


Saturated fatty acids (SFA) have been associated with heart disease; the ingestion of milk fat, with its high proportion of SFA (70–75%), has therefore been a subject of concern. About 2% of milk fatty acids (FA) are, however, polyunsaturated (PUFA) (Elgersma et al. Reference Elgersma, Tamminga and Ellen2006), which could help maintain human health. Different studies have examined the role of nutrition in lactating cows as a means of modifying the FA profile of milk, i.e., reducing the proportion of SFA and increasing that of PUFA, mainly conjugated linoleic acid (CLA). Diet plays an important role in determining the degree of unsaturation of cow milk fat. Long chain FA are particularly affected since ruminants cannot synthesize them; the diet is therefore the only source (Elgersma et al. Reference Elgersma, Maudet, Witkowska and Wever2005). It is well known that, compared with non-grazing cows, grazing cows produce milk with a FA profile favourable to human health, with higher proportions of CLA, vaccenic (VA) and linoleic acids (Dewhurst et al. Reference Dewhurst, Shingfield, Lee and Scollan2006). This invests fresh forage with added value and highlights grazing as a worthwhile natural production system. However, grazing alone cannot meet the nutritional requirements of high production dairy cows, particularly in early lactation, and supplementation is required to provide the energy required for an animal's genetic potential to be fully reached. Supplementation with total mixed ration (TMR) could help produce high yields of milk with enhanced protein and fat contents, with the grazing component allowing a better milk FA profile to be achieved, moreover, the economic cost of this production system could be lower. The effects of feeding TMR and pasture diets have been previously reported (Fontaneli et al. Reference Fontaneli, Sollenberger, Littell and Staples2005), and associate grazing with a smaller dry matter intake and lower milk production compared with intensive systems. In addition, the effects of pasture feeding and TMR feeding systems on milk FA composition have been reported by several authors (Kay et al. Reference Kay, Roche, Kolver, Thomson and Baumgard2005; Vibart et al. Reference Vibart, Fellner, Burns, Huntington and Green2008). However, the intensity of grazing that can be allowed for achieving a better FA profile without compromising milk yield has not been checked in areas with an oceanic climate. This is found along European west coasts and it is characterized by summers generally humid and warm, and winters fairly mild. These climatic conditions allow grazing all year round, and for this reason, pasture based systems have been traditional across the Atlantic Europe. Now, it is necessary to balance the production capacity of dairy cows with the use of grass in their diet without compromising milk production. This study has been directed to obtain milk with quality with low inputs, which makes farm exploitation based on pasture more profitable and sustainable. The aim of this study was to evaluate different strategies of limited grazing associated with TMR feeding on the FA profile of milk, feed intake and milk production.
Materials and Methods
Cows, diet and pasture
This work was undertaken in the spring of 2007, adhering to the standards of the European Union Animal Welfare Directive Number 86/609/EEC. Fifteen Holstein cows with 94±44 days in milk were selected. The milk production of these cows was 34·8±6·8 l/d in the month prior to the start of the experiment. TMR was formulated according to NRC (2001) requirements for dairy cattle (Table 1). Additionally, two concentrates, A and B, were distributed as energetic supplements in an automatic feeder. Concentrate A was offered at 2·0 kg/d per cow and concentrate B adjusted to 0·5 kg/l of milk produced above 30 l/d. Six 1·5 ha paddocks with a wide range of grasses (Agrostis tenuis, Dactylis glomerata, Lolium perenne), legumes (Trifolium repens, Viccia cracca) and other species provided the grazing area. Water was always available.
Table 1. Composition (% DM) of the total mixed ration (TMR) and concentrates

Experimental design
The animals were randomly assigned to three treatment sequences in a changeover design. Each trial period lasted 21 days, including 14 for adaptation and seven for data collection. The treatments were: 1) 12 h grazing (TMR12; 07·30 h to 19·30 h) plus TMR; 2) 6 h grazing (TMR06; 12·30 h to 18·30 h) plus TMR; and 3) no grazing time (TMR00) plus TMR. The TMR was offered ad libitum indoors in all treatments. Two adjoining paddocks were used in each experimental period, one for the adaptation stage, the other for the data collection stage. Both paddocks were divided into two plots by mobile electric fences, corresponding to the TMR06 and TMR12 treatments. On the day prior to the start of the trial, pasture yield and dry matter yield were measured to determine pasture availability and to set the grazing area of the paddocks (on the basis of herbage yield) according to the grazing time permitted. All cows were weighed on the first and last day of each period after morning milking. All were milked twice daily from 07:00–08:00 h and 19:00–20:00 h. After morning milking, the TMR12 cows were moved to their corresponding plot, where they stayed until evening milking. The TMR06 cows remained indoors until 12:30 h, and were then moved to their corresponding plot until evening milking. The TMR00 cows were kept indoors throughout the day.
The TMR intake of individual animals was recorded using a computerised system (Bach et al. Reference Bach, Iglesias and Busto2004). TMR refusals were removed and weighed daily. Herbage intake in the TMR06 and TMR12 treatments were estimated using NRC prediction equations (Macoon et al. Reference Macoon, Sollenberger, Moore, Staples, Fike and Portier2003). Briefly, energy requirements were recorded as net energy (NE Mcal/d) requirements for maintenance, lactation, body weight changes, walking and grazing. The NE from pasture intake was estimated as total NE requirements minus the NE supplied by the TMR and concentrate intakes.
Sample collection and analysis
TMR samples and orts were taken daily during the measurement period. Concentrates and pasture were sampled at the beginning of each trial period. TMR and forage samples were dried (60°C, 24 h) and ground (0·75 mm); concentrate samples were ground through a 1 mm screen. Dry matter (DM), ash, crude protein (CP), ether extract and starch were determined by near infrared spectroscopy (NIRS; FOSS NIRSystem 5000). Neutral and acid detergent fibres (NDF, ADF) were analysed as described by Van Soest et al. (Reference Van Soest, Robertson and Lewis1991). Calcium was determined by AA spectrophotometry (Perkin Elmer 3030B) and phosphorus by UV-VIS spectrophotometry (Perkin Elmer λ35). The FA content of the feeds was analysed according to Sukhija & Palmquist (Reference Sukhija and Palmquist1988) with the modifications of Palmquist & Jenkins (Reference Palmquist and Jenkins2003). The esterification of FA was performed using hexane and 10% methanolic HCl as follows: heating at 70°C in a water bath for 2 h, cooling and adding 2 ml of hexane and 5 ml of 6% K2CO3, and centrifuging for 5 min at 500 g.
Milk production was measured and the milk sampled daily at both milking times over the seven data collection days of each trial period. Milk samples were analysed for fat and protein contents (MilkoScan FT 6000). For milk FA analysis, the fat was isolated as described by Feng et al. (Reference Feng, Lock and Garnsworthy2004) and transesterified as described by Christie (Reference Christie1982) with the modifications of Chouinard et al. (Reference Chouinard, Louise, Barbano, Metzger and Bauman1999). Two millilitres of hexane were added to 40 mg milk fat. This was then added to 40 μl sodium methylate in methanol and incubated at room temperature for 10 min before adding 60 μl of a solution of 1 g oxalic acid in 30 ml diethyl ether. These samples were then centrifuged for 5 min at 2400 g at 5°C. FA methyl esters were determined by gas-liquid chromatography (Varian 4000 GC/MS, Inc. Palo Alto, CA, USA) in both milk and feed samples. Methyl esters were separated using a 100 m×0·25 mm i.d. fused silica capillary column (CP-Sil 88 Varian). Individual FA peaks were identified by comparison of their retention times and mass spectra with those of pure methyl ester standards (Matreya Inc., PA.; Sigma-Aldrich Inc.).
Statistical analysis
Differences in the chemical composition of the pasture between trial periods were examined using the GLM procedure (SAS, 1999), recording the effect of period and the residual error. Feed intake and milk data were analysed using the GLM procedure (SAS, 1999) according to the model: Yijk=μ+Ai+Pj+Tk+Eijk, where Yijk=the dependent variable, μ=the overall mean, Ai=the effect of treatment, Pj=the period, Tk=the effect of the animal, and Eijk=the residual error. Means were separated using the Tukey test.
Results
Chemical composition of feedstuffs and pasture
Table 2 shows the chemical compositions and FA profiles of the TMR, concentrates and herbage for each trial period. No differences were seen between the trial periods with respect to the nutritive values of the TMR or concentrates. The chemical composition of the pasture, however, changed over the course of the experiment. During trial period 3, the pasture showed its lowest NDF and ADF contents, while the lowest DM and organic matter levels and the highest CP levels were seen during trial period 2. Nearly 95% of the FA of the TMR and both concentrates was made up of palmitic (C16:0), oleic (C18:1) and linoleic (C18:2) acids. The main FA of the pasture consumed were linolenic (C18:3), C16:0 and C18:2. Linear increases in C18:1 and C18:2 were observed over the course of the experiment.
Table 2. Chemical compositions (% on DM) and fatty acid profiles of the total mixed ration (TMR), concentrates and pasture in the different trial periods
Values are means for n=3

a,b,c Values in the same row with different letters differ significantly
* =P<0·05; **=P<0·01
† NS: Not significant
§ ND: Not detected
¶ NA: Not analysed
Feed intake and body weight changes
Body weight changes ranged from −0·3 (TMR06) to 1·6 kg (TMR00), with no significant differences between treatments. The dry matter intake (DMI) associated with TMR decreased significantly when the time available for grazing increases (Table 3). No significant differences in the DMI associated with concentrate A or concentrate B were seen between any treatment. The pasture availability was 19·8 and 39·7 kg/d per cow for the TMR06 and TMR12 treatments respectively. The DMI afforded by grazing, estimated as the difference between requirements and that supplemented by TMR and concentrates, was 60% higher in the TMR12 treatment than in the TMR06 treatment. However, no significant differences were seen between treatments with respect to total DMI.
Table 3. Body weight, dry matter (DMI) and FA intake for the three treatments
Values are means for n=15 for body weight values, n=105 for the other variables

a,b,c Values with different letters differ significantly
** =P<0·01; ***=P<0·0001
NS: Not significant
The fat intake was significantly affected by the different treatments, ranging from to 0·71 kg/d in the TMR12 treatment to 0·83 kg/d in the TMR00 treatment (P<0·01). In fact, individual FA intake was different between treatments, except for lauric acid (C12:0), and myristic acid (C14:0). The TMR00 treatment was associated with higher intakes of C6:0, C16:0, C18:0, C18:1 and C18:2 than either of the grazing treatments; these intakes decreased progressively as grazing time increased. The C18:3 intake was directly related to its herbage content.
Milk production and milk composition
Table 4 shows the milk yield and composition results. No differences were seen between treatments with respect to milk production, milk protein content or protein yield. The milk fat content of the TMR12 milk was significantly lower than that of the TMR00 or TMR06 milk. This explains the low fat yield of the TMR12 milk.
Table 4. Milk production, composition and FA profile for the three treatments
Values are means for n=105
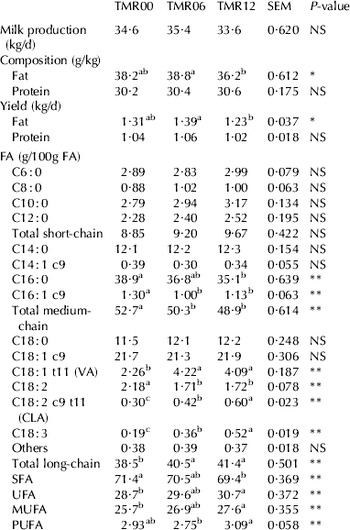
a,b,c Values in the same row with different letters differ significantly
* =P<0·05; **=P<0·01
NS: Not significant
Milk fatty acids
No significant differences were seen between treatments with respect to the short-chain FA content of the different milks. The medium chain FA content was highest in the TMR00 milk since the C16:0 and palmitoleic acid (C16:1) content decreased as grazing time increased. The long-chain FA content was higher in the TMR06 and TMR12 milk than in the TMR00 milk, a consequence of the formers' high VA (C18:1 t11), CLA (C18:2 c9 t11) and C18:3 contents. The VA content of the TMR06 and TMR12 milk was twice that of the TMR00 milk, with no significant difference between the TMR06 and TMR12 milk. The CLA content was higher in the milk produced in the grazing treatments than that produced in the TMR00 treatment. Indeed, the TMR12 milk had a 50% and 30% higher CLA content than the TMR00 and TMR06 milk respectively. In addition, the C18:3 content was 32% higher (P<0·01) than in the TMR06 milk, and nearly three times that of the TMR00 milk. However, the grazing treatments led to a reduction in milk C18:2 content compared with the TMR00 treatment. The proportion of unsaturated FA (UFA), including monounsaturated FA (MUFA) and PUFA, was higher in the TMR12 milk than in than TMR00 milk. The TMR06 milk showed an intermediate value.
Discussion
In the present work, the different treatments had no effect on milk production. This agrees with that reported by other authors who examined the feeding of cows with TMR plus the grazing of pasture (Loor et al. Reference Loor, Soriano, Herbein and Polan2003) or winter oats (Schroeder et al. Reference Schroeder, Delahoy, Vidaurret, Bargo, Gagliostro and Muller2003) grazing. This absence of any difference in milk yield might be explained by the adequate net energy intake of the grazing cows, confirmed by the absence of variation in live weight. The reduction in milk fat content seen with the TMR12 treatment has been reported by other authors (Loor et al. Reference Loor, Soriano, Herbein and Polan2003; Elgersma et al. Reference Elgersma, Ellen, Van der Horst, Mouse, Boer and Tamminga2004) after switching cows from TMR to fresh forage.
As seen in the present work, Schroeder et al. (Reference Schroeder, Delahoy, Vidaurret, Bargo, Gagliostro and Muller2003) reported a lower C16:0 and a higher C18:1 content in the milk of grazing cows than in that of cows fed TMR. Similarly, Elgersma et al. (Reference Elgersma, Ellen, Van der Horst, Mouse, Boer and Tamminga2004) reported that switching cows from fresh pasture to TMR during the winter increased the C16:0 content of their milk from 22·6 to 34·8 g/100 g FA. Ward et al. (Reference Ward, Wittenberg, Froebe, Przybylski and Malcolmson2003) observed lower concentrations of C16:0 in milk when increasing the DMI provided by fresh pasture from 50 to 80%. In the present study, the greater intake of C16:0 in the TMR00 treatment than in the TMR06 or TMR12 treatment might explain the slightly higher C16:1 content of the TMR00 milk; C16:1 can be produced through the desaturation of C16:0 via the action of delta-9 desaturase (Peterson et al. Reference Peterson, Kelsey and Bauman2002).
The low proportion of C18:2 in milk from the TMR12 cows can be explained by the fall in C18:2 intake and by its hydrogenation in the rumen. The latter could have contributed to an increase in the VA, CLA and stearic acid contents of the milk as a result of their synthesis from C18:2 in the rumen. Some authors have reported similar results when feeding cows fresh forage (Ferlay et al. Reference Ferlay, Martin, Pradel, Coulon and Chilliard2006), when grazing cattle (Bargo et al. Reference Bargo, Delahoy, Schroeder and Muller2006), and when feeding with fish and plant oils (AbuGhazaleh et al. Reference AbuGhazaleh, Felton and Ibrahim2007).
In this study, the pasture intake reduced the proportion of SFA and increased that of UFA (mainly MUFA) in the milk. This might be explained by the higher intake of C18:2 and C18:3 in fresh forage and their biohydrogenation in the rumen. With a pattern similar to that seen in the present work, Bargo et al. (Reference Bargo, Delahoy, Schroeder and Muller2006) reported high intakes of C18:3 and C18:2 in cows feeding on pasture. Bauman et al. (Reference Bauman, Baumgard, Corl and Griinari2000) showed that ruminal lipolysis and free FA biohydrogenation led to drastic reductions (70–90%) in dietary PUFA via their transformation into trans-isomers of MUFA (especially VA) or SFA (mainly stearic acid).
The synthesis of CLA is an important factor to consider in nutritional strategies involving high proportions of fresh forage since it provides PUFA, of which C18:3 makes up some 50–75% (Elgersma et al. Reference Elgersma, Maudet, Witkowska and Wever2005). The supplementation of TMR with pasture provides additional C18:2, a direct precursor of CLA and VA, while C18:3 increases the proportion of VA in the rumen. In turn, VA is an important substrate that can be converted to CLA in the mammary gland via the action of delta-9 desaturase (Griinari et al. Reference Griinari, Corl, Lacy, Chouniard, Nurmela and Bauman2000). This may also explain the high VA, CLA and C18:3 contents in the TMR12 milk. However, a slightly higher VA concentration was seen in the TMR06 milk than in the TMR12 milk, despite the cows of the latter treatment having 6 h more grazing time and therefore enjoying greater pasture availability and C18:3 intake. The TMR12 treatment should have provided an even higher C18:3 and C18:2 intake, leading to the highest levels of VA and CLA. However, the VA might have been more effectively absorbed and desaturated in the mammary tissue to produce CLA, thus explaining the higher concentration of CLA and slightly lower concentration of VA than seen in the TMR06 milk.
The relationships between forage quality and the meadow effect in each trial period are difficult to explain. The C18:3 concentration is higher in grass than in legumes. Moreover, the lipids of forage are located in the leaf chloroplasts; a high proportion of leaves in the diet might therefore lead to a high intake of C18:3. The paddocks used in the present study, although adjacent and with similar botanical compositions, differences in terms of species proportions or plant maturity might therefore have influenced the C18:1 and C18:2 contents of the forage (Elgersma et al. Reference Elgersma, Maudet, Witkowska and Wever2005).
In these climatic conditions, grazing is allowed all year round. This fact allows savings in the cost of food in the barn, which could provide increased profitability to dairy farms. Moreover, the results show that the use the grass does not reduce milk yield, and fat is healthier also, which could lead to an increase in value added of milk produced in this system.
In conclusion, the intake of fresh herbage through grazing as a supplement to TMR could be used as a strategy for modifying the FA profile of milk made by cows living under oceanic climatic conditions. Feeding strategies based on grazing could lead to substantial increases in MUFA in milk. Six hours of grazing leads to no reduction in the milk yield, and saves 3·5 kg of TMR/d per cow. Twelve hours grazing reduces the DMI associated with TMR without affecting milk yield, but provides milk with higher proportions of CLA and C18:3. Improving the FA profile of milk by grazing should be considered in strategies designed to produce healthier milk.
This work was supported by Spanish projects PCTI-PC06-006 and INIA-RTA2007-0058-C02. Mr. Morales-Almaráz was financed by CONACYT-México.




