


INTRODUCTION
Studies of the early ontogenic stages in benthic marine invertebrates are especially helpful for understanding the type of population control acting upon a species in a given area (Roughgarden et al., Reference Roughgarden, Gaines and Possingham1988). This is due to the fact that the larvae and early juvenile stages are the most important for dispersal and they have the highest level of mortality at the population level (Palmer et al., Reference Palmer, Allan and Butman1996; Gosselin & Qian, Reference Gosselin and Qian1997; Hunt & Scheilbling, Reference Hunt and Scheilbling1997). Additionally, in the case of species that can be exploited commercially, these studies allow us to understand the fluctuations in stock and recruitment (Wahle, Reference Wahle2003), and thus form a biological baseline useful in the sustainable management of fisheries (e.g. Eaton et al., Reference Eaton, Brown, Addison, Milligan and Fernand2003).
However, an important limitation for many field based ecological studies is the inability to correctly identify the early stages of many species. Larval phases such as the zoea, decapoditae and megalopae of decapod crustaceans frequently do not exhibit the specific diagnostic characteristics as evident in adults, but have cryptic morphologies within the related group. As a result individuals are assessed and assigned to higher taxa, genera or even families. However, this generates ambiguities and less precision in the interpretation of ecological data (e.g. recruitment events).
A classic tool for helping to identify larvae collected in the field is to use complete descriptions of larvae obtained from laboratory cultures. These descriptions have proved useful not only in ecological studies but also for systematic and evolutionary studies (Williamson, Reference Williamson and Abele1982; McHugh & Rouse, Reference McHugh and Rouse1998; Feldmann, Reference Feldmann2003). Despite the utility of this approach for groups such as euphausids and decapods, the larval morphology may vary depending on environmental conditions (Anger, Reference Anger and Vonk2001). Thus, the controlled conditions (temperature, salinity, density and absence of predators) of laboratory cultivation may reduce the morphological plasticity of the larvae compared with those that develop in the field. Indeed important morphological differences have been observed between brachyuran larvae raised in the laboratory and those encountered in the field. For example, Cuesta et al. (Reference Cuesta, Luppi, Rodriguez and Spivak2002) describe field collected megalopae of the graspid Neohelice granulata with morphological anomalies, in the form of additional cephalothoracic spines and a reduced number of setae compared to megalopae raised in the laboratory. Another species, Pilumnoides perlatus, differs in the pattern of setation and also demonstrates a noticeable difference in size; megalopae larvae from the laboratory are 30% smaller than those encountered in the field (Ampuero, Reference Ampuero2007).
When larval descriptions are not available, the stage being studied can be cultivated up to the identifiable stage (normally early juveniles) to generate a voucher collection which permits the rapid identification of the remaining individuals. This technique has been successfully used in a number of studies concerning recruitment dynamics of decapods (Palma et al., Reference Palma, Pardo, Veas, Cartes, Silva, Manríquez, Díaz, Muñoz and Ojeda2006; Pardo et al., Reference Pardo, Palma, Prieto, Sepúlveda, Valdivia and Ojeda2007; Negreiros-Fransozo et al., Reference Negreiros-Fransozo, Wenner, Knott and Fransozo2007). However, this form of larval identification is time consuming and carries with it a decided degree of uncertainty when the genetic diversity of the sampling area is high and/or the species are only differentiated by morphological details of appendages difficult to see without adequate dissection (i.e. mandible and maxilla).
The use of molecular markers has been demonstrated to be a powerful tool for the identification of cryptic species or ontogenetic stages exhibiting little differentiation (Hebert et al., Reference Hebert, Cywindka, Ball and deWaard2003a; Vences et al., Reference Vences, Thomas, Bonett and Vieites2005; Rao et al., Reference Rao, Liston, Crampton and Takeyasu2006). This applies especially to the sequencing of the COI gene of mitochondrial DNA as it has proved to be a useful character for distinguishing between malacostran crustaceans (Knowlton & Weigt, Reference Knowlton and Weigt1998; Barber & Boyce, Reference Barber and Boyce2006), but is not clear in other zoological groups (e.g. France & Hoover, Reference France and Hoover2002; Shearer et al., Reference Shearer, Van Oppen, Romano and Worheide2002; Hebert et al., Reference Hebert, Ratnasingham and deWaard2003b; Meier et al., Reference Meier, Shiyang, Vaidya and Ng2006). Thus the molecular markers can be a good alternative for the identification of cryptic larval stages of decapods, especially when various species of the same genera are collected simultaneously.
The study of the megalopae of the genus Cancer is complicated by the problems outlined above, due to a high degree of morphological similarity between the species (Iwata & Konishi, Reference Iwata and Konishi1981). Four species (Cancer edwardsii Bell, 1835, C. Coronatus Molina, 1782, C. setosus Molina, 1782 and C. porteri Rathbun, 1930) have overlapping geographical distributions along the east coast of the South Pacific (Nations, Reference Nations1975) so their megalopa larvae can be encountered sympatrically. The first three have a latitudinal distribution which extends from the equatorial region down to the Patagonian region (Garth, Reference Garth1957) and C. porteri has a discontinuous distribution in both hemispheres, except in the tropical region (Nations, Reference Nations1975), with a southern limit at approximately 35°S (Garth, Reference Garth1957). Despite their high abundances and high levels of commercial exploitation in the region (SERNAPESCA, 2006), little is known about their population ecology and even less about their recruitment dynamics (but see Jesse & Stotz, Reference Jesse and Stotz2003; Pardo et al., Reference Pardo, Palma, Prieto, Sepúlveda, Valdivia and Ojeda2007).
One of the major difficulties in advancing research into this group of crabs is the identification of the megalopa larvae, a key stage during which recruitment and the moult to the first juvenile stage takes place. There are two descriptions available of megalopae for the genus Cancer along the coast of the south-eastern Pacific, C. edwardsii and C. setosus (Quintana, Reference Quintana1983; Quintana & Saelzer, Reference Quintana and Saelzer1986), both are from laboratory cultures. These larvae are extremely similar and the comparison of the general morphology presented in the published figures does not allow for adequate identification of individuals taken from the field. Additionally, all four species can occur in the same habitat (Jesse & Stotz, Reference Jesse and Stotz2003; Muñoz et al., Reference Muñoz, Pardo, Henríquez and Palma2006) and three of the species (C. edwardsii, C. setosus and C. coronatus) have been recorded with high juvenile abundances in the estuaries of the north Patagonian region (Pardo L.M., unpublished data). This increases the possibility that megalopae of all three species may be present simultaneously in the same location. Without a clear description of these larvae taken from the natural environment, it is difficult to advance our understanding of the specific population ecology of these species throughout their geographical range.
Thus the objectives of this research are to: (1) describe the megalopa larvae of the three species found in the field; (2) determine the specific morphological characteristics (SMC) which permit rapid identification; (3) determine which of the SMC do not vary either spatially or temporally; and (4) corroborate, using molecular markers (the COI gene), the identification of specific megalopae which were identified by means of the SMC, contrasting them with adults of all the species of Cancer present in the region.
Taxonomic status of studied species
According to Ng et al. (Reference Ng, Guinot and Davie2008), some Pacific species of Cancer have been reassigned to other genera: Cancer edwardsii as Metacarcinus edwardsii and Cancer setosus as Romaleon polyodon. The criteria used by Ng et al. (Reference Ng, Guinot and Davie2008) for this reassignment of both Metacarcinus and Romaleon were fully based on the study carried out by Schweitzer & Feldmann (Reference Schweitzer and Feldmann2000). This is a palaeogeographical work, based only on the carapace morphology without including other morphological or genetic traits. At present molecular evidence analysed by one of our collaborators in a phylogenetic study (Mantelatto F.L.M., unpublished data) would not support new nominations at this time and pending a further set of species. In this same context, Harrison & Crespi (Reference Harrison and Crespi1999) showed also some controversy among morphological and genetic Cancer species status. Therefore, with this non-consensual idea and possible controversy, we decided to keep the previous taxonomy and nomenclature of this group. Additionally, we did not use Cancer plebejus (Poepping, 1836) and C. polyodon (Poepping, 1836) because they are considered as junior synonyms of C. coronatus Molina, 1782 and C. setosus Molina, 1782 respectively. In this sense and considering that species studied here are commercially important we think that this is the best way, at this time, to keep this name and avoid problems as recently reported for the shrimp species of the genus ‘Penaeus’ (Flegel, Reference Flegel2008).
MATERIALS AND METHODS
Collection of the megalopae
Larvae were collected in the subtidal of the San Carlos inlet in Bahía Corral, located on the south side of the mouth of the Rio Valdivia, Chile (39º 49′S 73º 14′W). In this area, the habitat consists of a mixture of boulders and coarse sand. Samples were obtained by means of an airlift manipulated by divers at depths of between 8 and 10 m. Because crab megalopae were found during spring and early autumn only, we analysed the morphological characteristics of larvae collected during October, December and April, in order to obtain a temporally representative analysis. In addition we analysed megalopae of C. setosus obtained from Punta Tralca located in central Chile (33° 35′S 71° 42′W), some 800 km north of the principal sampling sites.
Description of the megalopae
Between 10 and 20 specimens of each species from Bahía Corral were dissected and examined in detail to describe larvae morphology. Additionally, 10 specimens of C. setosus from central Chile (Pta de Tralca) were also analysed. The megalopae and their appendages were dissected in glycerin on microscope slides under a stereo microscope. The figures were made using a drawing tube mounted on a Zeiss AXIOSKOP microscope. The descriptions were made following the scheme proposed by Clark et al. (Reference Clark, De Calazans and Pohle1998). Typically the measurement of the cephalothorax length (CL) in species of Cancer is made from the distal end of the rostral spine to the mid-posterior margin of the carapace (DeBrosse et al., Reference DeBrosse, Sulkin and Jamieson1990). However, due to the damage found in the rostral spine on several individuals, in this study the crabs were measured from the base of the rostral spine to the mid-posterior margin of the cephalothorax. Cephalothorax width (CW) was measured on the midline of carapace.
DNA sequence based identification
After dissection of the megalopae, tissue samples from four individuals per species from Bahía Corral were preserved in 95% ethanol for subsequent genetic analyses. Two additional megalopae were also sampled from Punta de Tralca, only for C. setosus. As there is no baseline genetic information for the species of the genus Cancer from Chile, we obtained adult specimens from all Cancer crabs which inhabit the Chilean coast to corroborate the genetic identification of the larvae. Specifically adult specimens were collected from Bahía Corral (C. coronatus, N = 1; C. edwardsii, N = 2 and C. setosus, N = 1), Caleta Lenga, Concepción (36º 44′ S 73º 11′W) (C. coronatus, N = 1 and C. porteri, N = 2) and Punta de Tralca (C. setosus, N = 1). Additionally, one adult specimen of the Platyxhantid crab Homalaspis plana (Milne-Edwards, 1834) from Bahia Corral was also analysed. From each adult specimen a sample of muscle tissue from the chelipod was extracted, and was preserved in 95% ethanol for subsequent DNA extraction.
The DNA extraction, from both adults and megalopae, was conducted using the method described by Aljanabi & Martinez (Reference Aljanabi and Martinez1997). The mitochondrial COI gene was amplified using the protocol and primers described by Folmer et al. (Reference Folmer, Black, Hoek, Lutz and Vrijenhoek1994) and the PCR (polymerase chain reaction) used the following conditions: 1 × buffer (Invitrogen), 3.2 nM MgCl2, 0.2 U/μL dNTP, 5 pmol forward and reverse primers and 0.1 U U/μL of Taq polymerase. The PCR cycle consisted of 3 minutes at 94ºC, 30 cycles of 30 seconds at 94ºC, 90 seconds at 42ºC and 90 seconds at 72ºC with a final elongation of 7 minutes at 72ºC. The PCR product was cleaned using QIAQuick columns (QIAGen, Mississaga, Ontario, Canada) and sequencing was performed at Macrogen Inc (www.macrogen.com). Sequences were then aligned by eye using the ProSeq v.2.9 software (Filatov, Reference Filatov2002). All haplotype was deposited in Genbank (Accession Numbers: FJ155371 to FJ155382).
In order to determine the nucleotide relationship among samples (megalopae and adults crabs), a neighbour-joining based phylogenetic (NJ) analysis was performed using Mega 4.0 software (Tamura et al., Reference Tamura, Dudley, Nei and Kumar2007). Using a bootstrap of 10,000 replicates, the analysis tested the consistency of each branch in the tree, grouping sequences with similar nucleotide composition. Using this method, unidentified megalopae larvae could be grouped with conspecific adults. Finally, to root the tree, sequences of Homalaspis plana were used as outgroup (Genbank Accession Number: FJ155383).
RESULTS
To avoid long and repetitive description, the megalopa of Cancer coronatus is described completely whereas only differences are described in detail for the rest of the species.
Cancer coronatus Molina, 1782
Cephalothorax (Figure 1A): length to width ratio of 1.4, narrowing towards the anterior margin terminating in a ventrally directed rostral spine. The posterior margin is smooth with a medial spine. The surface of the carapace is almost devoid of setae.

Fig. 1. Comparative illustration of megalopae. General dorsal view of (A) Cancer coronatus, (B) Cancer edwardsii and (C) Cancer setosus.
Antennule (Figure 2A): penduncle 3-segmented, with 2, 3, 0 simple setae plus 2 plumose setae in the proximal segment. Unsegmented endopod with 2 subterminal and 2 terminal simple setae. Exopod 4-segmented with 0, 6, 10, 3 and 0, 0, 3, 1 simple setae; and 1 long terminal plumose seta on the distal segment.

Fig. 2. Comparative illustration of the cancridae megalopae appendages. Antennule, antenna, mandible and maxillule of Cancer coronatus (A, D, G, J); Cancer edwardsii (B, E, H, K); Cancer setosus (C, F, I, L).
Antenna (Figure 2D): peduncle 3-segmented; setation 4, 2, 4. Flagellum composed of 8 segments, with a setation pattern of 0, 0, 4, 0, 5, 0, 4, 3 simple setae, proceeding from the proximal to distal segment.
Mandible (Figure 2G): a developed cutting plate. Three-segmented mandibular palp with 7 plumodenticulate cuspidate and 3 plumose setae on the distal segment.
Maxillule (Figure 2J): coxal endite with 11 simple, 13 plumodenticulate cuspidate and 1 plumose seta. Basial endite with 8 plumodenticulate cuspidate and 6 marginal simple setae. Endopod 2-segmented with 2 simple setae on the proximal segment and 2 on the distal segment. Exopodal setae present.
Maxilla (Figure 3A): coxal endite bilobed. Proximal and distal lobe with 4 + 0 plumodenticulate cuspidate, 2 + 1 plumose setae and 3 + 0 simple setae. Basial endite bilobed, the proximal and distal lobe with 7 + 8 plumodenticulate cuspidate and 1 + 1 simple setae. Endopod with 5 plumose setae along the external margin. Scaphognathite with 86 plumose and 2 simple marginal setae, plus 6 simple setae on the internal surface.

Fig. 3. Comparative illustration of the cancridae megalopae appendages. Maxille, first maxilliped and second maxilliped of Cancer coronatus (A, D, G); Cancer edwardsii (B, E, H); Cancer setosus (C, F, I).
First maxilliped (Figure 3D): triangular epipod at the proximal end narrowing rapidly towards the distal end, with 10 evenly distributed long simple, and 5 shorter setae close to the proximal end. Coxal and basial endite with 13 + 22 simple setae. Unsegmented endopod with 4 terminal simple setae. Exopod 2-segmented with 4, 4 plumose setae.
Second maxilliped (Figure 3G): epipod elongated with 13 long filiform setae. Gill present between the epipod and exopod. Endopod 4-segmented, where the proximal segment has an obvious suture line. Setation pattern of 3, 1, 7 8 simple setae proceeding from the proximal to distal segment. Exopod 2-segmented with 1 simple seta on the proximal segment and 5 terminal plumose setae on the distal segment. Protopodite with 4 simple setae close to the base of the endopod.
Third maxilliped (Figure 4A): elongated epipod with 24 evenly spread long simple setae and 2 plumose setae located at the proximal end. Protopod with 13 simple and 4 plumose setae. Five-segmented endopod with 23 + 9 simple setae in the isquium and merus; and carpus, propodus and dactylus with 14 + 13 + 9 plumose setae. Exopod 2-segmented with 4 simple setae on the proximal segment and 5 terminal plumose setae on the distal segment.

Fig. 4. Comparative illustration of the cancridae megalopae appendages. Third maxilleped, cheliped and second pereiopod of Cancer coronatus (A, D, G); Cancer edwardsii (B, E, H); Cancer setosus (C, F, I).
Chelipeds (Figure 4D): surface covered in a large number of simple setae. Exhibits a prominant spine on the internal margin of the isquium. Merus with an acute projection at the distal end. Propodus subquadrate in form with a length to width ratio of 2.0, logitudinal grooves ill defined, internal chela fixed with shallow denticulation.
Second pereiopod (Figure 4G): surface covered in setae, the majority of which are simple in form. Coxa of second pereiopod with a thick cuticular spine. Fifth pereiopod (Figure 1A) with 3 long terminal setae on the dactylus.
Abdomen (Figure 1A): composed of 6 somites plus the telson, and dorsal surface with 2, 4, 6, 8, 8, 3 simple setae. Second and fifth segments possess biramous pleopods, fifth pleopod (Figure 5A) exopods have 21 terminal long plumose setae, endopods with 4 terminal cincinnuli. Sixth segment with a pair of uniramous uropods with 1 plumose seta on the external margin of the protopodite and 14 long plumose setae on the exopodite (Figure 5D).
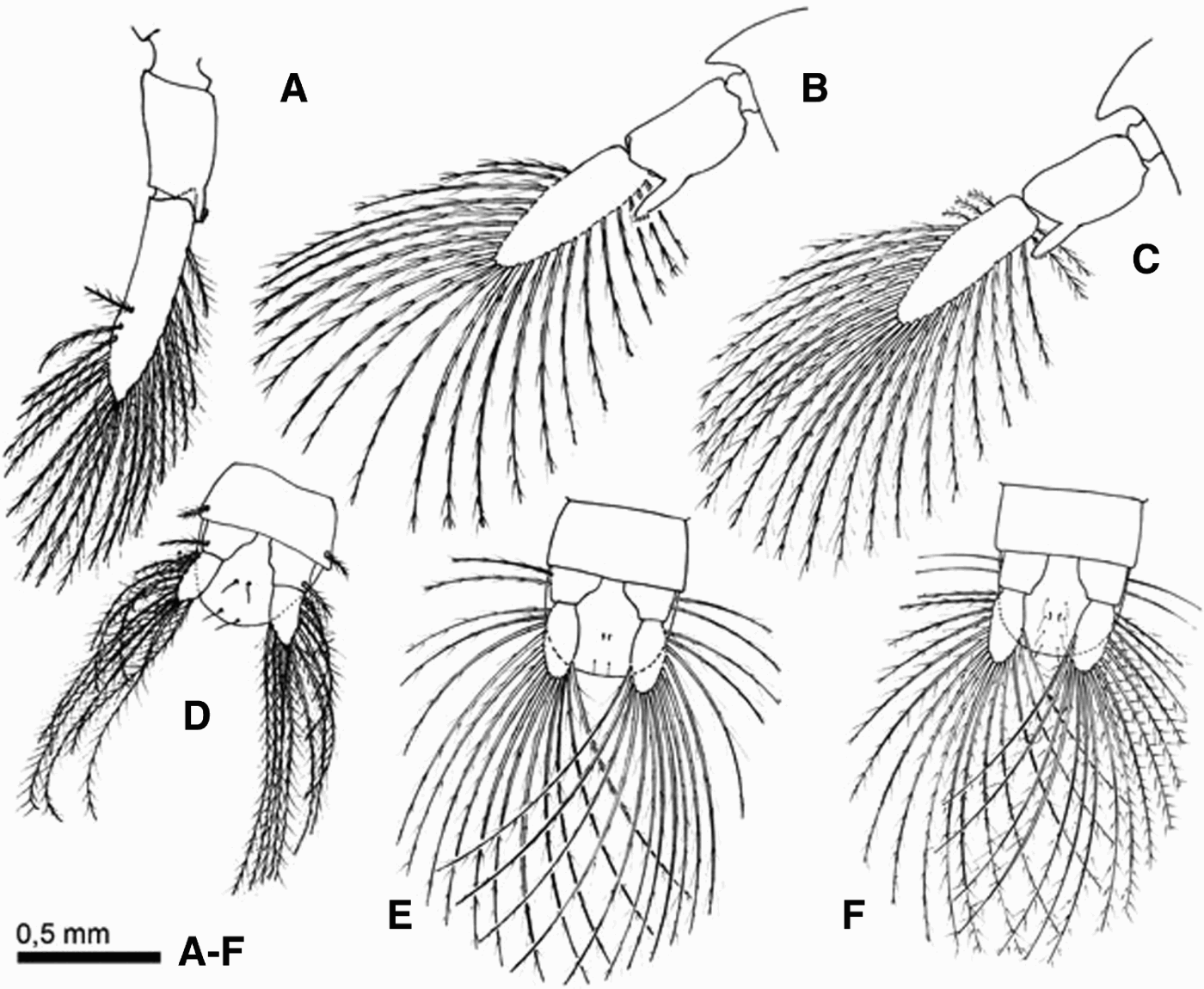
Fig. 5. Comparative illustration of the cancridae megalopae appendages. Second pleopod and telson of Cancer coronatus (A, D); Cancer edwardsii (B, E); Cancer setosus (C, F).
Telson (Figure 5D): semicircular posterior margin, with 2 pairs of simple setae on the ventral surface and a single pair on the dorsal surface.
Cancer edwardsii Bell, 1835
Cephalothorax (Figure 1B): the ratio length/width was approximately 1.3. Sixty-two surface setae and 58 ventral marginal setae.
Antennule (Figure 2B): penduncle with 4, 9, 2 simple setae plus 8 plumose setae. Endopod has 5 simple setae. Exopod with a setation of 0, 14, 10, 6 aesthetascs and 0, 0, 3, 1 simple setae on each segment.
Antenna (Figure 2E): peduncle setation 4, 2, 4 (the interior seta of the second segment is plumose); and flagellum with 0, 0, 4, 0, 5, 0, 4, 3 simple setae per segment.
Mandible (Figure 2H): mandibular palp with 13 plumodenticulate cuspidate setae on the distal segment.
Maxillule (Figure 2K): coxal and basial endite with 12 (7 plumodenticulate cuspidate, 5 plumose) +19 (plumodenticulate cuspidate) setae and 5 + 4 simple marginal setae. Epipodal and exopodal setae present.
Maxilla (Figure3B): proximal and distal lobe of the coxal endite with 7 + 6 plumose setae. Proximal and distal lobe of the basial endite has 8 + 10 plumodenticulate cuspidate and 1 + 1 simple setae. Scaphognathite with 77 plumose setae.
First maxilliped (Figure 3E): epipod with 12 very long simple setae along the mid to distal exterior margin, and 2 shorter simple setae on the proximal end. Coxal endite with 14 plumose setae and the basial endite with 25 plumodenticulate cuspidate setae. Endopod with additional plumose setae. Exopod with 4, 5 plumose setae and 1, 1 simple seta on the proximal and distal segment respectively.
Second maxilliped (Figure 3H): epipodite with 15 long simple setae. Endopod with 5, 2 (1 simple, 1 plumose), 7 (2 plumodenticulate cuspidate, 5 plumose), 4 (5 plumodenticulate cuspidate, 4 plumose) setae. Exopod with 2 simple setae on proximal segment and 5 long plumose setae on the distal segment.
Third maxilliped (Figure 4B): epipod with 20 long simple setae, and 6 plumose setae located on the proximal margin. Protopod with 25 plumose setae. Endopod, 27 (plus 6 plumose), 7, 6, 12, 9 plumodenticulate cuspidate setae and 1, 4, 8, 0, 0 simple setae. Exopod with 5 simple setae on the proximal segment, and 8 long plumose setae on the distal segment.
Chelipeds (Figure 4E): propodus globular in form with a length to width ratio of 1.5 and well defined longitudinal grooves.
Abdomen (Figure 1B): somites dorsal surface with 4, 16, 14, 14, 14, 6 simple setae. Biramous pleopods, fifth with exopods having 24 terminal long plumose setae (Figure 5B). Uropod with 2 long plumose setae on the protopodite (Figure 5E).
Telson (Figure 5E): 2, 2 simple setae on the ventral and dorsal surface respectively.
Cancer setosus Molina, 1782
Cephalothorax (Figure 1C): length to width ratio of 1.4. 32 surface setae and 16 ventral marginal setae.
Antennule (Figure 2C): penduncle with 7 plumose setae and 1 simple seta on the first segment: and 4 simple setae on the second one. Endopod with 6 simple setae. Exopod with 0, 18, 12, 10 aesthetascs.
Antenna (Figure 2F): peduncle with 4, 1, 5 setae (the seta on the second segment is plumose) and flagellum with 0, 0, 4, 1, 5, 2, 3, 2 simple setae.
Mandible (Figure 2I): mandibular palp proximal segment with 1 plumose seta; distal segment with 15 plumodenticulate cuspidate setae.
Maxillule: (Figure 2L): coxal endite with 2 plumose, 7 plumodenticulate cuspidate and 7 simple setae. Basial endite with 24 plumodenticulate cuspidate and 5 simple setae.
Maxilla (Figure 3C): coxal endite proximal and distal lobe with 3 + 5 plumodenticulate cuspidate, 3 + 0 plumose and 1 + 1 simple seta. Basial endite proximal and distal lobe with 10 + 12 plumodenticulate cuspidate and 1 + 1 simple seta. Endopod with 7 plumose setae. Scaphognathite with 95 marginal plumose setae.
First maxilliped (Figure 3F): endopod with 22 long simple setae. Coxal and basial endite with 21 + 36 plumodenticulate cuspidate plumose setae. Endopod with 3 plumose setae and 5 simple setae. Exopod with 4, 7 plumose setae.
Second maxilliped (Figure 3I): epipodite with 18 long simple setae. Endopod with 2 (plus 2 plumose), 1, 8, 9 plumodenticulate cuspidate, and 6, 1, 0, 0 simple setae. Exopod with 1, 7 plumose setae plus 2 simple setae. Protopod with 4 simple setae and 4 plumose setae close to the base of the endopod.
Third maxilliped (Figure 4C): epipodite with 30 long simple setae and 6 marginal plumose setae located near the proximal end. Protopod with 18 plumose setae and 2 simple setae. Endopod with 37, 16, 20, 18, 9 plumodenticulate cuspidate setae. Exopod with 3 simple setae on the proximal segment and 10 plumose setae on the distal segment.
Chelipeds (Figure 4F): without isquial spine. Tubular propodus with a length to width ratio of 1.8 and several well defined longitudinal grooves.
Abdomen (Figure 1C): somites dorsal surface with 4, 18, 18, 18, 14, 6 simple setae. Biramous pleopods, fifth (Figure 5C) with exopods having 24 long and 3 short terminal plumose setae. Uropod with 2 long plumose setae on the protopod and 16 long plumose setae on the exopod (Figure 5F).
Telson (Figure 5F): 2, 8 simple setae on the ventral and dorsal surface respectively.
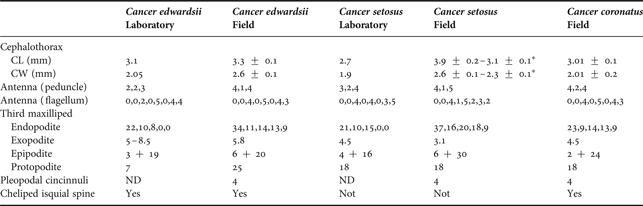
Intraregional variation: larvae collected around 800 km between them did not show differences in SMC, but southern were noticeably larger than northern megalopae (Table 1).
Table 1. Variation in biometric measures of Cancer megalopae from field and laboratory. All setal counts are listed from proximal to distal. CL is measured from base of rostral spine to middle of the posterior margin. From megalopae described in the literature (reared at the laboratory), CL and CW were estimated from illustrations, using the graphic scales provided by the authors (Quintana Reference Quintana1983; Quintana & Saelzer Reference Quintana and Saelzer1986). Abbreviations: CL, cephalothorax length; CW, cephalothorax width; ND, no data. * indicate megalopae from Punta de Tralca.
Molecular identification
The sequences of 554 bp obtained from the COI gene of both megalopae and adults of the studied species of Cancer, exhibited a high level of differences between species. We observed around 60 base pair differences between species and a maximum of 4 between individuals of the same species (Table 2). These clear differences between the sequences of each species were the basis for grouping the megalopae with the adults, without ambiguity, demonstrated by tree nodes with high support values between different species (Figure 6).

Fig. 6. Neighbour-joining tree analysis of the COI sequences for the Cancer species. The number at the tree nodes indicates the bootstrap values from 10,000 replicates. The Figure shows also the sample site and stage that illustrated these haplotypes. Me, megalopae; Ad, adult—most part of individuals from Bahía Corral, except (*) from Punta de Tralca and (**) from Concepción.
Table 2. Summary of base pair difference between COI sequences (554 bp) showing maximum and minimum difference within and among species. The mean of differences is expressed in per cent.

DISCUSSION
Several characters have been used to identify megalopae of the genus Cancer to the species level. Specific diagnostic characteristics that have been considered previously include: the number of plumose setae present on the uropods and cincinnuli present on the pleopods (Poole, Reference Poole1966; Trask, Reference Trask1970), the carapace length and presence or absence of a lateral knob (Orensanz & Gallucci, Reference Orensanz and Gallucci1988), and the setae and spines over the antenna and the endopod of the third maxilliped (Iwata & Konishi, Reference Iwata and Konishi1981). However, we found a considerable amount of overlapping in these characteristics between species (Table 1).
The general morphology of the megalopae studied display a high degree of intra-specific similarity. However, certain morphological features in the chelipeds such as the presence or absence of the isquial spine allow for a rapid and precise identification of each of the three species. These diagnostic characteristics have not been commented upon previously in the descriptions of C. edwardsii and C. setosus (Quintana, Reference Quintana1983; Quintana & Saelzer, Reference Quintana and Saelzer1986), based on laboratory reared larvae, and have been recorded for C. coronatus for the first time. In addition, these features appear to be highly conserved on both the spatial and temporal scales. For example, megalopae of C. setosus collected at locations separated by more than 800 km were identified to the species level and were later corroborated by the DNA analyses, despite differences in their size and date of collection.
The scarce morphological differentiation between the megalopae of the genus Cancer has been noted previously. DeBrosse et al. (Reference DeBrosse, Sulkin and Jamieson1990) did not find invariable morphological features which allowed for the identification of three sympatric species of Cancer (C. magister, C. oregonesis and C. productus) collected in the Puget Sound basin on the north-east Pacific coast. As in our case, the megalopae presented a high degree of morphological similarity (except for the considerably larger size of C. magister) and it was not possible to easily distinguish the larvae maintained in the laboratory using the published descriptions.
Another interesting finding was the differences in larval morphology between field and laboratory reared megalopae (Table 1). The larvae of C. edwardsii and C. setosus collected in the field are considerably larger than those raised in the laboratory. In the case of C. coronatus there were no available descriptions for comparison. In addition to the size, the larvae from the field exhibit a marked increase in the number of setae on their cephalothoracic appendages and in general all the megalopae analysed appear to have more setae than their conspecifics reared in the laboratory. Considerable variation has been described in the megalopae of other species of the genus Cancer obtained in the field (Orensanz & Gallucci, Reference Orensanz and Gallucci1988; DeBrosse et al., Reference DeBrosse, Sulkin and Jamieson1990) and also in other brachyuran species (Cuesta et al., Reference Cuesta, Luppi, Rodriguez and Spivak2002; Ampuero, Reference Ampuero2007).
The discrepancies between larvae collected in the field and those cultivated in the laboratory could be explained by two non-exclusive factors. First, the descriptions based on cultivated larvae normally utilize a single or few reproductive females to provide the individuals described, reducing the genotypic variation available for study. Second, the controlled conditions under which the larvae are cultivated (temperature, salinity and food availability) may restrict phenotypic expression.
The analysis of molecular markers in larvae collected from the field are few, but with a high degree of certainty in terms of identification. For example, Webb et al. (Reference Webb, Barnes, Clark and Bowden2006) conducted genetic analyses on Antarctic marine invertebrate larvae, but found that the high resolution of their analysis was limited by the general lack of genetic information for the adult organisms present in the area. In the present study, the lack of information on adults was not limiting for our analyses as they were collected and analysed at the same time as the megalopae. Thus, we were able to assign the megalopae of the genus Cancer on the coast of Chile to specific species using diagnostic morphological features which were later corroborated with the sequencing of the COI gene of mitochondrial DNA.
This study affirms the necessity of including morphological variation in the descriptions of larvae, and of analysing larvae collected in the field, along with those raised under different laboratory conditions. Furthermore, the present study provides clear evidence of the utility of molecular markers in the study of larvae where previous descriptions are unavailable or where there are significant morphological differences between the adults and the larvae of a species. These types of techniques are also useful for identifying the different stages of development of a species, and for those species that exhibit little or no diagnostic characters (Burton, Reference Burton1996; Neigel et al., Reference Neigel, Domingo and Stake2007).
ACKNOWLEDGEMENTS
We thank Dr Elena Clasing, Elena Gessler, Constanza Ceroni, Juan Pablo Fuentes, Matthew Lee and Simon Cardyn for their active participation in this research, from sampling to drawing the megalopae. Thanks to Marcela Espinoza for helping with DNA extraction. L.M.P. thanks the Direction de Investigation and Development (DID) of the Universidad Austral de Chile and FONDECYT 11070024 for financial support. D.V. thanks the Basal Grant PFB 023, CONICYT, Chile and Iniciativa Científica Milenio Grant ICM P05-002. Thanks to two anonymous referees who have greatly helped us in improving the clarity of the presentation. All experimental work was conducted in Chile and complied with its existing laws.








