


Introduction
The prevalence of infertility worldwide is estimated to affect around one in seven couples (NICE, 2004). The proportion of babies born from assisted reproductive technologies (ART) is increasing rapidly, and the numbers have quadrupled in the last 20 years (HFEA, 2009). To date, at least 3.5 million babies worldwide have been born following ART (de Mouzon, Reference de Mouzon2008). Despite these numbers, the impact of these treatments on the general human population is poorly understood.
There are three commonly used methods of ART: Intra-Uterine Insemination (IUI), In Vitro Fertilization (IVF) and Intra-Cytoplasmic Sperm Injection (ICSI). Intra-Uterine Insemination requires a catheter to deposit sperm directly into the uterus, which then swim through the fallopian tubes towards ovulated eggs. During IVF or ICSI, cumulus–oocyte complexes are aspirated from the ovaries after ovarian stimulation. Under IVF, oocytes are incubated with a number of motile spermatozoa. During ICSI the operator selects a single spermatozoon for direct injection into an egg that has been stripped of its cumulus cells. Embryos arising from IVF or ICSI are cultured up to 6 days in vitro (termed cleavage stage at 1–3 days and blastocyst stage thereafter) and are then transferred back to the patient (Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014). These methods of ART have been reported to differentially affect the sex ratio at birth (Tarin et al., Reference Tarin, Bernabeu, Baviera, Bonada and Cano1995, Reference Tarin, García-Pérez, Hermengildo and Cano2014; Menezo et al., Reference Menezo, Chouteau, Torello, Girard and Veiga1999; Luna et al., Reference Luna, Duke, Copperman, Grunfeld, Sandler and Barritt2007; Dean et al., Reference Dean, Chapman and Sullivan2010; Fedder et al., Reference Fedder, Loft, Parner, Rasmussen and Pinborg2013; Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014; Murakami et al., Reference Murakami, Egasghira, Tanaka, Mine, Otsubo and Kuramoto2014; Zhu et al., Reference Zhu, Zhuang, Chen, Liu and Qiao2015), with a general tendency for more male offspring to be born following IVF, and more females to be born after ICSI. Further, under IVF and ICSI, sex ratios have been reported to be more male biased after blastocyst-stage transfer compared with after cleavage-stage transfer (Chang et al., Reference Chang, Lee, Jee, Suh and Kim2009; Dean et al., Reference Dean, Chapman and Sullivan2010; Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014).
At reproductive age, sex ratio bias has the potential to generate substantial public health concerns (Pyeritz, Reference Pyeritz1998; Hesketh & Xing, Reference Hesketh and Xing2006; Dean et al., Reference Dean, Chapman and Sullivan2010; Hesketh & Min Reference Hesketh and Min2012; Shrivastava et al., Reference Shrivastava, Shrivastava and Ramasamy2014, Reference Shrivastava, Shrivastava and Ramasamy2015; Guilmoto, Reference Guilmoto2015), leading, for instance, to increased socially disruptive behaviour, aggression, transmission of sexually transmitted diseases and mental health problems (Tucker et al., Reference Tucker, Henderson, Wang, Huang, Parish and Pan2005; Zhou et al., Reference Zhou, Wang, Li and Hesketh2011, Reference Zhou, Wang, Zheng, Zhou, Li and Hesketh2012a, Reference Zhou, Yan and Heskethc; Bhattacharya, Reference Bhattacharya2013; Madan & Breuning, Reference Madan and Breuning2014; Moss & Maner, Reference Moss and Maner2016). Given that ART methodologies differentially affect the sex ratios of offspring produced, the focus here is on whether the sex ratios of ART-produced babies might adversely cause a general public health concern via indirect effects on the sex ratios produced by untreated members of the local population.
Models of population sex ratio
From the evolutionary perspective, much of the understanding of population sex ratios derives from the Düsing–Fisher theory of equal investment, until the cessation of parental expenditure, which is equivalent to equal sex ratio when sons and daughters are similarly costly to produce (Fisher, Reference Fisher1930; Bull & Charnov, Reference Bull and Charnov1988; Seger & Stubblefield, Reference Seger and Stubblefield2002; Mace & Jordan, Reference Mace and Jordan2005; West, Reference West2009; Song, Reference Song2014; Orzack et al., Reference Orzack, Stubblefield, Akmaev, Colls, Munné, Steinsaltz and Zuckerman2016). In essence, the ‘Fisherian’ argument is that in a population with a sex ratio that is biased (either towards males or towards females), offspring belonging to the rarer sex will have better mating prospects than those of the more common sex. Thus, parents with a genetic predisposition to produce more rare-sex progeny, whether facultatively or to a fixed degree, attain a higher than average number of grandchildren (≈evolutionary fitness), leading to the tendency to produce the rare sex becoming more widespread in the population. This response decreases the population sex ratio bias and also decreases the advantage associated with the production of rare sex progeny. Hence, frequency-dependent selection returns sex ratio biases to equality.
The Fisherian prediction only applies if a number of assumptions are met. These include that populations are large with mature offspring finding mates from throughout the population (panmixis) and that the relationship between fitness returns and resource allocation is identical for both offspring sexes (Fisher, Reference Fisher1930; Bull & Charnov, Reference Bull and Charnov1988; Seger & Stubblefield, Reference Seger and Stubblefield2002; Mace & Jordan, Reference Mace and Jordan2005; West, Reference West2009). Modifying the assumptions of the Düsing–Fisher approach has led to a large body of sex ratio theory covering the complexities of a range of organismal life-histories and predicting how reproducing individuals should respond to a range of scenarios including variations in, and perturbations to, local conditions (West, Reference West2009; Argasinki, Reference Argasinki2013).
A model of constrained sex allocation
Sex ratio models usually assume that parents are able to produce both sexes of offspring, but this may not always be the case (Gardner, Reference Gardner2014). Using essentially Fisherian assumptions, Godfray (Reference Godfray1990) developed a model that predicts the Evolutionarily Stable Strategy (ESS) progeny sex ratio of unconstrained (normal) mothers in populations that contain different proportions of mothers that are constrained to produce only sons, and with equivalent numbers of offspring produced by constrained mothers and by unconstrained mothers. While Godfray’s (Reference Godfray1990) model predictions are independent of the genetic mechanism of sex determination, constraints on sex allocation can be particularly apparent in haplodiploid species, which include many invertebrates (Godfray, Reference Godfray1990; Gardner, Reference Gardner2014). Under haplodiploidy unmated females can reproduce but can produce only male offspring, from unfertilized eggs, while mated mothers can produce both fertilized and unfertilized eggs, which develop into daughters and sons respectively. Godfray’s (Reference Godfray1990) model has hitherto been employed to understand sex allocation strategies in haplodiploid invertebrates but can also be applied to vertebrate species with non-haplodiploid sex determination (e.g. those with chromosomal [XX, XY] sex determination) to explore the consequences of mechanistic constraints on sex ratios produced by some mothers (Godfray, Reference Godfray1990).
Godfray’s (Reference Godfray1990) model predicts that in the absence of constrained females in the population, the ESS sex ratio of unconstrained mothers is 0.5 (as expected from Fisherian theory), with sex ratio defined as the proportion of a mother’s offspring that are male. More generally, when the proportion of females in the population constrained to produce only sons is p, the ESS sex ratio, r, of unconstrained mothers is given by:
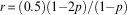 $$r\, {\equals}\, (0.5)(1{\minus}2p)/(1{\minus}p)$$
$$r\, {\equals}\, (0.5)(1{\minus}2p)/(1{\minus}p)$$
which is obtained by solving:
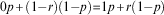 $$0p{\plus}(1{\minus}r)(1{\minus}p){\equals}1p{\plus}r(1{\minus}p)$$
$$0p{\plus}(1{\minus}r)(1{\minus}p){\equals}1p{\plus}r(1{\minus}p)$$
This predicts that unconstrained mothers should respond to reproduction by constrained females by producing progressively more daughters among their own offspring (lower sex ratios) as the proportion of females that are constrained increases. The model assumes that individual unconstrained mothers employ fixed sex allocation and the mating environment is constant but the same predictions are recovered if it is assumed that unconstrained mothers have facultative sex allocation and the mating environment is variable (Gardner, Reference Gardner2014). Godfray (Reference Godfray1990) considered proportions up to p=0.5, in which case half of the mothers in the population are constrained and producing only sons and the other half are unconstrained but are selected to produce only daughters (Fig. 1, upper panel, where the boldest solid line meets the x-axis).

Fig. 1 Evolutionarily Stable Strategy (ESS) sex ratios of unconstrained females in the presence of a range of proportions of constrained females in the population and to varying degrees of constraint, c, of these females. Constrained females may be limited to produce more sons than daughters (c>0.5) or more daughters than sons (c<0.5), with c=0.5 representing no constraint. The upper panel shows predictions for the full range of possible values of c: for c=0 constrained females can only produce daughters while for c=1 they can only produce sons (the latter matching the assumptions and predictions of Godfray’s Reference Godfray1990 model). The lower panel shows results for values of c close to c=0.5, similar to those estimated from clinical data (Fig. 2).
Methods
A model of partial-constraint
Here Godfray’s (Reference Godfray1990) model is modified to take into account the fact that the degree of constraint experienced by mothers may not be all-or-nothing and that constraints may operate in either direction. Specifically, situations are considered where some females are constrained to produce abnormally male-biased or female-biased progeny sex ratios, as observed under human ART treatments (Dean et al., Reference Dean, Chapman and Sullivan2010; Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014; Zhu et al., Reference Zhu, Zhuang, Chen, Liu and Qiao2015): this is termed ‘partial-constraint’. Godfray’s assumption that the numbers of offspring produced by unconstrained and constrained mothers are equivalent is retained for simplicity. We note that in practice, couples reproducing by ART are likely to have fewer offspring than couples that do not require ART, but also the incidence of monozygotic twinning can be increased by ART (Chang et al., Reference Chang, Lee, Jee, Suh and Kim2009), which will act to increase any sex ratio biasing effect of treatment. While this modelling is developed with reference to human sex ratios, it may also be applicable to invertebrates exhibiting intermediate degrees of constraint (Chevrier & Bressac, Reference Chevrier and Bressac2002). The purpose is to explore how (1) the degree to which some females are constrained and (2) the direction of the constraint, as well as (3) the proportion of constrained females in a population, might influence the sex ratios produced by unconstrained females, via frequency-dependent natural selection.
The modified model contains a term to represent the degree to which constrained females are constrained, and the direction of the constraint, c (0≤c≤1), is the proportion of males produced by constrained females. Thus c=1 indicates a constraint to produce males only, as assumed by Godfray (Reference Godfray1990), and c=0.5 indicates that nominally ‘constrained’ females are effectively unconstrained and c=0 indicates a constraint to produce females only. The term c is therefore also equal to the progeny sex ratio of constrained females when expressed as the proportion of offspring that are male. The modified model is:
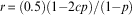 $$r\, {\equals}\, (0.5)(1{\minus}2cp)/(1{\minus}p)$$
$$r\, {\equals}\, (0.5)(1{\minus}2cp)/(1{\minus}p)$$
which is obtained by solving:
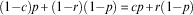 $$(1{\minus}c)p{\plus}(1{\minus}r)(1{\minus}p)\, {\equals}\, cp{\plus}r(1{\minus}p)$$
$$(1{\minus}c)p{\plus}(1{\minus}r)(1{\minus}p)\, {\equals}\, cp{\plus}r(1{\minus}p)$$
Note that when c=1, Eqn (4) simplifies to Eqn (2).
Results
The modified model predicts that the proportion of constrained females in the population, the degree to which these females are constrained and whether any constraint is towards the production of male or female offspring, will all influence the sex ratios produced by unconstrained females (Fig. 1). If constrained females can only produce sons, the results are identical to Godfray’s original model (Godfray, Reference Godfray1990), but if the degree of constraint to produce males is less extreme, such that constrained females are producing some daughters along with a majority of sons, then the sex ratios of unconstrained females are less affected (Fig. 1, upper and lower panels, lines for c>0.5 to c=0.9).
For cases of partial-constraint (0<c<1) it is informative to consider the influence of larger proportions of constrained females in the population than under Godfray’s assumption of c=1 (in Godfray’s model, when p>0.5 all unconstrained females are selected to produce only sons, Fig. 1): the less the degree of constraint (values closer to c=0.5), the larger the proportion of constrained mothers needs to be for unconstrained females to be selected to produce only one sex of offspring (Fig. 1). Further, considering constraints to produce female-biased sex ratios (c<0.5) as well as constraints to produce predominantly sons (c>0.5), shows that the sex ratio response of unconstrained mothers is symmetrical around c=0.5 (Fig. 1).
The indirect effects of ART-induced sex ratio biases
Further to illustrating sex ratio responses to some ‘round figure’ values of c (Fig. 1), values of c estimated from clinical studies of ART can be used. The upper panel of Fig. 2 shows predictions for five estimates of c from data collated across all fertility clinics in Australia and New Zealand on 13,368 babies born following treatment from 2002 to 2006 (Dean et al., Reference Dean, Chapman and Sullivan2010): the overall sex ratio of babies born following single-embryo transfer (SET) ART (0.513) and the four estimates for the specific ART regimes (ICSI and IVF, ranging from 0.487 to 0.561). Similarly, the lower panel of Fig. 2 shows predictions for six values of c estimated from data collected from 106,066 babies born between 2000 and 2010 in the United Kingdom following ART (Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014). These comprise the five estimates for the specific ART regimes (ICSI, IVF and IUI, ranging from 0.488 to 0.539) and the overall mean (0.507). Both panels also show a reference line for c=0.5, which is predicted to elicit no change in the sex ratio produced by unconstrained females. Note that only ICSI using cleavage stage embryo transfer is predicted to select for male-biased sex ratios among unconstrained mothers (because this is the only ART technique that generates a female bias among patients’ progeny) and that because a mixture of techniques are employed in each country the overall effect of utilizing ART will typically be to select for male bias among the untreated population (Fig. 2).

Fig. 2 Evolutionarily Stable Strategy (ESS) sex ratios of unconstrained females in the presence of a range of proportions of constrained females in the population with degrees of constraint, c, estimated from national-scale clinical data following different ART procedures. Predictions for the mean values of c and for c=0.5 (no constraint) are also shown. Values of c in the upper panel derive from clinics across Australia and New Zealand (Dean et al., Reference Dean, Chapman and Sullivan2010) and values in the lower panel derive from clinics across the UK (Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014).
Discussion
Human sex ratio research must be interdisciplinary if it is to be successful (Lazarus, Reference Lazarus2002).
Assisted reproductive technologies have been reported to directly affect the sex ratios of babies born (Dean et al., Reference Dean, Chapman and Sullivan2010; Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014; Zhu et al., Reference Zhu, Zhuang, Chen, Liu and Qiao2015; but see Orzack et al., Reference Orzack, Stubblefield, Akmaev, Colls, Munné, Steinsaltz and Zuckerman2016). There are numerous ways in which sex ratio bias could affect public health and social well-being (Pyeritz, Reference Pyeritz1998; Tucker et al., Reference Tucker, Henderson, Wang, Huang, Parish and Pan2005; Hesketh & Xing, Reference Hesketh and Xing2006; Zhou et al., Reference Zhou, Wang, Li and Hesketh2011, Reference Zhou, Wang, Zheng, Zhou, Li and Hesketh2012a, Reference Zhou, Yan and Heskethc; Hesketh & Min, Reference Hesketh and Min2012; Bhattacharya, Reference Bhattacharya2013; Madan & Breuning, Reference Madan and Breuning2014; Shrivastava et al., Reference Shrivastava, Shrivastava and Ramasamy2014, Reference Shrivastava, Shrivastava and Ramasamy2015; Guilmoto, Reference Guilmoto2015). Here, potential influences of the observed sex ratio effects of ART (on the offspring of treated patients) on the sex ratios produced by the general (unassisted) population were explored employing an evolutionary approach based on frequency-dependent sex allocation strategies. This modelling suggests that even if the ART treatments carried out were of the type that leads to the greatest sex ratio bias (IVF at the blastocyst stage in Australian and New Zealand populations, c=0.561, Fig. 2), well over 20% of mothers in the population would have to reproduce via ART for the unconstrained mothers to be selected to produce progeny sex ratios that would be noticeably deviant from equality. This is greater than the estimated prevalence of human infertility problems (one in seven couples, 14.3%; NICE, 2004). It would take almost 90% of reproduction in the Australian and New Zealand population, and more than 90% in the UK population, to be via this specific type of ART before untreated mothers would be selected to produce female offspring only. Given that several different ART techniques are utilized, each leading to different degrees and directions of sex ratio bias and that, currently, at most 4% of babies are born following ART treatment (Dean et al., Reference Dean, Chapman and Sullivan2010), the putative indirect influence of ART on the birth sex ratio of untreated members of the population can be considered currently negligible (see also Orzack et al., Reference Orzack, Stubblefield, Akmaev, Colls, Munné, Steinsaltz and Zuckerman2016).
The model used to predict the response of unconstrained mothers to assisted reproduction by other mothers adopts Fisherian assumptions concerning population mating patterns and evolutionary fitness returns on investment. Full conformity to Fisherian assumptions is probably a biological rarity (Bull & Charnov, Reference Bull and Charnov1988). In particular, human sex allocation may be affected by sexually differential fitness returns (Bereczkei & Dunbar, Reference Bereczkei and Dunbar1997; Lazarus, Reference Lazarus2002; Mace & Jordan, Reference Mace and Jordan2005; Almond & Edlund, Reference Almond and Edlund2007; James, Reference James2012, Reference James2013): predicting how these might influence the sex ratio response of unconstrained parents to the presence of individuals reproducing via ART would not be straightforward (West, Reference West2009) and key information on parental investment is currently lacking (Orzack et al., Reference Orzack, Stubblefield, Akmaev, Colls, Munné, Steinsaltz and Zuckerman2016). Further, human population sex ratios at conception may adhere to the ‘baseline’ expectation of 0.5 (Orzack et al. Reference Orzack, Stubblefield, Akmaev, Colls, Munné, Steinsaltz and Zuckerman2016) but at birth are typically slightly male biased, ca. 0. 513 (Mace & Jordan, Reference Mace and Jordan2005; Almond & Edlund, Reference Almond and Edlund2008; Dean et al., Reference Dean, Chapman and Sullivan2010; ONS, 2011; James, Reference James2013; Maalouf et al., Reference Maalouf, Mincheva, Campbell and Hardy2014).
Theory developed to complement the Düsing–Fisher approach indicates that sex ratios of local sub-populations should influence sex ratio evolution (Argasinski, 2013). Moreover, human reproductive behaviour has been reported to respond to local sex ratio bias in a range of ways (Chipman & Morrison, Reference Chipman and Morrison2013) including overproduction of the rarer sex (Lummaa et al., Reference Lummaa, Merila and Kause1998; Ranta et al., Reference Ranta, Lummaa, Veijo and Merila2000; Lazarus, Reference Lazarus2002; Helle et al., Reference Helle, Kaar, Helama and Jokela2008; but see James, Reference James2000), overproduction of the rarer sex particularly supporting the notion that sex ratios of untreated members of the population could be affected indirectly by the practice of ART. These reports derive from studies within the framework of evolutionary ecology but analogous frequency-dependent responses to sex ratio bias are also reported by social scientists whose discipline encompasses the complex array of behavioural and social processes that shape human reproductive decisions (Bhattacharya, Reference Bhattacharya2013; Zhou et al., Reference Zhou, Zhou, Wang, Zheng and Hesketh2013). Of partiular note is evidence that human birth sex ratio perturbations in China between 1962 and 1964 caused substantial and opposite effects among the progeny of mothers born during this period, indicating adaptive inter-generational sex ratio adjustment (Song, Reference Song2014).
The proximate (physiological) mechanism(s) by which individual humans might adjust sex allocation in response to local population sex ratios are not well understood (Lummaa et al., Reference Lummaa, Merila and Kause1998). There could be assessment of the current adult sex ratio, perhaps based on time to fertilization (e.g. delayed mating) (Werren & Charnov, Reference Werren and Charnov1978; Godfray, Reference Godfray1990; West, Reference West2009), coupled with hormonal changes influencing offspring gender (James, Reference James2011, Reference James2012; Setti et al., Reference Setti, Figueira, Braga, Iaconelli and Borges2012). Alternatively, parents may respond to the sex ratio of the preceeding chort (James, Reference James2000; Helle et al., Reference Helle, Kaar, Helama and Jokela2008). Current data support that human birth sex ratios are affected by the childhood experience of parents (Song, Reference Song2014). Further, members of human communities are often concious of, and concerned about, local sex ratio bias (Mackenzie et al., Reference Mackenzie, Lockridge and Keith2005; Shrivastava et al., Reference Shrivastava, Shrivastava and Ramasamy2014, Reference Shrivastava, Shrivastava and Ramasamy2015), which can lead to deliberate measures to alter sex ratios towards equality (Hesketh et al., Reference Hesketh, Lu and Xing2011; Hesketh & Min, Reference Hesketh and Min2012; Zhou et al., Reference Zhou, Wang, Zhou and Hesketh2012b; Bhattacharya, Reference Bhattacharya2013), while other members of a population may be seeking to increase the probablity of producing offspring of a particular sex by the various means available to them (Madan & Breuning, Reference Madan and Breuning2014; Guilmoto, Reference Guilmoto2015). Sex-specific abortion has been a common method in some societies and has led to sex ratio skew in several countries (Hesketh & Min, Reference Hesketh and Min2012; Zhou et al. Reference Zhou, Wang, Zhou and Hesketh2012b; Madan & Breuning, Reference Madan and Breuning2014; Song, Reference Song2014) but ART could potentially be empoyed, generating considerable ethical concerns (Guilmoto, Reference Guilmoto2015).
This study has used an evolutionary ecology approach to explore potential responses to sex ratio bias, i.e. one that predicts, using principles of genetic evolution, how individuals would be selected to behave in order to maximize their fitness. There is ongoing debate over the extent to which this approach can be applied to humans and human sex ratios due, for example, to the importance of cultural factors and conscious decision-making (Frank, Reference Frank2002; Mace & Jordan, Reference Mace and Jordan2005; West & Burton-Chellew, Reference West and Burton-Chellew2013) and recent analysis has shown how cultural effects can confound and obscure adaptive sex ratio adjustment (Song, Reference Song2014). Thus, we do not claim that our model applies exactly to human sex ratios. Rather, predictions are presented heuristically to provide a tractable guide to the approximate degree of effect that might be expected in the human population generally in response to ART being carried out on a specific proportion of mothers. Dean et al. (Reference Dean, Chapman and Sullivan2010) cautioned that increasing use of ART may have a major public health impact via the sex ratio (Dean et al., Reference Dean, Chapman and Sullivan2010). The new model suggests that such impact will be largely confined to effects on the sexual composition of the families receiving treatment (who should be made aware during counselling; Chang et al., Reference Chang, Lee, Jee, Suh and Kim2009) and that the treatment they undergo will not have appreciable indirect effects on members of the wider population.
In conclusion, ART procedures are associated with deviations in the sex ratios of babies born. However, unless ART becomes very considerably more common in human populations, these gender biases are unlikely to constitute a major public health concern, at least in terms of reproduction by unconstrained parents, because any sex ratio response by unassisted members of the population is expected to be too small to discern.
Acknowledgments
For help and discussion thanks are due to Simon Gates, W. H. James, John Lazarus, Adrian Pratt and an anonymous referee.


