



Introduction
Reintroduction biology has developed into a scientific discipline and has become a powerful conservation technique. The earliest reintroductions were primarily for game management and restocking but these early efforts have developed in sophistication to provide generic multi-taxa guidelines (IUCN 2012). Successful reintroduction practitioners used a multi-disciplinary approach, exploiting a species’ full adaptive potential to survive in human-dominated landscapes (Jones et al. Reference Jones, Heck, Lewis, Mungroo, Slade and Cade2008; Ewen et al. Reference Ewen, Armstrong, Parker and Seddon2012a; Jones and Merton Reference Jones and Merton2012).
The Southern Ground-hornbill Bucorvus leadbeateri occurs from southern Kenya to the Eastern Cape province of South Africa. The species is globally ‘Vulnerable’ but regionally listed as ‘Endangered’ in South Africa, Lesotho, Swaziland (Taylor and Kemp Reference Taylor, Kemp and Taylor2015) and Namibia (Simmons et al. Reference Simmons, Brown and Kemper2015), primarily due to high rates of unnatural mortalities. Known threats that contribute to adult mortalities include electrocution by energy infrastructure, poisoning by agro-chemicals, lead toxicosis, and trade for ritual and medicinal purposes (Morrison et al. Reference Morrison, Daly, Burden, Engelbrecht, Jordan, Kemp, Kemp and Potgieter2005). In South Africa, the population is estimated at 2,000 adults, with adult survival being the most sensitive parameter for maintaining a stable population (Morrison et al. Reference Morrison, Daly, Burden, Engelbrecht, Jordan, Kemp, Kemp and Potgieter2005). There is an apparent lag effect in the population decline, caused by local loss of individuals due to habitat fragmentation, anthropogenic threats and local extirpation (Rudolf et al. Reference Rudolf, Conner, Carrie and Schaefer1992). This suggests that losses went unnoticed until populations were already severely depleted of dispersing individuals and successful breeding groups.
The species exhibits demographic traits such as being long-lived, having low reproductive rates and being the largest obligate cooperative breeding bird species (Kemp Reference Kemp1995, Reference Kemp1988), which make it extinction-prone. Individuals live in socially complex groups, comprising an alpha breeding pair, with predominantly male offspring as non-breeding helpers. The helpers assist with territorial defence, predator vigilance, feeding the nest-bound female during incubation and early brooding, and later with the nestling and post-fledging care - essentially alloparenting (Kemp and Kemp Reference Kemp and Kemp1980, Kemp Reference Kemp1988, Reference Kemp1995a).
Given the species’ extensive territorial requirements, with territories ranging from 80 to 250 km2 (Kemp Reference Kemp1988, Knight Reference Knight1990, Theron et al. Reference Theron, Jansen, Grobler and Kotze2013a. Zoghby et al. Reference Zoghby, Ryan, Little, Reid and Hockey2015), groups outside protected areas often span several privately-owned properties, which complicates implementing census or conservation actions. Each group is highly territorial, with low dispersal rates between groups (Kemp Reference Kemp1995a, Wilson and Hockey Reference Wilson and Hockey2013, Carstens et al. Reference Carstens, Kassanjee, Little, Ryan and Hockey2019b). At any one time, less than 3% of the population lives alone or in small unisexual groups (Kemp Reference Kemp1988). However, being resident on permanent year-round territories also has advantages for the conservation manager. It eliminates concerns about seasonal movements, changes in social and habitat requirements, and exposure to, and mitigation of, threats across entire landscapes, which are issues that challenge reintroduction programmes for many other species (e.g. Ogada and Buij Reference Ogada and Buij2011). This allows spatially focused interventions for population reintroduction or expansion and compensates for other challenges when managing ground-hornbill biology.
Natural recolonisation of now-vacant areas should be possible (Theron et al. Reference Theron, Dalton, Grobler, Jansen and Kotze2013b) since habitat quality in savannas or grasslands appears to be viable, provided that a sufficiently short basal vegetation layer of < 50 cm is maintained (Knight Reference Knight1990), woody plants are not too dense (Loftie-Eaton Reference Loftie-Eaton2014) and there is no persecution. However, the species requires large trees or cliff ledges for roost sites (Zoghby et al. Reference Zoghby, Little, Ryan and Hockey2016), and sufficient shade to mitigate the effects of heat stress (Kemp and Kemp Reference Kemp and Kemp1980, Kemp Reference Kemp1995a, Zoghby et al. Reference Zoghby, Ryan, Little, Reid and Hockey2015). They nest in tree cavities, cliffs crevices and earth-bank burrows (Kemp Reference Kemp1995a), but readily accept artificial nest boxes (du Plessis et al. Reference du Plessis, Erlich, Ross-Gillespie, Rode and Kemp2007, Wilson and Hockey Reference Wilson and Hockey2013, Carstens et al. Reference Carstens, Kassanjee, Little, Ryan and Hockey2019a). No nesting attempt has yet been observed to fledge more than a single chick (Kemp and Kemp Reference Kemp, Kemp, Kemp and Kemp2007), despite more than 60 nests being continually monitored for over a decade, and many more monitored opportunistically since 1969. Harvesting of either the first-, second- or third-hatched redundant egg or chick was first attempted in 1969 (Kemp et al. Reference Kemp, Kemp, Turner, Kemp and Kemp2007). Breeding success and juvenile survival is affected by nest predation, but is largely environmentally driven with habitat productivity closely linked to rainfall patterns (Kemp et al. Reference Kemp, Joubert and Kemp 1989 ), and breeding is only attempted in productive seasons (Kemp Reference Kemp1988, Wilson and Hockey Reference Wilson and Hockey2013). Juveniles are dependent on adult care for at least a year after fledging, or longer if no subsequent juvenile has fledged (Kemp Reference Kemp1995a).
Considering these biological needs, reintroductions may be supported with habitat management, threat mitigation and, where necessary, the provision of artificial nests, which can increase productivity and survival. Conservation management of the Southern Ground-hornbill was first proposed in 1996 (Kemp and Begg Reference Kemp and Begg1996) and began in 1999 (Theron and Turner Reference Theron, Turner and Soorae2008). Initially, it was thought that the lack of suitable nest sites was the primary factor limiting population growth, but since then it has been found that anthropogenic threats such as secondary poisoning, electrocution on energy infrastructure and direct persecution are also significant drivers of decline (Kemp Reference Kemp1995a, Jordan Reference Jordan2011, Kemp and Verdoorn Reference Kemp and Verdoorn2013), particularly in landscapes outside formally protected areas. During reviews of conservation actions necessary to sustain the population, reintroduction was modelled as one of several available conservation tools and was considered to be as effective for population growth as the provision of artificial nest boxes (Morrison et al. Reference Morrison, Daly, Burden, Engelbrecht, Jordan, Kemp, Kemp and Potgieter2005, Spear Reference Spear2005). In 2011, a national Single Species Recovery Plan for the Southern Ground-hornbill retained reintroduction as a conservation action (Jordan Reference Jordan2011). Much has been learnt about the biology of the species since the first experimental reintroductions in 1995, through in- and ex-situ research, opportunistic insights and subsequent reintroduction attempts. There is now a need to ensure that these lessons are available to other reintroduction practitioners, both for this species, its congener the Northern Ground-hornbill Bucorvus abyssinicus, and for other cooperative breeding species with similar complex social structures.
Southern Ground-hornbill translocations in South Africa to date are defined as true reintroductions (Figure 1), rather than as reinforcements as suggested by Cilliers et al. (Reference Cilliers, Evans, Coetzee and van Rensburg2013). There are no common surrogate species on which these reintroduction or husbandry techniques could have been tested, as commonly done for other species (e.g. Andean Condors Vultur gryphus for California Condors Gymnogyps californianus, and Common Ravens Corvus corax for Hawaiian Crows C. hawaiiensis; Williams et al. Reference Williams, Pople, Showler, Dicks, Child, Ermgassen and Sutherland2013), nor are there methods that have been used for other species that could be easily adapted (e.g. Peregrine Falcon Falco peregrinus methods used for Aplomado Falcon F. femoralis; Heinrich Reference Heinrich and Soorae2008). The intention of these Southern Ground-hornbill reintroductions was not specifically to restore ecosystems, as has been successful for wolves Canis lupus (Ripple and Beschta Reference Ripple and Beschta2007) and dingoes Canis lupus dingo (Dickman et al. Reference Dickman, Glen and Letnic2009), nor to restore populations to ‘wilderness’ levels, but simply to slow, or in the long-term reverse, population declines where sufficient wild dispersal or recruitment is unlikely.
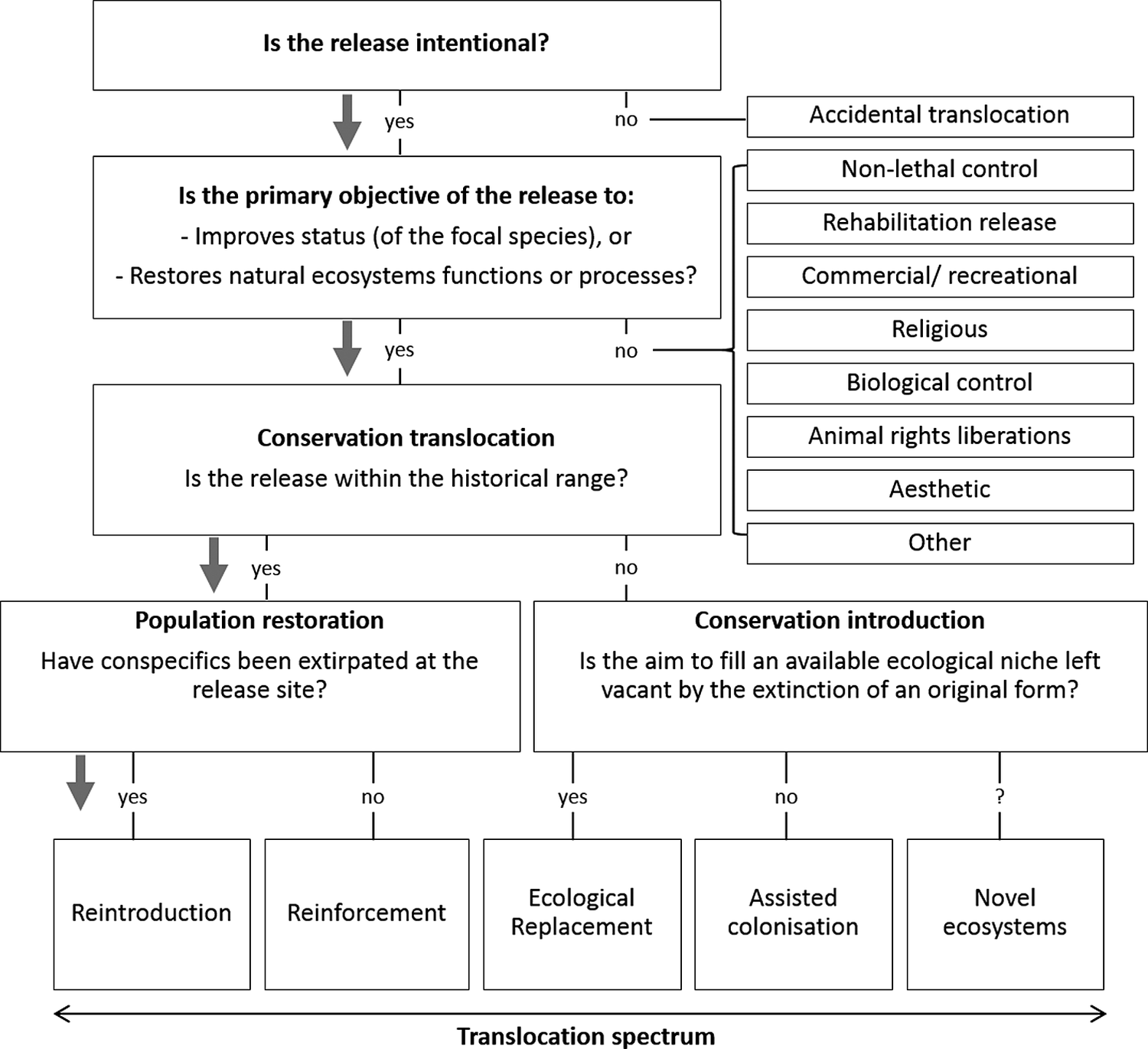
Figure 1. The translocation spectrum, from the reviews of Seddon et al. (2014) and the IUCN Reintroduction Guidelines (2012), with Southern Ground-hornbill reintroduction rationale indicated by arrows.
Pioneering efforts for ground-hornbills tested the feasibility of reintroductions but did not meet all criteria of earlier (Black Reference Black1991) or current guidelines (IUCN 2012). However, since 2011, once reintroductions proved feasible, these reintroduction criteria were largely addressed, and the guidelines were followed. Experimental reintroductions are increasingly seen as opportunities to better understand a species' ecology (Ewen et al. Reference Ewen, Armstrong, Parker and Seddon2012b), but these later reintroductions may be viewed as quasi-experimental, given that sample sizes are not yet sufficient for replicated, controlled experiments. K-selected species such as ground-hornbills display traits common to many threatened species, which also make them difficult to study in the field, while their conservation status also limits manipulation of wild populations. These logistical and statistical difficulties suggest alternative approaches, such as adaptive management where actions are designed as ‘experiments’ to gain information and allow decisions to be reviewed continually (Seddon et al. Reference Seddon, Armstrong and Maloney2007, Taylor et al., Reference Taylor, Canessa, Clarke, Ingwersen, Armstrong, Seddon and Ewen2017).
Gaps in knowledge, such as dispersal dynamics, can be investigated because reintroductions are de facto forced-dispersal events. Using redundant and harvested release stock, with no negative impacts on source populations, allows the use of individuals to understand habitat ecology, and individual, group and population behavioural ecology (Sarrazin and Barbault Reference Sarrazin and Barbault1996). The value of this approach has been recognised by legislation in the United States of America where such populations are listed as ‘non-essential experimental populations’ (Bartel and Rabon Reference Bartel, Rabon and Soorae2014).
Although there is a reasonable understanding of the complexity required for reintroductions of obligate cooperatively breeding species, there are few transferable techniques for such long-lived, socially complex birds as the Southern Ground-hornbill. Allee effects are also critical for population restoration of cooperative breeders (Rudolf et al. Reference Rudolf, Conner, Carrie and Schaefer1992, Courchamp et al. Reference Courchamp, Grenfell and Clutton-Brock1999) and when sourcing individuals for release (Rudolf et al. Reference Rudolf, Conner, Carrie and Schaefer1992). A bias in production towards male offspring requires management (Rapaport et al. Reference Rapaport, Kloc, Warneke, Mickelberg and Ballou2013) since dispersal abilities are generally low and female offspring tend to disperse earlier and further than males (Williams et al. Reference Williams, Pople, Showler, Dicks, Child, Ermgassen and Sutherland2013, Carstens et al. Reference Carstens, Kassanjee, Little, Ryan and Hockey2019b). New territories are likely formed when a dispersing female recruits one or two unrelated adult males to join her in an uninhabited territory (Courchamp and Macdonald Reference Courchamp and Macdonald2001), but translocated males potentially returned to their natal group if not transported far enough away due to strong territory fidelity (for Southern Ground-hornbill 70 km was insufficient; Rudolf et al. Reference Rudolf, Conner, Carrie and Schaefer1992) and re-establishment is slow (Williams et al. Reference Williams, Pople, Showler, Dicks, Child, Ermgassen and Sutherland2013).
We extracted lessons from the quasi-experimental Southern Ground-hornbill reintroduction programme in South Africa to investigate the viability of redundant second-hatched chicks as release birds; to establish how best to reintroduce captive-reared birds back into the wild, and to assess the feasibility of continuing reintroductions as a conservation tool for the species. We provide a sound opportunistic overview of specific parameters determining success or failure of past releases. This provides a framework for engaging other complementary disciplines to ensure that sound evidence-based plans can be developed. We also present six lessons learnt from implementing various reintroduction techniques and from investigating aspects of the species’ ecology during these reintroductions.
Methods
We followed standards suggested by Sutherland et al. (Reference Sutherland, Armstrong, Butchart, Earnhardt, Ewen, Jamieson, Jones and Lee2010) for documenting and monitoring avian reintroduction projects, ensuring that failures are as well documented as successes. Data were extracted from newsletters, diaries, photographic archives, press releases, reintroduction monthly reports, published accounts, spreadsheets and archived programme emails. From this, a database was constructed to profile all reintroductions. To fill data gaps or seek clarification, contact was made where possible with the original staff and partners involved. This assessment spanned reintroductions from 1995 to 2016, with ongoing reintroductions indicated. Two releases conducted in 1995 were a collaboration between the Transvaal Museum (now Ditsong Museum of Natural History) and Swaziland National Trust Commission. All subsequent post-1999 releases were conducted by the Mabula Ground Hornbill Project, in collaboration with multiple private, government and non-profit partners.
Releases focussed largely on private protected areas that approached ‘pristine’ habitat as closely as possible (Table 1). Most previous research had been conducted in the Kruger National Park, South Africa, and habitat suitability was initially based on comparable habitat type and condition. Since ground-hornbill groups are resident on fixed territories, release sites needed to be extensive enough to sustain at least one territory.
Table 1. Description of individual release sites for Southern Ground-hornbills in South Africa (SA) and Swaziland (SW). Land tenure was private except for Malalotja Nature Reserve in Swaziland. Land use was game reserves except for Haenertsburg, which included silviculture, small scale cropping and dairy practices.

Founder groups were reintroduced at eight sites in the Limpopo, Mpumalanga and Eastern Cape Provinces of South Africa and in Swaziland (Figure 2; Table 1). The size of each release site was considered suitable to hold at least one group, based on an estimated territory density of 100 km2 per group (Kemp Reference Kemp1995a), but where surrounding areas for the establishment of neighbouring groups existed these were favoured. Natural predators were present at all sites and low levels of persecution and limited (or no) use of poisons by landowners suggested. Loan agreements with landowners were used to ensure that released birds were cared for and monitored.

Figure 2. Map of eight reintroduction sites within the historical and current distribution ranges of Southern Ground-hornbills in South Africa and Swaziland, across an annual average rainfall gradient (ARC-ISCW 2014). Release sites listed in chronological order of the release attempts: 1. Malatoja Nature Reserve; 2. Mabula Private Game Reserve; 3. Haenertsburg; 4. Marakele Buffelspoort natural area; 5. Shamwari Game Reserve; 6. Madikwe Conservancy; 7. Thaba Tholo Game Farm and 8. Loskop Dam Nature Reserve.
Release birds for the reintroduction programme were redundant second-hatched chicks, harvested and captive-reared in compliance with protocols that ensure the harvest had no negative impact on existing wild populations. The stock was predominantly chicks harvested from wild nests, in areas where populations were deemed to be at carrying capacity, and where breeding success was monitored. However, some ground-hornbills were captive- or parent-reared and bred in captivity from birds that were themselves wild-harvested second-hatched chicks. In addition, the stock was opportunistically sourced as rehabilitated wild-reared birds: poisoning survivors (n = 2), illegal capture (n = 1), or experimental translocations of helper beta-males captured from wild groups (n = 2).
The quality of reintroduction stock varied and included captive-reared birds socialised to humans and/or conspecifics to varying degrees, and parent-raised captive-bred birds (Appendix S1 in the online supplementary material). Since 2012, only birds considered sufficiently ‘wild’ were used. These originated from a captive rearing centre at the Loskop Dam Nature Reserve (LDNR), Mpumalanga Tourism and Parks Agency (Table 1), which allowed for maximum conspecific socialising and minimal human intervention due to its rural location. Although efforts were made to improve socialisation at zoo-based facilities, those birds were still largely unsuitable due to insufficient conspecific socialisation, despite attempts to move chicks at fledging age to the rural facility. The rationale was that moving fledging chicks into existing groups in a ‘wilder’ environment would counter inadequate socialising during rearing but proved largely unsuccessful. If socialising during rearing was inadequate, it could not be corrected later as the birds were already too socialised to humans. Sample sizes were too small (n = 5) and sex-biased (female 4:1 male) to assess if the captive parent-reared stocks were of higher quality, although their behaviour matched that of captive-reared stock from LDNR. Parent-reared chicks received antibodies or microbes administered through the female's saliva. In all other respects, the diet of captive- and parent-reared chicks was the same, as captive pairs were fed the same food for chick provisioning as the hand-reared chicks received. Releases were conducted with birds ranging from fledging age (~ 90 days) to 21 years old. The average age was three years, with a median of two years.
To compound the analysis, the artificially reared birds were raised in various environments. One rural facility was able to simulate a natural environment, while others were more typical sterile captive conditions. Stock at the rural facility was held, as far as possible, in groups with ‘normal’ demographics, and parent-reared male chicks were left with their parents for a number of years to gain helping experience. Previous analysis suggested that age-at-harvest was the major determinant for survival during captive rearing and that the production of release-quality birds improved with time and with an enhanced understanding of socialising requirements (Kemp et al. Reference Kemp, Kemp, Turner, Kemp and Kemp2007).
Initially, the transportation of harvested chicks from wild nests to a captive-rearing facility was by road, but later sponsored flights reduced the time and stress of long road trips (> 4 hrs). For fledged ground-hornbills moved from holding aviaries to soft-release aviaries at release sites, all efforts were taken to minimise transport time. Birds were transported individually on soft bedding in crates with good ventilation and darkened with shade-cloth or a linen sheet. Transport-time was an average of five hours, with a maximum of eight hours to return a rehabilitated female to the wild. Captive-reared birds were initially caught by hand in holding aviaries, and later by using large gillie catch-nets to minimise capture time. All processing of individuals (weighing, measuring, drawing blood and ringing) was conducted prior to transport and release. Birds were moved from the captive holding site at dawn, to allow for most of the travel period to occur in the cool of the day. On arrival at release sites, birds were left crated for 30 minutes to settle down before being released into a soft-release aviary, always well before sunset. Behavioural observations were made for several days to ensure that the birds had settled, and then only the human ‘shepherds’ (appointed to walk with them, keep them from harm and provide general care) remained on site.
One wild subadult male was caught in a large walk-in trap for use as an experienced mentor for augmentations. The targeted wild group responded to pre-recorded play-back calls and was attracted into the trap by a life-sized fibreglass decoy and bait items. Once the target bird was in the trap, a drawstring was pulled, and the bird was swiftly caught by hand. The bird was processed, its health assessed, and then crated for transport. From trap-closure to crating took no longer than 10 minutes.
Release birds of varying ages between fledging and 21 years considered ready for release were moved to release sites, usually into an on-site aviary for between 2 and 61 days, and then released. After release, supplementary feeding of day-old poultry chicks, chicken heads, mice or mealworms Tenebrio molitor beetle larvae were offered to keep the birds attracted to the release site and to prevent starvation if individuals were not accepted by the group or were unable to forage successfully.
Most releases were to form a founder group where no birds had been released previously or were to augment an already established reintroduced group. A human ‘shepherd’ was initially employed to guide and protect released birds. Subsequently, adult male wild ground-hornbill mentors were used to ‘lead’ and 'educate' individuals, with human intervention limited to daily monitoring.
Stainless steel rings (size G, internal diameter 26 mm), engraved with a unique number and address of the South African Bird Ringing Unit (SAFRING), were applied on the right leg for individual identification and registered with SAFRING and the Pan-African Association of Zoos and Aquaria (PAAZA) Southern Ground-hornbill studbook. Plastic colour bands in various combinations were used on the left leg, and a unique ten-digit microchip transponder was inserted into the muscle between the shoulder blades (for youngsters under four months) or into the left pectoral muscle. Prior to release, 15 g, tail-mounted, very high frequency (VHF) telemetry transmitters (Holohil Model R1-2C, with an estimated battery life of 24–36 months) were fitted.
A total of 47 individuals were released between 1995 and 2016, with an additional 15 cases where birds were moved between release sites. The first five birds released in 1995 were not sexed (immature birds only show sexually dimorphic colours after two years of age and DNA sexing was not available), but subsequent releases comprised 18 females and 24 males. Reintroduction techniques were amended with time and experience, and three distinct phases of reintroduction management were defined (Table 2). In addition, management of reintroductions changed in 1999, 2009 and 2010.
Table 2. The basic characteristics of three distinct management phases of Southern Ground-hornbill reintroductions.

Statistical analyses
Birds of varying quality for reintroduction were released under various environmental, biological, social and management conditions, with protocols amended for each subsequent release. Adaptive management was necessary but made comparisons among attempts difficult. Due to the varied methods of the releases, it was not possible to assess the primary success criterion of survival to maturity. This would ensure that an individual had survived to sexual maturity, learnt by observation and experienced appropriate hunting, defence, courtship and breeding behaviours, likely assisted in caring for subordinate members of a group, and avoided natural predation and anthropogenic threats. Therefore, we used two alternative binary response-variables to quantify reintroduction success for analytical purposes of available data: 1) survival to six months post-release, to account for any immediate health and behavioural inferiorities in release stock that would affect survival (n = 62), and 2) survival to one-year post-release, to account for an individual’s acceptance into the group social structures, which affords enhanced survival (n = 60).
Recent reintroductions were excluded from the analysis if individuals were yet to reach these temporal benchmarks. To analyse data across varied release types, with limited controls due to lack of experimental design and ongoing adaptive management, generalized linear mixed models (GLMMs) were fitted with bird identification (bird ID) as a random effect (Bolker et al. Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009), using R (v. 3.0.2, R Core Team, Vienna, AT). To assess survival probabilities, models were fitted with a binomial distribution (logit link function) using the package ‘lme4’ (Bates et al. Reference Bates, Mächler, Bolker and Walker2015).
The analysis was exploratory, trying to find common denominators of success, and several a priori hypotheses for a set of 12 predictor variables (Appendix S2) were tested in the models to represent alternative explanations of how predictor variables influenced the response variable. These were defined according to both physical and psychological profiles of each individual bird, release and existing group demography, age at release, the season of release, release method(s), release site, management, and monitoring types. Several of these variables were also found to be important in other reintroductions (Meretsky et al. Reference Meretsky, Snyder, Beissinger, Clendenen and Wiley2001). Variables were selected for their relevance to the species' biology (Morrison et al. Reference Morrison, Daly, Burden, Engelbrecht, Jordan, Kemp, Kemp and Potgieter2005), and for where reliable data existed for each release. This allowed us to extract the greatest heuristic value from the available data and to establish a comprehensive baseline for future analysis, on which to set a priori hypotheses for sound experimental design in the future. Sample sizes were not sufficient to exclude individual events that occurred in the same year, site or using the same methodology, but variability within these temporal, spatial and methodological categories was likely sufficient to ensure no pseudo-replication. Categorical data was simplified to be able to retain discriminatory value.
Single linear models were used due to the relatively small sample sizes. In each model, the null model (intercept-only) was also included. Correlated variables were not used in the same models (Appendix S2). An information-theoretic approach, using log-likelihoods and comparisons of Aikake’s Information Criterion (AIC), was used for final model selection (Bolker et al. Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009), where the lower the AIC value, the better the model fit. An alpha P < 0.05 indicated significance.
Results
Pre-release factors
Choice of release stock:
Initial releases used any available ground-hornbills but later expanded harvest and captive-breeding programmes allowed for the selection of more suitable individuals. Age was not a significant determinant of initial survival (six months post-release).
Season of release:
Season of release was positively correlated with survival after release (Table 3). Fledging occurs in January to March when food productivity is at its highest in the austral summer rainfall season. Twenty releases were conducted in the cool dry austral autumn and winter (April-July), 13 during the hot dry spring and early summer (August-October), and 33 during the mid-summer rainy season (November-March; peak breeding season). Three birds that succumbed to starvation or dehydration had been released during the hot dry period (with no group or mentor support). No detrimental trends were observed for birds released during the cool dry period. Release in the rainy season contributed to successful survival of individuals up to six-months.
Table 3. Information-theoretic model selection for linear regression models relating to predictor variables with proportion of individuals surviving to six months for (a) variables where n was comparable and (b) where n varied by linear models but analysis could still be conducted (though none were significant). Significant variables (P < 0.05) are highlighted in grey.

K = total number of parameters (explanatory terms + random term + residual variance); AIC = Akaike’s Information Criterion; ∆AIC = difference between the AIC value for a given model and the model with the lowest AIC; w i = weight of model; Deviance = -2 log-likelihood.
Type of release and aviary security:
Hard-releases, when five individuals were released immediately into the landscape on arrival at a release site (1995–1999), all failed. This was likely compounded by the release of poorly socialised birds. Soft releases, using an on-site enclosure for pre-release captivity, were used for all subsequent releases so the birds could acclimate and interact, through the mesh of the enclosure, with previously released birds. The mean acclimation period was 62 days (range = 2–292 days), but 21 days was considered adequate as the longer the birds were in the aviary the greater was the risk of injury. Improved aviary security (both holding and soft release) would have avoided losses to predation by leopard Panthera pardus (n = 1) and honey badger Mellivora capensis (n = 3), and escape from the holding aviaries (n = 4). Once aviaries were predator-proof (electrification and/or base secured from digging predators with a 1-m deep concrete or metal-mesh skirting) no further losses occurred. The latest aviaries were built with a catch-cage atrium to reduce stress when capturing the birds, which also functioned as double-door access to prevent accidental escapes.
No instances of inadequate biosecurity were detected. No trespassing occurred at any of the holding or soft-release aviaries, despite the species having human-related value for traditional medicine and ritual purposes (Williams et al. Reference Williams, Cunningham, Kemp and Bruyns2014). However, caution must be taken as one incident occurred in 1992 at a South African zoo when a male was caught and killed through the enclosure fencing, with body parts removed, purportedly for traditional medicine.
Enrichment and pre-release training:
Pre-release enrichment is necessary to develop natural behaviours, and to prevent boredom. Early releases of human-habituated individuals utilised no enrichment, but later more effort was made to include daily enrichment. This included the provision of mealworm beetle larvae hidden in leaf bundles or fruits to encourage self-foraging and extend feeding time, dung from African elephant Loxodonta africana and white rhinoceros Ceratotherium simum to allow for practice at sifting through dung piles for insects, and insect larvae in Combretum and Kigelia spp. seed pods to allow for smashing and breaking techniques to be honed. Being social, it was noted that the best enrichment was by having conspecifics to engage with and mimic. Ground-hornbills held alone in soft-release aviaries showed elevated stress behaviours, such as increased pacing and prolonged contact calling.
Conditioning to facilitate post-release management, using an audio cue at feeding time (a whistle or recorded call) and associated visual cue when being fed (a red bowl), if applied consistently, was successful, even after several years without use.
Genetic screening:
Records of natal nests were maintained in the PAAZA studbook and genetic samples bio-banked for analysis prior to pairings and release-bird choice.
Post-release factors
Survival to adulthood:
Data were insufficient to perform rigorous statistical analyses, but Phase I saw survival to adulthood (taken as year 6) of 6%, rising to 15% in Phase II and 31% for Phase III, with an additional seven immature birds still alive, which if they survive, will enhance survival to adulthood to 38%. This excludes birds released as adults. Average natural survival in a protected area was estimated to be 30% (Kemp Reference Kemp1988).
Survival to six months:
Surviving for six months post-release, or necessary removal from a site due to aggression, indicates that there are no immediate problems with the release candidate (excessive habituation to humans, physiological or psychological issues, socially accepted and adequate foraging ability). Only two predictor variables, the season of release and phase of management, were significant to survival when comparing relative contributions in explaining variation in the data (Table 3). Probability of an individual surviving six months post-release was greatest for releases during the rainy season (November to March, Z = 2.27, P = 0.02; Table 3, Figure 3) and under current management (Phase III, Z = 2.20, P = 0.03; Tables 3 and 4, Figures 3 and 4). The remaining predictors were not significant (P > 0.05).

Figure 3. Differential effects of the factors within significant determinants of survival of reintroduced Southern Ground-hornbills to six-months post-release with single linear models for the season of release and project management phase. (Values are given as a back-transformed mean ± SE; n = 62 individuals).
Table 4. Summary results of the binomial Generalised Linear Mixed Model showing the effects of season of release and management phase on the likelihood of a reintroduced Southern Ground-hornbill surviving to six months after release (n = 62 individuals).


Figure 4. Differential effects of significant determinants of survival of reintroduced Southern Ground-hornbills to one year or longer post-release with single linear models for wild experience. (Values are given as a back-transformed mean ± SE; n = 60 individuals).
Survival to one year:
Surviving until one year after release indicates an individual’s ability to survive a dry season, also suggesting group acceptance and experience in predator avoidance. The only significant determinant of survival to one year was whether they had wild experience (Z = 1.97, P = 0.05), which was learnt from wild mentors in ‘bush-schools’ (Tables 5 and 6, Figure 4). In contrast to survival to six months, neither season of release nor phase of management affected survival to one year, although management phase ranked as the second-most important determinant after wild experience and, given larger sample size, may have had a stronger effect. Although the remaining predictor variables were not significant singular determinants of success, they, and some other aspects of reintroductions, were still important components, and their documentation and trends are presented as the basis for a priori hypotheses in the future.
Table 5. Information-theoretic model selection for linear regression models relating to predictor variables with the proportion of individuals surviving to one year for (a) variables where n was comparable and (b) where n was varied by linear models could still be conducted (though none were significant).

K = total number of parameters (explanatory terms + random term + residual variance); AIC = Akaike’s Information Criterion; ∆AIC = difference between the AIC value for a given model and the model with the lowest AIC; w i = weight of model; Deviance = -2 log-likelihood.
Table 6. Summary results of the binomial GLMM showing the effects of season of release and management phase on the likelihood of a reintroduced Southern Ground-hornbill surviving for one year after release (n = 60 individuals).

1 No wild experience was the reference category
Contingency tables, essentially categorical logistic regression models, confirmed these results using cell count and standard residuals. Survival to six months was significantly higher than expected in the rainy season. Survival to one year was higher than expected for birds that had gained some wild experience in a ‘bush-school’ and lower than expected for entirely naive birds (Table 7).
Table 7. Contingency analysis for logistic regression models relating predictor success with the probability of a successful reintroduction.

Group formation versus augmentation:
Releases were divided into two categories; formation of a new group in areas where wild birds no longer existed, and augmentation to expand initially reintroduced groups or later to 'wild' naive stock, but never into existing wild groups. In total, 29 birds were used to form nine new founder groups, while a further 33 were released into previously released groups (Appendix S3). The first three reintroductions (1995, 1999, 2000) consisted of eight naive hand-reared birds with no wild experience. These failed due to the birds experiencing predation (n = 4) or being unable to forage well enough (n = 2), while the remaining two birds probably survived solely due to the opportunistic subsequent augmentation of a rehabilitated sub-adult wild male bird as a mentor. The sub-adult wild male was a rehabilitated bird from a group which was poisoned and became the alpha male establishing a territory on Mabula Private Game Reserve (MPGR).
Seventeen augmentations were attempted with male birds. A wild adult male that was rehabilitated after a wild poisoning event had eight hand-reared males added into his group, for which he took on the role of mentor, including kin; a wild-bred son and a second son reared artificially after its mother was killed during incubation. The only aggression shown was to another rehabilitated adult male excluded from the territory. A further nine augmentations of juvenile and sub-adult males to other experienced males were successful.
Nineteen augmentations were attempted with females. Of the seven instances where a female was added to an all-male group, only one behaviourally compromised individual was not accepted. In the remaining 12 augmentations of females to groups, where an older alpha female was already present, the introduced female was immediately or with time (< 8 months) rejected (n = 10), and even moved out of the group territory on to other sites due to aggression from the alpha male (n = 8). The excluded females were too young (range = 3–5 months old) to show distinctive sexual colouration, which is only evident, on average, at 14 months of age. A second female accepted into the group was mentored by a beta male but died by electrocution just 93 days post-release at the age of five months, while another was accepted into a group that consisted of just an older female and a juvenile male. No second-female augmentations were attempted post-2012.
Two clear factors emerged from attempted augmentations and group formations that lowered the probability of success: (1) lack of an experienced male mentor, and (2) the failure of second-female augmentations.
Mortalities of reintroduced ground-hornbills:
No mortalities or injuries occurred during capture or transport. Six mortalities occurred during holding in soft-release aviaries due to leopard (1) and honey badger (3) predation, starvation due to cage-mate competition (1), and lead toxicosis (1) from spent lead-ammunition fragments in provided food. A total of 28 ground-hornbills died post-release (Appendix S4). Of these, nine were due to natural causes (predation, a falling branch, disease), while the remaining 19 were all human-induced, either through improper management (habituation during rearing, lack of sufficient survival skills, not recognising unsuitable birds; n = 5) or due to deaths from anthropogenic threats at release sites (poisoning, electrocution, persecution; n = 14). Predation occurred either in groups with no experienced mentor (n = 4), or when juveniles of one-month post-fledging were predated where a mentor was present (n = 2). In only seven cases was a mortality recorded without knowing the cause of death (three birds were assumed dead after an escape from holding aviaries; one with no transmitter banished from a group; three suspected poisonings after disappearing on a farm where the farmer acknowledged indiscriminate use of aldicarb-laced bait; and a carcass found decomposed in a field together with several poisoned Helmeted Guineafowl Numida meleagris). Post-mortems were not always conducted, but since 2012 were mandatory and identified the cause of death, even if the cause of death was obvious, e.g. predation.
Movement post-release:
Systematic monitoring of early releases was patchy and so the reasons for and when birds chose to move where they did was poorly understood. Many of the earlier release sites were chosen opportunistically, rather than according to a strict protocol, and group and individual movements often forcibly restricted by catching birds and moving them back to the central release site to ensure their safety and reduce logistical expenses. Since 2012, releases were generally unforced in this respect but still monitored.
In every case, reintroduced birds ranged widely post-release, usually beyond the borders of the ‘safe’ areas selected, even if such sites were considered large enough to hold several groups. In four cases where groups moved beyond the boundaries of the release site, they suffered mortalities (Malalotja Nature Reserve 1, MPGR 1, Thaba Tholo Nature Reserve 3) or were caught for the aviculture trade but recovered (n = 2). The furthest single movement was 40 km. Prior to that, an alpha pair traversed an area approximately 100 km across, but it is unknown if this was to investigate their surroundings, to find their way back to the original release site or to find conspecifics.
An alpha male, which had been moved to a second site after he had established his territory, traversed the equivalent of approximately four average-sized territories (100 km2 group density) to his natal group site. This return route was across mostly cattle and game farms. Four beta males of natural dispersal age (4–6 years) were translocated successfully over 70–250 km with no signs of 'homing' behaviour. An extraordinary incident occurred when an alpha female was moved to a nearby town for veterinary care. Her mate was found to have followed the vehicle containing the female, and this was only realised by the researchers once she had been treated and when he was following the vehicle back to the release site, each bird keeping contact through repeated calling; traversing 200 km over a 24 hour period (Turner Reference Turner2005).
Long-term food availability and supplementary feeding
Ground-hornbills are faunivorous and opportunistic in their diet preferences, including scavenging if the opportunity presents itself. The only reintroduced ground-hornbills unable to forage sufficiently well were two naive, mal-imprinted to humans, hand-reared juveniles, released with no mentor bird, and a behaviourally compromised female that should not have been released. Another case of starvation occurred in a soft-release aviary, where a bird was out-competed at feeding times by an individual from the same age cohort.
At Malalotja Nature Reserve (MNR) release 1, supplementary food was supplied daily for 10 days, and thereafter at 2- to 3-day intervals until, within a month, they did not always return. No food was supplied to MNR release 2 because they immediately dispersed separately on release and could not be tracked down in time before they starved. At MPGR, daily supplementation was initially provided, since the naive hand-reared birds needed time to learn to forage, and food was used to keep them bound to a pre-defined territory area (Theron and Turner Reference Theron, Turner and Soorae2008). Once a wild male was included, and group augmentation increased beyond natural levels (wild groups would only ever need to support a maximum of one chick per year), the provision of additional food helped him to feed additional immatures. A wild adult male, banished from the territory to an adjoining area, survived alone with no supplementary feeding for at least seven years. The second group with a wild mentor at Thaba Tholo Nature Reserve (TT) did not make use of regular supplementary feeding and the whole group, including a yearling, survived an atypically dry winter with minimal support. Despite release sites having sufficient prey available, supplementary feeding continues to be used as a technique for anchoring birds to safe areas while territory formation and social bonding occurs and, in the case of one site, to exclude the use of part of a territory where a farmer was known to use poisons regularly and indiscriminately.
Pair bonds and breeding
Pair bonds were managed, largely eliminating natural mate choice. Although many of the reintroduced pairs were not sexually mature, they exhibited strong bonds (n = 4 pairs). Bonding was reflected by courtship behaviour (gifting food items, courtship preening, spending time together away from the rest of the group and at the nest, and by receiving nest lining carried by the males). Even when not offered a mate choice, managed pairings have been largely successful (78%) in at least initiating courtship. In the few instances when pairings were not successful (n = 7 of 25, 22%), poor husbandry management (unsuitable nest structure, no nest lining provided, no small food items provided regularly for gifting) was apparent. No differences in successful bonding were observed whether the male had any wild experience or not, and only three of the 25 pair-bonded females had any wild experience. Territoriality is evident even in captivity, with one male and one female killed when two pairs were housed together in the same aviary. Captive facilities generally reared same or similar age cohorts together, sometimes as siblings, which did not impact future mate bonds.
Provision of artificial nests:
Artificial nests were provided at MPGR (one hollow Eucalyptus sp. log, floor diameter 40 cm), HBG (two wooden boxes, specifications unknown), LDNR (one Plastiwood and one insulated-resin box, both with 50 cm diameter hollow) and TT (one Plastiwood box with 50 cm diameter hollow). The nest at MPGR yielded two chicks and was central to the group’s activities. Even during the non-breeding season, when the group foraged at the extremes of its territory, they flew over 3 km to briefly visit the nest before returning to the foraging area. One bird at HBG was observed carrying nest lining, and the group showed interest in both nests until bees occupied them (N. Thomson 2012 pers. comm.). At LDNR, a year after the female joined the group, the nest sites became of interest. The nest at TT was visited almost daily by the group and became the centre of much of their social behaviour.
Veterinary management
Since two released individuals succumbed to Newcastle Disease Virus (NDV: pigeon paramyxovirus type 1), a reintroduction-tailored vaccine was developed (Koeppel and Kemp Reference Koeppel and Kemp2019). No other vaccines were administered.
Veterinary interventions were required for 14 birds. Sick birds masked initial symptoms, and even subtle changes in behaviour, appearance or appetite turned out to be significant indicators of declining health. Monitors assigned to each group were thus important for picking up subtle signs that enabled timely examination and intervention. Typical behaviour of sick birds included hiding in dense vegetation and withdrawing socially. They did not vocalise, stopped foraging and would often not accept supplementary food. Faeces and urates may be abnormal in colour and/or consistency, together with increased respiratory rate.
Post-release monitoring and maintenance:
Ground-hornbills are difficult to recapture and so, unlike other programmes where birds can be recaptured annually (e.g. Californian Condors; Walters et al. Reference Walters, Derrickson, Fry, Haig, John and Wunderle2008) for lead chelation and to replace transmitters and monitor the physical condition, it is not always possible to maintain these levels of monitoring. However, group movements are mainly cohesive on a permanent territory, so it is only necessary to track a single individual fitted with a telemetry unit to locate the group and check their daily status. Monitoring has been varied over time but, where possible, a daily visual check is a minimum requirement. Release birds remain wild and would not allow a monitor within 100 m of them before flying away. This flight distance hinders monitoring of these birds for foraging and social behaviour but ensured that they are unlikely to encounter direct harm from humans, although still vulnerable to indirect threats such as poisoning and electrocution.
Discussion
A review of a reintroduction programme is usually intended to assess the success of the species recovery programme, but this is not yet possible for such a long-lived species. It has been estimated that successfully reintroduced populations reach stability after 8–10 times the age of first breeding of the species concerned (Ewen et al. Reference Ewen, Soorae and Canessa2014), which for ground-hornbills would be 65–80 years. Defining the success of reintroductions is fraught with complexity, but it should focus on the success at the population level rather than at the individual level. A review of what defines reintroduction success by Jule et al. (Reference Jule, Leaver and Lea2008) found a consensus that a combination of criteria should be considered. However, not all these criteria appear assessable for the Southern Ground-hornbill at this stage.
Breeding by the first wild-born population: Since the only surviving offspring had not yet reached sexual maturity, it would take at least another five years (2020) before this can be tested.
An unsupported wild population of at least 500 individuals: This is now considered an arbitrary threshold as it does not take into account life-history traits, habitat quality or eventual meta-population structure, which varies widely between reintroduced populations. This is especially unlikely for top-order predators, where reattainment of criteria for a global conservation status of ‘Vulnerable’ is preferred (Hayward and Somers Reference Hayward and Somers2009).
Establishment of a self-sustaining wild population: This is a more pragmatic goal that would require production and recruitment to exceed adult mortaltity rates. However, since the Southern Ground-hornbill reintroduction is largely establishing founder groups this is realistically achievable.
The principal aim of any reintroduction is the long-term establishment, preferably without further intervention (Seddon et al. Reference Seddon, Sarrazin, Barbault, Beck, Griffith, Wolf, May and Gorman1999), and Gusset (Reference Gusset2009) argued that this is the survival of released individuals to breeding in the short-term, and persistence of the population in the long-term. The short-term objective has been achieved for the Southern Ground-hornbills. These can be modelled on the life-history traits of the target species to set realistic temporal goals for achievement, taking into account aspects of release sites and circumstances of release events. In addition, individual survival rates should be as great as, or greater than, source populations to ensure overall population growth (Gusset Reference Gusset2009). For Southern Ground-hornbills total annual mortalities (59%) were below expected losses of juveniles and sub-adults in the wild (70%, although these data may include dispersal; Kemp Reference Kemp1988), and include naive birds released unsuccessfully in Phases I and II. Phase III management matches the wild survival rate to adulthood. At maturity, annual turnover should drop to less than 1.5% per year if unnatural causes of mortality are eliminated (Kemp Reference Kemp1988).
The estimated release of just one founder group per year was sufficient to have a positive impact on the South African ground-hornbill population of 2,000 adults (Morrison et al. Reference Morrison, Daly, Burden, Engelbrecht, Jordan, Kemp, Kemp and Potgieter2005) but was considered unrealistic (Spear Reference Spear2005). However, this goal has been achieved successfully for three consecutive years, even in this experimental phase. Production can now be escalated to ensure that these criteria can be met at a larger and higher scale, after which the building blocks will be in place for reaching the third stage of success, population growth and persistence.
Population viability, however, requires more than just a sufficient population size and involves demographic, genetic, behavioural and ecological processes (Sarrazin and Barbault Reference Sarrazin and Barbault1996, Wynhoff Reference Wynhoff1998). Current success criteria rely heavily on reaching numerical indicators, but Reading et al. (Reference Reading, Miller and Shepherdson2013) went so far as to say that behaviour is the most important aspect influencing a reintroduced animal’s early post-release survival. Even success criteria that include behaviour are, however, deficient. Real-world practicalities, such as the bio-political environment and need for long-term funding, are possibly more important since even a complete knowledge of a species' complexities cannot be overcome without these factors.
Reintroductions of any long-lived species are management intensive, prolonged, and therefore costly over time. Even the highly successful Peregrine Falcon recovery programme in the United States needed 25 years to release 5,000 juveniles and establish the 100 breeding pairs that achieved a downlisting by IUCN Red Data List criteria (Brauning et al. Reference Brauning, Barber and McMorris2013).
Even if reintroductions may be discontinued in time, valuable conservation, behavioural and ecological lessons have already been learnt about the species through this process, which can be expeditiously applied where necessary in the future.
Lessons learnt from the reintroductions
Lesson 1: ‘Bush-schools’ can address the need for male helpers:
Group augmentation theory (e.g. Kokko and Johnstone Reference Kokko and Johnstone2001, Santema et al. Reference Santema, Taborsky, Kingma, Santema, Taborsky and Komdeur2014, Graf et al. Reference Graf, Gusset, Reid, Jansen van Rensburg, Slotow and Somers2006) explains why unrelated individuals in a cooperative group assist in rearing offspring other than their own, even when there are no kin-benefits. Groups benefit from the recruitment of young born into a group, or from unrelated dispersers, by their delayed reciprocity as helpers. This is sometimes taken to extremes, where non-kin are kidnapped to boost group size, as observed in White-winged Choughs Corcorax melanorhamphos (Heinsohn Reference Heinsohn1991), meerkats Suricata suricatta (Clutton-Brock Reference Clutton-Brock, Kappeler and van Schaik2006) and possibly Southern Ground-hornbills (A. Kemp 2009 in litt.). Wild Southern Ground-hornbill alpha males accept augmented non-kin young males. This was successfully done twice in Phase III and was evidenced by wild groups, including wild alpha males feeding in-situ captive birds. This suggests that group size is positively related to individual fitness of the alpha breeding pair, and probably also of other existing group members, and that group augmentation maintains groups at viable sizes.
The acceptance of non-kin individuals by wild or experienced mentor birds was used to train naive captive-reared birds in the wild, in what we term ‘bush-schools’. One juvenile male is added to each ‘bush-school’ group annually, where they follow the alpha male’s lead and mimic his behaviour. Mentor birds have been observed to ‘teach’ (Thornton and Raihani Reference Thornton and Raihani2010) young birds how to tackle difficult or dangerous prey items, such as tortoises or venomous snakes. The accepted juvenile male benefits from extensive social (e.g. territorial defence, dominance establishment) and reproductive (e.g. courtship and copulation behaviour, female and chick provisioning) learning, as they would in the wild. Once they have 3-4 years of experience, they are removed from the group and used as experienced mentors in the formation of new groups, augmented with a female and their own naive male helpers. Earlier experience suggested that alpha males are unable or unwilling to maintain more than one juvenile male at a time (Theron and Turner Reference Theron, Turner and Soorae2008). Furthermore, acceptance of non-kin individuals only holds true for young introduced birds (the oldest successful augmentation was an individual of 510 days old), while two attempts to augment wild-living groups with more mature birds led to the intruder being expelled or killed. Augmentation should, therefore, occur at the natural time of fledging in late austral summer or when the rains return the following season as success was significantly enhanced during the rainy season.
Lesson 2: The fate of females in male-skewed social groups:
Attempts to add a second female to a group with an established alpha female failed due to aggression from the alpha male, regardless of the season of release. No attempts were made to add second females to groups in Phase III.
Alliances were occasionally formed by subordinate females with beta-male group members, but these did not last as the alpha male would not tolerate beta males that showed interest in secondary females. Augmented females were excluded and, with time, chased to the periphery of the group territory. They then either left the group or trailed behind a group at a safe distance, behaviour that has also been seen in wild groups with more than one adult female (Kemp Reference Kemp1995).
Female exclusion happened to individuals from as young as post-fledging, which then had to survive alone, probably experiencing higher mortalities than males. The skewed adult sex ratio (Kemp and Kemp Reference Kemp and Kemp1980; Kemp Reference Kemp1988) suggests differential mortality of females, probably because they are evicted earlier and are more vulnerable out of the security of a group, but with the likelihood that they form the main dispersing sex.
Lesson 3: Novel threats to wild populations:
Three previously unknown threats to the wild population were discovered by frequent monitoring of reintroduced ground-hornbills. These threats were not taken into account during an initial Population and Habitat Viability Analyses (PHVA) (Spear Reference Spear2005, Morrison et al. Reference Morrison, Daly, Burden, Engelbrecht, Kemp, Kemp, Potgieter, Turner, Kemp and Kemp2007), or any other formal conservation planning for the species (Jordan Reference Jordan2011). Nor were they considered during earlier reintroduction plans.
Captive-reared birds were found to be susceptible to Newcastle Disease Virus (NDV: pigeon paramyxovirus type 1), with two mortalities related to birds scavenging on sick or dead wild doves (Abolnik et al. Reference Abolnik, Gerdes, Kitching, Swanepoel, Romito and Bisschop2008). It is unknown what the titres are of wild populations or if they have natural immunity, and whether NDV is a risk to wild populations.
Four birds were lost to electrocution on electrical transformer boxes, with three birds from the same group electrocuted at the same transformer box in five months, suggesting that other group members were unable to understand and/or learn about the threat. In each case, a beehive or paper-wasp nest was present on the transformer box, which may have initiated ground-hornbill curiosity since the group losing three birds had been unaffected in their previous years on the territory.
Three ground-hornbills tested positive for high lead levels and one was critically ill until chelation therapy was applied (Koeppel and Kemp Reference Koeppel and Kemp2015). Although it is known that vultures are susceptible to toxicosis via ingestion of lead fragments from hunting offal (Piper Reference Piper, Houston and Piper2005), this had not been recorded previously for ground-hornbills.
Lesson 4: Using redundant second-hatched chicks as release stock:
Southern Ground-hornbill reintroduction stock was either harvested wild redundant second-hatched chicks or the offspring from captive pairs. These nestlings were captive reared to fledging for release. To survive, candidates for release must be able to avoid predators, acquire and process food, interact appropriately with conspecifics, find suitable nests and orient and navigate in a complex and changing environment. In addition, fear and avoidance of humans are advantageous, a condition many animal husbandry managers find difficult to promote (Kleiman Reference Kleiman1989).
Captive-bred stock is widely used for reintroductions, but significantly more success is achieved with wild-to-wild translocations (Jule et al. Reference Jule, Leaver and Lea2008; Seddon et al. Reference Seddon, Armstrong and Maloney2007). Captive breeding is expensive, especially for a bird that takes nearly a decade to mature, and can erode the genetic basis for important morphological, physical and behavioural traits through artificial selection. The effects of improper socialising during captive-rearing negatively influenced survival ability. The species' natural learning and strong social preferences make them susceptible to habituation, both pre- and post-release, with reports of ready habituation even in the wild (e.g. KNP groups begging from tourist vehicles, a group in KwaZulu-Natal that shared the shade of a labourer's quarters in the heat of the day, or seeking the company of other animals e.g. impala Aepyceros melampus herds and chacma baboon Papio ursinus troops rather than being alone). The earlier released birds were habituated and although this did not lead to severe imprinting on humans, it did mean that release candidates showed little fear of humans. This led them to associate with humans and human-related objects such as vehicles, building windows and transformer boxes, which led to mortalities. Habituated birds became aggressive only at sexual maturity, attacking both people and moving vehicles, as also reported for captive Aceros hornbills (Coupe Reference Coupe1967).
Restoration of Californian Condors has shown that the quality of released individuals is fundamental for long-term sustainability (Meretsky et al. Reference Meretsky, Snyder, Beissinger, Clendenen and Wiley2001). Poor-quality release candidates were only successful with the addition of a wild mentor or mate and constant supplementary management. In Phase III, only ‘wild’ Southern Ground-hornbill candidates were released, which improved the reintroduction success.
While earlier captive-reared stock lacked behavioural competence, changes in husbandry in holding aviaries have seen improvements. Where possible, pre-release birds are kept in naturally structured social groups, with chicks fledged into captive groups. Here they learn how social dominance structures operate, being exposed to normal social, courtship and mating interactions. Pre-release artificial enrichment provided was aimed at the improvement of foraging skills. Providing individuals with opportunities to develop and improve such behavioural skills as predator avoidance, habitat selection, physical conditioning, problem-solving, hunting and appropriate imprinting conditioning, should increase reintroduction success (Reading et al. Reference Reading, Miller and Shepherdson2013).
Lesson 5: How to anchor a group:
Reintroductions of large, wide-ranging birds into selected ‘safe’ sites will be more complex than for mammals since they cannot be constrained by fences (Kemp Reference Kemp1995a). It is expected that re-introduced individuals will explore their surroundings, regardless of the quality of the habitat that they are released into. A buffer zone of 20 km around each release site is used to ensure that landowners and their staff are aware of, and support the programme, so cost-reduction for monitoring is an added incentive in assessing how best to anchor groups in ‘safe’ areas.
In soft-release aviaries, an effort is made to ensure that stress is kept to a minimum and social aggregation and compatibility promoted (as for social carnivores). Ground-hornbills were not released if they did not appear to be bonded with their cage contemporaries or, in the case of augmentations, with any previously released individuals.
Artificial nest structures are commonly used in reintroduction programmes, e.g. raptors (Clarke Reference Clarke and Soorae2008, Heinrich Reference Heinrich and Soorae2008) and parakeets (Ortiz-Catedral et al. Reference Ortiz-Catedral, Adams, Hauber, Brunton and Soorae2010). The nest site is important to ground-hornbills as a centre for social behaviour, forming the centre of their territorial movements, particularly in the breeding season (Zoghby et al. Reference Zoghby, Ryan, Little, Reid and Hockey2015). Nest sites may be used for many years, particularly where high-quality sites are at a premium (Kemp and Begg Reference Kemp and Begg1996). Artificial nests have been readily accepted by wild (Wilson and Hockey Reference Wilson and Hockey2013) and released groups (Theron and Turner Reference Theron, Turner and Soorae2008, Carstens et al. Reference Carstens, Kassanjee, Little, Ryan and Hockey2019a).
Lesson 6: Awareness raised:
The amount of interest in the species created by the reintroduction programme increases continuously. Even the earliest releases in Swaziland saw communities drawn into finding the poacher of a Southern Ground-hornbill, and interest expressed by local schools (Kemp Reference Kemp1995b). Decision-making processes for reintroduction were the catalyst for the first conservation-planning workshop in 2005, which included the formation of a national species Action Group and drafting a PHVA, which led ultimately to a Single Species Recovery Plan (Jordan Reference Jordan2011). The reintroduction programme provided the stimulus and momentum for extensive research into the genetics, behaviour, breeding biology, spatial land-use, cultural importance and health of Southern Ground-hornbills. For many of the land-owners engaged prior to a reintroduction or harvest, it boosted their interest in conservation in general and obtained their help in threat mitigation (use of poisons and lead ammunition, and the insulation of transformer boxes) and changes in land-use management, which also enhances these habitats for other species.
Conclusion and recommendations
No impact or reduction in viability of source populations has been found following the removal of redundant second-hatched chicks (Carstens et al. Reference Carstens, Kassanjee, Little, Ryan and Hockey2019a), supporting the 'insurance' hypothesis for obligate brood reduction (Morandini and Ferrer Reference Morandini and Ferrer2014). These chicks, despite captive rearing, were able to function as founder and mentor stock, bred successfully and reared a new generation, provided that behavioural factors were adequately addressed, and that the reintroduction happened during the rainy season.
Initial reintroductions showed that naive stock had a low chance of survival without constant management and that, without prevention of habituation to humans, behaviours were too aberrant. However, the use of wild-experienced alpha males to mentor a naive mate and helpers in ‘bush-schools’ has been done successfully three times. As a technique for ensuring correct behaviour in artificially reared individuals, this has considerable promise in supporting complex social learning requirements. Too few birds have been released so far to form viable subpopulations and so the isolated groups established need to be managed as part of a meta-population until core sub-populations can be developed. Establishment of new neighbouring groups to existing 'bush schools' becomes the next challenge.
There are compelling arguments to expand our understanding of demographics, social structures and reintroduction techniques for the species through carefully planned experimental reintroductions (as recommended by Taylor et al. Reference Taylor, Canessa, Clarke, Ingwersen, Armstrong, Seddon and Ewen2017). Uncertainties in reintroduction management explicitly tested either in the field or using modelling, should initiate full-scale production of Southern Ground-hornbill recruits. This would enable quantifiable growth at a population level by increasing the numbers of groups being released to establish viable core sub-populations (matching those suggested by Morandini and Ferrer Reference Morandini and Ferrer2017).
Supplementary Materials
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0959270920000131
Acknowledgements
This work is based on the pioneering studies of Alan and Meg Kemp, parents of the lead author. Kim Labuschagne and Thoko Rapinga assisted with permits. Angela Ferguson provided statistical analysis support. This research was funded by the National Zoological Gardens of South Africa (NZG), then under the management of the National Research Foundation and now the South African National Biodiversity Institute, as part of the Professional Development Programme of the National Research Foundation and National Department of Science and Technology (Grant number IUD80356). Further support was from the Mabula Ground Hornbill Project, chiefly funded by SASOL Ltd., Mabula Private Game Reserve, Disney Conservation Fund, Seaworld and Busch Gardens Conservation Fund, Rufford Grants and San Diego Zoo Global.













