



Introduction
Ramalina Ach. is a cosmopolitan genus with c. 150–200 species (Kashiwadani & Nash Reference Kashiwadani and Nash2004; Fletcher et al. Reference Fletcher, James and Purvis2009; Sérusiaux et al. Reference Sérusiaux, van den Boom and Ertz2010) which are known to be variable both morphologically and chemically (e.g. Culberson Reference Culberson1967; Sheard & James Reference Sheard and James1976; Rundel Reference Rundel1978; Krog & Østhagen Reference Krog and Østhagen1980; Stevens Reference Stevens1987). However, recent studies have shown that different morphological and/or chemical characteristics, once thought to be caused by environmental factors, can also indicate distinct species (Kashiwadani et al. Reference Kashiwadani, Nash and Moon2007; Ohmura et al. Reference Ohmura, Moon and Kashiwadani2008; Hayward et al. Reference Hayward, Blanchon and Lumbsch2014; Pérez-Vargas & Pérez-Ortega Reference Pérez-Vargas and Pérez-Ortega2014).
Anatomical analyses can contribute to an improved understanding of Ramalina taxonomy; however, anatomical traits were not always mentioned in descriptions and consequently this information is frequently missing in the literature for several species. However, Kashiwadani and collaborators have ensured that the taxonomic significance of the cortex and chondroid tissue in the identification of Ramalina species has now been recognized (e.g. Kashiwadani Reference Kashiwadani1986, Reference Kashiwadani1987, Reference Kashiwadani1992; Kashiwadani & Kalb Reference Kashiwadani and Kalb1993; Kashiwadani & Nash Reference Kashiwadani and Nash2004; Kashiwadani et al. Reference Kashiwadani, Moon and Lai2006, Reference Kashiwadani, Nash and Moon2007).
In Brazil, Ramalina species occur in almost all environments, but especially in areas with high humidity, such as near the mountains and sea. Southern and south-eastern regions, characterized by a transitional subtropical/tropical climate, present the highest number of species and biomass (e.g. Kashiwadani & Kalb Reference Kashiwadani and Kalb1993; Spielmann Reference Spielmann2006). Most species have a corticolous habit, occurring in forest borders and open areas. A few saxicolous species are found mainly along the coast on rocky shores, and on rocks that accompany the rivers in the plateau (Gumboski Reference Gumboski2016). Kashiwadani & Kalb (Reference Kashiwadani and Kalb1993) provided a good overview of the Brazilian species but, nonetheless, many regions and environments still lack adequate information about Ramalina species.
The aims of this work are to present a description of a new Ramalina species using morphological, anatomical, chemical and molecular data, and to provide a key to 32 neotropical saxicolous species based on a literature review and examination of herbarium material.
Material and Methods
Study region
The study region is located in the southern Brazilian highlands (>800 m a.s.l.), characterized by mosaics of grasslands (called ‘Campos de Cima da Serra’) and an Atlantic mixed forest (known as the Araucaria forest owing to the dominance of Araucaria angustifolia) (e.g. Rambo Reference Rambo1953; Boldrini Reference Boldrini1997; Behling et al. Reference Behling, Pillar, Orlóci and Bauermann2004; IBGE 2004). Collection points were situated in waterfall areas: ‘Cachoeirão dos Rodrigues’ (Silveira River; 28°35'59·85''S, 49°59'19·89''W, c. 1150 m, municipality of São José dos Ausentes) and ‘Cachoeira dos Venâncios’ (Camisas River; 29°01'03·72''S, 50°15'31·55''W, c. 850 m, municipality of Cambará do Sul) (Fig. 1). According to Köppen (Reference Köppen1936), the climate is categorized as Cfa (humid subtropical). The annual average temperature is 14·8°C and the relative humidity c. 80% (Moreno Reference Moreno1961). In winter, the temperature falls to −8 °C, with frequent frosts and occasional snow formation. The annual rainfall is spread throughout the year and varies from 1500 to 2000 mm (Boldrini Reference Boldrini1997).

Fig. 1 A, map of the study region in Rio Grande do Sul State, southern Brazil, with collection area ‘Cachoeirão dos Rodrigues’ (1) and ‘Cachoeira dos Venâncios’ (2); B, Silveira River, Cachoeirão dos Rodrigues, habitat of the type specimen of Ramalina fleigiae sp. nov. In colour online.
Molecular data
Small samples (0·01–0·05 g) of Ramalina thalli were soaked in acetone overnight. The acetone was then removed with a pipette and the samples were reduced to powder with a ball mill and subsequently used for DNA extraction following the protocol described in Cubero et al. (Reference Cubero, Crespo, Fatehi and Bridge1999). The markers selected were the ITS rDNA and IGS. PCR amplifications were performed in a Veriti Thermal Cycler (Applied Biosystems) using the primers ITS1F (Gardes & Bruns Reference Gardes and Bruns1993) and ITS4 (White et al. Reference White, Bruns, Lee and Taylor1990) for the ITS rDNA region and IGSf and IGSr (Wirtz et al. Reference Wirtz, Printzen and Lumbsch2008) for IGS. Each 25 µl reaction contained: 1× Taq buffer (Promega), 0·2 mM of dNTP set, 0·2 µM of each primer, 25 mM of MgCl2, 1U of DNA Polymerase (Promega) and c. 20–50 ng of DNA. The PCR conditions for ITS amplification were: 2 min at 95°C for initial denaturation, followed by 30 cycles of 30 s at 95°C, 1 min at 50 °C, 1 min at 72 °C, with a final elongation of 10 min at 72 °C. For IGS amplification the conditions used were: 2 min at 95°C for initial denaturation, followed by 30 cycles of 30 s at 95°C, 45 s at 46°C, 1 min at 72°C, with a final elongation of 10 min at 72°C. PCR products were purified with ammonium acetate and ethanol, and sequenced in an ABI PRISM 3500 Genetic Analyzer (Applied Biosystems).
Sequences from nine specimens of Ramalina fleigiae were obtained. We also analyzed sequences from material collected in southern Brazil: the saxicolous R. laevigata (three specimens) and the corticolous species R. exiguella (three specimens), R. gracilis (one) and R. sprengelii (two). In addition to their phylogenetic proximity, these species were selected as they share morphological, chemical (e.g. R. exiguela) and ecological (e.g. R. laevigata) features with R. fleigiae. Sequences from R. celastri, R. glaucescens, R. inflexa, R. ovalis and R. siliquosa used in the comparisons were obtained from GenBank. Lecania erysibe (Ach.) Mudd and L. leprosa Reese Næsb. & Vondrák were used as outgroup. Geographical origin, voucher details and additional information about the specimens are provided in Table 1.
Table 1 Specimen information together with the GenBank Accession numbers for the collections used in Fig. 2 Newly obtained sequences are in bold
Forward and reverse reads were assembled using Geneious 9.1.2 (http://www.geneious.com; Kearse et al. Reference Kearse, Moir, Wilson, Stones-Havas, Cheung, Sturrock, Buxton, Cooper, Markowitz and Duran2012). Sequences were aligned using the MAFFT algorithm (Katoh et al. Reference Katoh, Misawa, Kuma and Miyata2002) in Geneious 9.1.2 (auto function for the best algorithm choice, gap penalty=1·53, offset value=0·123 and scoring matrix=200PAM/k=2) and manually adjusted. The analyses were performed with two data sets: ITS only and ITS/IGS sequences combined. The best model for each matrix was obtained using the jModelTest2 program (Guindon & Gascuel Reference Guindon and Gascuel2003; Darriba et al. Reference Darriba, Taboada, Doallo and Posada2012). Maximum likelihood analyses were performed in PhyML (Guindon & Gascuel Reference Guindon and Gascuel2003) with 1000 bootstrap replicates for calculation of the nodal support, using the TrNef+G substitution model for the combined data set and TiM2+G for the ITS data set. Bayesian analyses were estimated using the software BEAST 1.8.2 (Drummond et al. Reference Drummond, Suchard, Xie and Rambaut2012) with the substitution model TN93+G for both data sets. A Yule tree prior and a random starting tree were selected and the MCMC runs were performed using four chain lengths of 10000000, sampling trees every 1000th generation. In TreeAnnotator v1.8.2 (part of BEAST v1.8.2 ), the first 20% of trees was discarded as burn-in and the maximum clade credibility tree generated. The results were checked in Tracer v1.6 (Rambaut et al. Reference Rambaut, Suchard, Xie and Drummond2014) for the MCMC convergence and the effective sample sizes (≥200). Conflicts between maximum likelihood and Bayesian analyses were assumed to be significant if two different relationships for the same taxa were supported with bootstrap values >75% and posterior probability >0·95 (Sérusiaux et al. Reference Sérusiaux, van den Boom and Ertz2010).
Additional tools, such as maximum likelihood (ML) distance (number of substitutions per site, s/s) calculations in PAUP* v4.0b10 (Swofford Reference Swofford2003) and the Automatic Barcode Gap Discovery (http://wwwabi.snv.jussieu.fr/public/abgd/; Puillandre et al. Reference Puillandre, Lambert, Brouillet and Achaz2012), were used to delimit R. fleigiae. The default settings (Pmin=0·001, Pmax=0·1, and 20 numbers of distribution) and four gap width values were tested (0·5, 0·7, 1·0 and 1·2) using Jukes-Cantor distances.
Morphological, anatomical and chemical analyses
Specimens were removed with a knife and dried at room temperature. Previously collected specimens from herbaria ICN and UFP were also studied, allowing us to identify the collection site and obtain fresh specimens.
Specimens were examined using standard techniques with stereoscopic (×20–40) and light microscopes (×400–1000). Freehand sections of the thalli and apothecia were mounted in water. Spot tests were conducted according to Huneck & Yoshimura (Reference Huneck and Yoshimura1996) and Orange et al. (Reference Orange, James and White2001), including observation under UV light and thin-layer chromatography (TLC) using solvent C.
The key to the 32 saxicolous species of Ramalina from the Neotropical region was based partially on literature (e.g. Nylander Reference Nylander1870; Malme Reference Malme1934; Grassi Reference Grassi1950; Kashiwadani Reference Kashiwadani1987; Kashiwadani & Kalb Reference Kashiwadani and Kalb1993; Marcano & Morales Méndez Reference Marcano and Morales Méndez1994; Aptroot & Bungartz Reference Aptroot and Bungartz2007; Kashiwadani et al. Reference Kashiwadani, Nash and Moon2007) and partially on herbarium specimens. The material examined was sourced from the following regions and herbaria: Argentina (CTES, MIN, MPUC, MSC), Bahamas (MSC), Brazil (BM, BHCB, CGMS, CTES, F, G, H, HAS, ICN, MPUC, SP, UFP, UPCB, TUR, W), Chile (BM, CTES, G, H, MIN, MSC, O, WRSL), Colombia (H), Cuba (G, H, MICH, MSC), Dominican Republic (MSC), Ecuador (MSC), Falkland Islands (MSC), Guadalupe (MICH, MIN), Guatemala (F, MSC), Guyana (BM), Haiti (MSC), Honduras (BM), Jamaica (H, MIN, MSC), Mexico (WIS, WRSL), Panama (F), Paraguay (CTES, H), Peru (DUKE, H, M, W), Puerto Rico (F, MICH, MSC), Uruguay (DUKE, ICN, MPUC) and Venezuela (MIN). In addition to the species described as exclusively saxicolous, we also included species rarely recorded as corticolous. Ramalina siliquosa (Huds.) A. L. Sm, a doubtful record for South America (e.g. Argentina; Calvelo & Liberatore Reference Calvelo and Liberatore2002), was integrated into the key.
Results and Discussion
Molecular data
The ITS and combined sequence data sets contained 549 and 958 unambiguously aligned sites, respectively. The specimens Ramalina fleigiae 1 to 8 presented identical sequences (in both markers) and thus only one sequence was used in the analyses. Phylogenetic reconstructions generated by maximum likelihood and Bayesian analyses, for both data sets, did not show incongruences and so only the Bayesian phylogenetic inferences are shown in Fig. 2. The species clustered in the two main clades have distinct morphological and anatomical features, as well as different geographical origins and habitats. Therefore, there are species with very distinct characteristics in the same clade, such as very long and somewhat cylindrical branches to mainly short and clearly flattened branches, as well as distinct patterns of chondroid tissue. The phylogenetic relationships in these Ramalina groups lack clear phenotypic and/or biogeographical affinities.
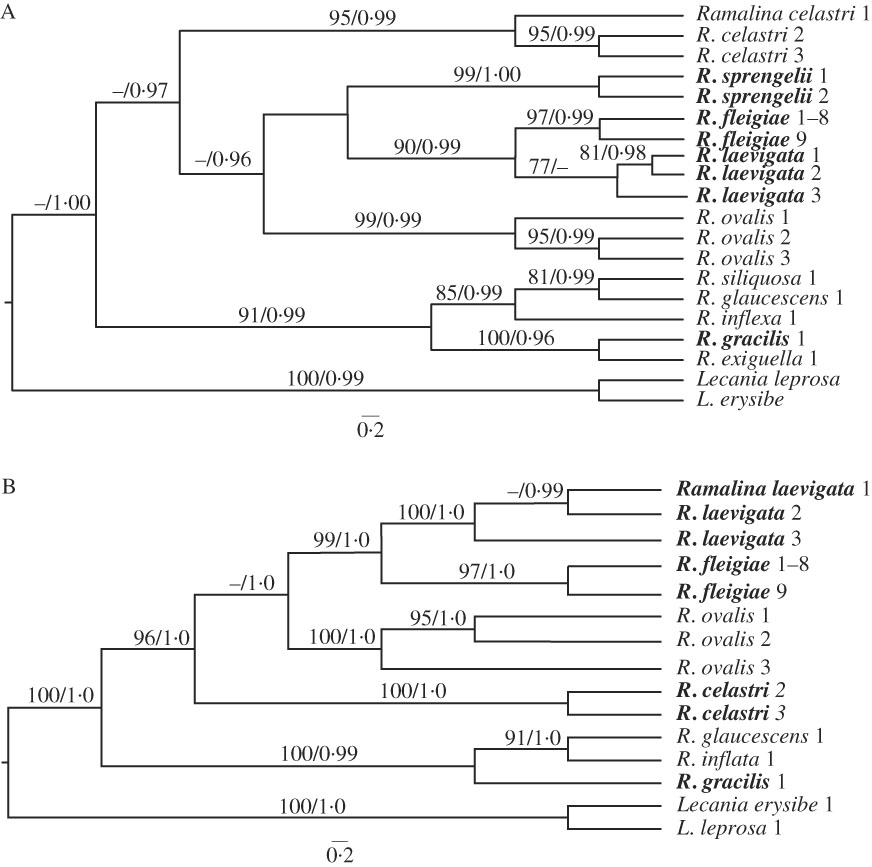
Fig. 2 Bayesian maximum clade credibility tree based on (A) ITS sequences and (B) the concatenated data set (ITS and IGS sequences) showing the phylogenetic relationships among Ramalina species, including R. fleigiae sp. nov. ML boostrap ≥70 (before the slash) and posterior probability ≥0·95 (after the slash) are shown above branches. Specimen details are given in Table 1. Specimens for which new sequences were obtained in this study are shown in bold.
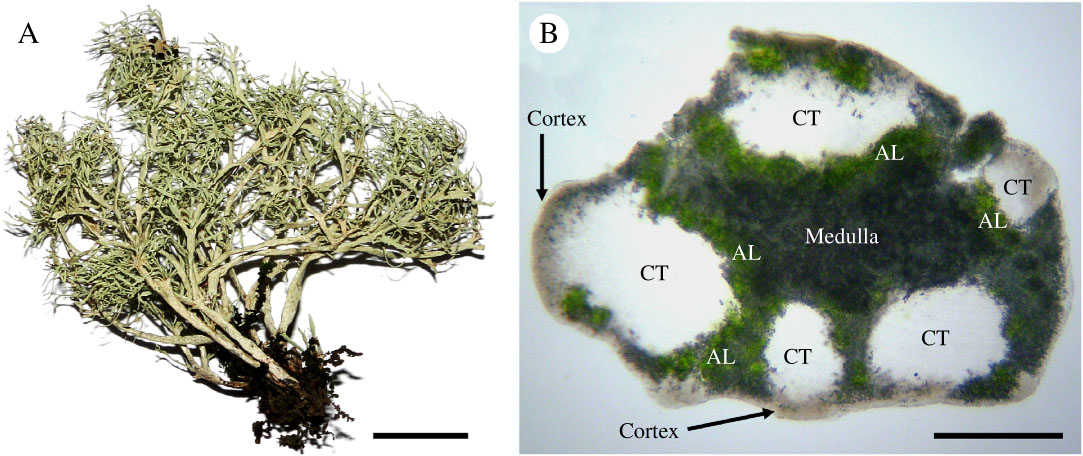
Fig. 3 Ramalina fleigiae. A, thallus (holotype) with solid, irregularly flat to subterete branches; B, TS main branch (holotype) showing the distinct cortex, the discontinuous, uncracked chondroid tissue (CT), algal layer (AL) below the cortex and around the chondroid tissue, and the medulla in the grey central portion. Scales: A=1 cm; B=200 μm. In colour online.
The sequences generated using R. fleigiae specimens clustered in well-supported clades with R. laevigata. Both of these species occur in riparian and saxicolous habitats, produce usnic acid only, and also present clearly distinct morphological and anatomical features. Ramalina fleigiae is densely branched in the upper half, irregularly flat to subterete, up to 1·5 mm wide, while R. laevigata is moderately branched and has distinct dorsiventral branches up to 2·0(–4·0) mm wide (Fries Reference Fries1825; Malme Reference Malme1934). In contrast, R. fleigiae is not closely related to R. exiguella and R. gracilis, two morphologically and chemically similar species. All have shrubby thalli, irregularly flat to subterete, branches both symbiotic propagules and a lack of medullary acids. While R. fleigiae occurs only on rocks above 1100 m, R. exiguella and R. gracilis occur mainly on trunks, twigs or rocks, and are strictly coastal species. Furthermore, R. exiguella and R. gracilis have linear to long linear pseudocyphellae while R. fleigiae is characterized by mainly irregularly ellipsoid to short linear pseudocyphellae.
Analyses of the ITS region revealed that R. fleigiae specimens presented ML distances between 0 (identical) and 0·001 s/s. The distances among the other species analyzed varied between 0·023 s/s (R. fleigiae×R. laevigata) and 0·210 s/s (R. fleigiae×R. inflexa; R. fleigiae×R. gracilis). For comparison, the mean distance between R. ovalis and R. celastri (recognized as distinct species by Hayward et al. Reference Hayward, Blanchon and Lumbsch2014) was 0·107 s/s, much higher than the distances between R. fleigiae and R. laevigata. All the settings tested in the ABGD analysis resulted in 10 distinct groups, corresponding to the species identification and corroborating the delimitation of R. fleigiae as a new species.
Morphological, anatomical and chemical analyses
Ramalina fleigiae Gumboski, Eliasaro & R. M. Silveira sp. nov.
MycoBank No.: MB 824641
Differing from Ramalina exiguella Stirt. by the thallus with branches originating from a single holdfast, most densely branched in the upper half of the thallus, and further distinguished by numerous irregular ellipsoid to short linear pseudocyphellae on the surface and margin of its branches.
Type: Brazil, Rio Grande do Sul State, municipality of São José dos Ausentes, locality of ‘Cachoeirão dos Rodrigues’, on a rock in the middle of Silveira River, c. 1150 m alt., 28°35'59·85''S, 49°59'19·89''W, 20 January 2015, E. Gumboski 5050 (ICN—holotype; UPCB, SP, H, F—isotypes). GenBank Accession no.: KY171855
(Fig. 3)

Fig. 4 Ramalina fleigiae (arrow) growing on mid-river rock, very near the water indicating they may be prone to temporary submergence (Silveira River, Cachoeirão dos Rodrigues, southern Brazil). In colour online.
Thallus saxicolous, shrubby, up to 5·0 cm tall and 6·0 cm wide, whitish green in the field, becoming stramineous in the herbarium; densely branched in the upper half in most well-developed thalli; branches originating from a single holdfast, consisting of dense groups of 2–7 branches rooting in a common, necrotic base up to 0·7 cm high; branching dichotomously anisotomic; main branches ramify mostly from the upper half; branches solid, rigid, irregularly flat to subterete, somewhat inflated, but not hollow, distinctly flattened mainly at their tips; main branches 0·4–1·5×0·2–1·1 mm broad; secondary branches 0·15–0·60×0·1–0·5 mm wide; apex somewhat truncated to slightly acute, sometimes curved appearing hook-shaped; surface opaque, irregular, ±striate owing to the presence of abundant pseudocyphellae resembling maculae; symbiotic propagules absent; pseudocyphellae numerous on lamina and margins, present across both the main and secondary branches, depressed on level surface, irregular ellipsoid to shortly linear, 0·15–1·0×0·05–0·15 mm, rarely orbicular, 0·1–0·2 mm diam.; cortex distinct, 10–35µm thick; chondroid tissue not cracked, discontinuous, in irregular bundles intermixed with medulla (but sometimes the major portion of the branches); main branches 80–620(–700) µm thick; secondary branches usually with one ±circular to irregular bundle, 30–210µm thick; medulla compact; algal layer below the cortex and around the chondroid tissue, 15–70µm thick.
Ascomata and conidiomata not found.
Chemistry. Cortex and medulla: K−, C−, KC−, PD−, UV−. TLC: usnic acid.
Etymology. This new species is dedicated to Dr Mariana Fleig, who devoted much of her life to the study of lichens and contributed enormously to the knowledge of Brazilian lichenology. Fleig also collected the oldest known herbarium specimens (in ICN) of this new species.
Distribution and habitat. The species described here is known only from two localities in the north-eastern plateau of Rio Grande do Sul State (Fig. 1). Ramalina fleigiae colonizes only rocks in the riverbed and waterfalls that are exposed to direct sunlight; some thalli were almost submerged in the water (Fig. 4) at c. 1150 m elevation. All thalli are probably temporarily submerged during occasional floods. No thallus was found on rocks that are further from the river. The species probably occurs in nearby rivers but due to very small population sizes these might easily have been overlooked.
The species was not found in similar environments visited by the first author in the adjacent states of Santa Catarina and Paraná.
Discussion. Ramalina fleigiae is a distinct species, known mainly for its saxicolous habit and branching pattern (originating from the same base, densely branched upper half). Branches are solid, irregularly flat to subterete (Fig. 3A), up to 1·5 mm wide, with numerous irregular ellipsoid to short linear pseudocyphellae on the lamina and margins. Anatomically, a distinct cortex with a discontinuous and not cracked chondroid tissue characterizes the species (Fig. 3B). There are no secondary metabolites in the medulla.
Similar species are R. gracilis (Pers.) Nyl. and R. exiguella Stirt. but both have thalli growing from a delimited holdfast, angular branches in cross-section, long linear pseudocyphellae and no distinct cortex; these two species are also strictly coastal (Fleig Reference Fleig1988; Kashiwadani & Kalb Reference Kashiwadani and Kalb1993). Ramalina osorioi Kashiw. et al. is a similar saxicolous species and has a shrubby thallus with solid branches. However, it is moderately branched, has distinctly dorsiventral branches and produces psoromic acid (Kashiwadani et al. Reference Kashiwadani, Nash and Moon2007). Ramalina continentalis Malme is a saxicolous species that inhabits rocks along rivers in Brazil, Paraguay and Uruguay (e.g. Malme Reference Malme1934; Osorio Reference Osorio1972). However, it has distinctly flattened and has much larger branches of 5–10(–20) mm width (Malme Reference Malme1934).
Ramalina furcellangulida Aptroot, endemic to the Galapagos Islands, has no distinct holdfast. Branches are irregularly angular in section, typically not flattened, often with rounded, irregular, laminal warts and linear pseudocyphellae, rarely with few punctiform tuberculate pseudocyphellae. This species produces divaricatic or salazinic and/or sekikaic acid (Aptroot & Bungartz Reference Aptroot and Bungartz2007).
Ramalina siliquosa (Huds.) A. L. Sm. is shrubby to pendulous and its main branches are more or less flattened, rarely entirely terete; however, its secondary branches are mostly furcate, rarely terete, and the surface is more or less matt, rugose, or frequently even foveolate, occasionally with well-developed furrows (Krog & James Reference Krog and James1977). In this species the chondroid tissue is clearly to moderately cracked (Kashiwadani Reference Kashiwadani1992). Ramalina cuspidata (Ach.) Nyl. has simple or sparingly furcate branches which rarely become densely branched towards the apices. Branches are mostly terete towards the base and remain so above or gradually become flattened (Krog & James Reference Krog and James1977). The holotype (H!) has a distinct 8–10µm thick cortex and the chondroid tissue is clearly cracked, continuous, and 50–220µm thick.
Ramalina geniculatella Aptroot, endemic to the Galapagos Islands, is initially shrubby, becoming pendant with age, branches are mostly flattened and partly terete, and most thallus parts have conspicuous whitish linear pseudocyphellae (Aptroot Reference Aptroot2008). An examination of the holotype (B!) showed that the cortex is absent, and the chondroid tissue is not cracked and forms almost regular bundles mixed with medulla.
Ramalina elegantula P. M. Jørg. has a caespitous thallus with linear thin branches, up to 0·5 mm wide. The pseudocyphellae are linear and marginal (Jørgensen Reference Jørgensen1977). The cortex is distinct, 10–20µm thick, and the chondroid tissue is continuous and uncracked, 60–105µm thick (O—holotype!). Ramalina leptocarpha Tuck. has a shrubby to subpendulous thallus. The branches are lanceolate, plane to somewhat canaliculated, and the pseudocyphellae are linear to ellipsoid, laminal, or rarely marginal. Ramalina bajacalifornica Bowler & Rundel has a shrubby to subpendulous thallus and flat or weakly canaliculated branches; pseudocyphellae are laminal or rarely marginal, short linear, somewhat elevated. This species produces salazinic acid in the medulla (Kashiwadani & Nash Reference Kashiwadani and Nash2004).
Two endemic saxicolous species of the Azores are morphologically similar to R. fleigiae. They differ by their branching and the pattern of pseudocyphellae, and by the production of secondary metabolites. Ramalina azorica Aptroot & Schumm has few antler-like branches or dense branches that become terete towards the tips. The pseudocyphellae are linear. It produces divaricatic acid together with an unknown pigment. Ramalina wirthii Aptroot & Schumm has branches that are mostly flattened, or in parts irregularly rounded, with several of the branches usually becoming terete toward the tips. It produces salazinic acid, usually together with protocetraric acid (Aptroot & Schumm Reference Aptroot and Schumm2008).
Ramalina jamesii Krog, R. portosantana Krog and R. timdaliana Krog, all endemic to Porto Santo Island near Madeira, are saxicolous and have shrubby thalli. However, R. jamesii has subterete and uneven branches, and fragmentation areas are common. The pseudocyphellae are shortly linear to ellipsoid. It produces divaricatic acid, salazinic acid and triterpenoids. Ramalina portosantana has subterete or more or less flat branches, and the surface is rugulose with irregularly reticulate ridges which can develop pseudocyphellae. The cortex is very thick, up to 80µm. The chondroid tissue occasionally adjoins the cortex but mostly forms numerous strands intermixed with the medulla. This species produces salazinic acid. Ramalina timdaliana is richly branched and the branches are subterete to somewhat flat, often with shortly linear pseudocyphellae. It produces divaricatic acid and triterpenoids (Krog Reference Krog1990).
Ramalina pluviariae Krog & Østh. is saxicolous and has subterete branches, and the pseudocyphellae are mainly linear, more or less anastomosing (Krog & Østhagen Reference Krog and Østhagen1980). It has no cortex and the chondroid tissue is discontinuous and clearly cracked, 70–350µm thick (O—holotype!). Ramalina hamulosa Krog & Østh. has subterete, angular to flattened branches, with numerous short branchlets terminating in nodules or hook-shaped structures. The pseudocyphellae are linear, restricted mostly to the margin of flattened branches (Krog & Østhagen Reference Krog and Østhagen1980). The cortex is distinct, 30–40µm thick, and the chondroid tissue is discontinuous, cracked, and 10–275µm thick (BM—isotype!).
Ramalina australiensis Nyl., a rarely saxicolous species, has thalli that are 3–10(–20) cm long, with sparse branching into the basal region of the thallus which becomes densely branched at the tips. The primary branches are usually subterete and thin, rarely thick, compressed and coarse (Stevens Reference Stevens1987). The chondroid tissue is clearly cracked, discontinuous, and 140–300µm thick (H-NYL—holotype!). Ramalina tropica G. N. Stevens has branches arising from the base with common lateral branchlets. The pseudocyphellae are linear, within which tuberculate extensions are sometimes present. The presence of salazinic and protocetraric acids is characteristic for this species (Stevens Reference Stevens1987).
Additional specimens examined. Brazil: Rio Grande do Sul State: municipality of Cambará do Sul, Cachoeira dos Venâncios, on rock, 2000, W. B. Sanders 00108.3B (UFP 29592); municipality of Jaquirana, locality of Cachoeira dos Venâncios, on rock, c. 850m, 29°01'03·72''S, 50°15′31·55''W, 2015, E. Gumboski 5054, 5055 (ICN), 5056, 5057 (SP); municipality of São José dos Ausentes, source of Rio do Marco, riverbed, on rocks, 1994, M. Fleig 6643, 6660 (ICN 99838); locality of ‘Cachoeirão dos Rodrigues’, on rock in the middle of the river (Silveira River), c. 1150 m, 28°35'59·85''S, 49°59'19·89''W, 2015, E. Gumboski 5049, 5051, 5052 (ICN), 5053 (JOI).

We are grateful to the curators of the herbaria B, BM, CGMS, CTES, DUKE, F, G, H, ICN, M, MICH, MIN, MPUC, MSC, O, SP, TUR, UFP, W, WIS and WRSL for the loan of specimens; to the curator and staff of ICN for allowing access to their facilities; to Dr André Aptroot for providing literature that was difficult to access; and to CAPES (Coordenadoria de Aperfeiçoamento do Pessoal do Ensino Superior) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) for financial support to ELG.





