


1. Introduction
The early Cambrian Period witnessed a profound shift from a marine ecosystem dominated by cyanobacteria to one similar to a modern-day ecosystem represented by eukaryotic algae and animals (Butterfield, Reference Butterfield2007, Reference Butterfield2011). This biotic event, which was also characterized by the rapid emergence and diversification of complex skeletal animals, was known as the ‘Cambrian Explosion’ (Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Zhu, Reference Zhu2010; Shu et al. Reference Shu, Isozaki, Zhang, Han and Maruyama2014). Strata of this age exhibit a pronounced spatial heterogeneity in the distribution of body fossils and bioturbation (Zhu, Reference Zhu2010; Buatois et al. Reference Buatois, Mángano, Olea and Wilson2016). For example, during Cambrian Age 3 (c. 521–514 Ma), shallow-water sections such as Xiaotan (Yunnan) yield abundant planktonic and benthic trilobites and intensive bioturbation, whereas deeper-water sections such as Wangjiaping (Hubei) yield few benthic but abundant planktonic trilobites without evidence of bioturbation (Zhu, Reference Zhu2010).
Animal evolution during early Cambrian time is widely inferred to have been linked to rising oxygen levels in the atmospheric–oceanic system (Nursall, Reference Nursall1959; Knoll & Carroll, Reference Knoll and Carroll1999; Sperling et al. Reference Sperling, Frieder, Raman, Girguis, Levin and Knoll2013). Evidence in support of this inference includes increases in marine sediment Mo and U concentrations and in δ98/95Mo values (to +2.34‰, which is close to modern values), suggesting seawater oxygen concentrations close to those of the modern ocean (Chen et al. Reference Chen, Ling, Vance, Shields-Zhou, Zhu, Poulton, Och, Jiang, Li, Cremonese and Archer2015b ). This nearly full oceanic oxygenation coincided with the appearance of nearly all metazoan body plans and a rise of metazoan body size represented by the Chengjiang Biota of Cambrian Age 3. However, other studies making use of geochemical redox proxies have inferred that early Cambrian deep-water settings remained anoxic and ferruginous (e.g. Canfield et al. Reference Canfield, Poulton, Knoll, Narbonne, Ross, Goldberg and Strauss2008; Sperling et al. Reference Sperling, Wolock, Morgan, Gill, Kunzmann, Halverson, Macdonald, Knoll and Johnston2015) or even euxinic (e.g. Goldberg et al. Reference Goldberg, Strauss, Guo and Liu2007; Wille et al. Reference Wille, Nagler, Lehmann, Schroder and Kramers2008). Recent geochemical investigations have documented highly heterogeneous redox conditions for the Yangtze Sea during early Cambrian time, in which mid-depth euxinic waters existed dynamically between oxic surface waters and ferruginous deep waters (Feng et al. Reference Feng, Li, Huang, Chang and Chu2014; Jin et al. Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016; Och et al. Reference Och, Cremonese, Shields-Zhou, Poulton, Struck, Ling, Li, Chen, Manning and Thirlwall2016). This pattern, dubbed the ‘euxinic wedge model’ (Li et al. Reference Li, Love, Lyons, Fike, Sessions and Chu2010, Reference Li, Cheng, Algeo and Xie2015 a; Li, Zhu & Chu, Reference Li, Zhu and Chu2016), may provide new insights regarding the influence of spatially heterogeneous redox conditions on early animal evolution.
In order to investigate the relationship between redox heterogeneity and early animal evolution, we conducted a high-resolution Fe-S-C-Al-Ti chemostratigraphic study of the outer-shelf Wangjiaping section from lower Cambrian strata of the Yangtze Block. The record of animal evolution at Wangjiaping is different from that at typical inner-shelf sections, such as Xiaotan (Yunnan Province) and Shatan (Sichuan Province). At Wangjiaping, the Shuijingtuo Formation yielded a few benthic trilobites of Metaredlichia sp. belonging to the Redlichiacea but abundant planktonic trilobites represented by Eodiscina, which are different from the abundant planktonic and benthic trilobites belonging to the Redlichiacea from the correlative strata at Xiaotan and Shatan (Xiang et al. Reference Xiang, Zhou, Ni, Zeng, Xu, Lai, Wang, Li, Wang, Xiang, Ni, Zeng, Xu, Zhou, Lai and Li1987). Our new data, in combination with published Fe-speciation data from the inner-shelf Xiaotan and Shatan sections, provided us with an opportunity to evaluate the relationship between redox heterogeneity and animal spatial distribution on the early Cambrian Yangtze Block.
2. Materials and methods
A total of 34 samples were collected from fresh outcrop at Wangjiaping in July 2015. Large rock blocks (>200 g) were sampled and all potentially weathered surfaces, post-depositional veins, and visible pyrite nodule and bands were trimmed away prior to powdering. Each sample was crushed into powders for measurement of total organic (TOC) and carbonate (TIC) carbon, iron speciation data, Al, Ti and pyrite sulphur isotopic compositions (δ34Spy). All geochemical data were generated at the State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences (Wuhan).
TIC was determined by the difference between total carbon (TC) and TOC, measured using a Jena Multi-EA 4000 carbon-sulphur analyser. For TC analysis, c. 100 mg of powder were weighed into a ceramic boat, combusted at 1350°C and measured using calibrated infrared sensors. For TOC analysis, c. 500 mg of powder were treated with 4-N HCl to remove carbonate, washed several times with deionized water to remove HCl residue, and finally dried overnight at 50°C. About 100 mg of the residue were analysed for TOC content following the same procedures as for TC. Analytical uncertainty is better than ±0.2% for TC and TOC based on replicate analyses of Alpha Resources standard AR-4007 (TC = 7.62%).
Highly reactive iron (FeHR) is the sum of carbonate-associated Fe (Fecarb), Fe oxides (Feox), magnetite Fe (Femag) and pyrite Fe (Fepy) (i.e. FeHR = Fecarb+Feox+Femag+Fepy). Fe sequential extractions were conducted following standard procedures for Fecarb, Feox and Femag (Poulton & Canfield, Reference Poulton and Canfield2005). About 100 mg of sample powder was treated with a sodium acetate solution buffered to a pH of 4.5 by addition of analytical-grade acetic acid at 50°C for 48 h to extract Fecarb. The sample residue from the first step was treated with a 50 g L−1 sodium dithionite solution buffered to a pH of 4.8 by addition of 0.2 M sodium citrate and analytical-grade acetic acid at room temperature for 2 h to extract Feox. The sample residue from the second step was treated with a 0.2 M ammonium oxalate and 0.17 M oxalic acid solution buffered to a pH of 3.2 by addition of analytical-grade ammonium water at room temperature for 6 h to extract Femag. After dilution with 2% HNO3 all extracts were measured for Fe content using atomic absorption spectroscopy (AAS), yielding an RSD of <5%. Fepy was stoichiometrically determined from chromium reducible pyrite (Canfield et al. Reference Canfield, Raiswell, Westrich, Reaves and Berner1986). Unreactive iron (FeU) was determined from the difference between total iron (FeT) and FeHR. A more detailed description of the Fe speciation methods can be found in Li et al. (Reference Li, Planavsky, Shi, Zhang, Zhou, Cheng, Tarhan, Luo and Xie2015b ).
FeT, Al and Ti were determined by X-ray fluorescence spectrometry (XRF) of fused glass beads. About 0.6 g of sample powders, a 6 g mixture of Li2B4O7, LiBO2 and LiF, and 0.3–0.4 g of ammonium nitrate as a fluxing medium were loaded into a porcelain crucible and mixed well with a spatula. The mixture was loaded into a platinum crucible, to which was added 5–6 drops of lithium bromide solution, before being finally transferred to the fusion machine to produce a glass bead. Analytical errors are better than ±5% for FeT, Al and Ti based on replicate analyses of two Chinese (GBW07107, GBW07108) standards.
3. Geological background
Palaeogeographic reconstructions show that South China was an isolated, peri-equatorial continental block close to northeastern Gondwana (modern Australia) during early Cambrian time (Fig. 1a; Yang, Steiner & Keupp, Reference Yang, Steiner and Keupp2015; Zhang et al. Reference Zhang, Li, Jiang, Evans, Dong, Wu, Yang, Liu and Xiao2015). The South China Block was composed of the Yangtze and Cathaysia blocks, which had been sutured together during the early Neoproterozoic Era, but continued to undergo some compressional deformation as late as the Late Ordovician ‘Caledonian’ Orogeny (Wang & Li, Reference Wang and Li2003). During the Ediacaran–Cambrian transition, the Yangtze Block consisted of the shallowly submerged Yangtze Platform bordered on the SE by a ramp descending into the deep-water Nanhua Basin, which had developed over a Neoproterozoic failed rift initialized at c. 820 Ma (Wang & Li, Reference Wang and Li2003).

Figure 1. Geological background of study section. (a) Global palaeogeographic reconstruction for Cambrian Stage 3 (modified from Yang, Steiner & Keupp, Reference Yang, Steiner and Keupp2015). (b) Palaeogeography of Yangtze Block during early Cambrian Fortunian and early Stage 2 ages (modified from Goldberg et al. Reference Goldberg, Strauss, Guo and Liu2007). The study section Wangjiaping and two correlated inner-shelf sections Xiaotan and Shatan are marked. (c) Lithological and stratigraphic profiles of the lower Cambrian Wangjiaping section. The U–Pb age is from the nearby Wuhe section (Okada et al. Reference Okada, Sawaki, Komiya, Hirata, Takahata, Sano, Han and Maruyama2014). ED – Ediacaran; F – Fortunian Stage; S2 – Stage 2; DY – Dengying Formation; YJH – Yanjiahe Formation; LM – Lower Member; and UM – Upper Member.
During the Cambrian Fortunian Age to early Age 2 (c. 541–529 Ma), sedimentation on the Yangtze Block was characterized by mainly carbonate deposits on the platform to the NW, interbedded shale, chert and carbonate deposits on the ramp, and mudstone/shale or chert deposits in the basin to the SE (Fig. 1b; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Zhu, Zhang & Yang, Reference Zhu, Zhang and Yang2007). A major transgression since the middle Cambrian Age 2 caused widespread deposition of black shale across the Yangtze Platform.
The Wangjiaping section (30°48′41″N and 111°11′12″E) is sited west of Yichang city in central Hubei Province. Palaeogeographically, this area represented outer-shelf facies during the early Cambrian Period (Zhu et al. Reference Zhu, Zhang, Yang, Li, Steiner and Erdtmann2003). The section consists of the Yanjiahe and Shuijingtuo formations (Fig. 1c). The Yanjiahe Formation, which disconformably overlies the late Ediacaran Dengying Formation, consists of sandy dolomite, phosphorite layers, black shale and dolomite. The small shelly fossil assemblages 1 (SSFA1, Anabarites trisulcantus–Protohertzina unguliformis) and 3 (SSFA3, Watsonella crosbyi) have been identified in the correlative Yanjiahe section, c. 15 km to the SW (Chen, Reference Chen1984). The overlying Shuijingtuo Formation is divided into two members based on lithology: (1) a lower member (LM; 0–15 m) consisting of black shale interbedded with calcareous mudstone and muddy limestone, in which are found abundant planktonic trilobites such as S. chanyangensis and H. orientalis; and (2) an upper member (UM; 15–21 m) consisting of muddy limestone, in which are found planktonic trilobites such as H. orientalis (Xiang et al. Reference Xiang, Zhou, Ni, Zeng, Xu, Lai, Wang, Li, Wang, Xiang, Ni, Zeng, Xu, Zhou, Lai and Li1987). Only one benthic trilobite fossil of Metaredlichia sp. was found in the muddy limestone layer at c. 14.8 m close to the LM–UM boundary (Fig. 1c; Xiang et al. Reference Xiang, Zhou, Ni, Zeng, Xu, Lai, Wang, Li, Wang, Xiang, Ni, Zeng, Xu, Zhou, Lai and Li1987). The lithological change from LM to UM is consistent with the marine regression recorded basinwide (Yang, Zhu & Zhang, Reference Yang, Zhu and Zhang2005). A U–Pb zircon age from ash beds at the base of the Shuijingtuo Formation black shale in the Wuhe section located c. 30 km to the SW of Wangjiaping provides a maximum age of 526.4±5.4 Ma (i.e. mainly of Age 3) for the Shuijingtuo Formation (Okada et al. Reference Okada, Sawaki, Komiya, Hirata, Takahata, Sano, Han and Maruyama2014).
4. Results
The Fe-S-C-Al-Ti data for the Wangjiaping section are provided in Table 1 and Figures 2–4. TOC concentrations range over 0.1–3.6% (mean 2.0%) and exhibit a stepwise decreasing trend upsection (Fig. 2). TIC concentrations are relatively variable, ranging over 0.8–11.6‰ (mean 4.3%). δ34Spy ranges from –14.1 to +19.3‰ (mean ‒0.1‰). Al ranges over 1.1–10.0% (mean 6.4%), roughly increasing upsection in the LM (0–15 m) of the Shuijingtuo Formation; it ranges over 0.7–4.1% (mean 2.2%), decreasing upsection in the UM (15–21 m) of the Shuijingtuo Formation. Ti ranges over 0.05–0.53% (mean 0.33%) and over 0.03–0.19% (mean 0.10%) in the LM and UM, respectively. FeU ranges over 0–2.4% (mean 0.68%) and over 0.07–0.71% (mean 0.30%) in the LM and UM, respectively. Ti and FeU exhibit similar patterns of stratigraphic variation to Al: increasing upsection within the LM and then decreasing upsection within the UM.
Table 1. Key geochemical data in the outer-shelf Wangjiaping section

a BS – black shale; CM – calcareous mudstone; ML – muddy limestone; bFor samples with measured FeU < 0, these values are reported as 0; c For samples with measured FeHR/FeT > 1, these values are reported as 1
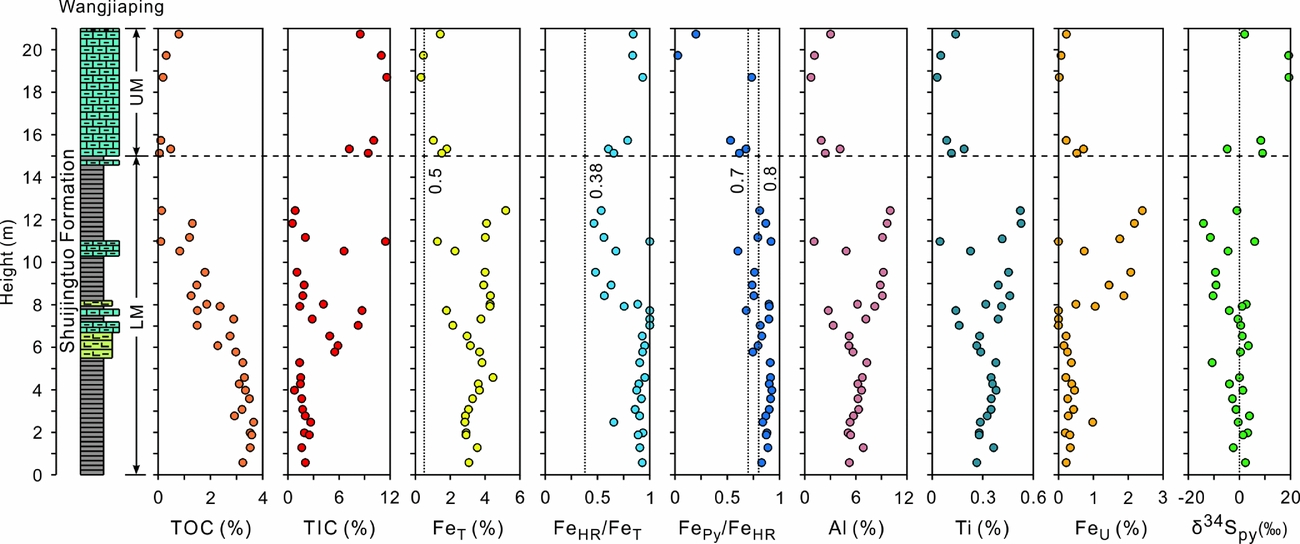
Figure 2. Composite chemostratigraphy of Wangjiaping section. The vertical dashed lines (from left to right) for each section indicate key values of FeT (0.5 wt%), FeHR/FeT (0.38) and Fepy/FeHR (0.7–0.8) used in redox interpretations (see text for details).
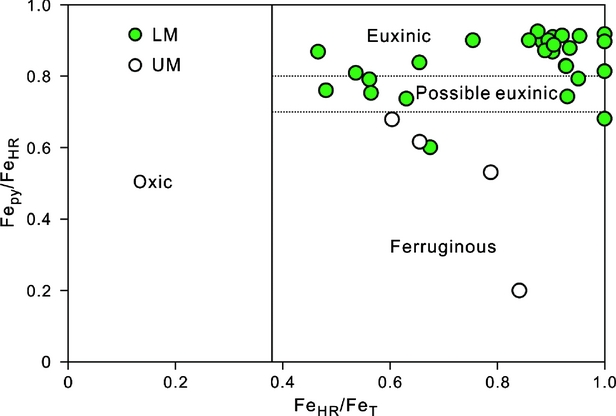
Figure 3. Cross-plot of FeHR/FeT v. FePy/FeHR for samples with FeT >0.5 wt% from the Wangjiaping section. Threshold values of FeHR/FeT (0.38) for oxic-anoxic conditions and FePy/FeHR (0.7-0.8) for ferruginous-euxinic conditions from Poulton & Canfield (Reference Poulton and Canfield2011).

Figure 4. Cross-plot of TOC v. FePy/FeHR for Wangjiaping section. Note that the three samples which are considered as outliers (unfilled symbols) were not included in the correlation.
FeT ranges over 1.27–5.18% (mean 3.40%), roughly increasing upsection in the LM; it ranges over 0.32–1.8% (mean 1.09%), decreasing upsection in the UM. FeHR/FeT ratio over the range 0.47–1.00 (0.82±0.17; mean±1σ), decreasing upsection in the LM; it varies over 0.60–0.93 (0.78±0.12), increasing upsection in the UM. FePy/FeHR ratios vary over the range 0.03–0.93 (0.77±0.20), and show a generally decreasing trend upsection.
5. Discussion
5.a. Basis for stratigraphic correlation of lower Cambrian deposits in South China
The stratigraphic division and correlation of lower Cambrian units on the Yangtze Block are mainly based on fossils such as small shelly faunas (SSFs) and trilobites (Yuan & Zhao, Reference Yuan and Zhao1999; Yang et al. Reference Yang, Zhu, Zhang and Li2003; Zhu et al. Reference Zhu, Zhang, Yang, Li, Steiner and Erdtmann2003; Yang, Zhu & Zhang, Reference Yang, Zhu and Zhang2005; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007), distinctive marker beds such as a black shale and Ni–Mo layer at the base of the Cambrian (Yang et al. Reference Yang, Zhu, Zhang and Li2003; Zhu et al. Reference Zhu, Zhang, Yang, Li, Steiner and Erdtmann2003; Jiang et al. Reference Jiang, Wang, Shi, Xiao, Zhang and Dong2012; Chen et al. Reference Chen, Zhou, Fu, Wang and Yan2015 a), and some zircon ages (Wang et al. Reference Wang, Shi, Jiang and Zhang2012; Chen et al. Reference Chen, Zhou, Fu, Wang and Yan2015 a). The presence of SSFA1 and SSFA3 in the lower and upper Yanjiahe Formation suggest the Cambrian Fortunian Age (c. 541–529 Ma) and Age 2 (c. 529–521 Ma), respectively (Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Peng, Babcock & Cooper, Reference Peng, Babcock, Cooper, Gradstein, Ogg, Schmitz and Ogg2012). The U–Pb zircon age of 526.4±5.4 Ma from ash beds at the base of the Shuijingtuo Formation black shale at nearby Wuhe section also indicates a Cambrian Age 2 (Okada et al. Reference Okada, Sawaki, Komiya, Hirata, Takahata, Sano, Han and Maruyama2014). The Shuijingtuo Formation contains trilobites such as S. chanyangensis, H. orientalis and Metaredlichia sp., which are of Cambrian Age 3 (c. 521–514 Ma), in which the first appearance of H. orientalis from the calcareous mudstone and muddy limestone marks the base of Yunnanaspis–Yiliangella trilobite Zone in the Hongjingshao Formation from some sections of Yunnan Province (e.g. Xiaotan; Yang, Zhu & Zhang, Reference Yang, Zhu and Zhang2005). We should note that, although the Ni–Mo ore layer is the marker bed for the boundary of Cambrian ages 2 and 3 (c. 521 Ma; Zhu et al. Reference Zhu, Zhang, Yang, Li, Steiner and Erdtmann2003), it was not found at Wangjiaping. However, this layer was reported at the base of the Shuijingtuo Formation in the nearby Jijiawan (c. 10 km to the west) and Yanjiahe (c. 30 km to the SW) sections of the Yangtze Gorges region (Chen et al. Reference Chen, Zhou, Fu, Wang and Yan2015 b). At Wangjiaping, the Ni–Mo ore layer may be located in an inaccessible covered interval. The framework of stratigraphic correlations mentioned above, which was discussed in greater detail by Jin et al. (Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016), provides a solid basis for the stratigraphic correlations among the study sections in Figure 5.
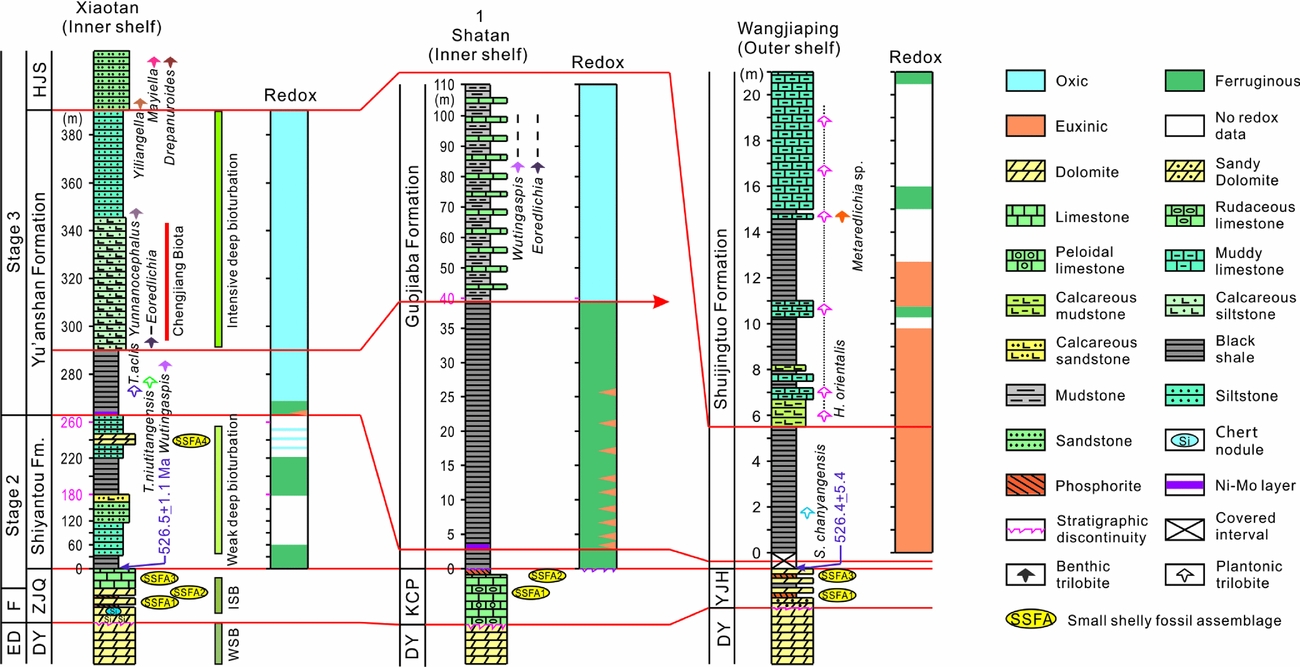
Figure 5. Comparison of fossil records with water-column redox conditions across the Yangtze Block, as reconstructed from geochemical data for Xiaotan, Shatan and Wangjiaping. The U–Pb age at Xiaotan is from the nearby Meichucun section (Compston et al. Reference Compston, Zhang, Cooper, Ma and Jenkins2008). The lithostratigraphy, biostratigraphy and redox conditions of the Xiaotan and Shatan sections were discussed in detail by Feng et al. (Reference Feng, Li, Huang, Chang and Chu2014) and Jin et al. (Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016), while related data of the Wangjiaping are from this study. See Section 5.a. for the basis of stratigraphic correlation between these sections. Abbreviations: ZJQ – Zhujiaqing Formation; HJS – Hongjingshao Formation; and KCP – Kuanchuanpu Formation.
5.b. Marine redox conditions at Wangjiaping
Sedimentary iron speciation has been widely used to distinguish oxic, ferruginous and euxinic conditions in siliciclastic rocks as well as in carbonate rocks with >0.5% FeT (e.g. Lyons & Severman, Reference Lyons and Severmann2006; Canfield et al. Reference Canfield, Poulton, Knoll, Narbonne, Ross, Goldberg and Strauss2008; Li et al. Reference Li, Love, Lyons, Fike, Sessions and Chu2010, Reference Li, Love, Lyons, Scott, Feng, Huang, Chang, Zhang and Chu2012; Poulton & Canfield, Reference Poulton and Canfield2011; Clarkson et al. Reference Clarkson, Poulton, Guilbaud and Wood2014). Modern and ancient anoxic sediments yield FeHR/FeT ratios >0.38 through addition of FeHR from the anoxic water column (Raiswell & Canfield, Reference Raiswell and Canfield1998). Under anoxic and sulphidic (euxinic) conditions, FeHR tends to be converted to Fepy through its reaction with H2S to form pyrite, resulting in Fepy/FeHR ratios of >0.7–0.8 (Poulton & Canfield, Reference Poulton and Canfield2011).
All samples in the lower Shuijingtuo Formation (0–15 m; i.e. LM) are characterized by >0.5% FeT, elevated FeHR/FeT ratios (>0.38) and high Fepy/FeHR ratios (0.60–0.93; mean 0.84), suggesting dominantly euxinic conditions punctuated by ferruginous episodes (Figs 2, 3). Samples from the upper Shuijingtuo Formation (15–21 m; i.e. UM) exhibit FeT >0.5% (except for two at 18.12 and 19.15 m), elevated FeHR/FeT ratios (>0.38) and low Fepy/FeHR ratios (<0.7), indicating ferruginous conditions. Iron speciation data at Wangjiaping therefore suggest a redox transition from dominantly euxinic conditions in the LM to ferruginous conditions in the UM of the Shuijingtuo Formation.
5.c. Interpreting the pyrite sulphur isotope record
The isotopic composition of sulphur in sedimentary pyrite (δ34Spy) has been widely used to explore biogeochemical cycling of sulphur in the Earth-surface system (e.g. Scott et al. Reference Scott, Wing, Bekker, Planavsky, Medvedev, Bates, Yun and Lyons2014). Among other things, pyrite δ34S can provide important insights into the influence of seawater sulphate concentrations on the development of mid-depth euxinia (e.g. Feng et al. Reference Feng, Li, Huang, Chang and Chu2014). Variation in δ34Spy depends mainly on the fraction of pyrite burial (f py), the isotopic composition of sulphur entering ocean (δ34Sin) and fractionation during microbial sulphate reduction (MSR) (εMSR). MSR enriches sulphide in the light isotopes of sulphur relative to the seawater sulphate source. Many factors influence εMSR in nature, including aqueous sulphate concentrations, sulphur oxidation and disproportionation, environments of pyrite formation, and MSR rates (e.g. Canfield & Teske, Reference Canfield and Teske1996; Habicht et al. Reference Habicht, Gade, Thamdrup, Berg and Canfield2002; Jones & Fike, Reference Jones and Fike2013; Leavitt et al. Reference Leavitt, Halevy, Bradley and Johnston2013; Algeo et al. Reference Algeo, Luo, Song, Lyons and Canfield2015). MSR can produce a fractionation of ≥60‰ under sulphate-replete conditions (Canfield, Farquhar & Zerkle, Reference Canfield, Farquhar and Zerkle2010; Sim, Bosak & Ono, Reference Sim, Bosak and Ono2011), whereas sulphate-poor conditions yield minimal fractionation (e.g. <10‰ at 200 μM SO4 2‒; Habicht et al. Reference Habicht, Gade, Thamdrup, Berg and Canfield2002; Gomes & Hurtgen, Reference Gomes and Hurtgen2013; Crowe et al. Reference Crowe, Paris, Katsev, Jones, Kim, Zerkle, Nomosatryo, Fowle, Adkins, Sessions, Farquhar and Canfield2014). In addition to MSR, sulphur oxidation and disproportionation can lead to a significant increase in εMSR (Canfield & Teske, Reference Canfield and Teske1996). With other factors held constant, high and low MSR rates result in reduced and increased isotopic fractionation, respectively, yielding values ranging over 0–70‰ (Sim, Bosak & Ono, Reference Sim, Bosak and Ono2011).
Our iron speciation data suggest that the Wangjiaping environment was continuously anoxic throughout the study interval, with samples in the LM and UM of the Shuijingtuo Formation accumulating under dominantly euxinic and ferruginous conditions, respectively (Section 5.b). Sulphur oxidation and disproportionation would therefore have had little to no influence on δ34Spy values in the study section. Given dominantly euxinic conditions in the LM of the Shuijingtuo Formation, pyrite would have formed mainly in the water columns. This unit shows intermediate δ34Spy values at 0–8 m (‒10.7 to +3.9‰, mean ‒0.2‰), followed by a shift to more negative values at 8–12 m (‒14.1 to 6.0‰, mean ‒6.7‰; Fig. 2). Relative to early Cambrian seawater sulphate, which had a δ34S value of +35 to +40‰ (Kampschulte & Strauss, Reference Kampschulte and Strauss2004; Goldberg, Poulton & Strauss, Reference Goldberg, Poulton and Strauss2005), these δ34Spy values indicate large MSR fractionations, that is, εMSR ≈ 29–54‰. This range of fractionations yields estimated seawater sulphate concentrations of c. 8–30 mM based on the MSR trend method of Algeo et al. (Reference Algeo, Luo, Song, Lyons and Canfield2015), that is, close to or somewhat lower than the modern seawater level of 29 mM. The negative shift in δ34Spy values upsection in the LM is roughly correlated with a decrease in organic matter as suggested by TOC, as well as the enhanced riverine flux as suggested by a rise of Al, Ti and FeU (Fig. 2), suggesting that the reduced MSR rate caused by a decrease in organic matter loading, or/and a rise of relatively 34S-depleted riverine sulphate to MSR, may have led to the decreasing δ34Spy values. The later mechanism has been proposed to explain the decrease of δ34Spy values from the correlative strata in the inner-shelf Shatan, outer-shelf Yangjiaping and Weng'an sections of the Yangtze Block (Feng et al. Reference Feng, Li, Huang, Chang and Chu2014; Jin et al. Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016).
Given ferruginous conditions in the UM of the Shuijingtuo Formation (Section 5.b), the study section was probably located around the chemocline (e.g. in the upper region of the oceanic thermocline), and pyrite formation increasingly occurred in the sediments. Low sulphate concentrations can develop within the sediment when sulphate consumption exceeds supply through diffusion, a condition that limits the degree of sulphur isotopic fractionation expressed between pyrite and porewater sulphate (e.g. Lyons et al. Reference Lyons, Werne, Hollander and Murray2003). Reduced diffusion of sulphate from the overlying water column into the sediment may therefore have caused the δ34Spy values to shift positively and to fall in a wide range (‒4.8 to +19.3‰, mean +8.9‰) in the UM of the Shuijingtuo Formation (Fig. 2).
5.d. Controls on development of euxinia at Wangjiaping
The tendency of anoxic waters toward euxinic versus ferruginous conditions depends on the relative fluxes of FeHR and H2S, with production of the latter controlled mainly by microbial sulphate reduction (MSR) linked to organic matter decay (Raiswell & Canfield, Reference Raiswell and Canfield2012; Feng et al. Reference Feng, Li, Huang, Chang and Chu2014). Given a certain FeHR supply in a marine system, the spatiotemporal occurrence of euxinia may have been controlled mainly by the local availability of marine organic matter and/or seawater sulphate (Li et al. Reference Li, Love, Lyons, Fike, Sessions and Chu2010, Reference Li, Love, Lyons, Scott, Feng, Huang, Chang, Zhang and Chu2012, Reference Li, Cheng, Algeo and Xie2015 a).
A large change in δ34Spy values from proximal to distal areas of a marine system is indicative of a gradient in seawater sulphate concentrations. The early Cambrian Yangtze Block exhibits such a gradient, with a mean δ34Spy of ‒12.0‰ in the inner-shelf Xiaotan section, shifting progressively to a mean of +22.5‰ in the basinal Longbizui section (Feng et al. Reference Feng, Li, Huang, Chang and Chu2014; Jin et al. Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016). Within this interpretative framework, the outer-shelf Wangjiawan section of the present study exhibits intermediate δ34Spy (‒2.1±5.3‰) and TOC (2.4±1.1%) values (Fig. 2). These values are nonetheless substantially lower than in the correlative outer-shelf Yangjiaping (δ34Spy = +8.3±4.7‰, TOC = 10.7±2.6%), slope Songtao (δ34Spy = +8.6±7.7‰, TOC = 8.3±4.0%) and slope Longbizui sections (δ34Spy = +21.0±6.8‰, TOC = 6.2±1.7%) (Feng et al. Reference Feng, Li, Huang, Chang and Chu2014). Note that all sections accumulated under euxinic conditions except for Longbizui, where ferruginous conditions prevailed. In view of these relationships, the relative fluxes of sulphate to organic matter at Wangjiaping are inferred to have been higher than in the other, deeper-water sections. When anoxic conditions are indicated in palaeomarine systems, the relative availability of organic matter versus sulphate is therefore likely to play a key role in the development of euxinia.
If our hypothesis is correct, we should expect to see increasing euxinic tendency along with increasing productivity at Wangjiaping instead of at those deeper sites. Indeed, a strong positive correlation between TOC and Fepy/FeHR (R 2 = 0.66; p < 0.01) was observed for samples (note that all have FeT > 0.5%) at Wangjiaping (Fig. 4). However, only weak correlations between TOC and Fepy/FeHR (R 2 ≤ 0.22) for anoxic samples were observed from correlative strata at the outer-shelf Yangjiaping, slope Songtao and Longbizui sections (Feng et al. Reference Feng, Li, Huang, Chang and Chu2014). These observations provide compelling evidence for organic control on the local euxinic formation at Wangjiawan and the high spatial heterogeneity of marine sulphate concentration recently recognized in Neoproterozoic–Cambrian oceans (Li et al. Reference Li, Love, Lyons, Fike, Sessions and Chu2010, Reference Li, Love, Lyons, Scott, Feng, Huang, Chang, Zhang and Chu2012; Feng et al. Reference Feng, Li, Huang, Chang and Chu2014; Jin et al. Reference Jin, Li, Peng, Cui, Shi, Zhang, Luo and Xie2014, Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016).
5.e. Spatial variation of early Cambrian redox conditions and metazoan evolution
The early Cambrian evolution of metazoans recorded by fossil distributions is characterized by a highly spatial heterogeneity in South China as well as in the global ocean (e.g. Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Zhu, Reference Zhu2010). In Cambrian Fortunian and Stage 2 strata, SSFA1 and SSFA3 were widely found in shelf settings on the Yangtze Block as well as on other cratons (Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Peng, Babcock & Cooper, Reference Peng, Babcock, Cooper, Gradstein, Ogg, Schmitz and Ogg2012; Landing et al. Reference Landing, Geyer, Brasier and Bowring2013). However, SSFA2 (represented by Paragloborilus subglobosus – Purella squamulosa) and SSFA4 (represented by Sinosachites flabelliformis – Tannuolina zhangwentangi) are restricted to a few locales on the Yangtze Block (e.g. eastern Yunnan and Sichuan provinces; Steiner et al. Reference Steiner, Li, Qian, Zhu and Erdtmann2007). In Cambrian Stage 3 deposits, planktonic and benthic trilobites appear in shallow-water sections on the Yangtze Block (e.g. in Yunnan, Sichuan and parts of Guizhou Province; Zhu, Reference Zhu2010), whereas few benthic but abundant planktonic trilobites were found in relatively deeper-water settings (e.g. in the Yangtze Gorges region of Hubei Province and the Weng'an and Songtao sections in Guizhou Province; Xiang et al. Reference Xiang, Zhou, Ni, Zeng, Xu, Lai, Wang, Li, Wang, Xiang, Ni, Zeng, Xu, Zhou, Lai and Li1987; Yang et al. Reference Yang, Zhu, Zhang and Li2003; Zhu et al. Reference Zhu, Zhang, Yang, Li, Steiner and Erdtmann2003). During early Cambrian time, shallow-water settings globally are characterized by more intensive bioturbation than deep-water settings (Buatois et al. Reference Buatois, Mángano, Olea and Wilson2016). A similar pattern exists on the Yangtze Block, as shown by a transition from weak to intensive bioturbation beginning at the base of Cambrian Stage 3 strata in the inner-shelf Xiaotan section, but a complete lack of bioturbation in coeval strata of the outer-shelf Wangjiaping section (Fig. 5; Xiang et al. Reference Xiang, Zhou, Ni, Zeng, Xu, Lai, Wang, Li, Wang, Xiang, Ni, Zeng, Xu, Zhou, Lai and Li1987; Shields-Zhou & Zhu, Reference Shields-Zhou and Zhu2013).
Because animals need oxygen to sustain their metabolism, the spatial variability of redox conditions in early Cambrian oceans may have directly controlled the spatial pattern of metazoan distribution. Where euxinic or ferruginous conditions existed, rare benthic fossils are found. The appearance of benthic trilobites such as Eoredlichia and Wutingaspis coincided with the development of persistently oxic bottom-water conditions, for example, beginning in Cambrian Stage 3 deposits of the inner-shelf Xiaotan and Shatan sections. However, only planktonic trilobites such as S. changyangensis and H. orientalis are present in correlative strata deposited under euxinic bottom-water conditions in the outer-shelf Wangjiaping section (Fig. 5). Given that benthic trilobites have been found in black shale, mudstone, calcareous siltstone, muddy limestone and sandstone facies (Yang et al. Reference Yang, Zhu, Zhang and Li2003; Zhu et al. Reference Zhu, Zhang, Yang, Li, Steiner and Erdtmann2003; Zhu, Reference Zhu2010; this study), the spatial distribution of these fossils does not appear to have been controlled by lithology. The fact that euxinic conditions inhibit organic decay and enhance fossil preservation (Gaines et al. Reference Gaines, Hammarlund, Hou, Qi, Gabbott, Zhao, Peng and Canfield2012; Wang et al. Reference Wang, Guan, Zhou, Wan, Tang, Chen, Chen and Yuan2014) suggests that the spatial heterogeneity of fossil distribution on the early Cambrian Yangtze Block does not reflect differential taphonomic effects, because benthic trilobites should have been preserved if they indeed lived in euxinic waters. We therefore infer that spatial variability in marine redox conditions was a key control on metazoan distribution and marine ecosystem development in early Cambrian oceans (cf. Gilleaudeau & Kah, Reference Gilleaudeau and Kah2015; Li et al. Reference Li, Cheng, Algeo and Xie2015 b).
Variability in patterns of bioturbation in early Cambrian oceans (Buatois et al. Reference Buatois, Mángano, Olea and Wilson2016) may also have been linked to spatial heterogeneity in redox conditions (Jin et al. Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016). Reduced benthic oxygen levels have been shown to correlate with shorter burrow lengths and lower burrow densities (Sturdivant, Diaz & Cutter, Reference Sturdivant, Diaz and Cutter2012). In the Yangtze Block, bioturbation occurred in the shallow-water setting (e.g. Xiaotan; Shields-Zhou & Zhu, Reference Shields-Zhou and Zhu2013) with oxic conditions, but no bioturbation was found in the relatively deeper-water settings (e.g. Wangjiaping) with euxinic conditions (Fig. 5). Furthermore, similarly, widespread anoxic conditions during early Cambrian time, possible due to low oxygen levels of less than 40% present in the atmosphere (Sperling et al. Reference Sperling, Wolock, Morgan, Gill, Kunzmann, Halverson, Macdonald, Knoll and Johnston2015), could have resulted in the protracted development of bioturbation (Tarhan et al. Reference Tarhan, Droser, Planavsky and Johnston2015), whereas the expansion of shelf oxic waters during Cambrian Age 3 (Jin et al. Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016) might be responsible for the increase in bioturbation (Buatois et al. Reference Buatois, Mángano, Olea and Wilson2016).
6. Conclusions
This study presents Fe-C-S-Al-Ti geochemical data from lower Cambrian strata of the outer-shelf Wangjiaping section on the Yangtze Block of South China. Iron speciation data indicate a redox change from dominantly euxinic to ferruginous conditions during Cambrian Age 3, accompanied by a fall in sea level. A good correlation between TOC and Fepy/FeHR suggests that organic matter availability limited development of euxinia at Wangjiaping. Pyrite δ34S is more negative than that in correlative deeper-water sections, suggesting higher sulphate concentrations at Wangjiaping. These observations are consistent with the hypothesis that the relative availability of sulphate versus organic matter controlled development of euxinia in the lower Cambrian strata of South China, and that organic matter was relatively limiting at Wangjiaping whereas sulphate was relatively limiting in deeper-water sections. Our work therefore provides evidence for the marine sulphate gradients from proximal to distal oceans previously proposed for Neoproterozoic–Cambrian oceans.
Redox conditions in comparison with fossils in shelf sections, including inner-shelf Xiaotan and Shatan and outer-shelf Wangjiaping, show that planktonic and benthic trilobites lived in oxic conditions, whereas only planktonic trilobites thrived in water masses having anoxic or euxinic bottom-water conditions. Further correlation suggests that bioturbation occurred in conjunction with oxic conditions, whereas no bioturbation developed under anoxic to euxinic bottom-water conditions. Similar patterns of spatial heterogeneities are inferred to have existed for redox conditions and animal evolution globally. These findings therefore suggest that the spatial heterogeneity of redox conditions in the shelf settings may have played a key role in the spatial distribution of early animals during early Cambrian time.
Acknowledgements
We thank Shida Tang and Haiyang Wang for laboratory assistance. This study was supported by the Chinese 973 program (grant no. 2013CB955704) and the NSFC (grant no. 41172030). Research by TJA is supported by the US National Science Foundation (Sedimentary Geology and Paleobiology program, grant no. EAR-1053449), the NASA Exobiology program (grant no. NNX13AJ1IG) and the China University of Geosciences-Wuhan (SKL-GPMR program GPMR201301 and SKL-BGEG program BGL21407).






