


The clade Chiroptera is the second most diverse order of mammals and has a nearly global distribution (Brown et al. Reference Brown, Cashmore, Simmons and Butler2019; Frick et al. Reference Frick, Kingston and Flanders2019; Simmons & Cirranello Reference Simmons and Cirranello2019). According to Marchese (Reference Marchese2015), 80 %, of extant bats need conservation or research attention. The study of the fossil record provides unique information that is of great value in the conservation of the species under threat and the environmental ecosystems (Jablonski & Shubin Reference Jablonski and Shubin2015). Furthermore, bat conservation is important not only for the maintenance of biodiversity, but also for the role in pest control performed by these flying mammals, which thus provide important ‘ecosystem services’ for humans (Riccucci & Lanza Reference Riccucci and Lanza2014).
Time-calibrated molecular and genomic phylogenies and analyses of the morphological data locate the origin of bats at around the Cretaceous–Paleogene boundary, but it is not clear when they diversified in relation to the boundary. The first unequivocal bats appear at the early Eocene with the remains of Icaronycteris index Jepsen, Reference Jepsen1966 and Onychonycteris finneyi Simmons, Seymour, Habersetzer & G. F. Gunnell, Reference Simmons, Seymour, Habersetzer and Gunnell2008, both from the early Eocene Fossil Butte Member of the Green River Formation, Wyoming, USA (Simmons et al. Reference Simmons, Seymour, Habersetzer and Gunnell2008). There is thus a ten-million-year gap between the apparent genetic origin of the lineage and the first recognisable bat fossils (Gunnell & Simmons Reference Gunnell and Simmons2005). The scarce fossil record of this clade is made up of isolated teeth, indeterminate fragments and some articulated skeletons (Brown et al. Reference Brown, Cashmore, Simmons and Butler2019).
There are five volcanic archipelagos (the Azores, Madeira, the Savage Islands, the Canary Islands and Cape Verde) in the mid-Atlantic Ocean. Geographically, the Azores is furthest north and is composed of nine islands (Flores, Corvo, Terceira, Graciosa, São Jorge, Pico, Fayal, Santa María and São Miguel). Further S is Madeira, which includes Porto Santo and the Desertas, and the archipelago of the Savage Islands. Near to the African coast is the archipelago of the Canary Islands, with seven main islands (Fuerteventura, Lanzarote, Gran Canaria, Tenerife, La Gomera, La Palma and El Hierro) and several islets (La Graciosa, Alegranza, Isla de Lobos, Montaña Clara, Roque del Oeste and Roque del Este; Fig. 1). Finally, furthest S is the Cape Verde archipelago, with ten islands divided into two areas: the Barlovento Islands comprising Santo Antão, São Vicente, Santa Luzia, São Nicolau, Sal and Boa Vista; and the Sotavento Islands comprising Maio, Santiago, Fogo and Brava. Across this region there are up to 15 different species of bats. These include three endemic species: Plecotus teneriffae Barret-Hamilton, 1907 in the Canary Islands; Pipistrellus maderensis Dobson, 1878 in the Azores, Madeira and the Canary Islands; and Nyctalus azoreum Thomas, 1901 in the Azores; and two endemic subspecies, Barbastella barbastellus guanchae Trujillo, Ibañez & Juste (Reference Trujillo, Ibáñez and Juste2002) in the Canary Islands and Nyctalus leisleri verrucosus Bowdich, 1825 in Madeira (Fajardo & Benzal Reference Fajardo and Benzal2002; Lopes & Medeiros Reference Lopes and Medeiros2011; Trujillo & González Reference Trujillo and González2011; Vasconcelos Reference Vasconcelos2018; Borloti et al. Reference Borloti, Dinis and Vasconcelos2020; Table 1).
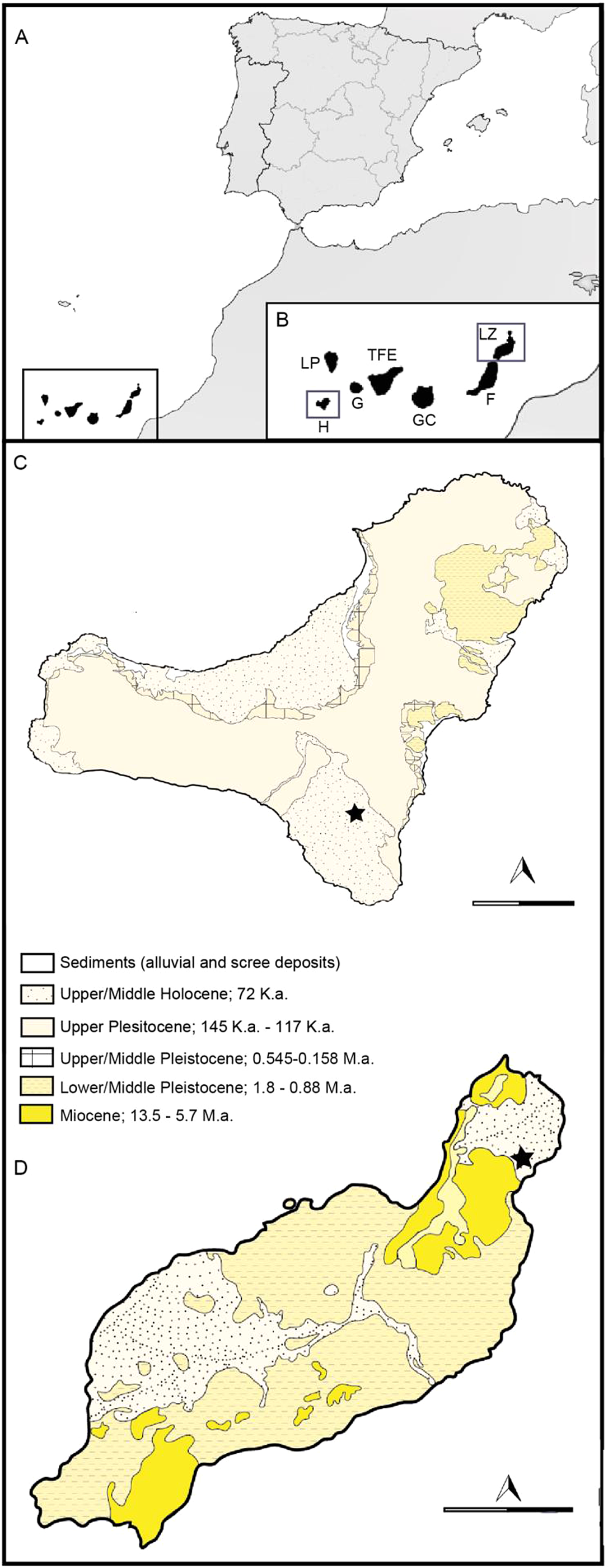
Figure 1 Map showing the location of the Canary Islands (A); the location of the islands of El Hierro and Lanzarote (B); geology and situation of the palaeontological sites in El Hierro (C) and Lanzarote (D). Star indicates location of the sites. Abbreviations: LP = La Palma; TFE = Tenerife; G = La Gomera; H = El Hierro; GC = Gran Canaria; F = Fuerteventura; LZ = Lanzarote.
Table 1 Compilation of all extant bats from the volcanic islands of the mid-Atlantic Ocean (*: currently extinct on this island; **: in press release).

In the Canary Islands, the fossil record is defined by the short geological history of the archipelago, its volcanic nature and the fragility of the remains. There are many current and fossil endemic species that have evolved in the fragile and changing island ecosystems.
Fossil bats are common components of Quaternary palaeontological cave sites (Galán et al. Reference Galán, Núñez-Lahuerta, López-García and Cuenca-Bescós2019a, Reference Galán, Núñez-Lahuerta, Sauqué, Cuenca-Bescós and López-García2019b). In the case of the Canary archipelago these include several lava tubes. The Canary Islands are the only archipelago in the mid-Atlantic Ocean where fossil bat remains are found, appearing in lava tubes and at archaeological sites. In concrete terms, they are found in four sites: Cueva de la Enladrillada (800 BP ± 50 years, Holocene) in Tenerife, Cueva Honda del Bejenado (700–750 ka, Pleistocene) in La Palma, Cueva de los Verdes in Lanzarote and Cueva Roja in El Hierro (Diego Cuscoy Reference Diego Cuscoy1972; Trujillo Reference Trujillo1991; Castillo et al. Reference Castillo, López, Martín and Rando1996; Govantes & García Reference Govantes and García2013). The distribution of this group is currently more extended, with seven indigenous species spread across all the islands (Trujillo Reference Trujillo1991; Asociación Española para la Conservación y el Estudio de los Murciélagos 2019; https://www.biodiversidadcanarias.es/biota/; Table 1). Of these seven species, six are from the Vespertilionidae family (Plecotus teneriffae; Hypsugo savii Bonaparte, 1837; Pipistrellus maderensis; Pipistrellus kuhlii Kuhl, 1817; Barbastella barbastellus Schreber, 1774; and Nyctalus leisleri Kuhl, 1817), and one from the Molossidae family (Tadarida teniotis Rafinesque, 1814). The distribution of these species among the Canary Islands is uneven, the western islands (Tenerife, La Gomera, La Palma and El Hierro) having greater diversity than the eastern islands (Gran Canaria, Graciosa, Lanzarote and Fuerteventura).
The main aim of the present work is to contribute to what is known of bat palaeobiodiversity in the volcanic oceanic archipelagos of the mid-Atlantic and to undertake the first taxonomic description of diagnostic bat fossil remains (teeth and humeri) from the mid-Atlantic volcanic islands, including the Canary Islands. To this end, remains from the islands of El Hierro and Lanzarote will be studied. All these data will provide the basis for the development of plans for the conservation of biodiversity in the future.
1. Geographical and geological context
The oldest sediments from the Canary archipelago are the Jurassic oceanic and clastic deposits of the island of Fuerteventura. These were produced by seafloor spreading events in the central Atlantic and the southwestern Moroccan continental margin (Steiner et al. Reference Steiner, Lausanne and Hernandez1998). The archipelago began to be formed during the Palaeocene (60 Ma) from a long-lived magma source, and its volcanic activity has continued through to the present in all the islands except La Gomera (Troll & Carracedo Reference Troll and Carracedo2016). The most recent historic eruptions (in the last 500 years) occurred onshore in Lanzarote, Tenerife and La Palma, and on the ocean floor off El Hierro (Troll & Carracedo Reference Troll and Carracedo2016).
Lanzarote and Fuerteventura are the easternmost islands in the archipelago. Between these islands there is a shallow stretch of sea called La Bocaina, which during the glacial periods of marine regression functioned as a land bridge. Lanzarote is a mature island dating from 16 to 22 Ma, mainly formed by two basalt volcanic edifices from the Neogene, each located at one end of the island: the Famara edifice (ca.15.5–12.3 Ma) in the N and the Los Ajaches edifice (ca.15.5–12.3 Ma) in the S (Lomoschitz et al. Reference Lomoschitz, Sánchez Marco, Huertas, Betancort, Isern, Sanz and Meco2016). Located to the E of Famara is an area of historic volcanism, where the Cueva de los Verdes is situated, a lava tube inside which the bat remains under study were collected.
At the other end of the archipelago is the island of El Hierro, which emerged 1.2 Ma. El Hierro saw the most recent volcanic activity in the Canary Islands, with the submarine eruption of the La Restinga (Pérez-Torrado et al. Reference Pérez-Torrado, Carracedo, Rodríguez-Gonzalez, Soler, Troll and Wiesmaier2012). The emergence of the island was a result of three overlapping volcanic systems (Tiñor, El Golfo and the Rifts; Pérez-Torrado et al. Reference Pérez-Torrado, Carracedo, Rodríguez-Gonzalez, Soler, Troll and Wiesmaier2012). Tiñor was the first to emerge, with an age of 1.12 to 0.88 Ma; it is located in the NE of the island. After the collapse of Tiñor, a new volcano, El Golfo, appeared, filling the collapsed area. Finally, the Rift System appeared; this does not form a central structure since the volcanic activity is simultaneous in the three arms of the rift (WNW, SSE and NE; Pérez-Torrado et al. Reference Perez-Torrado, Rodriguez-Gonzalez, Carracedo, Fernandez-Turiel, Guillou, Hansen and Rodríguez Badiola2011; Troll & Carracedo Reference Troll and Carracedo2016). In the southeastern sector of the Rift System, in an area with Holocene eruptions, is the lava tube known as Cueva Roja, where the bat remains under study were gathered.
A general and geological description of the two palaeontological sites where the studied remains were found follows below.
1.1. Cueva de los Verdes
The site of Cueva de los Verdes is situated on the island of Lanzarote. It is part of the lava tube from Corona Volcano known as the Corona lava tube. This is 7.5 km long and up to 35 m high and is considered one of the longest and largest lava tunnels in the world. The Cueva de los Verdes opens to the outside by a natural skylight (locally called a ‘jameo’), the product of a natural collapse of the lava tube roof (Troll & Carracedo Reference Troll and Carracedo2016).
The chemical composition of the lavas from Corona Volcano corresponds to an olivine basalt similar to the lavas ejected by the volcanoes that erupted during the prehistory of Lanzarote (Bravo Reference Bravo1964). A recent dating of the lava by Argon–argon (40Ar/39Ar) yielded an age of 21,000 ± 6500 years for the eruption of the volcano, near the last glacial maximum in the Upper Pleistocene (18,000–21,000 years ago; Troll & Carracedo Reference Troll and Carracedo2016).
1.2. Cueva Roja
The site of Cueva Roja is located on the island of El Hierro. It is a hole at a height of 350 m connecting with a 300-m-long tube in the hillside of El Julan. The tube is divided into two branches: one to the right with a 70 % slope at some points, which then continues level until its furthest point. The second, left-hand branch is narrow and with only a slight slope; it finishes in a volcanic cavern of about 12 m depth. In some sections the walls have collapsed (Hernández et al. Reference Hernández, Medina and Izquierdo1992). The geological materials of the Soleiman Volcano in San Andrés (El Hierro, Canary Island), which belong to the same geological series as Cueva Roja, have been carbon–14 dated to 4230 years BP (Fuster Reference Fuster1993), so we may assume that Cueva Roja is around 4000 years old (Holocene).
Institutional abbreviations. PCCRULL = Paleontology Collection – Carolina Castillo Ruiz; TFMC = Museo de Ciencias de Tenerife.
2. Material and methods
The studied fossils from Cueva Roja are kept at the Paleontology Collection – Carolina Castillo Ruiz, Universidad de La Laguna (Tenerife, Canary Islands, Spain), with the label PCCRULL; and the material from Cueva de los Verdes is kept at the Museo de Ciencias de Tenerife (Canary Islands, Spain), with the label TFMC.
The method of dental nomenclature is from Crespo et al. (Reference Crespo, Sevilla, Montoya and Ruiz-Sánchez2020) and the humerus nomenclature is from Salles et al. (Reference Salles, Arroyo-Cabrales, do Monte Lima, Lanzelotti, Perini, Velazco and Simmons2014). The measurement method is from Sevilla (Reference Sevilla1988) and Galán et al. (Reference Galán, Núñez-Lahuerta, López-García and Cuenca-Bescós2019a, Reference Galán, Núñez-Lahuerta, Sauqué, Cuenca-Bescós and López-Garcíab). The lower teeth are indicated as i1, i2, i3, c, p2, p3, p4, m1, m2 and m3, and the upper teeth as I1, I2, I3, C, P2, P3, P4, M1, M2 and M3.
The current material used for comparison with the bat remains is deposited in the Museo de Naturaleza y Arqueología from Tenerife.
3. Systematic palaeontology
Order Chiroptera Blumenbach, 1779
Suborder Yangochiroptera van den Bussche & Hoofer, 2004
Family Vespertilionidae (Gray, 1821)
Genus Plecotus (Geoffroy, 1818)
Plecotus teneriffae Barrett-Hamilton (1907)
(Figs 2, 3a)
Locality. Cueva Roja, El Hierro, Canary Islands, Spain.
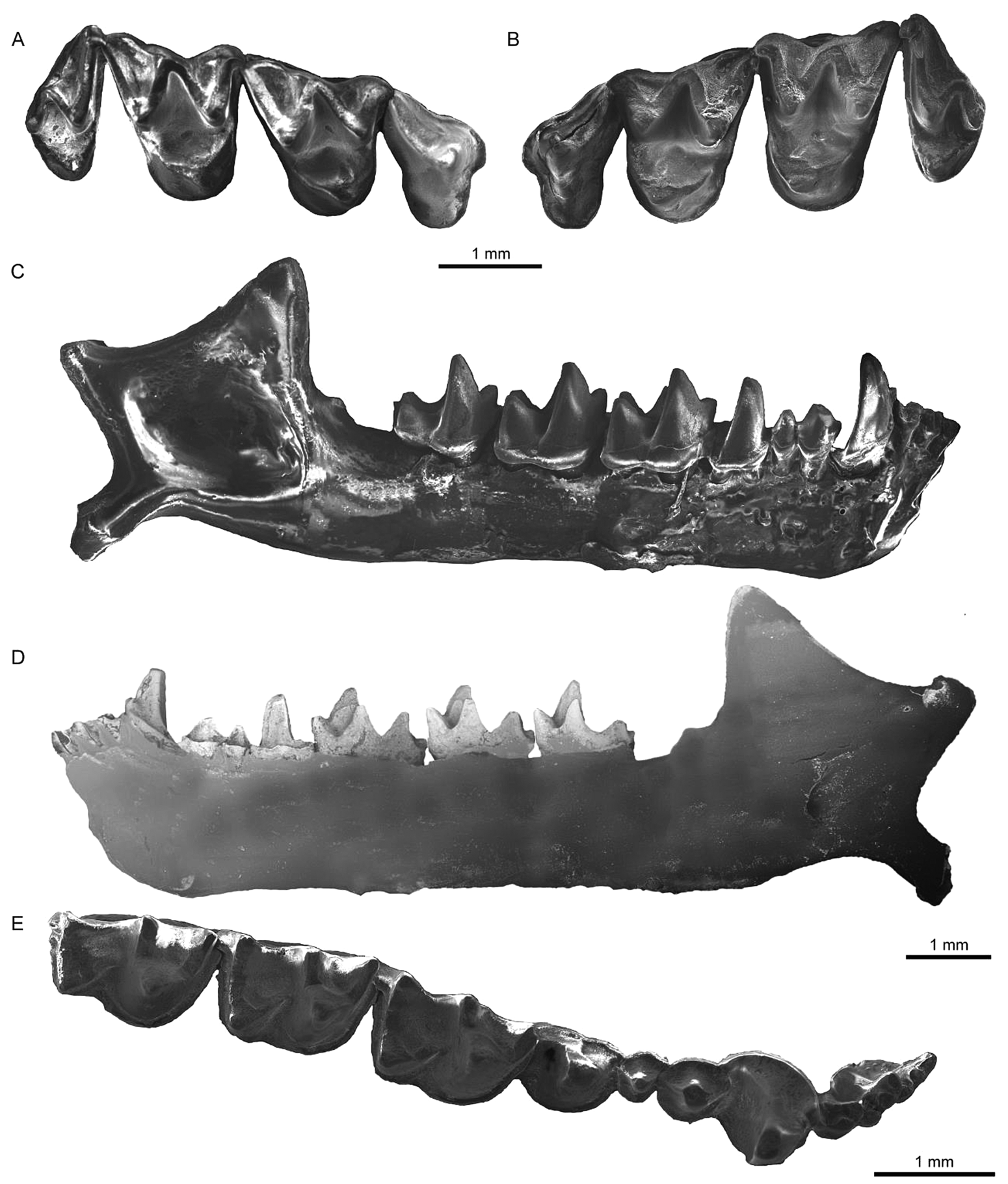
Figure 2 Fossil cranial of Plecotus teneriffae from Cueva Roja. (A) Left maxilla (PCCRULL1197-1) in occlusal view. (B) Right maxilla (PCCRULL1197-1) in occlusal view. (C) Right mandible (PCCRULL1197-2) in lateral view. (D) Right mandible (PCCRULL1197-2) in lingual view. (E) Right mandible (PCCRULL1197-2) in occlusal view.

Figure 3 Fossil humeri from Cueva Roja. (A) Right distal humerus of Plecotus teneriffae (PCCRULL1197-5) in anterior view. (B) Right distal humerus of Pipistrellus maderensis (PCCRULL1199-5) in anterior view.
Material. PCCRULL1197: maxilla with P4, M1, M2, M3 on both sides (PCCRULL-1197-1), two mandibles with all teeth (PCCRULL-1197-2 and PCCRULL-1197-3) and two humeri (PCCRULL-1197-4 and PCCRULL-1197-5).
Description. P4: P4 is a triangular tooth. There is a main cusp which is positioned towards the mesiolabial side. The anterior margin is very concave. The paracingulum is short and thick. The labial cingulum of the paraflexus is narrow, while the lingual cingulum is slightly thicker towards the anterior side than towards the posterior side. The talon is slightly developed.
M1: M1 is a sub-triangular tooth. The well-developed parastyle forms a right angle to the preparacrista. The precingulum is narrow. The metastyle is straight. The metacingulum is thin and does not connect with the metastyle. The metacone is slightly higher than the paracone. The ectoloph is asymmetrical, with a well-developed postmetacrista. The labial cingulum of the metaflexus is well developed. The paraloph, metaloph, paraconule, metaconule and talon are absent. The postprotocrista is not connected to the metacingulum. The lingual cingulum is slightly developed.
M2: M2 is similar to M1, with the difference in occlusal view; the paracingulum is thin but not thinner than the M1 and has a slightly more symmetrical ectoloph.
M3: M3 is a triangular tooth. The postparacrista and premetacrista are strongly reduced. The metacone is reduced. The metastyle and the postmetacrista are absent. The mesostyle is reduced to one-third of the paracone. The precingulum and the lingual cingulum are very reduced.
i1: in occlusal view, i1 is oval-shape, there are three cuspids in line. There is a small cingulid on the anterolabial side. There is a small cingulid on the posterior side.
i2: in occlusal view, i2 is oval-shape, there are four cuspids, three of which are in line. The posterior cusp is not well developed. There is a small anterior cingulid.
i3: in occlusal view, the tooth is subquadrangular. There are four cuspids of similar size, three of which are in line. The anterolabial cingulum is well developed.
c: the outline is circular and slightly mesiolingually elongated. The mesiolabial cuspid has four sides: while the lingual and the labial are convex, the anterior and the posterior are concave. It has a well-developed cingulid in the mesial side and there is a mesiolingual cuspule. There is a wide lingual cingulid. The labial cingulid is poorly developed.
p2: the outline is oval. There is a well-developed lingual and labial cingulid, with an mesiolingual thickening, which is slightly reduced on the anterior and posterior side. There is only a single cuspid.
p3: p3 is similar to p2, but smaller.
p4: p4 is a quadrangular tooth. The cuspid is in a mesiolingual position. There is a cingulid with a large expansion on the distolingual side and another smaller one on the mesiolingual side. In labial view, the cingulid is straight and very oblique, after Sevilla & López Martínez (Reference Sevilla and López Martínez1986).
Lower molars: the protoconid is the highest cuspid, being slightly higher in m3 regarding the other cuspids. The metaconid is higher than the paraconid; the paraconid in m3 is slightly smaller than in m1 and m2. The hypoconid is higher than the metaconid in m1 and m2, while in m3 it is as low as the entoconid. This latter cuspid is the lowest; in particular in m3 it is lower than in the anterior molars. In occlusal view, the paraconid, metaconid and entoconid in m1 and m2 are in line, whereas in m3, the entoconid is slightly displaced to the labial side.
m1: the molar is myotodont. The labial cingulum is thick and in labial view it is divided by a concave notch between trigonid and talonid. The trigonid is open. The paraconid notch is not present. The paralophid is angular in lateral view. The entocristid is concave. The oblique crista is connected to the middle of the posterior wall of the trigonid. The hypoconulid is in line with the entoconid.
m2: m2 is similar to m1, but the trigonid is closed.
m3: m3 is similar to m2; the talonid is smaller than in the other two molars and it is reduced by displacement of the hypoconid labially. The oblique crista is connected to the trigonid well below the middle of the trigonid.
Humerus: the spinous process of the humerus is short and blunt. The epitrochlea is slightly enlarged. The transition from the trochlea to the medial ridge of capitulum forms a slightly concave valley and the proximal edge is convex between the trochlea and the capitulum. The transition from the capitulum to the lateral ridge of capitulum forms a more concave groove than in the anterior area; this part is narrower than the anterior area. The lateral ridge of capitulum is visible in anterior view.
Remarks. The material from Cueva Roja studied here is similar to current populations. Plecotus teneriffae differ from the Plecotus austriacus and Plecotus gaisleri from Europe and Africa, although they are genetically closely related (Pestano et al. Reference Pestano, Brown, Suárez and Fajardo2003; Benda et al. Reference Benda, Kiefer, Hanák and Veith2004). The differences are as follows. (i) In their dental morphology, they are very similar in the upper teeth, although in the M3 of Pl. austriacus and Pl. gaisleri the metacone is much lower than the paracone (Sevilla Reference Sevilla1988; Benda et al. Reference Benda, Kiefer, Hanák and Veith2004), whereas in Pl. teneriffae they are practically at the same height. (ii) In the lower teeth, there are more differences between Pl. austriacus and Pl. teneriffae. In labial view, the cingulid of p4 in Pl. austriacus has two concavities, one in each root, whereas in Pl. teneriffae it may be curved or straight and oblique. The lack of the paraconid notch distinguishes Pl. teneriffae from Pl. austriacus (Sevilla Reference Sevilla1986, Reference Sevilla1988). The cingulid in labial view is thicker in Pl. teneriffae, whereas in Pl. austriacus it is irregular, being thicker on the trigonid side (Sevilla Reference Sevilla1988; Lindenau Reference Lindenau2005). Plecotus austriacus does not have the small notch in the labial cingulid of the lower molars. (iii) The specimens of Pl. teneriffae from El Hierro have a longer forearm than the specimens from Tenerife and La Palma (Palomo et al. Reference Palomo, Gisbert and Blanco2007).
Genus Pipistrellus (Kaup, 1829).
Pipistrellus maderensis (Dobson, 1878)
(Figs 3b, 4)
Locality. Cueva Roja, El Hierro, Canary Islands, Spain.

Figure 4 Fossil cranial of Pipistrellus maderensis from Cueva Roja. (A1) Left maxilla (PCCRULL1200-1). (A2) Right mandible (PCCRULL1200-3). (B1) Left maxilla (PCCRULL1201-1). (B2−4) Left mandible (PCCRULL1201-3). (A1, A2, B1, B4) in occlusal view; (B2) in labial view; (B3) in lingual view.
Material. PCCRULL-1198: maxilla with P2, P4 on both sides (PCCRULL1198-1), right mandible with i1, c, p2, p4, m3 (PCCRULL1198-2), left mandible with m3 (PCCRULL1198-3), two humeri (PCCRULL1198-4 and PCCRULL1198-5), 2C (PCCRULL1198-6 and PCCRULL1198-7), I1 (PCCRULL1198-8 and PCCRULL1198-20), M1 (PCCRULL1198-9), two M2 (PCCRULL1198-10 and PCCRULL1198-11), two M3 (PCCRULL1198-12 and PCCRULL1198-13), two m1 (PCCRULL1198-14 and PCCRULL1198-18), two m2 (PCCRULL1198-15 and PCCRULL1198-19), c (PCCRULL1198-16), p4 (PCCRULL1198-17); PCCRULL-1199: two humeri (PCCRULL1199-4 and PCCRULL1199-5), M3 (PCCRULL1199-6), m2 (PCCRULL1199-8), c (PCCRULL1199-9), m1 (PCCRULL1199-10), m3 (PCCRULL1199-11); PCCRULL-1200: left maxilla with I1, I2, C, P2, P4, M1, M2 (PCCRULL1200-1), right maxilla with I1, I2 (PCCRULL1200-2), right mandible (PCCRULL1200-3), left mandible with i2, i3, c, p2, p4, m1, m2, m3 (PCCRULL1200-4), two humeri (PCCRULL1200-5 and PCCRULL1200-6), M3 (PCCRULL1200-7), C (PCCRULL1200-8), P4 (PCCRULL1200-9), M2 (PCCRULL1200-10), M3 (PCCRULL1200-11); PCCRULL-1201: left maxilla with I1, P4, M1, M2, M3 and right maxilla with I2, P2, P4, M2 (PCCRULL-1201-1), mandible with i2, c, p2, p4 (PCCRULL-1201-2), mandible with all teeth (PCCRULL-1201-3), two humeri (PCCRULL-1201-4 and PCCRULL-1201-5), right C (PCCRULL-1201-6); PCCRULL-1202: right mandible with i2 to p4 (PCCRULL-1202-2), left mandible with p2 and p4 (PCCRULL-1202-3) and right humerus (PCCRULL-1202-4).
Description. I1: the tooth is elongated and oval in occlusal view. There is a main cusp on the anterior side, which has a distolabial cuspule. A small accessory distal cusp may be present (two out of five; PCCRULL1198-8 and PCCRULL1200-1) or not (three out of five). In one specimen, the right maxilla has an accessory cusp, whereas in the left maxilla this is absent. There is a thick cingulum that narrows lingually. There is one root.
I2: the tooth is circular in occlusal view. The main cusp is on the anterolingual side, which is connected by a crest to a cuspule on the labial side. There is another cuspule, on the lingual side that is more developed than the cuspule labial. There is a small cingulum on the labial side. There is one root.
C: subquadrangular tooth in occlusal view. On the posterolabial side there is a small talon. There is a groove on the labial side. The cingulum is thicker on the lingual side and thinner on the anterolabial side. There is one root.
P2: the tooth is small and circular in occlusal view. It is situated in a position lower than the talon of the canine. The cusp is on the anterior side. There is a crest that connects and starts with the cusp and then extends and ends towards the distal side. A cingulum surrounds the tooth, narrower on the distolingual side. There is one root.
P4: subquadrangular tooth. The labial margin is straight. The main cusp is on the mesiolabial side. The paracingulum is thick. The labial cingulum is narrow. There is an accessory cuspule on the anterolingual side. The mesiolabial side is slightly concave. The talon has an intermediate development with a very rounded base. There are three roots.
M1: trapezoid tooth. The parastyle is at a right angle to the preparacrista. The paracingulum is wide, but narrows near the parastyle, and they are poorly connected. The precingulum is short. The metastyle is slightly curved. The metacingulum is connected to the metastyle and is not variable in thickness. The ectoloph is slightly asymmetrical, with the metacone larger than the paracone. The labial cingulum of the metaflexus is well developed and the labial cingulum of the paraflexus is narrow. The paraloph may be well developed (two out of three; PCCRULL1200-1 and PCCRULL1201-1) or not (one out of three). The metaloph is well developed. The paraconule may be present (one out of three; PCCRULL1201-1) or not (two out of three). The metaconule is present. The postprotocrista is connected to the metaconule. The talon is absent. The lingual cingulum is narrow. There are three roots.
M2: this tooth is similar to M1 but is more rectangular. The ectoloph is slightly asymmetrical, the metacone being larger. The paraloph and the metaloph are very well developed. The metaconule is well developed. The paraconule may be present (three out of six; PCCRULL1198-11, PCCRULL1200-1 and PCCRULL1201-1) or not (three out of six). In one specimen, there is a paraconule in the left maxilla, but it is absent in the right one.
M3: triangular tooth. The parastyle is short but well developed, and at a right angle to the preparacrista. The paracingulum is short but thick and is connected to the parastyle. The postparacrista and the premetacrista are slightly reduced. The metacone is present and it protrudes slightly distally. The metastyle is absent. The postmetacrista is reduced to a small spur on the metacone. The mesostyle is reduced to half the height of the paracone. The paraloph is narrow. The precingulum and lingual cingulum are very short and reduced. There are three roots.
i1: in occlusal view, there are three cuspids in line. There is a small labial cingulum. There is one root.
i2: this tooth is similar to i1, but there is a small distal cingulum.
i3: this tooth is similar to i2.
c: the tooth is oval in occlusal view. The lingual and labial sides are convex while the anterior and posterior are concave. The main cuspid is on the mesial side and has a cingular cuspule on the mesiolingual side. The distolingual cingulid has a thickening. The precingulid is short and poorly developed. There is one root.
p2: the tooth is very reduced and is oval in occlusal view. There is a mesial main cuspid and two small accessory cuspules on the anterolingual and distolingual sides. The labial and the lingual cingulids are narrow. There is one root.
p4: the tooth is rectangular in occlusal view. There is a mesiolingual main cuspid. The precingulid is thick. There is a well-developed accessory anterolingual cuspule. The lingual cingulid has a thickening distally. This cingulid is straight and very oblique in labial view after Sevilla & López Martínez (Reference Sevilla and López Martínez1986). There are two roots.
Lower molars: the protoconid is the highest cuspid, and it decreases in size from m1 to m3. The hypoconid is the second largest cuspid on m1 and m2, whereas on m3 it is more reduced and is similar in height to the paraconid and the metaconid. The paraconid and metaconid are similar in height. The entoconid is slightly lower than the paraconid and metaconid. There are two roots.
m1: the tooth is nyctalodont. The labial cingulid is irregular in lateral view. This cingulid is thicker in the trigonid region than in the talonid region. The paralophid notch is not present. The trigonid is closed. The paralophid is angular. The entocristid is concave–convex. The oblique crista is connected to the middle of the posterior wall of the trigonid. The hypoconulid is in line with the entoconid.
m2: this tooth is similar to m1. The labial cingulid is thinner than in m1.
m3: m3 is similar to m2. The hypoconid in m3 is clearly more labial than in m2, thick and irregular after Sevilla (Reference Sevilla1988). It presents a labial reduction of the talonid with a more lingual entoconid.
Humerus: the triangular styloid process of the humerus is short and blunt and is moderately developed. The epitrochlea is slightly enlarged. The transition from the trochlea to the medial ridge of capitulum has a markedly concave groove in anterior and proximal view. The transition from the capitulum to the lateral ridge of capitulum is short and slightly concave. The lateral ridge of capitulum is hidden by the capitulum in anterior view. The radial fossa in anterior view is well developed.
Remarks. The dental morphology of current populations is similar to that of the fossil specimens. Pipistrellus kuhlii and Pipistrellus maderensis are genetically confirmed as sister species (Pestano et al. Reference Pestano, Brown, Suárez and Fajardo2003; Jesus et al. Reference Jesus, Teixeira, Freitas, Teixeira and Brehm2013). There are some differences in dental morphology: for example, in the upper molars, the paraloph, metaloph and metaconule are better developed in P. maderensis than in P. kuhlii. There is a small paraconule in P. maderensis whereas in P. kuhlii this is variable (see remarks on P. kuhlii below). The morphology of these latter cusps and crests is similar to that of Pipistrellus rouresi Crespo, Sevilla, Mansino, Montoya, & Ruiz-Sánchez, Reference Crespo, Sevilla, Mansino, Montoya and Ruiz-Sánchez2018 from the latest Miocene of Venta del Moro (Crespo et al. Reference Crespo, Sevilla, Mansino, Montoya and Ruiz-Sánchez2018), showing the primitive character of P. rouresi, although M1 is clearly narrower in the latter species. In p4, the labial cingulid is straight and very oblique in labial view in P. maderensis, whereas in P. kuhlii it is concave in labial view (Sevilla Reference Sevilla1986). In the lower molars, the talonid is larger in P. kuhlii (Sevilla Reference Sevilla1988) than in P. maderensis. Moreover, in P. kuhlii, the labial cingulid is thicker, showing the same thickness in the three lower molars, whereas in P. maderensis m1 presents a thicker on the trigonid than on the talonid. The trigonid of the m1 of P. maderensis is more open than in P. kuhlii (Sevilla Reference Sevilla1986).
The two species have a cranium with a similar total length (Trujillo Reference Trujillo1991). However, the specimens from Cueva Roja are slightly smaller than those found on the other islands. In the measurements of the forearm, the specimens from the Azores are slightly smaller than the material from the Canary Islands and Madeira (Trujillo & González Reference Trujillo and González2011). The total length of the mandible in specimens of P. maderensis is similar, although there is one individual from Cueva Roja with a much smaller mandible than the other individuals. Pipistrellus maderensis has a slightly lower mean talonid width of m3 compared to present-day Pipistrellus pipistrellus (0.597 mm; Sevilla Reference Sevilla1988) and P. kuhlii from Cueva Roja. However, we consider it very difficult to separate these species on the basis of biometry (see Table 2).
Table 2. Measurements of the cranial, mandible, teeth and humerus of Cueva Roja and Cueva de los Verdes. L: Length; W: Width.

Abbreviations: CBL = condyle basal length; CC = distance between the upper canines; CM3 = length of upper tooth row from canine to third molar; CrB = maximum width of the cranium base; CrL = total cranial length; IC = interorbital constriction width; M1M3 = length of upper molar row; M3M3 = distance between the upper third molars; ZB = maximum width between the two zygomatic arches; M1, M2 and M3: L1 = greater length, distance from the parastyle to the metastyle; L2 = medium length, measuring the constriction of the protoconal basin; and W = width, distance from the parastyle to the base of the protocone; cm3 = length of lower tooth row from canine to third molar; conan = distance between the condylar and the angular processes of the ramus; conW = width of the condylar process of the ramus; coran = distance between the coronoid and the angular processes of the ramus; m1m3 = length of lower molar row; mdl = total mandible length; raH = height of mandibular ramus; m1, m2 and m3: l = distance from the paraconid to the hypoconulid; w1 = trigonid width, distance from the metaconid to the protoconid; and w2 = talonid width, distance from the entoconid to the hypoconid; HL = length of humerus; EW = width of the humeral epiphysis.
In reference to the humerus, the lateral ridge of capitulum is hidden by the capitulumin anterior view, as in P. kuhlii and Pipistrellus nathusii, differing from P. pipistrellus where the lateral ridge of capitulum is visible above the capitulum (Dupuis Reference Dupuis1986).
The significant differences observed between the upper molars of current individuals and fossils of P. maderensis and P. kuhlii suggest that these materials should be ascribed to P. maderensis.
Pipistrellus kuhlii (Kuhl, 1817)
(Fig. 5)
Locality. Cueva de los Verdes, Lanzarote, Canary Islands, Spain.

Figure 5 Fossil cranial of Pipistrellus kuhlii from Cueva de los Verdes. (A1) Right maxilla (TFMC VO 35-23). (A2) Left maxilla (TFMC VO 35-23). (B1) Right maxilla (TFMC VO 35-7). (B2) Left maxilla (TFMC VO 35-7). (C1, C2) Right mandible (TFMC VO 35-11). (D1−D3) Left mandible (TFMC VO 35-12). (A1, A2, B1, B2, C2, D3) in occlusal view; (C1, D1) in lateral view; (D2) in lingual view.
Material. TFMC VO 35: left maxilla with C, M1, M2, M3 and right maxilla with P4, M1, M2, M3 (TFMC VO 35-2 (a)), mandible with i1, i2, i3, p4, m2, m3 (TFMC VO 35-2 (b)), right maxilla with M2, M3 and left maxilla with M2 (TFMC VO 35-4), right maxilla with C, P2, P4, M1, M2, M3 and left maxilla with I2, C, P2, P4, M1, M2, M3 (TFMC VO 35-6), left maxilla with C, P2, P4 and right maxilla with I1, I2, C, P2, P4, M2 (TFMC VO 35-7), left mandible with c, p2, p4, m1, m2, m3 (TFMC VO 35-9), right maxilla with all teeth and left maxilla with C, P2, P4, M1, M2, M3 (TFMC VO 35-11 (a)), right mandible with all teeth (TFMC VO 35-11 (b)), left maxilla with P2, P4, M2 and right maxilla with P2, P4, M1, M2, M3 (TFMC VO 35-12 (a)), left mandible with all teeth (TFMC VO 35-12 (b)), maxilla with all teeth (TFMC VO 35-23), right maxilla with P4, M1 and left maxilla with P4, M2, M3 (TFMC VO 35-24), left maxilla with I2, C, P2, P4, M1, M2, M3 (TFMC VO 35-25), left maxilla with worn M3 (TFMC VO 35-29), right maxilla with C, M1, M2, M3 and left maxilla with P2, P4, M2 (TFMC VO 35-35), right maxilla with P2, P4, M3 (TFMC VO 35-37), right maxilla with M2 (TFMC VO 35-38), right maxilla with P4, M3 and left maxilla with M3 (TFMC VO 35-40), left maxilla with C, M2 (TFMC VO 35-0).
Description. I1: the tooth is elongated with an oval outline in occlusal view. The main cusp is on the anterior side. The distal cingulum is present and is narrower on the anterolingual side. It has an accessory distal cusp of variable size. It may present an accessory cusp that is well developed (two out of four; TFMC VO 35-7 and TFMC VO 35-23) or poorly developed (two out of four); in one specimen, the right I1 has an accessory cusp, whereas in the left I1 it is poorly developed. There is one root.
I2: the tooth is circular in occlusal view with an anterolingual main cusp, which is connected by a crest to a cuspule on the labial side. There is a lingual accessory cusp. The labial cingulum is thicker distally. There is one root.
C: subquadrangular tooth in occlusal view, with a talon protruding on the posterolabial side. There is a groove on the labial side. There is a thick cingulum that starts on the lingual side and ends on the distolabial side, becoming narrower on the anterior side and almost disappearing on the anterolabial side. There is one root.
P2: P2 is small and circular in occlusal view and is situated in a lower position than the talon of the canine in posterolabial view. The main cusp is on the anterior side. A crest begins in the cuspid and then it extends to the distal side and ends with an enlargement. A narrow cingulum surrounds the tooth. There is one root.
P4: P4 is a quadrangular tooth. The labial margin is concave. There is a main mesiolabial cusp. The paracingulum is thick. The lingual cingulum is narrow and there is an accessory cusp on the anterolingual side. The talon is moderately developed and at its base it is slightly rounded. There are three roots.
M1: M1 is a trapezoid tooth. The parastyle is very well developed and is at a curved angle to the preparacrista. The mesostyle is of a similar size to the parastyle. The paracingulum is somewhat narrow and is connected to the parastyle. The precingulum is thin. The metastyle is at a right angle to the postmetacrista. The metacingulum is connected to the metastyle and does not vary in thickness. The ectoloph is slightly asymmetrical, and the metacone is the larger cusp. The labial cingulum of the paraflexus is well developed. The paraloph and the metaloph are small. Almost all the molars have a metaloph, although it is not very well developed (ten out of 11), and only in one specimen is there neither a paraloph nor a metaloph (one out of 11; TFMC VO 35-6). The metaconule is well developed. The talon and the paraconule are absent. The postprotocrista is not connected to the metacingulum. The lingual cingulum is thick. There are three roots.
M2: M2 is similar to M1 but shows greater variability in the characters of the paraloph and the metaloph. The metaloph may be well developed (one out of 21; TFMC VO 35-7), poorly developed (14 out of 21) or absent (six out of 21). Only in one specimen are both the paraloph and the metaloph absent (one out of 21; TFMC VO 35-35). Finally, in two individuals (two out of 15), the metaloph is absent in the right or left maxilla, whereas in the other maxilla it is present, albeit not very well developed (Fig. 5b). The postprotocrista is connected to the metaconule.
M3: M3 is a triangular tooth. There is a rounded and well-developed parastyle connected at a curved angle to the preparacrista. The paracingulum is thick and is connected to the parastyle. The postparacrista and the premetacrista are small. The metacone is present and there is a distal spur. The metastyle is absent. The paraloph may be present (nine out of 14) or not (five out of 14; TFMC VO 35-2 (a), TFMC VO 35-4, TFMC VO 35-29, TFMC VO 35-35 and TFMC VO 35-37). The labial cingulum of the metaflexus is very thin. In one specimen, the left M3 has a small part of the postmetacrista, whereas in the right M3 it is absent. The mesostyle is reduced to half of the paracone. The precingulum and the lingual cingulum are short. There are three roots.
i1: In occlusal view, there are three cuspids in line. It has a well-developed labial cingulid. There is one root.
i2: in occlusal view, it has three cuspids in line and a small distal cingulid and has other cingulid at the labial side. There is one root.
i3: in occlusal view, it has a trapezoidal outline with three cuspids in line and a well-developed distal cingulid, and has another cingulid at the labial side. There is one root.
c: the tooth is oval in occlusal view. The lingual and the labial sides are convex and the anterior and the posterior sides are concave. The main cusp is mesial and there is an enlargement of the cingulid on the mesiolingual side. On the distolingual side of the cingulid there is a thickening. The labial cingulid is well developed. There is one root.
p2: the tooth is oval in occlusal outline and is very reduced. There is a main mesial cuspid and two small accessory cuspules directed to the anterolingual and distolingual sides. A thick cingulid encircles the tooth. There is one root.
p4: p4 is a quadrangular tooth. There is a main mesiolingual cuspid, and there is a thick cingulid surrounding the tooth. There is an accessory cuspule on the anterolingual side. The lingual cingulid is greatly thickened distally. The labial cingulid is concave in labial view in the area of the root. There are two roots.
Lower molars: the protoconid is the highest cuspid. The hypoconid is the second largest cuspid in m1 and m2, but in m3 it is reduced and is similar in height to the paraconid and the metaconid. The paraconid, metaconid and entoconid are similar in height in m1 and m2, but not in m3, where the entoconid is slightly lower. The lower molars slightly decrease in size towards m3. There is a sizeable labial cingulid that is larger on the anterior side and narrower distally, especially in m3. The labial cingulid is variable in width. It may be intermediate or wide, but the thickness is always consistent among the three lower molars. There are two roots.
m1: the tooth is nyctalodont. In one specimen (TFMC VO 35-12), the labial cingulid is very well developed and in three more it is less well developed. The paraconid notch is absent. The trigonid is closed. The paralophid is not angular. The entocristid is concave–convex. The oblique crista ends in the middle of the posterior wall of the trigonid. The hypoconulid is slightly more lingual than the entoconid.
m2: this tooth is similar to m1, but the paralophid is angular.
m3: this tooth is similar to m1. The talonid is slightly reduced toward the entoconid. The location of the hypoconid in m3 is more labial than in m2.
Remarks. Current populations of this species show variability in the accessory cusp of I1. Regarding the morphology of the upper molars, the paraconule is absent in current Iberian and Canary Island populations (Sevilla Reference Sevilla1986; pers. obs. 2021), whereas in Central and Eastern European populations it is well developed (Rosina & Sinitsa, Reference Rosina and Sinitsa2014). The variability in the development or absence of the metaloph of Pipistrellus kuhlii fossil individuals is also observed in the present-day individuals. Menu (Reference Menu1987) proposes that P. kuhlii presents a monocuspid I1. By contrast, Sevilla (Reference Sevilla1986) defines P. kuhlii as having a variable accessory cusp, as occurs in the material studied here.
In comparison with the dental morphology of other species of this genus, Pipistrellus pipistrellus presents a well-developed secondary cusp in I1 (Menu Reference Menu1987), whereas in P. kuhlii or Pipistrellus maderensis it is variable. As in P. pipistrellus, the lower incisor teeth of P. kuhlii and P. maderensis do not have a diastema, whereas Pipistrellus nathusii has diastemata between i2 and i3 and between i3 and the lower canine (Dupuis Reference Dupuis1986). In both P. kuhlii and P. maderensis the outline of p2 is oval, whereas in P. nathusii and P. pipistrellus it is triangular, the p2 of P. nathusii being more elongated than that of P. pipistrellus (Dupuis Reference Dupuis1986). Also notable is that unlike the species of this genus present in the Canary Islands, P. nathusii presents myotodont lower molars (Sevilla Reference Sevilla1986). As in P. maderensis, both P. nathusii and P. pipistrellus have a smaller talonid than that observed in P. kuhlii (Sevilla Reference Sevilla1986).
In a preliminary analysis, Trujillo (Reference Trujillo1991) identified P. kuhlii in this site, and our studies now confirm the presence of this species. Given the absence of the paraconule, the population of the Canary Islands may be descended from Iberian populations.
4. Discussion
4.1. Palaeobiogeographic implications
The bat fossil record studied here stems from the oldest (Lanzarote) and youngest (El Hierro) islands of the Canary archipelago (Trujillo Reference Trujillo1991; Castillo et al. Reference Castillo, López, Martín and Rando1996). It is composed of three species with extant populations, two of which are endemic: one, Plecotus teneriffae, from the Canary Islands; and another, Pipistrellus maderensis, from the Canary Islands, Madeira and the Azores (Fajardo & Benzal Reference Fajardo and Benzal2002; Lopes & Medeiros Reference Lopes and Medeiros2011). The third species has a distribution across North Africa and southern Europe (Pipistrellus kuhlii; Palomo et al. Reference Palomo, Gisbert and Blanco2007; Juste & Paunović Reference Juste and Paunović2016a, Reference Juste and Paunović2016b, Reference Juste and Paunović2016c).
Bats of the genus Plecotus are known as the long-eared bats. They have a wide distribution predominant in the Palearctic and at the margins of the Afrotropical and Indo-Malayan regions (Fukui et al. Reference Fukui, Tu, Thanh, Arai, Harada, Csorba and Son2020). The genus Plecotus has split into two clades: the ‘austriacus group’ distributed across Europe and north Africa, and the ‘auritus group’ with a distribution across the Middle East and Europe. This separation took place during the Miocene/Pliocene boundary at least 5 Ma (Juste et al. Reference Juste, Ibáñez, Muñoz, Trujillo, Benda, Karataş and Ruedi2004; Spitzenberger et al. Reference Spitzenberger, Strelkov, Winkler and Haring2006). The ‘austriacus group’ includes the species Plecotus austriacus found in Cape Verde and Madeira (where it arrived from Europe), and Pl. teneriffae of the Canary Islands (Tenerife, La Palma and El Hierro), related to Plecotus kolombatovici Dulic, 1980 from the N of Africa (Spitzenberger et al. Reference Spitzenberger, Strelkov, Winkler and Haring2006). This indicates the African origin of the endemic Pl. teneriffae, which may have become differentiated during the early Pleistocene (2.3 Ma; Juste et al. Reference Juste, Ibáñez, Muñoz, Trujillo, Benda, Karataş and Ruedi2004). At this point, two of the westernmost islands, El Hierro and La Palma, had not yet emerged (Troll & Carracedo Reference Troll and Carracedo2016).
Like the genus Plecotus, the species Pipistrellus kuhlii has a Palearctic distribution with a possible origin in Central Asia (Hukov et al. Reference Hukov, Timofieieva, Prylutska, Rodenko, Moiseienko, Bohodist, Domanska and Vlaschenko2020). In the mid-Atlantic islands this species can today be found in the Canary Islands (Fuerteventura, Gran Canaria and Tenerife; Fajardo & Benzal Reference Fajardo and Benzal2002) and in the archipelago of Cape Verde (São Vicente, Santo Antão, São Nicolau, Fogo and Santiago; Vasconcelos Reference Vasconcelos2018; Borloti et al. Reference Borloti, Dinis and Vasconcelos2020). However, its fossil record shows that in the Canary Islands it could have been more widely distributed in the past, with populations in Lanzarote (Trujillo Reference Trujillo1991; Masseti Reference Masseti2010). In contrast, Pipistrellus maderensis is found in the western Canary Islands, coinciding with P. kuhlii only in Tenerife (Fajardo & Benzal Reference Fajardo and Benzal2002), as well as being found in Madeira and the Azores (Flores and Santa Maria; Lopes & Medeiros Reference Lopes and Medeiros2011; Trujillo & González Reference Trujillo and González2011; Torre et al. Reference Torre, Fernández-Lugo, Guarino and Fernández-Palacios2018).
Genetic studies undertaken by Pestano et al. (Reference Pestano, Brown, Suárez and Fajardo2003) indicated that the two extant species of the genus Pipistrellus in the Canary Islands (P. kuhlii and P. maderensis) form a monophyletic clade. The age of the fossil remains from El Hierro and Lanzarote precludes inferences about the arrival of these microbats in the archipelago, but we can raise the hypothesis of colonisation. The studied P. kuhlii bones were found in Lanzarote in a lava tube dated by 40Ar/39Ar to 21 ± 6.5 ka. This age coincides with a drop in sea level associated with a glacial maximum (Carracedo et al. Reference Carracedo, Singer, Jicha, Guillou, Rodriguez, Meco, Pérez-Torrado, Gimeno, Socorro and Lainez2003), and suggests migration from the Iberian Peninsula/North Africa.
This age suggests that Lanzarote was the first island to be colonised when the species arrived in the Canary Islands. Nowadays, the current population is distributed across all altitudinal gradients in Fuerteventura (807 m maximum altitude in Pico de la Zarza), as well as in medium and high mountainous areas in Tenerife and Gran Canaria, and without record in Lanzarote (Fajardo & Benzal Reference Fajardo and Benzal2002).
Due to the fact that the coexistence of the two species of the genus Pipistrellus has only been cited on one island, and given that there are problems in differentiating them using the phenotype and echolocation, it is necessary to improve our knowledge of their teeth. In this context, the fossil record has proven to be a very useful tool for differentiating the two species on the basis of their dental morphology. In addition, the data obtained indicate that their former distribution extended to an area where there is no current record. In the case of Plecotus teneriffae, the fossil record coincides with the current distribution (https://www.biodiversidadcanarias.es/biota/).
4.2. Palaeoecological implications
Plecotus teneriffae is a current species endemic to the Canary Islands that lives at altitudes between 150 and 2300 m above sea level (Alcaldé & Juste Reference Alcaldé and Juste2016a, Reference Alcaldé and Juste2016b). Its distribution is linked to crevices and caves, whether natural or artificial, but never where humans live, unlike its most closely related species Plecotus auritus and Plecotus austriacus (Hutterer et al. Reference Hutterer, Ivanova, Meyer-Cord and Rodrigues2005). Tree holes and bat or bird boxes are never used (Benzal & Fajardo Reference Benzal and Fajardo1997, Reference Benzal, Fajardo, Mitchell-Jones, Amori, Bogdanowicz, Kryštufek, Reijnders, Spitzenberger, Stubbe, Thissen, Vohralík and Zima1999). However, in general it inhabits a wider range of roost sites than the aforementioned species because it has fewer competitors. Nowadays, this species does not appear on the eastern islands because it does not tolerate xeric environments or the absence of water, or because the insect assemblage is not suitable. The species is a forest dweller (especially coniferous and to a lesser degree laurel forest), although it occasionally forages in more open and arid areas, such as among fruit trees and vegetable gardens. It hunts in the above-mentioned habitats and in caves, in groups, stalking, on surfaces and rarely in flight, and it avoids hunting in open habitats (Benzal & Fajardo Reference Benzal and Fajardo1997, Reference Benzal, Fajardo, Mitchell-Jones, Amori, Bogdanowicz, Kryštufek, Reijnders, Spitzenberger, Stubbe, Thissen, Vohralík and Zima1999). The principal food source is lepidopterans, dipterans and a few coleopterans. It is a sedentary species (Hutterer et al. Reference Hutterer, Ivanova, Meyer-Cord and Rodrigues2005).
Pipistrellus maderensis is the most abundant species in the present-day assemblage, and the species with most contact with humans (Fajardo & Benzal Reference Fajardo and Benzal2002). It lives in a wide range of habitats, and roosts in a wide variety of places such as crevices in stone walls, bat and bird boxes, human dwellings, tree holes and caves, among others. It is often associated with human populations, although in contrast with Pipistrellus pipistrellus in Europe, it is not often found in parks, gardens and urban areas in the Canary Islands (Alcaldé & Juste Reference Alcaldé and Juste2016a, Reference Alcaldé and Juste2016b). Its habitat ranges from sea level to 2150 m above sea level. Its hunting area includes towns, forests, caves and ponds. Gregarious behaviour is poorly developed, although on some occasions several dozen specimens may gather. Its favourite prey is flying insects, including small moths and dipterans. It is a sedentary species (Trujillo & González, Reference Trujillo2011).
Pipistrellus kuhlii roosts in a wide range of habitats, including bird and bat boxes, stone wall crevices and houses, among others, and it is associated with human settlements (Juste & Paunović Reference Juste and Paunović2016a, Reference Juste and Paunovićb, Reference Juste and Paunovićc). Its habitat ranges from sea level to 1625 m above sea level, but it prefers low altitudes, being a thermophile species, and nowadays it is not found on El Hierro. It hunts in a wide range of areas, including ponds, towns, caves, agricultural areas, among others, like Plecotus teneriffae and Pipistrellus maderensis (Fajardo & Benzal Reference Fajardo and Benzal2002). It prefers open areas such as fields and waterways, although it is generally not far from tree vegetation. This bat feeds on small insects, including dipterans, psocopterans and coleopterans. In North Africa, it forages in forest as well as semi-desert environments, and it is found in temperate grassland and Mediterranean-type shrubland (Hutterer et al. Reference Hutterer, Ivanova, Meyer-Cord and Rodrigues2005). It is probably a sedentary species (Hutterer et al. Reference Hutterer, Ivanova, Meyer-Cord and Rodrigues2005; Palomo et al. Reference Palomo, Gisbert and Blanco2007).
In summary, the presence of Pl. teneriffae in Cueva Roja demonstrates the existence of a humid forest near the entrance to the cave since, as it is a sedentary species, they would not be transient specimens. The presence of only P. kuhlii in Cueva de los Verdes, without any of the other species, suggests a more xeric and thermophile environment than Cueva Roja. This is consistent with the current environment of El Hierro (where Cueva Roja is located), which is the most southwestern island in the archipelago, with a more humid and wooded environment due to the oceanic influence than Lanzarote (where Cueva de los Verdes is located). This latter island is one of the most northeastern islands, with a less forested and more xeric environment due to the Saharan influence (del Arco & Rodríguez Reference Del Arco and Rodríguez2018).
5. Conclusions
Fossil bat remains have been found and studied in the geological context of volcanic and oceanic islands (Canary Islands). These remains belong to three extant species, two of the genus Pipistrellus (Pipistrellus maderensis and Pipistrellus kuhlii) and one of the genus Plecotus (Plecotus teneriffae). These remains were found inside lava tubes dating to the upper Pleistocene and Holocene. The fossil remains of Plecotus teneriffae were found on the island of El Hierro and differ morphologically from other species of Plecotus present in Europe and North Africa, fundamentally in the height of the paracone and metacone of M3 and in the cingulid of p4 and the lower molars.
Our results make it possible to distinguish between extant and fossil specimens of P. maderensis and P. kuhlii present in the Canary Islands. The diagnostic characters that separate the two species relate to the differential development of the paraloph, metaloph and metaconule in M1 and M2, which in P. maderensis are more developed than in P. kuhlii. The presence of bone remains of P. kuhlii has been confirmed on the island of Lanzarote. However, at present there is no evidence for the existence of living populations on the island.
It will be necessary to undertake a comparison between the dentition of P. kuhlii specimens from the Canary Islands and from North Africa to confirm the presence or absence of the paraconule in the upper molars. This would help to establish the African or Iberian origin of this species in the Canary Islands.
The discovery of Pl. teneriffae in Cueva Roja indicates the presence of a humid forest near the entrance to the cave, whereas the presence of only P. kuhlii in Cueva de los Verdes, without any of the other species, suggests a more xeric and thermophile environment than Cueva Roja. This is consistent with the current environments of El Hierro and Lanzarote.
The palaeobiological data obtained in this work are relevant to the conservation of current species in the following respects: (i) they allow us to distinguish bat species from the dentition; (ii) they shed light on the ancient distribution, extirpations or extinctions of vulnerable endemic and non-endemic bat species in the islands of the mid-Atlantic Ocean; and (iii) they increase what is known of a group of flying animals that are of great benefit to humanity due to the resources and processes with which they provide natural ecosystems.
6. Acknowledgements
This work has benefited from the financial support of the research projects PROID1017010136 (C.C.R., P.C-C.), FCT-17-12775 (C. C. R., P. C.-C.); the Spanish Ministerio de Economía, Industria y Competitividad and the European Regional Development Fund of the European Union (MINECO/FEDER EU, project CGL2016-75062-P to C. C. R.). We acknowledge Dr María Esther Martín González, in charge of the area of Paleontology and Geology of the Museo de la Naturaleza y la Arqueología (MUNA), for providing the current material on bats in the Canary Islands. This work forms part of the activities of the Universidad de La Laguna research groups ‘Estudio de la fauna Fósil y actual de Canarias’ and ‘Crecimiento submarino y emersión de las Islas Canarias: estudio geológico de los Complejos Basales’. Rupert Glasgow revised the text in English. The support of the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) toward this research for J. González-Dionis and V. D. Crespo is hereby acknowledged.







