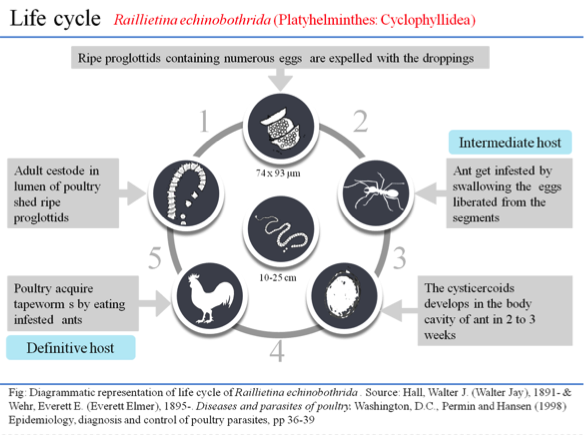
INTRODUCTION
Eucestodes fulfil their energy requirements essentially by metabolizing the glucose and other simple carbohydrate molecules, and in these parasites phosphoenolpyruvate carboxykinase (PEPCK) plays a significant role in glucose oxidation (Bryant, Reference Bryant1975; Roberts, Reference Roberts, Arme and Pappas1983; Bryant and Behm, Reference Bryant and Behm1989; Smyth and McManus, Reference Smyth and McManus1989). Understanding the metabolic processes and mechanisms by which the parasites derive energy for their survival is of prime importance in biomedicine and parasitism management. PEPCK catalyses the carboxylation step, i.e. production of oxaloacetate (OAA) from phosphoenolpyruvate (PEP), in eucestodes including Raillietina echinobothrida as well as in other helminths (Prichard and Schofield, Reference Prichard and Schofield1968; Saz and Lescure, Reference Saz and Lescure1969; Tandon and Das, Reference Tandon and Das2007). However, in higher vertebrates (including its definitive host, G. domesticus) it is functionally involved in gluconeogenesis (Nelson and Cox, Reference Nelson and Cox2012). There are two isoforms of PEPCK, mitochondrial PEPCK (PEPCK-M) and cytosolic PEPCK (PEPCK-C), found in animals, coded by different genes and expressed in very early stage of life. For the first time, PEPCK-M was reported by Utter and Kurahashi (Reference Utter and Kurahashi1953) in chicken liver mitochondria; whereas, PEPCK-C by Nordlie and Lardy (Reference Nordlie and Lardy1963) in rat tissues. PEPCK-C appears at birth in the livers of mammals for maintenance of glucose homeostasis (Ballard and Hanson, Reference Ballard and Hanson1967), while PEPCK- M gene is present in the liver before birth (Arinze, Reference Arinze1975).
Considering the importance of the enzyme in energy derivation for survival of the cestode parasites, inhibition of PEPCK might down-regulate the utilisation of glucose in the cestode parasites, which may help for parasitism management. The PEPCK/PK branch point in eucestodes is the first divergent step between metabolic pathways of the parasites and their vertebrate hosts and has been studied in various helminths (Prichard, Reference Prichard1976; Moon et al. Reference Moon, Mustafa, Hulbert, Podesta and Metrrik1977; Hoffmann et al. Reference Hoffmann, Mustafa and Jorgensen1979; Das et al. Reference Das, Ramnath, Dutta and Tandon2015). Owing to the differing primary functions of PEPCK in helminths and their hosts, this step provides an avenue for anthelmintic target (Reynolds, Reference Reynolds1980). In order to characterize this important enzyme, PEPCK has been biochemically purified and characterized from few helminths species including R. echinobothrida (Behm and Bryant, Reference Behm and Bryant1975a , Reference Behm and Bryant1982; Cornish et al. Reference Cornish, Wilkes and Mettrick1981a ; Wilkes et al. Reference Wilkes, Cornish and Mettrick1981, Reference Wilkes, Cornish and Mettrick1982; Rohrer et al. Reference Rohrer, Saz and Nowak1986; Das et al. Reference Das, Tandon, Saxena, Joshi and Singh2013). However, the copy number of PEPCK is not known in these helminths, but PEPCK(GTP) found in R. echinobothrida is generally present as a single copy number in all organisms known so far. Recently, PEPCK gene from the cestode parasite, R. echinobothrida, has been deduced and submitted to NCBI GenBank (accession no. KC252609.1) (Dutta et al. Reference Dutta, Ramnath, Tandon and Das2016a ).
Phytoestrogens (genistein and daidzein) isolated from the root-peel of Flemingia vestita (a medicinal plant found in North Eastern India) has vermifugal action against several intestinal cestodes including R. echinobothrida and trematodes (Tandon et al. Reference Tandon, Pal, Roy, Rao and Reddy1997) and have been reported to alter PEPCK activity in R. echinobothrida when treated in vitro (Das et al. Reference Das, Tandon and Saha2004a , Reference Das, Tandon and Saha b ; Tandon and Das, Reference Tandon and Das2007). In vitro treatment with genistein and its derivatives (Rm6423 and Rm6426) induces morphological and structural alterations leading to impair viability in Echinococcus multilocularis and E. granulosus metacestodes (Naguleswaran et al. Reference Naguleswaran, Spicher, Vonlaufen, Ortega-Mora, Torgerson, Gottstein and Hempill2006). Choi et al. (Reference Choi, Jung, Yeo, Kim and Lee2008) demonstrate that genistein and daidzein decrease blood glucose by lowering PEPCK activity in non-obese diabetic mice. Recently, biocomputational studies by Katiyar et al. (Reference Katiyar, Jain, Dhanjal and Sundar2015) reveal the mixed inhibition of human PEPCK-C by genistein. PEPCK-C has been found to alter by genistein in some cancerous cells by mediating DNA promoter methylation and RNA stability (Seenappa et al. Reference Seenappa, Das, Joshi and Satyamoorthy2016).
Despite considerable advances in understanding the kinetics of PEPCK from several sources, the nature, mechanism and mode of action correlating the parasite and its host are largely unknown. Hence, with the aim of ascertaining the role of PEPCK as a likely target site of anthelmintic drug action, the intent of the present study was to find out the biological significance of PEPCK in the parasite and to ascertain a comparative functional differences, if any, between PEPCKs from the cestode parasite R. echinobothrida (RePEPCK), and its host, Gallus domesticus (GdPEPCK). In search of possible modulators for the enzyme, phytoestrogens were also analysed on recombinant proteins from both sources.
MATERIALS AND METHODS
Chemicals
All chemicals used in this study were of highest analytical grade and purchased from Sigma (St Louis, USA) and Sisco Research Limited (SRL, India). Restriction enzymes and other enzymes were procured from New England Biolabs Ltd (UK).
The parasite
The cestode parasite, R. echinobothrida, was collected from the small intestine of naturally infected chicken (G. domesticus), and morphologically identified by the presence of a heavily armed rostellum with two rows of hooklets and four circular suckers on the scolex (Soulsby, Reference Soulsby1982). The segregated live parasites were washed in PBS (pH 7.4) and maintained in PBS (pH 7.4) at 37 °C prior to experiment.
siRNA treatment
siRNA against RePEPCK (NCBI GenBank accession no. KC252609.1) was commercially synthesized (Qiagen), and using soaking method (Geldhof et al. Reference Geldhof, Visser, Clark, Saunders, Britton, Gilleard, Berriman and Knox2007), different concentrations of siRNA (5, 25 and 50 nm) were added to the freshly collected live parasites along with positive- and negative- control. For each set of siRNA experiment, 2 numbers of live parasites (about 0.2 g wet weight, roughly same size and developmental stage) were incubated with siRNA and 5 mm D-glucose in a 6-well plate at 37 °C and three set of biological replicates were performed for each siRNA experiment. The positive control siRNA mix (50 nm of Premix FlexiTube siRNA Ctrl All stars II) targets some vital genes necessary for survival of an organism; whereas, the negative control mix contains the transfection reagent without siRNA against RePEPCK. The parasites were incubated at 37 °C, and onset of paralysis, i.e. absence of motility upon physical disturbance, in the parasites was monitored by visually examining them every 15 min. Their survival time due to siRNA was also monitored (data not shown). At the paralysis time, the parasites were collected and processed for analysis of RePEPCK mRNA expression using Real Time PCR and RePEPCK protein expression by western blot.
β-actin from R. echinobothrida as an endogenous control
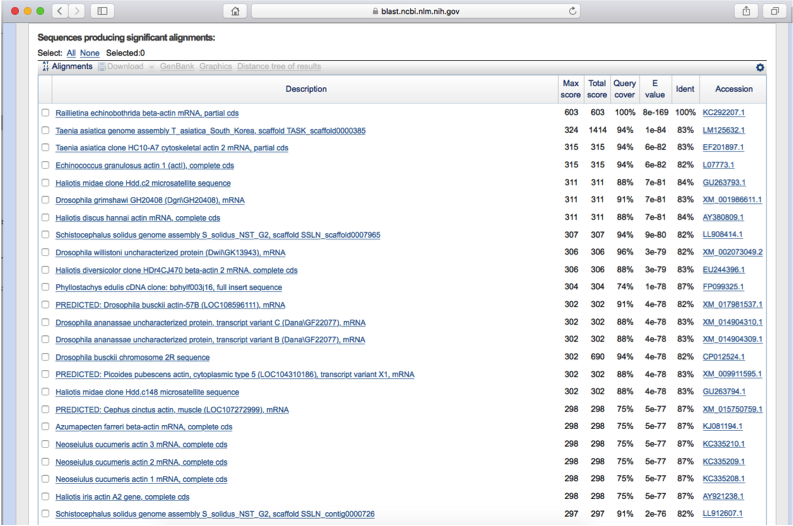
The housekeeping gene, β-actin, from R. echinobothrida was used as endogenous control for Real Time PCR for relative quantification of the expression of RePEPCK mRNA. β-actin gene sequence of R. echinobothrida was obtained by PCR amplification using degenerate primers (FP: GCTCGTCGTCGACAACGGCTC; and RP: CAAACATGATCTGGGTCATCTTCTC) designed from the conserved regions of the complete cDNA sequence of Homo sapiens β-actin gene (NCBI GenBank accession no. BC002409.2). In brief, total RNA was isolated from the parasite using Purelink RNA mini kit (Thermo Fisher Scientific) following manufacturers’ protocol. RNA was then quantified using Nanodrop 2000 (Thermo Scientific) and its integrity was checked on 1.2% agarose gel. 2 µg of total RNA from R. echinobothrida was taken, and the first strand synthesis (cDNA) was performed by using the Super Script III first strand reverse transcriptase polymerase chain reaction (RT-PCR) reaction kit (Thermo Fisher Scientific) following manufacturers’ protocol. Using cDNA as template, PCR was performed with the following cycle parameters: initial denaturation temperature of 95 °C for 5 min; 35 cycles of 95 °C for 30 s, 55 °C for 45 s and 72 °C for 1 min. 1x PCR reaction buffer contained 10 mm Tris-HCl (pH 8.3), 50 mm KCl, 1.5 mm MgCl2, 200 µ m dNTPs and 200 nm of each primer. The PCR product (334 bp amplicon; Fig. S1) was gel eluted and purified using Sigma GenElute gel extraction kit following manufacturer's instructions. The purified product was cloned into pGEM-T easy vector, sequenced and submitted to NCBI GenBank (accession no KC292207.1). The obtained sequence (partial CDS) was translated conceptually using ExPASy-Translate tool (http://web.expasy.org/translate/) and aligned against β-actin amino acid sequences from different species using Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo) (data not shown). Primers for β-actin from R. echinobothrida were designed (Table S1) from the obtained partial CDS of β-actin, which served as endogenous control for Real Time PCR.
Real Time PCR for analysis of expression of RePEPCK mRNA
For relative quantification of the expression of RePEPCK mRNA level, total RNA was isolated from the parasites and cDNA was synthesized as described above. Using cDNA as template, the relative expression of RePEPCK mRNA level in the siRNA treatment groups with respect to their respective controls was assessed by Real Time PCR on a 7500 Fast Real Time PCR System (Applied Biosystems) using the gene specific primers (Table S1). Real Time PCR was performed in a 20 µL final volume using Power SYBR Green master mix and the primers specific for RePEPCK. The reaction was carried out in triplicates for each sample in optical 96-well reaction plates using the following PCR conditions (predenaturation 95 °C for 2 min, 40 cycles of 95 °C for 15 s, 60 °C for 30 s) and relative RePEPCK mRNA levels were determined using the comparative threshold cycle method. Data were normalized using β-actin from the parasite and analysed using 7500 Fast system SDS software (Version 1.4).
Western blot analysis of RePEPCK protein
For expression of RePEPCK protein, western blot was performed using standard protocol. Isolated protein samples were resolved on SDS-10% (w/v) PAGE and electroblotted onto polyvinylidene difluoride membrane, the membrane was blocked for 1 h at room temperature in TBS [10 mm Tris-HCl (pH 7.4), 0.5 M NaCl] plus Tween 20 (0.2% v/v) containing 5% (w/v) nonfat dried milk and then incubated with anti-PEPCK-C antibody (Code no. sc-74823; Santa Cruz Biotechnology, U.S.A.) overnight at 4 °C. After washing, the membrane was incubated with the corresponding goat anti-rabbit IgG-HRP secondary antibody (Code no. sc-2004; Santa Cruz Biotechnology, USA) for 2 h. The membrane was incubated in Amersham™ ECL™ Prime western blotting detection reagent for 3 min, and the protein bands were captured using chemidoc (ImageQuant LAS 500, GE Healthcare Life Sciences).
Cloning, over-expression and purification of RePEPCK
To sub-clone RePEPCK into pE-SUMO vector (kindly provided by Dr Todd Holyoak, University of Waterloo, Canada), the ORF (1887 bp) of RePEPCK (NCBI GenBank accession no. KC252609.1) was amplified from pGEM-PEPCK construct, originally cloned in our laboratory, using the forward primer having a BsmB1 cut site (5′-AACACGTCTCAAGGTATGTCGCCACTCTTTAGG-3′) and the reverse primer having a Xho1 cut site (5′- GGCGCTCGAGTTACATTGCTTTAATGCG-3′). PCR was performed using 50 ng of DNA with the following cycle parameters: initial denaturation temperature of 95 °C for 60 s, 35 cycles of 95 °C for 30 s, 62 °C for 30 s and 72 °C for 2 min followed by a final extension of 72 °C for 10 min. The 1.8 kb RePEPCK amplicon was gel eluted and purified using GenElute™ gel extraction kit (Sigma NA1111) following manufacturers’ instructions.
The pE-SUMO vector was digested with Bsa1 and Xho1 in 1x CutSmart buffer for 2 h at 37 °C, dephosphorylated by alkaline phosphatase and purified using PCR purification kit. The insert was also digested with BsmB1 and Xho1 and purified. The digested insert and pE-SUMO vector was ligated at 3:1 molar ratio using DNA ligase. The RePEPCK sequence and orientation was confirmed by colony PCR, double digestion and direct sequencing, and the construct was transformed into Escherichia coli cells (BL21 DE3).
A single colony was picked up from the LB agar plate and inoculated in 5 mL LB broth containing 50 mg mL−1 kanamycin. It was incubated at 37 °C with continuous shaking (220 rpm) for overnight. Next day, 1 mL of this primary culture was inoculated in l00 mL culture and incubated at 37 °C with shaking until the OD600 reached about 0.8 and then induced with 0.5 mm IPTG at 37 °C for 8 h. The cells were harvested by centrifugation at 6000 rpm for 15 min and resuspended in lysis buffer (50 mm Tris-HCl, pH 7.4 containing 1 mm DTT, 10% Glycerol, 150 mm NaCl and 0.1 mm EDTA), sonicated and analysed by SDS-10% (w/v) PAGE. The recombinant His-tagged RePEPCK was then purified using Ni2+-nitrilotriacetate (Ni-NTA) agarose, protein was eluted at 300 mm imidazole and the N-terminal fusion part was removed by using SUMO protease (1 U µL−1) overnight at 4 °C. The protein (RePEPCK) was characterized using enzyme kinetics.
Cloning, over-expression and purification of GdPEPCK
Similarly, the ORF of GdPEPCK was also cloned into pE-SUMO vector and over-expressed. In brief, RNA was isolated from the liver of G. domesticus and cDNA was synthesized as described above. The ORF of G. domesticus cytosolic PEPCK (NCBI GenBank accession no. NM_205471.1) was cloned into pE-SUMO vector using the forward primer having a Sma1 cut site (5′-ATCCCGGGATGGCCCCAGAGCTGAAAAC-3′) and the reverse primer having a Xho1 cut site (5′-CGGTCTCGAGCTACAACTGTTTTATTCTCA-3′). Using the following PCR conditions (predenaturation 95 °C for 90 s, 35 cycles of 95 °C for 30 s, 63 °C for 30 s and 72 °C for 2 min followed by a final extension of 72 °C for 10 min), PCR was performed taking 50 ng of DNA as template. The GdPEPCK amplicon was gel eluted, purified and ligated. The construct (pE-SUMO-GdPEPCK) was then transformed into host cells (BL21 DE3) and over-expressed. The recombinant His-tagged protein was then purified using Ni-NTA agarose, eluted at 300 mm imidazole. After removal of the N-terminal fusion part, the protein was characterized using enzyme kinetics.
Biochemical characterization of RePEPCK and GdPEPCK
RePEPCK kinetics was assayed spectrophotometrically (Cary 60 UV-Vis, Agilent Technologies; USA) at 40 °C as described by Dutta et al. (Reference Dutta, Ramnath, Tandon and Das2016a ). The standard assay mixture of 1.5 mL for PEP carboxylation reaction contained 50 mm Tris-HCl buffer (pH 7·4), 5 mm PEP, 0·15 mm NADH, 0·5 mm GDP, 20 mm NaHCO3, 4 mm MnCl2 in addition 10 units of MDH. GdPEPCK carboxylation reaction was assayed as described by Colombo et al. (Reference Colombo, Carlson and Lardy1978); 1.5 mL of the reaction mixture contained 50 mm Tris-HCl buffer (pH 7·4), 8 mm PEP, 0·15 mm NADH, 0·6 mm GDP, 8 mm NaHCO3, 0.5 mm MnCl2 in addition 10 U of MDH. For RePEPCK decarboxylation assay, the modified method of Johnson and Holyoak (Reference Johnson and Holyoak2012) was followed; 1.5 mL of reaction mixture contained HEPES buffer (0.05 M, pH 7.6), OAA (0.35 mm), GTP (0.45 mm), ADP (1 mm), MgCl2 (6 mm), MnCl2 (0.05 mm), LDH (10 U), NADH (0.15 mm) and PK (5U). GdPEPCK decarboxylation activity was assayed using modified method of Noce and Utter (Reference Noce and Utter1975). For this assay, the reaction mixture (in a volume of 1.5 mL) contained HEPES buffer (0.05 m, pH 7.6), OAA (0.5 mm), GTP (0.5 mm), ADP (1 mm), MgCl2 (8 mm), MnCl2 (0.05 mm), LDH (10 U), NADH (0.15 mm), PK (5U). After 5 min pre-incubation, the above reactions were initiated by the addition of an appropriate volume of the recombinant protein in the reaction mixture.
The background and negative controls were maintained without NADH and the enzyme source, respectively. Protein content in the sample was determined using Bradford's reagent (Bradford, Reference Bradford1976), and bovine serum albumin was taken as the standard. Specific activity was expressed as U/mg of protein. One U of enzyme activity is defined as the amount of the enzyme that catalyses the oxidation of 1 µ m of NADH per min under standard assay conditions.
Several buffers (pH ranges from 5.2 to 9.6) were used to find out the optimal PEPCK activity. Apparent Michaelis-Menten constants (K mapp) for the substrates and co-factors; and inhibitory constants (K i) for the phytoestrogens (genistein and daidzein) were determined for carboxylation as well as decarboxylation reactions. K mapp and K i were calculated by plotting the reaction rates at varying concentrations of the substrate or co-factors (Michaelis-Menten and Lineweaver-Burk plots). The effect of these compounds on the enzyme activity was studied at three different concentrations (5, 10 and 20 µ m).
CD spectra analysis
The effect of the phytoestrogens on conformation of the purified recombinant proteins was analysed with and without the phytoestrogens using CD spectrophotometer (JASCO-815) following the method described by Dutta et al. (Reference Dutta, Ramnath, Dkhar, Tandon and Das2016b ).
Molecular docking
In order to find out the possible interactions of the phytoestrogens with PEPCK, molecular docking was performed essentially following Dutta et al. (Reference Dutta, Ramnath, Tandon and Das2016a ). Molecular docking of the possible modulators into the active site of PEPCK of the parasite and its host was performed. For host PEPCK, the available crystal structure of chicken mitochondrial PEPCK structure (PDB ID 2QZY) was selected. However, for parasite PEPCK, the homology model described by Dutta et al. (Reference Dutta, Ramnath, Dkhar, Tandon and Das2016b ) was used.
Biological significance of lead molecules
A comparative dosage- and time-dependent experiment was performed to find out the biological significance of genistein and tetrahydropalmatine (THP) and their effect on RePEPCK activity following the methods described by Dutta et al. (Reference Dutta, Ramnath, Tandon and Das2016a ). In brief, different concentrations (0.25–1.0 mm) of phytoestrogens or THP were added to each set of treatment with simultaneous maintenance of their corresponding controls. The treated parasites were also monitored for onset of paralysis in them as mentioned above (data not shown). At the onset of paralysis, the parasites were collected and a 10% homogenate was made from the treated parasites and their respective controls. RePEPCK activity in the tissue homogenate was assayed spectrophotometrically at 40 °C in the direction of PEP carboxylation following the method as mentioned in the preceding section above.
Statistical analysis
Data were represented as the mean ± s.em. (n = 3) or mentioned otherwise and probability values less than 0·05 were taken to be statistically significant. Co-efficient of bivariate regression line was calculated using OriginPro 8 software and the co-efficient for linear lines was not less than 0.9 (R 2 ⩾ 0·9).
RESULTS
siRNA induces paralysis in the parasite and inhibits RePEPCK expression
Biological significance of PEPCK in the parasite was anlysed using siRNA. siRNA caused onset of paralysis in the parasite at 28.0 ± 0.5 h, 13.0 ± 1.3 h and 5.0 ± 0.7 h with 5, 25 and 50 nm concentration of siRNA, respectively; however, onset of paralysis in the parasites occurred at 92.0 ± 1.2 h without siRNA (negative control). On the other hand, the parasites treated with positive control siRNA were paralysed at 3.0 ± 0.3 h (Table 1). At the onset of paralysis in the parasites, silencing of PEPCK gene by siRNA resulted in reduction of PEPCK mRNA level (Fig. 1A) and PEPCK protein (Fig. 1B). In corroboration, PEPCK kinetics also decreased at onset of paralysis in the siRNA treated parasites (Table 1).

Fig. 1. (A) RePEPCK mRNA levels in siRNA treated parasites were measured by Real Time PCR with respect to controls. Data were collected from 3 biological replicates and represented as mean ± s.e.m. (B) Western blot analysis of RePEPCK protein at the time of paralysis in R. echinobothrida treated with siRNA. Lane 1, negative control; lanes 2–4, represent PEPCK expression level at different concentrations (5, 25 and 50 nm) of siRNA; β-actin was used as loading control. (C) Differential specific activities of RePEPCK and GdPEPCK. Values were taken from 3 separate experiments and expressed as mean ± s.e.m. (D, E) Effect of different concentrations (1– 50 mm) of NaHCO3 − on RePEPCK- (D) and GdPEPCK- activity (E). Data were taken from 3 separate experiments and expressed as mean ± s.e.m. PEPCK, phosphoenolpyruvate carboxykinase.
Table 1. Effect of siRNAs on onset of paralysis and PEPCK activity in the cestode parasite, R. echinobothrida

Values are expressed as mean ± s.e.m. (n = 3).
a The negative control mix (50 nm of Premix FlexiTube siRNA Control All stars 1) contains the transfection reagent without siRNA against RePEPCK.
b The positive control siRNA mix (50 nm of Premix FlexiTube siRNA Ctrl All stars II) targets some vital genes necessary for survival of an organism.
*Signifies statistical significance in siRNA treated parasites compared with −ve control with p value of <0.001.
Biochemical characterization of RePEPCK and GdPEPCK
In order to find out functional differences and in search of probable inhibitors, PEPCK from both the parasite and its host was overexpressed, purified and characterized. The ORF of RePEPCK was cloned into pE-SUMO vector and the positive clones were confirmed by PCR (Fig. S2A). The induced recombinant protein was purified by using Ni-NTA agarose, and the protein gave about 70 kDa band on SDS-10% (w/v) PAGE (Fig. S2B).
The kinetic properties of recombinant enzymes were compared with those of native enzymes and summarized in Table S2, notably decarboxylation reaction was absent in the parasite (Fig. 1C). Tris-HCl buffer (50 mm) was found to be the ideal buffer for RePEPCK activity (Table S3) and maximal RePEPCK activity was observed at pH 7.4 (Fig. S3A). However, HEPES buffer was chosen as the more appropriate buffer for GdPEPCK decarboxylation reaction as it showed optimal activity at pH 7.6 (Table S3). Several salts of divalent cations were investigated in order to find out suitable divalent cations for the enzyme activity. MnCl2 was found to be appropriate showing 4 and 0.5 mm as the best concentrations for RePEPCK and GdPEPCK activities, respectively, for carboxylation reaction (Table S3; Fig. S3B, C). Different concentrations of exogenous HCO3 − (1–50 mm) were used for determining its optimal concentration for both enzyme activities. 20 and 8 mm of HCO3 − were found to be the appropriate concentration for RePEPCK and GdPEPCK carboxylation reaction, respectively (Fig. 1D, E). Among various nucleoside phosphates tested on RePEPCK and GdPEPCK activities in the carboxylation direction, GDP was found to be the suitable nucleoside di-phosphate (Table S4). 0.5 and 0.6 mm concentrations of GDP were found to be the appropriate concentrations for RePEPCK and GdPEPCK, respectively (Fig. S3D). In order to find out the appropriate co-factors, several nucleoside di-phsosphates and tri-phsosphates were used. ADP and GTP exhibited the maximal GdPEPCK activity in decarboxylation reaction, respectively. 1 and 0.5 mm were found to be the optimal concentrations for ADP and GTP, respectively, for GdPEPCK activity (data not shown). To fulfil the dual cation requirement for decarboxylation reaction (Lee et al. Reference Lee, Hebda and Nowak1981), several divalent cations were examined at constant MnCl2 (0.02 mm), MgCl2 (5.8 mm) was found to be the best cation; and similarly, at constant MgCl2 (5.8 mm), MnCl2 (0.02 mm) was found to be suitable cation (Table S3). Of different concentrations of MnCl2 (0.01–0.5 mm), 0.05 mm was found to be suitable for GdPEPCK optimal activity for decarboxylation reaction. MgCl2 at 8 mm was found to fulfil the requirement of the second divalent cation for GdPEPCK activity for decarboxylation reaction (Table S3).
In order to find out the influence of the substrate on RePEPCK and GdPEPCK activities, various concentrations of PEP (0.01–8 mm) were used. In the case of RePEPCK, a negative co-operative interaction was observed beyond 5 mm of PEP concentration. A similar pattern of kinetic behavior was also observed for GdPEPCK carboxylation reaction beyond 10 mm PEP. The V max for RePEPCK was found to be 23 U mg−1 of protein (Fig. S4A); however, for GdPEPCK it was 13 U mg−1 of protein in carboxylation reaction (Fig. S4B). For GdPEPCK decarboxylation reaction, Vmax was recorded as 33 U mg−1 of protein when OAA was used as substrate (Table S3; Fig. S4C). K mapp of RePEPCK and GdPEPCK were observed with K mapp of 46.9 and 22.9 µ m for RePEPCK and GdPEPCK, respectively, in the carboxylation reaction as determined by double reciprocal plot. K mapp for OAA was found to be 15.4 µ m for GdPEPCK for decarboxylation reaction (Table S5). K mapp for other co-factors was determined and is presented in Table S5. A comparative K mapp of recombinant and native PEPCKs from both sources have been summarized in Table S5.
Effect of phytoestrogens on RePEPCK and GdPEPCK activities and their conformation
In search for possible modulators of PEPCK, the phytoestrogens, genistein and daidzein, were tested on RePEPCK and GdPEPCK activities. It was observed that genistein and daidzein inhibited RePEPCK in a non-competitive manner with K i of 0.15 and 0.26 µ m, respectively (Fig. 2A, Fig. S5A). However, the phytoestrogens inhibited GdPEPCK activity in a competitive manner with K i of 0.20 and 0.35 µ m, respectively, in carboxylation reaction (Fig. 2B, Fig. S5B). Notably, genistein and daidzein altered GdPEPCK activity in a competitive manner accompanied with K i of 0.31 and 0.64 µ m, respectively, for decarboxylation reaction (Figs S5C, D). Results of the inhibition study revealed that genistein was more inhibitory than daidzein for RePEPCK and GdPEPCK.

Fig. 2. (A) Lineweaver-Burk plot showing inhibition of RePEPCK activity (carboxylation) by genistein at variable substrate concentrations. The plot was calculated from assays of seven different concentrations of PEP (0.01–8 mm) in variable concentrations of genistein {(*) control, (⚫) 5 µ m, (■) 10 µ m, and (▾) 20 µ m}. The replot of the slope of the lines in the Lineweaver-Burk plot vs. genistein concentrations shows Ki = 0.15 µ m with R 2 = 0.9972 (inset). (B) Lineweaver-Burk plot showing inhibition of GdPEPCK activity (carboxylation) by genistein. The replot shows Ki = 0.20 µ m with R 2 = 0.9990 (inset). (C) Near-UV CD spectra (250–350 nm) of RePEPCK (0.5 mg mL−1) in 10 mm Tris-HCl (pH 7.4) in presence of 20 µ m genistein. The CD panel was monitored by changes in near-UV CD ellipticity. The experiments were performed in the presence 20 µ m of genistein (dotted line) against control (solid line). PEPCK, phosphoenolpyruvate carboxykinase.
Far-UV CD spectra analysis depicted that genistein (20 µ m) (Fig. 2C) and daidzein (20 µ m) (data not shown) had no effect, within experimental errors, on the conformation of RePEPCK, and also on GdPEPCK (data not shown).
Differential binding of phytoestrogens onto RePEPCK and GdPEPCK
The PEPCK active site is relatively large and has a number of specific binding pockets. Out of the modulators tested, genistein had the lowest binding energy (CDocker energy) of 53.20 kcal per mol. Binding sites of genistein lied near the substrate-binding site (Fig. 3A, B). Analysis of the genistein-PEPCK complex revealed that the ligand bound to the R89 (critical amino acid responsible for PEP binding) via a hydrogen bond. Hydrophobic interactions were also observed with S288 and R407 in the genistein-PEPCK complex. However, daidzein showed no significant interactions with residues of active site of PEPCK. Surface model corresponding to the volume of binding sites were prepared to examine the difference in binding conformations of the host and parasite model in complex with genistein towards PEP (Fig. 3C). It was also predicted that the chicken PEPCK is likely to be a competitive inhibitor for PEP, whereas it is predicted to be non-competitive for parasite PEPCK, which was also observed in enzyme kinetics studies.

Fig. 3. (A) Overview of the predicted interactions between chicken mitochondrial PEPCK and genistein. The protein backbone is rendered as a gray ribbon and dotted lines represent the hydrogen bonds. The catalytic residues and the ligand are shown as thin and thick stick models, respectively. (B) Pharmacophoric overlay of the top poses of genistein upon PEP of the parasite and chicken mitochondrial PEPCK model. Surface structure of genistein and PEP in the substrate-binding site of the host enzyme showing similar pocket. (C) Surface structure of genistein and PEP in the substrate-binding site of the parasite enzyme showing different pocket. PEPCK, phosphoenolpyruvate carboxykinase.
Biological relevance of the modulators
When the parasites exposed to defined concentrations of possible modulators, onset of paralysis ensued in the parasites ~2–8 h of incubation, while the parasites survived up to ~90–96 h without the modulators. Dosage-dependent experiments revealed a significant decrease in RePEPCK activity at more than 0·25 mm of the modulators (Fig. S6A); while time-dependent experiments showed no significant change in the enzyme activity within 1 h of genistein treatment (Fig. S6B); while RePEPCK activity decreased by ~35% at the same time frame in the case of THP.
DISCUSSION
PEPCK has been considered as a potential anthelmintic target for a long time (Reynolds, Reference Reynolds1980); however, the present study provides the first direct report to prove that PEPCK is essentially required for the survival of the parasite. In platyhelminths, siRNA could be used for therapeutic control (Bai et al. Reference Bai, Zhang, Jin, Kang, Zhu, Zhang, Li, Ma, Zhao, Shi, Li, McManus, Zhang and Wang2014); in this study, a functional RNAi pathway by silencing of PEPCK was observed in R. echinobothrida. The first report on the delivery of siRNA in a cestode was in Moniezia expansa by silencing of neuropeptide gene using the soaking method (Pierson et al. Reference Pierson, Mousley, Devine, Marks, Day and Maule2010). As siRNA caused paralysis in R. echinobothrida in the present study, similar obeservations have been observed in other parasites too, e.g. pat-10 gene in Meloidogyne graminicola, cathepsin L and Z-like cathepsin protease in Onchocerca volvulus larvae, and helicase in Brugia malayi (Aboobaker and Blaxter, Reference Aboobaker and Blaxter2003; Lustigman et al. Reference Lustigman, Zhang, Liu, Oksov and Hashmi2004; Ford et al. Reference Ford, Guiliano, Oksov, Debnath, Liu, Williams, Blaxter and Lustigman2005; Nsengimana et al. Reference Nsengimana, Bauters, Haegeman and Gheysen2013; Singh et al. Reference Singh, Kushwaha, Mohd, Pathak and Misra-Bhattacharya2013). Silencing of actin gene of Litosomoides sigmodontis by siRNA also causes paralysis in the parasite within 48–72 h, and releases significantly less microfilariae (Pfarr et al. Reference Pfarr, Heider and Hoerauf2006).
A comparison of RePEPCK and GdPEPCK specific activities for carboxylation reaction provides a clue about the preferred reaction in the parasite and its host, the RePEPCK specific activity being approximately 2-folds higher than the GdPEPCK. Specific activity of GdPEPCK for decarboxylation reaction supports the earlier report that OAA to PEP formation is the main reaction for PEPCK in chicken liver (Noce and Utter, Reference Noce and Utter1975), notably decarboxylation reaction is completely absent in R. echinobothrida. In the present study, 4 mm MnCl2 was found to be suitable for RePEPCK optimal activity. In Ascaris suum, optimal activity is obtained with 2 mm MnCl2; but 4 mm CoCl2 has also been reported to activate PEPCK activity (Wilkes et al. Reference Wilkes, Cornish and Mettrick1982; Rohrer et al. Reference Rohrer, Saz and Nowak1986). For GdPEPCK carboxylation study, MnCl2 at 0.5 mm was found to be suitable co-factor; which is in slight deviation from an earlier report (Noce and Utter, Reference Noce and Utter1975). The role of cations in PEPCK activity is well documented (Lee et al. Reference Lee, Hebda and Nowak1981; Cornish et al. Reference Cornish, Wilkes and Mettrick1981b ; Rognstad, Reference Rognstad1981). Hebda and Nowak (Reference Hebda and Nowak1982a ) demonstrate that monovalent cations are not required for PEPCK activity and there is only one MnCl2 specific site available on the enzyme. However, other divalent cations have been reported to substitute for MnCl2, though to a lesser degree of activity (Schramm et al. Reference Schramm, Fullin and Zimmerman1981; Hebda and Nowak, Reference Hebda and Nowak1982a ). The data from previous investigations demonstrate that HCO3 − is essential for CO2 utilizing enzymes including PEPCK, and is responsible for the reaction proceeding in the direction of PEP formation in-vivo. It is also reported that a high concentration of CO2 in the cell may cause the enzyme to act only in the carboxylation direction under physiological conditions (Behm and Bryant, Reference Behm and Bryant1975b ). RePEPCK showed the optimal activity at 20 mm of HCO3 −, whereas GdPEPCK, at 8 mm.
Few inhibitors, such as, oxalate, 3-aminopicolinate, 3-mercaptopicolinic acid (MPA), sulfoacetate, phosphonoformate, phosphoglycolate and 3-phosphonopropionate etc., are reported to inhibit PEPCK differently (DiTullio et al. Reference DiTullio, Berkoff, Blank, Kostos, Stack and Sounders1974; Goodman, Reference Goodman1975; MacDonald, Reference MacDonald1979; Dayer et al. Reference Dayer, Dayer and Ghayour2013). Oxalate is known as a weak linear non-competitive inhibitor for PEPCK from chicken liver mitochondria with Ki value of 3.20 mm (Hebda and Nowak, Reference Hebda and Nowak1982b ); however, recently it has been found as competitive inhibitor and comparatively more inhibitory for rat PEPCK with Ki of 89 µ m (Dayer et al. Reference Dayer, Dayer and Ghayour2013. Recently, Balan et al. (Reference Balan, Mcleod, Lotosky, Ghaly and Holyoak2015) report that MPA inhibits PEPCK by binding to two discrete binding sites (substrate binding site and an unidentified allosteric site). Interestingly, the phytoestrogens, genistein and daidzein, inhibit RePEPCK in non-competitive manner; however, this inhibition is competitive for GdPEPCK. The rate and extent of inhibition and pattern for substrate protection for RePEPCK and GdPEPCK are variable for genistein and daidzein. Genistein more sturdily inhibits the enzyme activity than daidzein, which may be because of its spine similarity with PEP.
The possible mechanism of differential kinetics of genistein for PEPCK from both the parasite and its host may be explained in the light of molecular docking studies. Genistein was predicted to interact with residues of the nucleotide-binding site of PEPCK from the parasite suggesting a different binding pocket from its substrate like THP (Dutta et al. Reference Dutta, Ramnath, Tandon and Das2016a ). However, THP showed poor binding with the host enzyme, GdPEPCK. Recently, biocomputational study demonstrates the mixed inhibition mechanism on human PEPCK-C by genistein (Katiyar et al. Reference Katiyar, Jain, Dhanjal and Sundar2015). As observed from the structural studies, the nucleotide and substrate binding pockets of PEPCK are different. Interestingly, when genistein was docked into the chicken mitochondrial PEPCK model, it was predicted to bind near the substrate-binding site having interaction with R89 residue of R-loop. The significance of R89 for PEPCK optimal catalytic activity has clearly been elucidated by Johnson et al. (Reference Johnson, Mcleod and Holyoak2016). The present study suggests a differential binding affinity of genistein towards the parasite and its host, which needs to be validated in crystallographic studies.
In conclusion, from the data presented herein, we infer that there are significant differences between RePEPCK and GdPEPCK in overall catalytic properties (particularly absence of decaroxylation reaction by RePEPCK), Michaelis-Menten constants, nature of inhibition by phytoestrogens and their inhibitory constants. Further validation needs to be established, so as to exploit the enzyme as a robust therapeutic agent against cestodes for parasitism management. This study provides a basic background for in-vivo investigation using animal models for understanding efficacy of phytoestrogens as anthelmintic.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/S0031182017000518.
Acknowledgements
We are thankful to Dr Todd Holyoak, University of Waterloo, Canada, for kindly providing pE-SUMO vector and pE-SUMO protease. We thank the Head, Department of Zoology, NEHU, for providing infrastructural facilities for the study.
FINANCIAL SUPPORT
This work was financially supported by DBT, GoI, through a research project (BT/PR 12828/Med/30/221/2009, dated March 22, 2010). Ramnath, AKD and BD thank to DBT and University Grants Commission, New Delhi, for providing research fellowship.