparison of progeny production between superparasitism with and without intervals since a previous attack
This experiment compared progeny production between hosts superparasitized after two periods from a previous attack. The superparasitism was made in vials using P. xylostella fourth-instar larvae and female parasitoids without the experience of oviposition. A host larva was exposed to two consecutive attacks at either a 0 or 48 h (±5 min) interval by two different parasitoids, to which were referred as 0 or 48 h-elapsed superparasitism treatment. The host was then removed and individually reared in vials on the food plant leaf discs for the emergence of offspring parasitoids. The adult parasitoids at emergence were recorded as in the above experiment. Each treatment was replicated 30 times.
Comparison of progeny production between self- and conspecific superparasitism
This experiment compared progeny production from self-superparasitized hosts with that from conspecifically superparasitized ones. A fourth-instar larval host was introduced into a vial, then either one naïve parasitoid was released and observed to make two attacks, creating the self-superparasitism treatment, or two parasitoids were consecutively released with each making a single attack (the second one was made using the parasitoid with the experience of oviposition to be comparable to self-superparasitism), creating the conspecific superparasitism treatment. The host was then removed and individually reared on the food plant leaf discs for the emergence of offspring parasitoids. The adult parasitoids at emergence were recorded as in the above experiment. The self-superparasitism treatment was replicated 25 times and the conspecific superparasitism one 28 times.
Comparison of progeny production between superparasitism by parasitoids with and without oviposition experience
This experiment compared progeny production from the host superparasitized by the parasitoid with the experience of oviposition and one without it. A P. xylostella fourth-instar larva was introduced into a vial and then a naive parasitoid was released. Once the parasitoid finished an attack on the host and then removed, another parasitoid with or without the experience of oviposition was released to make the next attack. The superparasitized host was then removed and reared as in previous experiments. A total of 28 superparasitizing parasitoids with the experience of oviposition were tested and 77 ones without it.
Data analysis
The non-parametric Sign Test was used to examine the difference between choices for unparasitized and parasitized hosts in the dual-choice trial. Generalized linear model (GLM) was used to determine the effect of respective factors under the examination on brood size (Poisson distribution, link = log) or sex ratio (binomial distribution, link = logit). Overdispersion was corrected by multiplying the standard errors of all the coefficient estimates by the square root of the estimated overdispersion (Gelman & Hill, Reference Gelman and Hill2007). Likelihood ratio test was used to determine the influence of the individual predictor variables in the GLM. The significance level used in all analyses was 5%. Data analyses were performed using R statistical program (R Development Core Team, 2014).
Results
Host discrimination
Oomyzus sokolowskii females made 55 attacks on unparasitized hosts out of 80 dual-choice trials, showing a significant preference for unparasitoized host larvae to parasitized ones (P = 0.001), with 68.8% probability (95% CI 57.4–78.7%) to attack the former when given a choice.
Brood size significantly differed between parasitoids attacking unparasitized hosts and parasitized ones (χ2 = 22.72, P < 0.001). It decreased to an average of 7.1 by a factor of 0.74 (95% CI 0.65–0.83) for the parasitoid attacking a parasitized host compared with one attacking an unparasitized one (fig. 1a). Sex ratio (male proportion of progeny) significantly differed between parasitoids attacking unparasitized hosts and those attacking parasitized ones (χ2 = 150.18, P < 0.001). The odds of producing male offspring to a parasitized host was 6.74 times greater than that to an unparasitized one (fig. 1b).

Fig. 1. Brood size (a) and male proportion (b) of progeny allocated to a Plutella xylostella larva that was previously parasitized by a sterilized parasitoid (superparasitism) and to one that was not parasitized (single parasitism) in Oomyzus sokolowskii. Bars are mean ± 1 standard error.
Comparison of progeny production between superparasitism with and without intervals since a previous attack
Brood size differed significantly between 0 and 48 h-elapsed superparasitism treatments (χ2 = 4.97, P = 0.03). Brood size from the 48 h-elapsed superparasitism treatment (on average 10.8) decreased by 20% (95% CI 3–34) compared with that from the 0 h-elapsed treatment (fig. 2a). Sex ratio differed significantly between the two time intervals treatments (χ2 = 12.28, P < 0.001), with the odds of producing male offspring from the 48 h-elapsed superparasitism decreased by a factor of 0.46 (95% CI 0.29–0.72) of that from the 0 h-elapsed one (fig. 2b).

Fig. 2. Brood size (a) and male proportion (b) of progeny from Plutella xylostella larvae superparasitized with a 0 and 48 h elapsed since a previous attack in Oomyzus sokolowskii. Bars are mean ± 1 standard error.
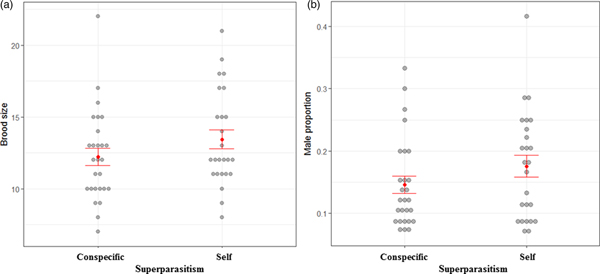
Comparison of progeny production between self- and conspecific superparasitism
Brood size did not differ between self- and conspecific superparasitism treatments (χ2 = 1.83, P = 0.18), with an average of 13.4 for self-superparasitism and 12.2 for conspecific one (fig. 3a). Sex ratio did not differ between self- and conspecific superparasitism treatments (χ2 = 2.24, P = 0.13), with an average of 18% for self-superparasitism and 15% for conspecific one (fig. 3b).

Fig. 3. Brood size (a) and male proportion (b) of progeny from Plutella xylostella larvae superparasitized by the same (self-superparasitism) and a different (conspecific superparasitism) parasitoid as in the previous attack in Oomyzus sokolowskii. Bars are mean ± 1 standard error.
Comparison of progeny production between superparasitism by parasitoids with and without oviposition experience
Brood size significantly differed between hosts superparasitized by parasitoids with and without the experience of oviposition (χ2 = 9.04, P = 0.002). It decreased to average 12.2 by a factor of 0.82 (95% CI 0.72–0.94) for the parasitoid with the experience of oviposition compared with one without it (fig. 4a). Sex ratio differed significantly between hosts superparasitized by parasitoids with and without the experience of oviposition (χ 2 = 13.18, P < 0.001), with the odds of producing male offspring by the parasitoid with the experience (sex ratio 15%) decreased by a factor of 0.56 (95% CI 0.40–0.77) compared with one without it (fig. 4b).

Fig. 4. Brood size (a) and male proportion (b) of progeny from Plutella xylostella larvae superparasitized by parasitoids with and without the experience of oviposition on unparasitized hosts in Oomyzus sokolowskii. Bars are mean ± 1 standard error.
Discussion
Our results suggest that O. sokolowskii females are able to discriminate in favor of unparasitized over parasitized hosts, not only in the willingness to attack under choice conditions but also in progeny allocation under no-choice conditions. The ability of host discrimination can be adaptive for female parasitoids in response to parasitized hosts, because superparasitizing a parasitized host would increase competition for resources among offspring in larval stage and decrease offspring fitness (Hardy et al., Reference Hardy, Griffiths and Godfray1992; Godfray, Reference Godfray1994; Koppik et al., Reference Koppik, Thiel and Hoffmeister2014). Our results from the experiment using sterilized parasitoids showed that O. sokolowskii females decrease their brood size while increasing sex ratio in response to a parasitized host as compared with an unparasitized one. Such oviposition strategy can be adaptive for the parasitoid to reduce the disadvantage of her offspring in competition with those laid by a previous parasitoid. The parasitoid progeny allocation is in line with the predictions of theoretical models (Suzuki & Iwasa, Reference Suzuki and Iwasa1980; Parker & Courtney, Reference Parker and Courtney1984). These predictions have also received the support from empirical studies of other parasitoids (Wylie, Reference Wylie1965; Werren, Reference Werren1980, Reference Werren1984; Ikawa & Suzuki, Reference Ikawa and Suzuki1982; Vet et al., Reference Vet, Datema, Janssen and Snellen1994; Shuker & West, Reference Shuker and West2004). It would be interesting and worthwhile to further investigate, though technically challenging, whether female parasitoids adjust their clutch decisions in superparasitism as a function of the number or even sex of offspring laid by previous parasitoids in the host.
In a superparasitized host, the secondary larvae are often more likely to be outcompeted by the older larvae of the first clutch, which are able to pre-empty some of the resources (Visser et al., Reference Visser, van Alphen and Nell1992c; Godfray, Reference Godfray1994; Lebreton et al., Reference Lebreton, Labarussias, Chevrier and Darrouzet2009). So, the secondary larvae suffer more from the competitive disadvantage with increasing time intervals between two attacks, if the later larvae are not able to destroy the offspring left by previous parasitoids. Our results offer support for this pattern, showing that a smaller brood with more females was produced from a host superparasitized with a 48 h interval from a previous attack than that without an interval. The support also comes from studies of other parasitoids, whereby the number of progeny from the second clutch that developed successfully on superparasitized hosts decreased with increasing time intervals between two oviposition bouts (Strand & Godfray, Reference Strand and Godfray1989; Baaren & Nénon, Reference Baaren and Nénon1996; Zaviezo & Mills, Reference Zaviezo and Mills2000). Therefore, female parasitoids are assumed to be under higher selective pressure to evolve mechanisms for avoidance of superparasitism when encountering a host with a longer time elapsed since first attack.
Competition is generally pursued to explain whether superparasitism is adaptive for superparasiting parasitoids (van Alphen & Visser, Reference van Alphen and Visser1990). An important distinction lies that while oviposition in a host previously parasitized by a conspecific leads to competition between conspecifics, oviposition in a host previously parasitized by the same female increases competition among sibs. This distinction is most obvious in solitary parasitoids, where only one of the eggs deposited in a host will be able to develop into an adult. For solitary parasitoids, self-superparasitism therefore often means a waste of the time and the egg, whereas conspecific superparasitism can be advantageous under a wider range of conditions because of the lower probability of elimination of the non-sib competitor from the parasitized host (van Alphen & Visser, Reference van Alphen and Visser1990). But for gregarious parasitoids, where more than one eggs deposited in a host will be able to develop into adults, the fitness consequences between the two kinds of superparasitism may not be as different as for solitary parasitoids. We therefore assume that gregarious parasitoids do not obtain selection favor for the discrimination in superparasitism between hosts previously parasitized by themselves and conspecifics. The assumption receives the support from our study, which showed that O. sokolowskii brood size and sex ratio did not differ between self- and conspecific superparasitism. Besides, the support also comes from studies of other gregarious parasitoids, including braconid C. glemorata attacking Pieris caterpillars (Gu et al., Reference Gu, Wang and Dorn2003) and the egg parasitoid T. evanescens attacking Ephestia kuehniell eggs (Dijken & Waage, Reference Dijken and Waage1987).
We showed that brood size and sex ratio decreased from the host superparaitized by O. sokolowskii females with the experience of ovipostion in comparison with those without it. The influence of prior oviposition experience on the willingness to superparasitization has been demonstrated in both solitary parasitoids (van Alphen et al., Reference van Alphen, Visser and Nell1992; Visser et al., Reference Visser, Luyckx, Nell and Boskamp1992a, Reference Visser, van Alphen and Hemerikb) and gregarious parasitoids as well (Ikawa & Suzuki, Reference Ikawa and Suzuki1982; Rosenheim & Rosen, Reference Rosenheim and Rosen1991; Rabinovich et al., Reference Rabinovich, Jorda and Bernstein2000). Our findings furthermore indicate that prior oviposition experience can affect progeny allocation in superparasitism by gregarious parasitoids. We assume that oviposition experience may provide the female wasp the cue about host patch quality – higher likelihood to encounter qualified hosts. After gleaning the information, parasitoids would increase their long-term fitness gains by laying a small clutch in a parasitized (thus low-quality) host to avoid running out of eggs when they are more likely to encounter high-quality hosts in the future. Theoretical and empirical studies indicate that parasitoids can make a better judge of the host encountered if they are able to learn from the past experience, and thus increase their fitness gains by adjusting oviposition strategy (Charnov et al., Reference Charnov, Los-den Hartogh, Jones and Assem1981; Turlings et al., Reference Turlings, Wackers, Vet, Lewis, Tumlinson, Papaj and Lewis1993; Henneman et al., Reference Henneman, Papaj, Figueredo and Vet1995; Flanagan et al., Reference Flanagan, West and Godfray1998).
In summary, our results showed that O. sokolowskii females discriminated in favor of unparasitized over parasitized P. xylostella larval hosts under choice conditions, and allocated a smaller brood with higher male proportion in the later than in the former hosts under no-choice conditions. Though progeny production of brood size and sex ratio did not differ between self- and conspecific superparasitism, it did for superparasitism with and without a time interval since previous attack or by parasitoids with and without oviposition experience. These performances are in line with the predictions of theoretical models. Our findings suggest that O. sokolowskii females may adjust their oviposition decisions on progeny allocation in response to a parasitized host to maximize their fitness gains from superparasitism. Our study contributes to an improved understanding of oviposition strategy for superparasitism by gregarious parasitoids.
Acknowledgements
The authors thank Yuanxing Sun for providing O. sokoloskii and Lintao Li for the help in the experiments. This study was supported by the National Key R&D Program of China (2017YFD0201000) granted to M. L.