The importance of plant foods to subsistence during North America's fluted-point period (FPP), ~13,500–12,000 cal BP, is contested. Many FPP sites containing subsistence remains are kill sites, and the era's few recorded camp sites rarely contain hearth features with preserved macrobotanical remains, facts that cloud the role of plants in the FPP diet (e.g., Cannon and Meltzer Reference Cannon and Meltzer2004; Waguespack and Surovell Reference Waguespack and Surovell2003). The near absence of plant remains in FPP sites may be due to poor preservation, but this does not mean that any evidence of plant use in these sites indicates a broad diet (e.g., Gingerich Reference Gingerich2011).
This study aims to understand both whether the scarcity of plant remains in FPP sites is attributable to poor preservation of archaeological features and what the few plant remains present in FPP hearths indicate about subsistence activities. In the Northeast (the New England states, New Jersey, New York, and Pennsylvania), there are eight FPP sites containing a total of 11 well-dated features (Figure 1; Table 1), six of which contain carbonized macrobotanical remains recovered via flotation (unfortunately, features from Debert, Neponset, and Vail were not floated and thus are not included in discussions of subsistence activities). Although the sample is small, it is the largest collection of macrobotanical remains associated with fluted-point technology from any comparably sized region in North America.

Figure 1. Location of FPP sites with radiocarbon dates from feature and nonfeature contexts included in this study (map by Nathaniel R. Kitchel).
Table 1. List of Fluted-Point Period Features Indicating the Presence or Absence of Plant Remains.

Note: At Shawnee-Minisink Gingerich (Reference Gingerich2011) has argued that only hawthorn (Crataegus sp.) and possibly blackberries (Rubus sp.) were consumed by the inhabitants of the site and that other plant remains were the product of accidental carbonization. At Shawnee-Minisink only hawthorn and hickory (Carya sp.) were recovered from secure feature contexts, whereas other small seeds were recovered from matrix samples (Gingerich Reference Gingerich2011:136) and are not included in this analysis.
Through an analysis of feature destruction over time, we show that FPP hearths are as common as expected given the small number of well-dated FPP sites in the Northeast. We also show that they contain macrobotanical remains at a higher frequency than hearths in a region with better preservation and where small seeds probably formed an important part of the diet. These findings indicate that plant remains are relatively common in Northeast FPP hearths and that the record is not biased against the preservation of plant macrobotanical remains. However, the plant remains recovered from Northeast FPP hearths show a preference for fleshy fruits, a pattern most consistent with a relatively narrow plant diet breadth. Although these results indicate that plant foods played a role in the diet during the Northeast FPP, they do not support the frequent use of labor-intensive plant foods like small seeds.
Plant Foods and Subsistence Specialization
The use of plant foods during the FPP intersects with an unresolved debate over whether terminal Pleistocene North American hunter-gatherers were specialized big game hunters (see Cannon and Meltzer Reference Cannon and Meltzer2004:1971; Waguespack and Surovell Reference Waguespack and Surovell2003). Subsistence specialization is rare and occurs in few ecological settings, usually in species-poor environments with an abundance of a single taxon, such as the Arctic tundra (Meltzer and Smith Reference Meltzer, Smith and Neusius1986:8–9). Paleoenvironmental reconstructions show that the terminal Pleistocene environment of northeastern North America was similar (though not identical) to those of modern high latitudes (Adovasio and Carr Reference Adovasio, Carr, Camps and Chauhan2009; Shuman et al. Reference Shuman, Newby and Donnelly2009) supporting populations of caribou (Rangifer sp.) that were hunted during the FPP (Robinson Reference Robinson and Chapdelaine2012). Although the applicability of ethnographic models derived from contemporary caribou hunters to these problems is contested (e.g., Levine Reference Levine, Jackson and Thacker1997), caribou hunting remains the most parsimonious explanation for many behaviors inferred from the FPP's archaeology (Ellis Reference Ellis2011). If any groups employing fluted-point technology are to be taken as specialized big game hunters, it is those of the Northeast. The role of plant foods among these groups is therefore important to the continental debate over the interpretation of plant remains from FPP hearths.
FPP Feature Taphonomy
It is not clear whether the pattern of plant remains from FPP hearths results from behavioral practices or taphonomic bias. Securely associating plant remains with human activities requires that a hearth must be (1) visible during excavation and (2) sampled via flotation. At FPP sites in the Northeast, macrobotanical recovery procedures varied considerably and are sometimes unreported. We thus restrict our analysis to presence/absence data. Other methods to identify plant remains at archaeological sites (e.g., phytoliths) have rarely been used at FPP sites and are not discussed here.
For charred macrobotanical remains to be securely associated with subsistence activities, they must be recovered from discernible cultural features (e.g., Spiess and Mosher Reference Spiess and Mosher1992). Hearths are the primary source of macrobotanical remains, because other potential features, such as storage pits, have not been identified at Northeast FPP sites. Although macrobotanical remains from hearths can be non-anthropogenic (Gramly and Funk Reference Gramly and Funk1990), if any plant remains are evidence of FPP subsistence practices, it is those found carbonized in hearths.
Features, like all archaeological remains, are subject to taphonomic bias: older materials are less common than younger materials, even if originally produced in equal numbers (Bluhm and Surovell Reference Bluhm and Surovell2019). If features are rendered visually indiscernible from the surrounding soil more rapidly than nonfeature charcoal, there should be a greater disparity between radiocarbon date frequencies from nonfeature contexts compared to those from features as sites get older. Note that we are concerned here with the characteristics that make features visually discernible, not only the carbon contained therein. If a feature is unidentifiable during excavation, then charred plant remains may be unattributable to subsistence. Alternatively, if taphonomic processes affect feature visibility and nonfeature carbon in a similar manner, then we should expect similar radiocarbon date frequencies from each context through time.
Are Dated Features Unexpectedly Rare during the FPP?
We used radiocarbon dates from the Northeast to compare the frequency of dates from feature and nonfeature contexts. We gathered dates from all time periods and contexts from the Canadian Archaeological Radiocarbon Database (Martindale et al. Reference Martindale, Morlan, Betts, Blake, Gajewski, Chaput, Mason and Vermeersch2016) and from a compilation project overseen by Robert Kelly. Before analysis we “cleaned” the database to remove spurious dates and assigned dates as “feature” or nonfeature dates from context information (Supplemental Text 1; Supplemental Table 1). Of 3,311 total dates, more than half (n = 1,864) came from features (Figure 2). The presence of more feature than nonfeature dates is expected because carbon from features is preferable for dating. Furthermore, our cleaning criteria removed some decades-old dates on dispersed carbon.

Figure 2. Uncalibrated radiocarbon dates used in this analysis from (a) nonfeature (n = 1,447) and (b) feature (n = 1,864) contexts.
To determine whether features were preferentially destroyed, we used two methods employing summed probability distributions (SPD). The first identifies significant divergences of feature dates by comparing the feature SPD to a null model created by iteratively reshuffling feature and nonfeature ages while accounting for sample size (e.g., Bevan et al. Reference Bevan, Colledge, Fuller, Fyfe, Shennan and Stevens2017; Crema et al. Reference Crema, Habu, Kobayashi and Madella2016). This permutation-based method tests whether the feature and nonfeature dates are from the same statistical population (permTest in rcarbon v1.4.1; Crema and Bevan Reference Crema and Bevan2021; Crema et al. Reference Crema, Habu, Kobayashi and Madella2016). The second method compares the feature and nonfeature SPDs themselves, accounting for taphonomic processes (Bluhm and Surovell Reference Bluhm and Surovell2019).
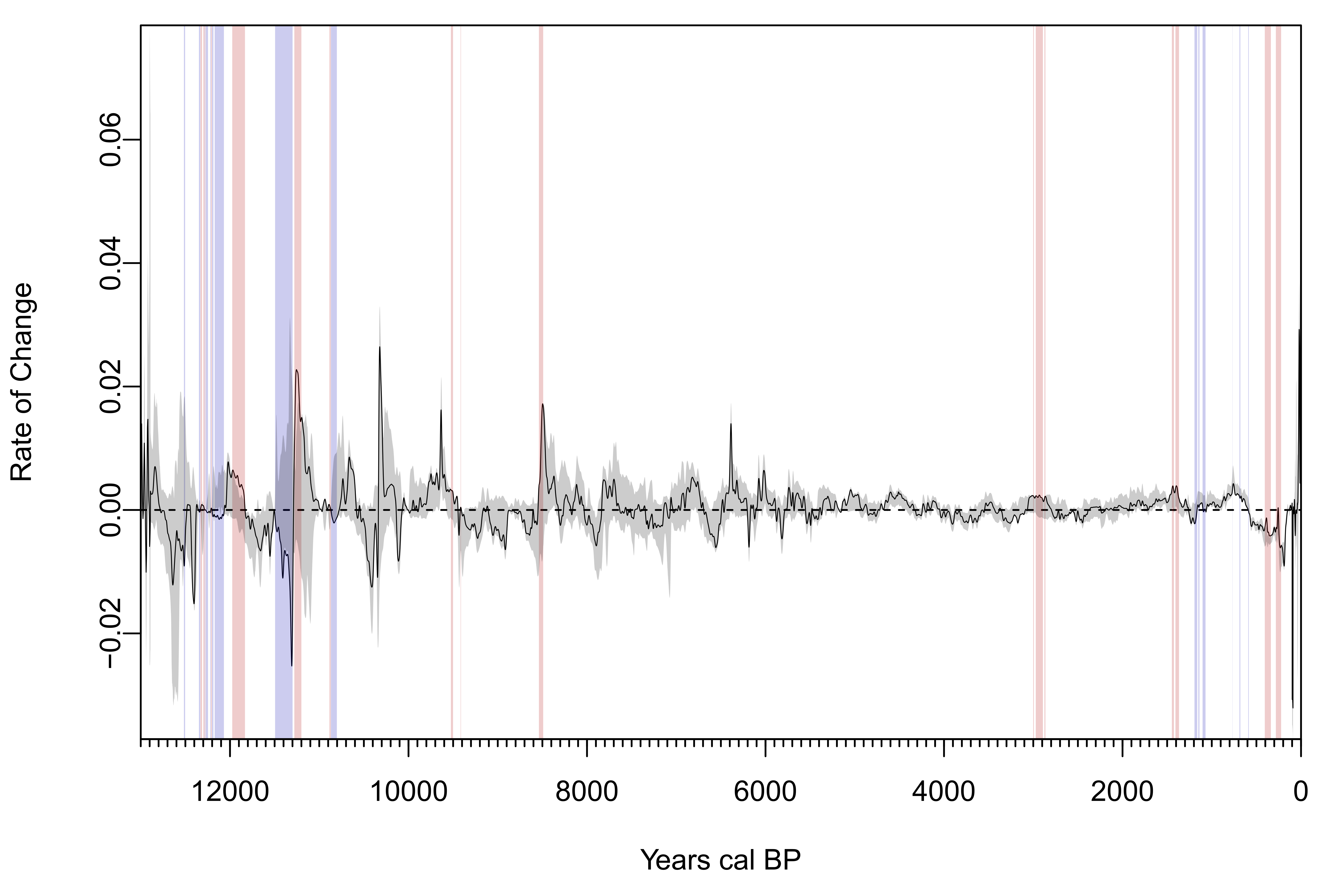
The feature SPD and the null model track each other, converging more frequently with age (Figure 3a). Nonetheless, although the feature SPD has statistically similar growth rates to the null model (rate of change p = 0.092; Supplemental Figure 1), it is significantly distinct overall (global p = 0.0002; Figure 3a). This difference is driven by divergences between the SPD and null model at ~250–990 cal BP, 1220–1340 cal BP, 2650–2920 cal BP, 3240–5030 cal BP, and 5930–5940 cal BP. Prior to 5030 cal BP we see convergence of feature and null model dates until ~9700 cal BP, before which feature dates are intermittently less frequent than expected at four brief time periods. The underrepresentation of feature dates during these oldest periods likely results from the small sample size of dates from features and nonfeature contexts during this time (Figure 2). Feature dates and null model dates do not differ significantly during the FPP.

Figure 3. Summed probability distributions of feature and nonfeature dates from the Northeast with 200-year smoothing and binned by site in 100-year intervals (feature ndates = 1,864, nsites = 565, nbins = 1,307; nonfeature ndates = 1,447, nsites = 493, nbins = 1,017). (a) SPD of feature dates compared to a 95% confidence interval of nonfeature dates; produced with 5,000 simulations of permTest function. (b) SPDs of feature and nonfeature dates taphonomically corrected (Bluhm and Surovell Reference Bluhm and Surovell2019). All dates were calibrated and summed in the rcarbon package v.1.4.1 (Crema and Bevan Reference Crema and Bevan2021) using the IntCal20 calibration curve (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey and Butzin2020) in R v.4.0.2 (R Core Team 2020).
The taphonomically corrected SPDs show a similar distribution (Figure 3b), with the differences between the feature dates and nonfeature dates during the Archaic (~3200–5000 cal BP) being particularly pronounced. We interpret the overall similarity of the feature and nonfeature dates as indicating that features are not preferentially destroyed compared to carbonized material within archaeological strata, although this initial test would benefit from higher resolution datasets (e.g., Edinborough et al. Reference Edinborough, Porčić, Martindale, Brown, Supernant and Ames2017). This analysis indicates that the small number of features (pre-6000 cal BP) in the record is not a reflection of feature-specific taphonomic bias. Therefore, we expect the record of plant materials from hearths during the FPP to be as representative of subsistence as any other archaeological materials from the same period.
Comparison to More Plant-Intensive Subsistence Systems
Although the sample of Northeast FPP hearths is small, it is no smaller than expected given the low number of dated sites in the region (Figure 1; Tables 1 and 2). These data suggest that the number of FPP hearths will always remain low, underscoring the need to draw conclusions from the extant sample of FPP hearths. As noted, 6 of 11 (55%) Northeast FPP features sampled via flotation contain carbonized plant remains, even though many (e.g., berries) may be accidental introductions (Walker et al. Reference Walker, Detwiler, Meeks and Driskell2001:174). To contextualize this information, however, it is instructive to compare them to a sample of hunter-gatherer hearths with good preservation and where small seeds are expected.
Table 2. List of Fluted-Point Sites with Dates from Nonfeature Contexts.

Although several early Holocene sites with multiple features, such as Barton Gulch (Armstrong Reference Armstrong1993) or Dust Cave (Walker et al. Reference Walker, Detwiler, Meeks and Driskell2001), do exist, a sample drawn from single sites is subject to bias by location-specific tasks, seasonal activities, or exceptional preservation. A larger sample prevents a single site from substantially biasing seed presence/absence information. Therefore, for the comparative sample we compiled a database of floated hearths from southwest Wyoming (Supplemental Table 2), a region that offers a searchable compliance database, the known use of seeds for subsistence, and good preservation due to aridity. The dataset includes the presence/absence of carbonized seeds from floated hearths found at sites containing metates from Lincoln, Sweetwater, and Uinta Counties (for methods, see Supplemental Text 1). We assume seeds were not processed by fire at these sites but were accidentally carbonized.
Of the 206 floated hearths sampled, 86 (37%) contained charred seeds, and only 11 (13%) contained more than five seeds each. These data suggest that, even where preservation is good and plants were an important part of diet, hearths often do not contain abundant macrobotanical materials. The results show that the sample of floated FPP hearths in the Northeast contain macrobotanical remains at a higher rate than those from Wyoming. This was surprising given that Wyoming hearths were expected to contain macrobotanical materials in greater abundance and more frequently than in Northeast FPP hearths. These results suggest that plant macroremains in Northeast FPP hearths are quite common.
Conclusion
Although feature dates are scarce during the FPP, they are no less common than expected given the small number of dates from this period. Additionally, the frequency of carbonized seeds in Northeastern FPP hearths is higher than at a time and place where preservation was better and where small seeds were important to the diet. In other words, although small, our sample of FPP macrobotanical remains is not substantially biased by taphonomic processes.
Although this finding might indicate that plants were fundamental to the diet during the Northeast FPP, the repeated presence of berries at these sites indicates that plant gathering was incidental to other concerns (Spiess et al. Reference Spiess, Wilson and Bradley1998:224). The scarcity of small seeds and the absence of grinding stones indicate that low-return-rate, high-processing-cost plant foods were not substantial components of the Northeast FPP diet (Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015). Rather, Northeast FPP groups focused on plant foods requiring no substantial technological investment (sensu Bettinger et al. Reference Bettinger, Winterhalder and McEleath2006) and providing essential nutrients (like vitamin C), such as berries.
Although plant foods augmented hunted foods (Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015), the evidence does not support intensive exploitation of technologically expensive, low-return-rate plant foods (see also Gingerich Reference Gingerich2011:139–140; Hofman and Todd Reference Hofman, Todd, Gerlach and Murray2001:204–205). This finding fits well with an interpretation of Northeast FPP groups as highly mobile hunter-gatherers (e.g., Ellis Reference Ellis2011), who offset local resource depression through mobility rather than dietary expansion and accompanying technological intensification. Ultimately, the plant foods most likely to have been exploited during the FPP are nutritionally important fleshy fruits, hickory nuts, and geophytes such as cattail (Typha sp.) rhizomes with high-caloric return rates (Figure 4; Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015). Although parenchymatous tissues are infrequently preserved in open-air hearths, efforts to identify the use of geophytes at FPP sites offers an additional test of this hypothesis.

Figure 4. Post-encounter return rates for select plant foods found in the Northeast. Black bars indicate geophytes. Some species, particularly hickory, were likely absent from the northern portion of the region during the Pleistocene (adapted from Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015).
The use of high-return-rate, low-processing-cost plant foods during this period does not mean these groups were unfamiliar with the utility of other plant resources for food or other purposes, but that those higher-cost resources were not regularly or intensively used under most circumstances. It is not necessary to invoke the habitual use of low-return-rate resources as a risk mitigation mechanism when maintenance of environmental knowledge fulfills this role (see Hofman and Todd Reference Hofman, Todd, Gerlach and Murray2001:201).
The overall caloric contributions of plants, particularly fleshy fruits during the FPP, were likely far smaller than that of meat, given a post-encounter return rate of caribou (Rangifer tarandus) that is nearly 4 times (25,370 kcal/hr; Smith Reference Smith1991) that of the most calorically dense plant food, winter cattail (7,000 kcal/hr; Madsen et al. Reference Madsen, Eschler and Eschler1997), and 18 times that of the hawthorn fruits found at Shawnee-Minisink (1,415 kcal/hr; Reidhead Reference Reidhead1981). Plant foods were likely periodically important to the FPP diet when meat was scarce and as a source of essential nutrients such as vitamin C. Still, our sample, though small, does not point to the systematic and intensive use of low-return-rate plant foods during the Northeast FPP. Instead, caribou hunting was most central and most likely structured many aspects of life at this time (Ellis Reference Ellis2011).
Acknowledgments
We thank Robert Kelly for pushing us to publish this research and for providing comments on various drafts of this article. Dr. Maria Fernanda (Mafe) Boza Cuadros aided with the translation of the abstract to Spanish. NK extends thanks to Richard (Dick) Doyle, who contributed to ongoing archaeological projects in northern New England but whose efforts were unacknowledged elsewhere. We also thank the anonymous reviewers for their helpful comments; their efforts substantially improved the final product. MM's work was conducted at the University of Wyoming. No permit was required for the analyses presented here.
Data Availability Statement
The radiocarbon database is included as Supplemental Table 1 and is available through the Canadian Archaeological Radiocarbon Database. The Wyoming data are available in published sources or from the Wyoming State Historic Preservation Office.
Supplemental Material
For supplemental material accompanying this article, visit https://doi.org/10.1017/aaq.2021.125.
Supplemental Text 1. Explanation of Methods for Radiocarbon and Comparative Hearth Analysis.
Supplemental Table 1. Cleaned Northeast Radiocarbon Database Used for Feature and Nonfeature Analysis.
Supplemental Table 2. Floated Features from Archaeological Sites in Wyoming Used in this Analysis.
Supplemental Figure 1. Rate of change analysis for feature permutation test using the same dataset and methods detailed in Figure 3.