1. Introduction
The biotic recovery after the end-Permian mass extinction is certainly not a synchronous event in all areas because its record strongly depends on the type of depositional setting (Pruss et al. Reference Pruss, Bottjer, Corsetti and Baud2006; Beatty, Zonneveld & Henderson, Reference Beatty, Zonneveld and Henderson2008; Zonneveld, Gingras & Beatty, Reference Zonneveld, Gingras and Beatty2010; Mata & Bottjer, Reference Mata and Bottjer2011). Among the various Early Triassic marine environments, the nature of the sedimentation and putative fossil record were controlled by sea-level changes (Heydari et al. Reference Heydari, Hassanzadeh, Wade and Ghazi2003; Weidlich, Reference Weidlich2007). The Triassic is marked by a global long-term rise in sea level with several second- and third-order transgressive-regressive sequences that can be identified in different localities throughout the world (Haq, Hardenbol & Vail, Reference Haq, Hardenbol and Vail1987; Hallam & Wignall, Reference Hallam and Wignall1999; Haq & Al-Qahtani, Reference Haq and Al-Qahtani2005). In SW Utah, the Lower Triassic sedimentary succession records three third-order transgressive-regressive cycles that originated in the Griesbachian, Smithian and earliest middle Spathian (Embry, Reference Embry1997; Paull & Paull, Reference Paull, Paull, Dickins, Zunyi, Hongfu, Lucas and Acharyya1997). If such long-term sea-level changes potentially controlled the evolution of the Triassic marine biota (Konstantinov, Reference Konstantinov2008; Ruban, Reference Ruban2008; Kelley et al. Reference Kelley, Motani, Jiang, Rieppel and Schmitz2013), higher-frequency sea-level fluctuations also impacted the distribution and record of the benthic marine faunas in the aftermath of the end-Permian mass extinction (Mata & Bottjer, Reference Mata and Bottjer2011). Although high-frequency sea-level changes are suspected in several Lower Triassic sedimentary successions (Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007; Chen, Fraiser & Bolton, Reference Chen, Fraiser and Bolton2012; Wu et al. Reference Wu, Zhang, Feng, Jiang, Li and Yang2012), they are rarely studied in detail because they cannot be easily dated and correlated (Embry, Reference Embry1997). The nature and influence of high-frequency relative sea-level changes on the Early Triassic marine biota distribution and its record are therefore still poorly known (Yang & Lehrmann, Reference Yang and Lehrmann2003).
During Early Triassic time, the Sonoma Foreland Basin of western USA was a place of continental and marginal marine sedimentation (McKee, Reference McKee1954; Blakey, Reference Blakey1974, Reference Blakey1977; Collinson, Kendal & Marcantel, Reference Collinson, Kendall and Marcantel1976). In SW Utah, the lowermost Lower Triassic stratigraphic unit belonging to the Thaynes Group (sensu Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007) is well exposed in Timpoweap Canyon near Hurricane, SW of Zion National Park (Gregory, Reference Gregory1950; Blakey, Reference Blakey1979; Nielson, Reference Nielson1991). Blakey (Reference Blakey1979) suggested that the Timpoweap Member was deposited in a transitional area between continental and marine settings. Lucas, Krainer & Milner (Reference Lucas, Krainer, Milner, Lucas and Spielmann2007) illustrated the type section of the Timpoweap Member (correlative of the Sinbad Formation of SE Utah) in Timpoweap Canyon. Although these authors demonstrated a general sea-level rise during Early Triassic time, they only suspected higher-frequency sea-level changes. The present study is a detailed analysis of an exceptional outcrop in Timpoweap Canyon that allows the illustration of high-frequency depositional sequences. The objectives of this paper are to (1) reconstruct the evolution of the depositional settings; (2) highlight high-frequency sea-level changes and shoreline migrations; and (3) discuss their potential origin and influence on the distribution and record of marine organisms.
2. Geological framework
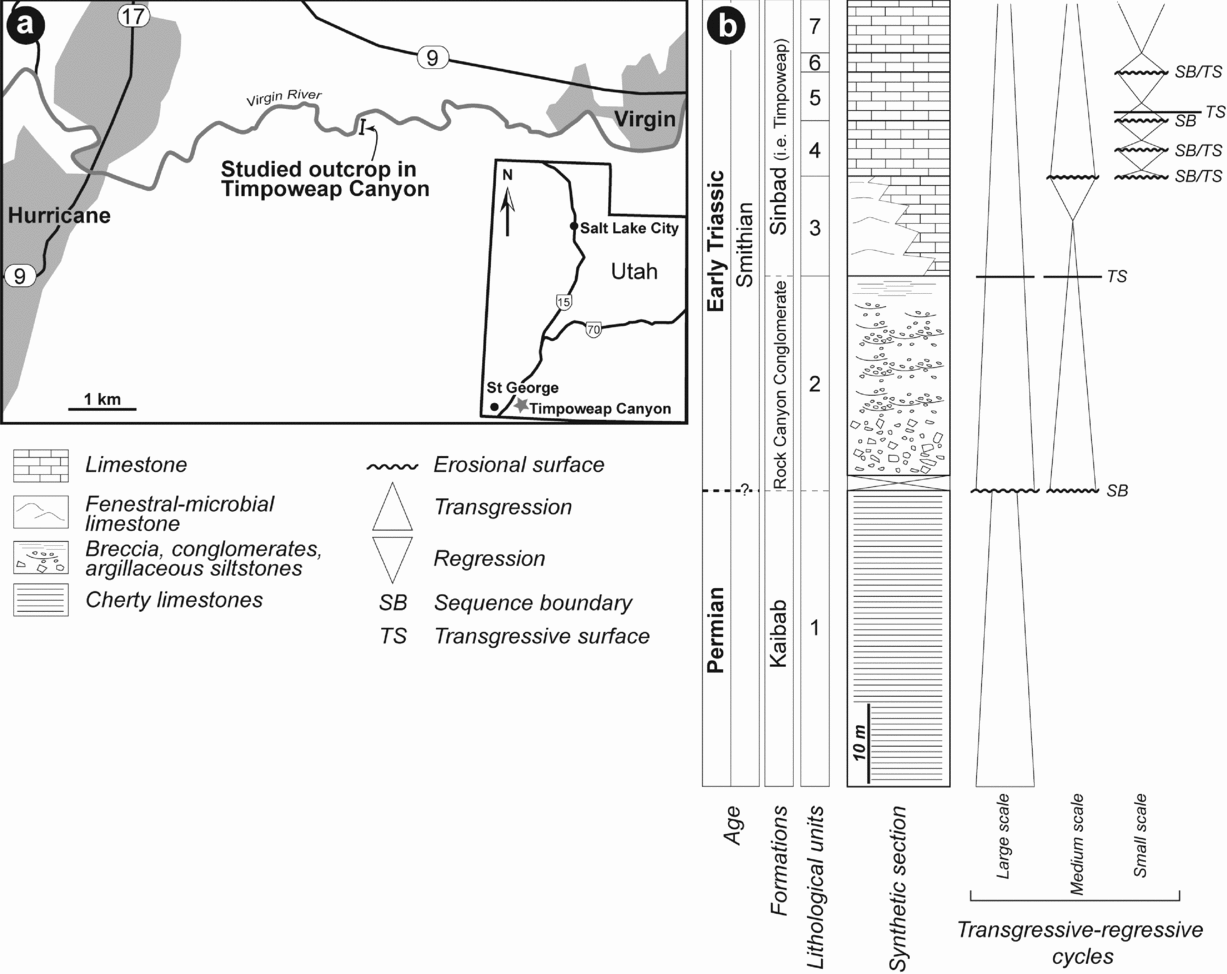
The study area is located in Washington County (SW Utah, USA), c. 30 km NE of St George (Fig. 1a). Directly south of the road that connects Hurricane to Virgin, the Timpoweap Canyon of the Virgin River reveals an impressive Permian–Triassic sedimentary succession (Blakey, Reference Blakey1979; Nielson, Reference Nielson1991). The river gorge, which ranges up to 120 m in depth, exposes the Late Permian Harrisburg Member of the Kaibab Formation, which is overlain by the Triassic Timpoweap Member of the Moenkopi Formation (Stewart, Poole & Wilson, Reference Stewart, Poole and Wilson1972; Blakey, Reference Blakey1979; Fig. 1b) or Thaynes Group (sensu Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). Regionally, the Kaibab Formation displays an upper contact that corresponds to an erosional unconformity (Jenson, Reference Jenson1986), which reveals a residual karst topography (Blakey, Reference Blakey1979). At most locations, erosion reaches a stratigraphic unit of the Harrisburg Member made of dolomitized fossiliferous limestones that grade upwards into chert and weather to form a cliff (Nielson, Reference Nielson1991). Overlying the deposits of the Kaibab Formation is a laterally discontinuous unit made of breccia and conglomerates initially defined by Reeside & Bassler (Reference Reeside and Bassler1922) as the Rock Canyon Conglomerate Member. In the literature, this lithostratigraphic unit is placed alternatively in the Kaibab Formation (Gregory, Reference Gregory1950) or in the Moenkopi Formation (Reeside & Bassler, Reference Reeside and Bassler1922; Blakey, Reference Blakey1979). The occurrence of conglomerates in both the Harrisburg and Timpoweap Members may explain the difficulty in delimiting the stratigraphic position of these deposits (Blakey, Reference Blakey1979). Gregory (Reference Gregory1950) assumed that the Rock Canyon Conglomeratic Member was equivalent in part to the Harrisburg Member of the Kaibab Formation and proposed that the term be abandoned. Alternatively, Nielson (Reference Nielson1991) proposed that the Rock Canyon Conglomerate should be considered as a separate Upper Permian – Lower Triassic formation. However, there is a facies gradation between the Rock Canyon Conglomerate and the limestones of the Timpoweap Member (Nielson, Reference Nielson1991). Such a gradation suggests that the conglomeratic deposits and overlying limestones belong to the same stratigraphic unit. The Timpoweap Member is therefore the first member of the Moenkopi Formation and corresponds to a lithologically variable unit composed mainly of sandy limestone to sandstones or siltstones, massive to thick-bedded grey limestone and chert pebble conglomerate lying above Permian rocks (Gregory, Reference Gregory1950; Stewart, Poole & Wilson, Reference Stewart, Poole and Wilson1972; Blakey, Reference Blakey1979; Nielson, Reference Nielson1991). More recently, Goodspeed & Lucas (Reference Goodspeed, Lucas, Lucas and Spielmann2007) and Lucas, Krainer & Milner (Reference Lucas, Krainer, Milner, Lucas and Spielmann2007) amended the regional stratigraphic nomenclature and assigned the Sinbad and Moenkopi to formation- and group-rank units, respectively. They also restricted the Moenkopi Group to continental deposits (e.g. Rock Canyon Conglomerate and Lower Red formations) and the Thaynes Group to marine deposits (e.g. Sinbad and Virgin formations). Although still under debate, such nomenclature was recently followed by Hofmann et al. (Reference Hofmann, Hautmann, Wasmer and Bucher2013) and Zatoń, Taylor & Vinn (Reference Zatoń, Taylor and Vinn2013) and is retained in this work. Moreover, Lucas, Krainer & Milner (Reference Lucas, Krainer, Milner, Lucas and Spielmann2007) considered the Timpoweap Member as a correlative and junior synonym of the Smithian Sinbad Formation.
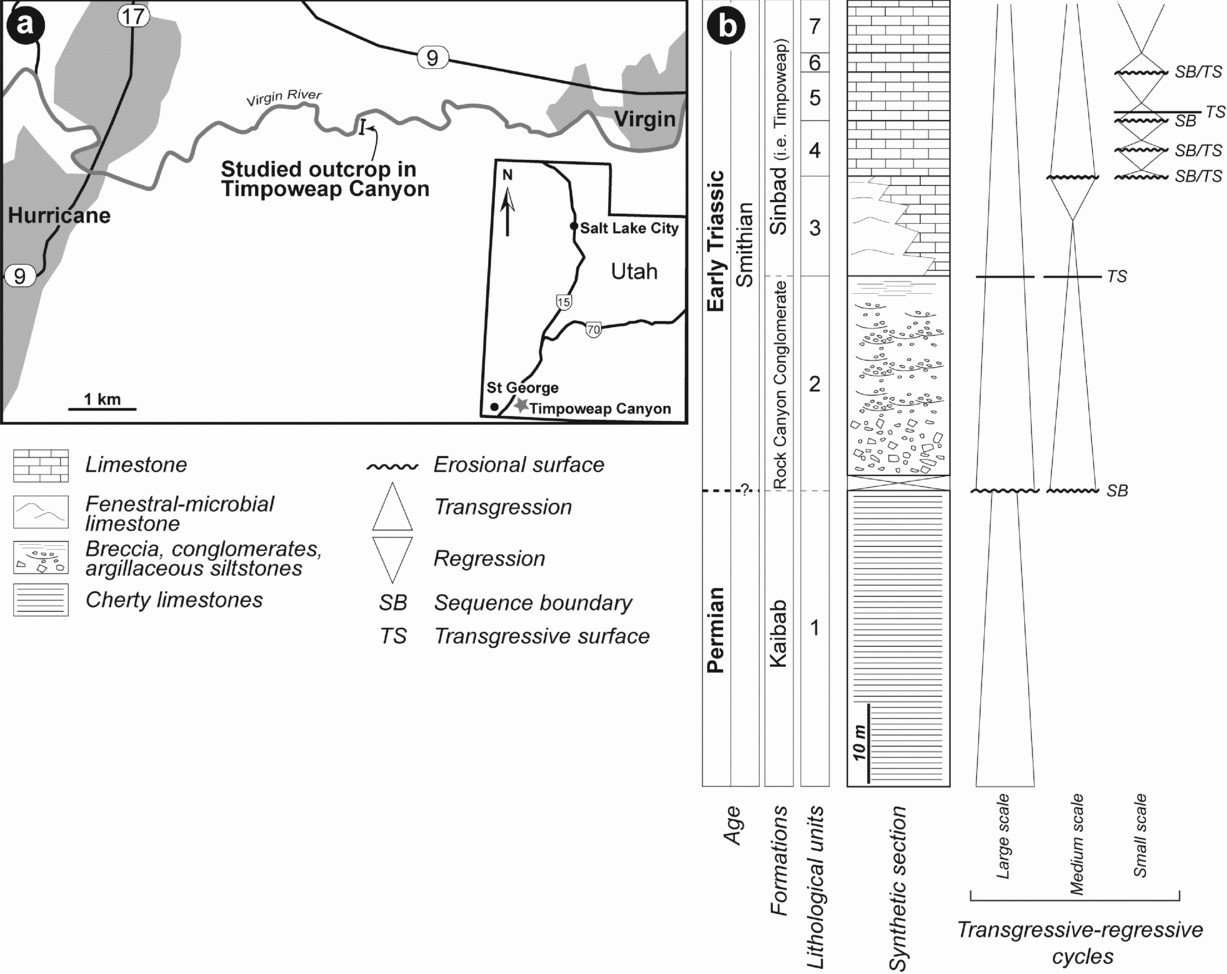
Figure 1. (a) Geographic location of Timpoweap Canyon in SW Utah. (b) Synthetic observed lithological succession with interpreted transgressive-regressive sequences.
The Sonoma Foreland Basin of the western USA was located at a near-equatorial position in eastern Panthalassa during the Early Triassic. Smithian marine outcrops are widely distributed within a large area covering Wyoming, Idaho, Utah and Nevada (Goodspeed & Lucas, Reference Goodspeed, Lucas, Lucas and Spielmann2007). They generally contain abundant fossils and a detailed regional biostratigraphic ammonoid-based zonation has been recently proposed for this substage (Brayard et al. Reference Brayard, Bylund, Jenks, Stephen, Olivier, Escarguel, Fara and Vennin2013). In the White Hills, the lower Spathian Virgin Formation is in disconformable contact with the Harrisburg Member, indicating that the youngest Permian–Triassic palaeogeographic highs were located west of St George (Jenson, Reference Jenson1986; Nielson, Reference Nielson1991). A previous facies analysis of deposits of the Smithian Sinbad Limestone (i.e. Timpoweap) Formation suggested that they were deposited along the shoreline of the seaway parallel to the present Hurricane Cliffs, and that the open sea was to the east (Blakey, Reference Blakey1979).
The Kaibab Formation in the region of Virgin is Permian in age, whereas the overlying Thaynes Group is largely Smithian and Spathian based on several index ammonoid occurrences (e.g. Meekoceras, Anasibirites, Tirolites and possibly Columbites; Stewart, Poole & Wilson, Reference Stewart, Poole and Wilson1972; Jenson, Reference Jenson1986; Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007; Hofmann et al. Reference Hofmann, Hautmann, Wasmer and Bucher2013). The Smithian Meekoceras gracilitatis and Anasibirites kingianus zones were also assumed to be locally present in the Sinbad Formation (i.e. Timpoweap Member of Stewart, Poole & Wilson, Reference Stewart, Poole and Wilson1972) of Timpoweap Canyon. The top of Timpoweap Canyon corresponds to the late Smithian Anasibirites kingianus zone. Dating of the uppermost units of the canyon by means of conodonts also indicated a late Smithian age (abundant ellisonids; see also Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). Dating of underlying units (i.e. microbial limestones of the Sinbad Formation; Fig. 1b) is difficult as conodonts are rare and fragmentary, but one ammonoid (Owenites sp.) gives a middle Smithian age for the deposits just above the fenestral-microbial limestone unit. Lithological and biostratigraphical correlations of the Timpoweap Canyon succession with neighbouring Smithian outcrops near Cedar City and Kanarraville is consistent with a middle Smithian age for the limestones of the base of the Sinbad Formation (Brayard et al. Reference Brayard, Bylund, Jenks, Stephen, Olivier, Escarguel, Fara and Vennin2013).
3. Lithological units
The studied area in Timpoweap Canyon corresponds to a N–S cliff that ranges up to c. 80 m in depth and c. 300 m in length (Figs 1a, 2a, b). Stratigraphically, the succession exposed in this cliff can be subdivided into seven lithological units (Fig. 1b). The first unit corresponds to the Permian limestones of the Kaibab Formation, while the second corresponds to sedimentary deposits of the Rock Canyon Conglomerate Formation and the last five units correspond to limestone deposits of the Smithian Sinbad Formation (Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). At the uppermost part of the cliff it is not possible to observe the contact with the overlying red siltstones and fine sandstones of the Spathian Lower Red Formation, which are present in the lectostratotype section (Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). These lithological units are described below according to their facies, stacking pattern and main stratigraphic surfaces.

Figure 2. (a) Photograph and (b) line drawing of the main stratigraphic units (U2–U7) at Timpoweap Canyon. (c–e) Close-ups of some stratigraphic architectures. (c) Dashed line represents lateral facies variation between microbial-fenestral limestones and dolomudstones (lithological unit 3); first bed of lithological unit 4 onlaps on the truncation surface at the top of lithological unit 3. Small black and white arrows: stratal terminations. (d) Truncation at the top of lithological unit 3 and high-angle clinoforms of the upper wedge (subunit 4a). (e) Truncated upper part of lithological unit 3 and high-angle clinoforms (subunit 4b).
3.a. Lithological unit 1
Even although it is beyond the scope of this study, lithological unit 1 is visible at the base of the gorge of the Virgin River and corresponds to the Permian limestones of the Kaibab Formation. Lucas, Krainer & Milner (Reference Lucas, Krainer, Milner, Lucas and Spielmann2007) gave a detailed description of the Kaibab Formation in the lectostratotype section of Timpoweap Canyon. It consists of silicified and recrystallized limestones (including only rare ostracods and brachiopods) associated with chert layers. In the study area, well-bedded deposits of this first lithological unit display abundant silicified burrows that reflect an intense infaunal activity. The contact with the overlying lithological unit was not observed due to vegetation cover and recent colluvium. However, this contact is regionally well known and corresponds to an erosional unconformity with a residual karst topography (Blakey, Reference Blakey1979; Jenson, Reference Jenson1986).
3.b. Lithological unit 2
Lithological unit 2 corresponds to the Rock Canyon Conglomerate Formation and can be subdivided into three subunits. A multi-metre-thick chert breccia is observed in the lowermost part of lithological unit 2. This subunit is composed of reworked, angular chert clasts up to 20 cm in diameter (Figs 1b, 3a). No sedimentary structures or clast organization were observed and clasts are embedded in an abundant matrix of coarse-grained sandstone. Overlying this breccia subunit is a multi-metre-thick conglomerate subunit made of well-rounded to subangular chert clasts. These deposits display low-angle cross-bedding and lateral accretion structures (Figs 1b, 3b). The uppermost part of lithological unit 2 displays a decimetre- to metre-thick interval of red mudstone to siltstone with sparse chert clasts (Figs 1b, 3c). In this third subunit, clasts are small (up to 2 cm in diameter) and well rounded. Silty lamina display small asymmetrical ripples. This last subunit shows lateral thickness changes and can be absent locally.

Figure 3. Outcrop close-up of depositional facies of lithological unit 2 and the base of lithological unit 3 at Timpoweap Canyon: (a) breccia (see hammer for scale); (b) conglomerate (see hammer for scale); (c) red sandstone; and (d) basal part of the fenestral-microbial limestones of lithological unit 3 (note the abundant extraclasts).
3.c. Lithological unit 3
The base of lithological unit 3 shows abundant cherty exotic clasts that gradually decrease in abundance in the first few metres of the unit (Fig. 3d). This indicates that the transition with lithological unit 2 reflects a facies gradation (as observed by Nielson, Reference Nielson1991) rather than a sharp surface (as suggested by Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). Lithological unit 3 also displays lateral facies variations. A first facies corresponds to a massive fenestral-microbial limestone interval in the northernmost part of the studied area (Fig. 2a, b). At the outcrop scale, several undulated surfaces can be laterally followed over several tens of metres, suggesting a stratified unit. These surfaces underline the presence of metre-scale patches (Figs 1b, 2b, 4a, b). This fenestral-microbial limestone interval has an upper truncation surface that gently dips to the south (Fig. 2). Two main subfacies comprise these fenestral-microbial limestones. The first subfacies is a fenestral packstone/bindstone with laminoid fenestrae, peloids, stromatolites, cortoids and micritic ooids (Fig. 4c). Its fauna is sparse, with some ostracods, gastropods and bivalve fragments. Very rare and poorly preserved meshworks of spicules suggest the presence of putative siliceous sponges. A second subfacies can be assimilated to a pisoid biocemenstone. In this subfacies, particularly large fenestrae display domal or columnar stromatolites with upward, lateral and downward growth directions (Fig. 4d). Other large voids are unfilled by a succession of sparry carbonate cements and micritic crusts (Fig. 4e). Some domal micritic crusts display laminated sheets (Fig. 4f). Micrite or cement crusts coat various grains (Fig. 4g). Meniscus cements occurred between pisoids (Fig. 4h). There are also subrounded to rounded aggregates and locally some muddy breccias. Subrounded and poorly sorted grains of detrital quartz are abundant in the first basal metre of fenestral-microbial unit 3, but they are rare in the rest of the unit.

Figure 4. (a, b) Field view and sketch of the internal structure of fenestral-microbial limestone of lithological unit 3; note the lateral accretion inside metre-scale patch reefs (black arrow). (c–h) Various microfacies of this fenestral-microbial limestone. (c) Fenestral packstone with a framework made of abundant peloidal grains and some bivalve fragments. (d) Columnar microbial growth at the roof and at the floor of a large fenestral void. (e) Succession of sparry carbonate cements and micritic crusts in a large void area. (f) Domal micritic crusts with laminated sheets. (g) Pisoids with directional elongations; latest micritic laminae show interpisoid linkage. (h) Meniscus cements between pisoids.
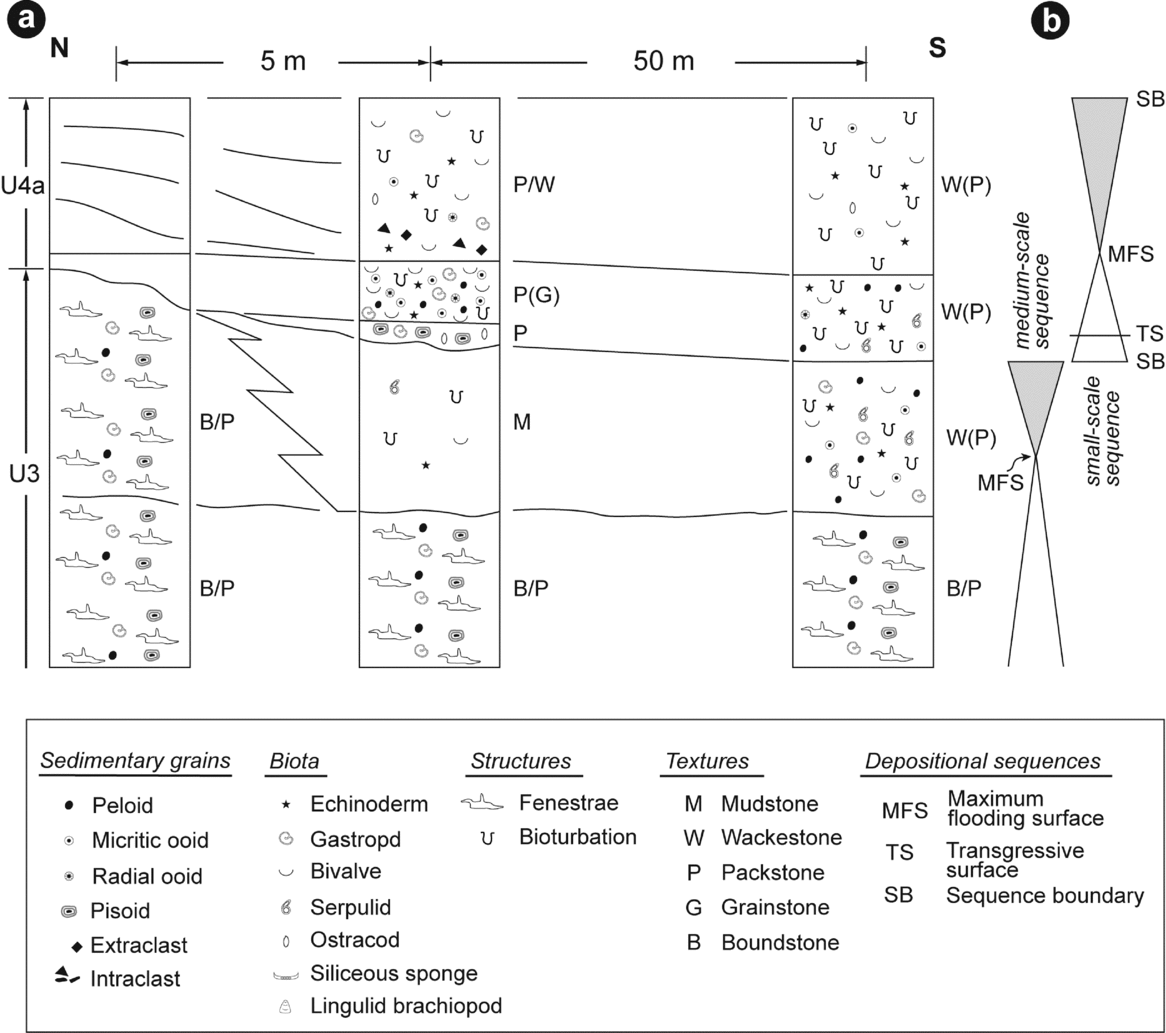
Lithological unit 3 displays the first N–S lateral facies variation between fenestral-microbial limestones and a dolomudstone (Figs 5a, 6a). The contemporaneity of these two facies belts is marked by the lateral continuity of some bed surfaces (Fig. 2c). The dolomudstone facies records a sparse fauna consisting of serpulids, bivalves and echinoderm plates (Fig. 5b). Quartz grains are relatively rare. The uppermost part of lithological unit 3 made of fenestral-microbial limestones is marked by a truncated surface. Laterally, the dolomudstone facies is also truncated and evolves in its uppermost part toward a subfacies that becomes rich in pisoids (Fig. 5c). A third facies is observed in lithological unit 3 in the southernmost part of the studied cliff and laterally to the dolomudstone facies. It corresponds to a bioturbated limestone with a wackestone (locally packstone) texture (Fig. 6). Associated with peloids and rare micritic ooids, the fauna is represented by gastropods, serpulids, bivalves and echinoderm plates. Quartz grains are present to common, rounded and moderately to poorly sorted.

Figure 5. Field view illustrating (a) lateral facies variation (dashed line) between fenestral-microbial limestones and (b) dolomudstones in lithological unit 3 (U3); white arrow indicates angular extraclasts. (c) Erosional surface at the top of lithological unit 3; note ostracods just above this surface. (d) Cracked pisoids and gastropods. (e) Peloidal and bioclastic packstone (lower right corner) and a burrow infilled by a grainstone with peloids and ooids. (f) Bioclastic wackestone to packstone with gastropods, bivalves and echinoderms. U4 – lithological unit 4; SB – sequence boundary; TS – transgressive surface; MFS – maximum flooding surface.
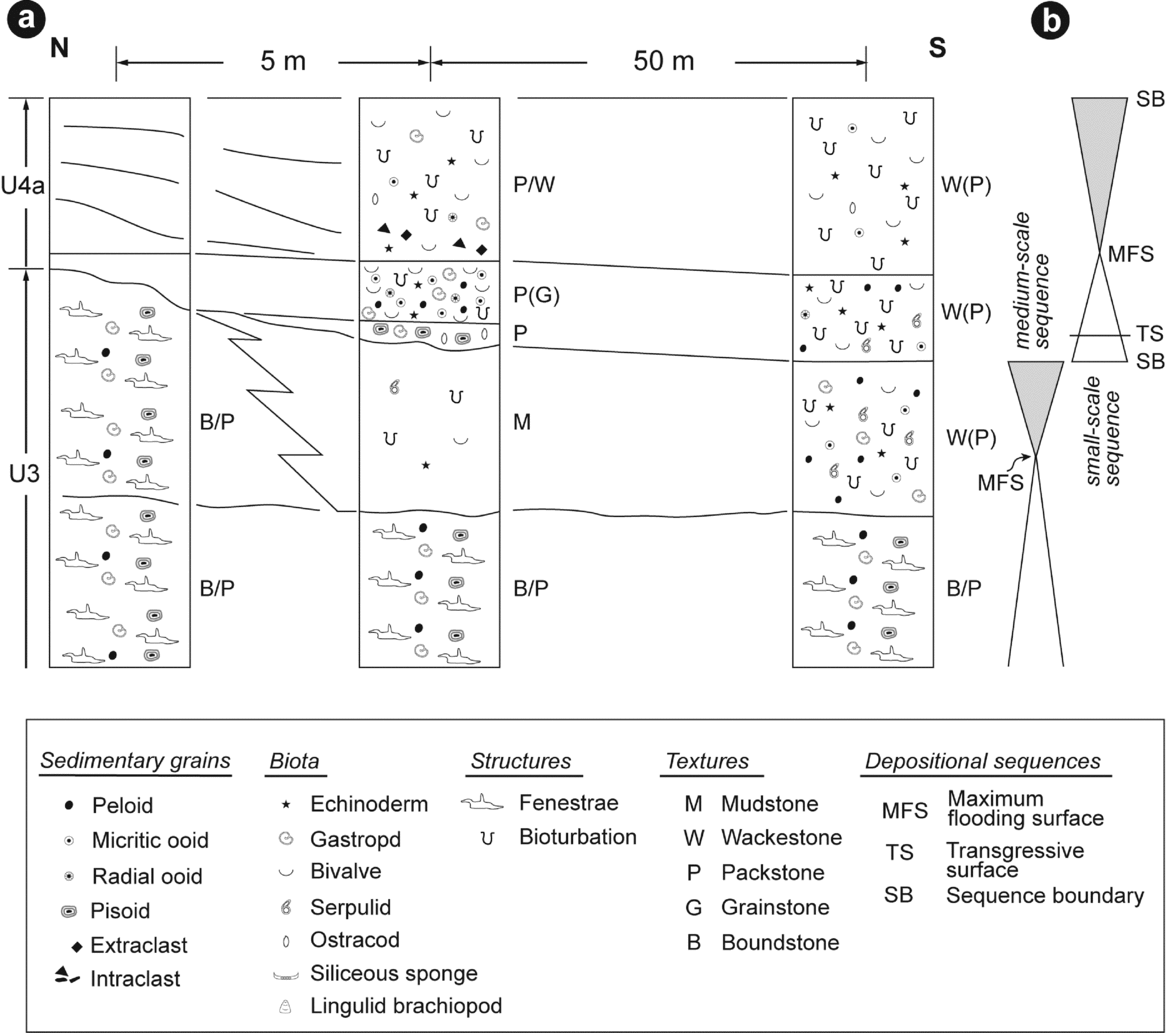
Figure 6. (a) Vertical and lateral facies variations at transition between lithological units 3 and 4. Not to scale. (b) Sequential interpretation.
3.d. Lithological unit 4
Lithological unit 4 is subdivided into two comparable subunits (4a and 4b), both of which display vertical and lateral facies changes (Fig. 6). In their northernmost part, these two subunits onlap on the truncated surface observed at the top of lithological unit 3, comprising the fenestral-microbial facies (Fig. 2). A few tens of metres south, the basal part of these two subunits displays centimetre-thick beds characterized by basal erosive surfaces (Fig. 5a, c). These thin beds consist of a grain-supported sediment particularly rich in large pisoids with some gastropods and ostracods (Figs 5c, d, 6a). Quartz grains are present to common, poorly to moderately sorted and subangular to subrounded. Several tens of metres south, the base of subunit 4b consists of sediments that have a mud-supported texture with only sparse fragments of echinoderms and bivalves (Figs 6a, 7a, i). Quartz grains are present to common, moderately sorted and well rounded, associated with some subangular centimetre-scale cherty clasts.

Figure 7. (a) Field and (b–i) microfacies illustrations of lithological units 4b and 5. (a) Lingulid brachiopods (white star) are present in subunit 4b, and small rounded siliceous sponges (black star) are present at the base of lithological unit 5. (b) Grainstone (locally packstone) consisting of abundant recrystallized bivalves, some echinoderms, gastropods, peloids and detrital quartz. (c) Dolomitized wackestone rich in quartz, with sparse echinoderm fragments. Dotted line in lithological unit 5 separates the two main subunits illustrated in (b) and (c). (d) Wackestone (locally packstone) composed of abundant echinoderms, bivalves and rare serpulids; quartz is abundant. (e) Packstone (locally wackestone) consisting of echinoderms, bivalves, gastropods, peloids and ooids; quartz is present. Note the shrinked structure of micritic ooids. (f) Wackestone composed of abundant echinoderms, bivalves, gastropods and rare serpulids; quartz grains are relatively large and abundant. (g) Packstone consisting of abundant peloids, echinoderms, gastropods, bivalves, serpulids and radial (and micritic) ooids; quartz is abundant. (h) Wackestone composed of echinoderms and bivalves; quartz is common. (i) Dolomitized mudstone-wackestone with sparse fragments of echinoderms and bivalves. Legend as in Figure 6.
Subunits 4a and 4b display a lower part characterized by one (subunit 4a) or several (subunit 4b) beds onlapping on the truncated surface at the top of lithological unit 3, comprising fenestral-microbial limestone facies (Fig. 2d, e). The onlapping bed of subunit 4a also shows lateral facies variations (Fig. 6). In a northern position, close to its onlap termination, the sediment displays a packstone (locally grainstone) texture and contains peloids, ooids (micritic and, to a lower extent, radial), undifferentiated bivalves, gastropods and echinoderm plates (Fig. 5e). Although preserved as incomplete internal moulds, numerous gastropods reach an unusually large size up to an estimated maximum of c. 9 cm (measured size c. 7 cm; Fig. 8). An ammonoid specimen (Owenites sp.) was observed associated with these gastropods. Beyond 50 m from the previous sampling zone, the sediment has a wackestone (locally packstone) texture with peloids, bivalves, serpulids, echinoderm plates and sparse micritic and radial ooids. In this lower part of subunit 4a, quartz grains are abundant, subrounded to subangular and moderately sorted.

Figure 8. (a) Field illustration of Smithian large-sized gastropod specimens from Timpoweap Canyon, lithological unit 4a. (b) Close-up of a single specimen. Dashed lines indicate shell-coiling axis. Specimen number: Université de Bourgogne Géologie Dijon, UBGD 275164.
The upper parts of subunits 4a and 4b display prograding sigmoids with downlap and toplap terminations (Fig. 2d, e). Sampled at the toe of the clinoform (sensu Quiquerez & Dromart, Reference Quiquerez and Dromart2006), the sediments exhibit a packstone-wackestone texture with bivalves, gastropods, echinoderms, ostracods and sparse radial and micritic ooids (Fig. 5f). Quartz grains are common, subrounded and moderately sorted. Subangular pebbles of cherty extraclasts are present at the base of the clinobeds (Fig. 5a). Several tens of metres south of the toe of the clinoform, subunit 4b can be subdivided into two main beds that can be regarded as distal parts of a clinobed (Fig. 7a). These distal clinobeds show a net vertical facies evolution. In their lower part, sediments display a mud-supported texture with echinoderm plates, bivalves, ostracods and some lingulid brachiopods (Fig. 7a, h, f). Quartz grains are common to abundant, subangular to subrounded and relatively well sorted. The uppermost part of the beds shows a grain-supported texture with gastropods, bivalves, echinoderm plates, ostracods, peloids and micritic and radial ooids (Fig. 7a, e, g). Quartz grains are abundant, subangular to subrounded and moderately sorted.
A few metres south of the toe of the clinoforms, both subunits are characterized by a dense and deep bioturbation marked by overabundant vertical trace fossils (Fig. 9). The entire bed thickness displays vertical or slightly oblique burrows that are circular in cross-section (Fig. 9a). Burrow diameters are between 0.5 and 2 cm. Burrow depth can reach up to 25 cm. Even if not always well preserved, these burrows show a U-shape apparently with no spreiten in between (Fig. 9b). Although a firm ichnogeneric assignment is difficult on the studied outcrop, these vertical burrows probably belong to the ichnogenus Arenicolites (R. Hofmann, pers. comm. Reference Hofmann, Hautmann, Wasmer and Bucher2013). Occasionally, some horizontal burrows (apparently not branching) are observed in these beds. Since outcrop conditions did not allow us to investigate their lining and sedimentary infill, these burrows belong either to the ichnogenus Palaeophycus or to Planolites (R. Hofmann, pers. comm. Reference Hofmann, Hautmann, Wasmer and Bucher2013).

Figure 9. Field observations of the intense bioturbation in lithological unit 4. (a) Basal and lower parts of subunit 4a; burrows are present in the entire bed thickness. Note the minor surfaces inside the bed that mark the successive phases of colonization by bottom dwellers. (b) Lower part of subunit 4b; note that burrows are up to 25 cm in depth and display a basal U-bend from pair association (white arrows, hammer for scale).
3.e. Lithological unit 5
Lithological unit 5 exhibits a vertical facies evolution (Fig. 7). Sediments of the thin (c. 30 cm thick) basal part are represented by a packstone (locally wackestone) texture with abundant and coarse fragments of echinoderms, bivalves and serpulids (Fig. 7a, d). This basal interval is strongly bioturbated and contains abundant, moderately well sorted and subrounded quartz grains. Notably, it also contains small isolated spheroid siliceous sponges that do not exceed c. 1 cm in diameter. These isolated sponges resemble lyssacine hexactinellids previously described from ramp environments in the Smithian of central Utah and Spathian of Nevada (Brayard et al. Reference Brayard, Vennin, Olivier, Bylund, Jenks, Stephen, Bucher, Hofmann, Goudemand and Escarguel2011 b). The lower part of lithological unit 5 consists of a thick interval of wackestone with peloids and sparse echinoderm plates. Quartz grains are very abundant, well sorted and subrounded. This interval is still densely bioturbated and shows low-angle cross-stratification. The upper part of lithological unit 5 consists of a bioturbated and bioclastic grainstone (locally packstone) with abundant bivalves, gastropods and peloids. Some fenestrae are visible within peloidal horizons. Quartz grains are common to abundant, well sorted and subangular. This facies is characterized by angular cross-stratifications and herringbone cross-bedding structures with high amalgamation.
3.f. Lithological unit 6
Lithological unit 6 corresponds to a metre-scale interval that generally forms the top of the studied cliff (Fig. 2). Along a N–S transect, this unit shows a marked lateral facies variation (Fig. 10). In the northernmost part of the outcrop, sediments display a wackestone (locally packstone) texture with common bivalves and gastropods, and some serpulids. There are also common micritic ooids, some cortoids and possible Favreina (Fig. 10b). Quartz grains are not observed. A second facies type rests vertically directly on the discontinuity observed at the top of the thickest interval of lithological unit 3, which constitutes a topographic high. This facies displays a grainstone texture with abundant recrystallized bivalves, gastropods and rare ostracods (Fig. 10c). Abundant peloids and micritic ooids are also present. No detrital quartz was observed. South of the topographic high, the sediments exhibit a packstone (locally wackestone) texture with abundant bivalves and gastropods (Fig. 10d). Echinoderm plates and serpulids represent additional fauna. Micritic ooids and peloids are also common. Quartz grains are present, rounded and well sorted. In the southernmost part of the study area, the sediment displays a packstone texture with abundant thin-shelled bivalves, some serpulids and echinoderm plates. These bivalves can be vertically oriented or show umbrella structures. In this later case, the valves shield underlying siliceous sponges (Fig. 10e). Quartz grains are rare and rounded.

Figure 10. (a) Depositional model for lithological unit 6. (b–e) Microfacies illustrations along a N–S transect in Timpoweap Canyon as illustrated in (f). (b) Wackestone with common bivalves and gastropods, some serpulids, common micritic ooids, some cortoids and possible Favreina. (c) Grainstone with abundant recrystallized bivalves, gastropods, rare ostracods, abundant peloids and micritic ooids. (d) Packstone with abundant bivalves and gastropods, common micritic ooids and peloids. (e) Packstone with abundant thin bivalves. Note that these bivalves show umbrella structures and that the pelecypod valves shielded underlying siliceous sponges. FWWB – fair-weather wave base. Legend as in Figure 6.
3.g. Lithological unit 7
Lithological unit 7 sets back from the cliff and can be observed only along local low-relief outcrop surfaces (Fig. 2a). Its facies corresponds to a bioclastic grainstone (locally packstone) with abundant bivalves, some ooids and peloids and possibly some Favreina. Locally, some fenestrae, mud clasts and mud draps are also observed. Quartz grains are common to abundant, well sorted and subrounded.
4. Depositional models
The progressive gradation between deposits of lithological units 2 to 3 is consistent with a first single depositional model (Fig. 11a). Previous studies interpreted the Rock Canyon Conglomerate (i.e. lithological unit 2) as palaeochannel deposits and thin regolithic breccias (Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). Nielson (Reference Nielson1991) considered that coarse to fine cross-bedded clastic rocks suggest deposition as a fan delta. Our observations reveal a lower breccia subunit with sandy matrix-supported clasts that lack imbrication and internal stratification (Fig. 3a), pointing to debris flows in an alluvial context. Even if a marked Permian palaeorelief was not observed in the studied area, the palaeogeographical reconstruction of Blakey (Reference Blakey1979) indicates a chert-pebble alluvial plain west of Hurricane at the front of low hills consisting of cherty Kaibab Formation in the area of the present Virgin Anticline. The second subunit of lithological unit 2 reveals conglomerates with planar cross-bedded gravels (Fig. 3b), which can reflect lateral out-building from longitudinal bars. This conglomeratic subunit is overlain by silty to muddy deposits of a third subunit (Fig. 3c). These later deposits suggest migration of active to inactive tracts.
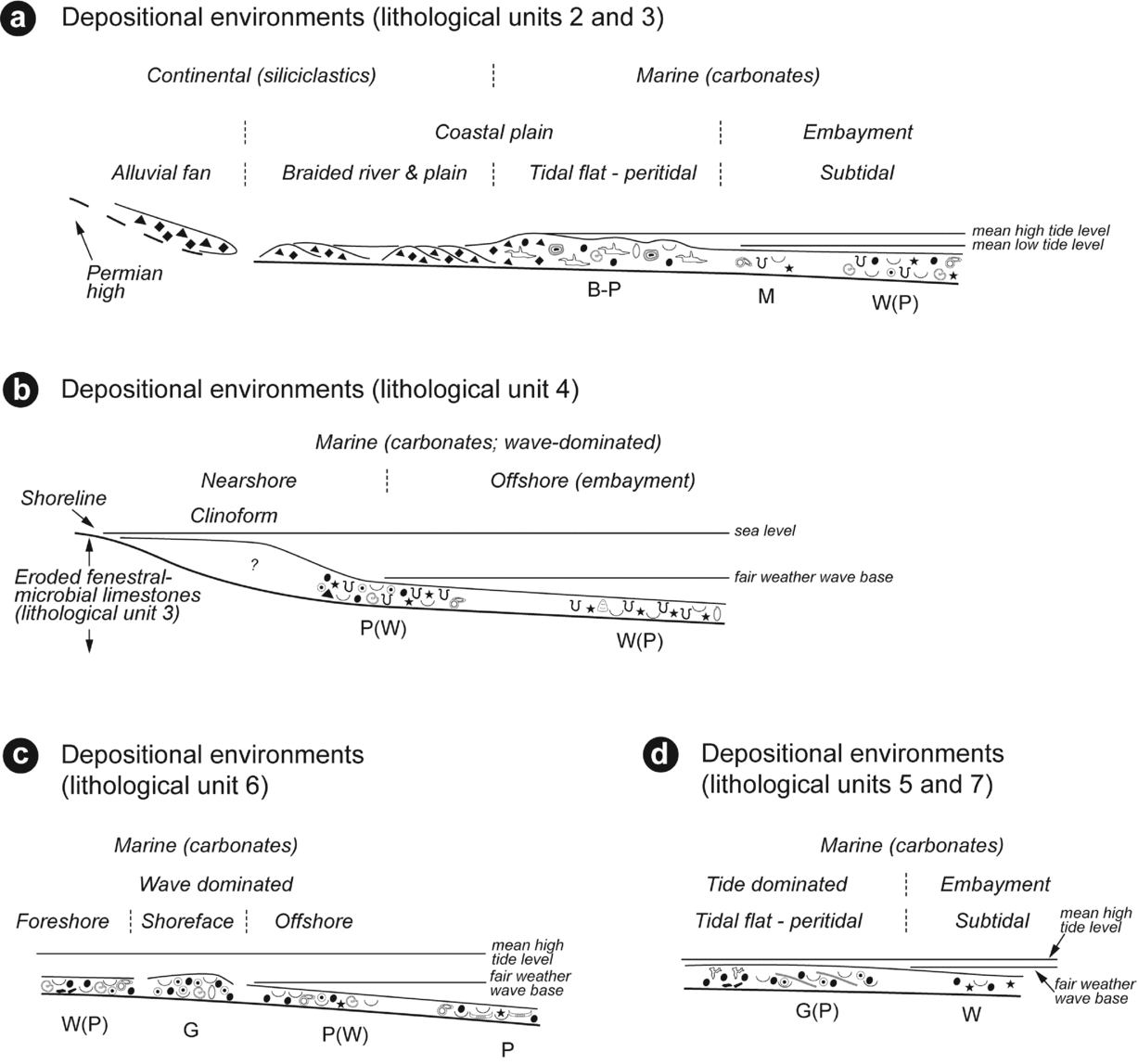
Figure 11. Depositional models deduced from the Timpoweap sedimentary succession. Depositional environments for: (a) lithological units 2 and 3; (b) lithological subunits 4a and 4b; (c) lithological unit 6; and (d) lithological units 5–7. Legend as in Figure 6.
The succession of the three subunits observed in lithological unit 2 (i.e. breccia, conglomerates and silts) therefore reflects an evolution from an alluvial fan to more distal rivers and flood plains. The transition to a tidal flat is progressively recorded at the base of lithological unit 3 where fenestral-microbial carbonates still contain abundant cherty clasts (limestone conglomerate of Blakey, Reference Blakey1979; Fig. 3d). Lithological unit 3 is characterized by a lateral facies zonation from peritidal to shallow subtidal settings (Fig. 11a). Peritidal carbonates (sensu Flügel, Reference Flügel2004), that is, shallow-subtidal, intertidal and supratidal sediments formed in marginal-marine and shoreline depositional environments, correspond to the fenestral-microbial facies. The association of stromatolites, fenestral pores and in situ brecciation are consistent with frequently exposed parts of tidal flats (Flügel, Reference Flügel2004). The observed fauna (i.e. sparse ostracods, gastropods and bivalves) is also consistent with an intertidal environment (Flügel, Reference Flügel2004). Pisoids and meniscus cement suggest vadose-marine environments in supratidal areas (Esteban and Pray, Reference Esteban, Pray and Peryt1983; Peryt, Reference Peryt and Peryt1983). Lateral accretion and minor truncation surfaces observed in low-relief patch-reef structures indicate reduced accommodation and recurrent erosional phases (Figs 2, 4a, b). Bioturbated dolomudstones with rare organisms (i.e. serpulids, bivalves and echinoderms) observed laterally to the fenestral-microbial facies (Figs 5, 6) reflect a more important available space for sedimentation in a shallow subtidal marine setting (Fig. 11a). At a more distal position (a few tens of metres to the south), the sediments become more bioclastics with gastropods, serpulids, bivalves and echinoderms. Even though few ooids are observed, the mud-supported texture still implies a low-energy setting protected from the open ocean. Such protection can result from the existence of a wide epeiric platform dampening incoming waves, positioned behind a barrier or within a restricted embayment. The presence of equivalent deposits (i.e. Sinbad and Timpoweap formations) between the eastern and western parts of southern Utah (Blakey, Reference Blakey1979; Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007) suggests an embayment that opened to the north to a larger epicontinental basin (Brayard et al. Reference Brayard, Bylund, Jenks, Stephen, Olivier, Escarguel, Fara and Vennin2013). This first facies model deduced from lithological units 2 and 3 highlights a fluvial-marine transitional environment from a coastal plain (see also Blakey, Reference Blakey1979) to an embayment system.
The analysis of the two subunits of lithological unit 4 indicates a significant change in topography that strongly modified the depositional settings (Fig. 11b). Attached to an antecedent topography that resulted from truncation of the fenestral-microbial limestones (lithological unit 3), subunits 4a and 4b can be assimilated to clinothems that are made of clinobeds bound by hiatal surfaces (Fig. 2d, e). If the flat-topped upper and middle slope parts of the clinoforms could not be sampled, the toe-of-slope part of the first clinothem (i.e. subunit 4a) consists locally of grain-supported sediments (Figs 5f, 6a). These latter sediments are also strongly bioturbated and composed of bivalves, gastropods, peloids, ooids and echinoderms. Such facies suggests deposition in an open marine setting at the transition between nearshore and offshore settings (Fig. 11b). The presence of large limestone gravels in the lower part of subunit 4a indicates that erosion was still active at the top of the underlying fenestral-microbial limestones of lithological unit 3 (Fig. 5a). Sampled in a more distal position (Fig. 7), sediments of the second clinothem (i.e. subunit 4b) display a mud-supported fabric with bivalves, echinoderms, sparse ostracods and rare ooids. The observed ichnoassemblage, with a high density of putative Palaeophycus or Planolites, reflects an intense colonization by bottom dwellers between storm periods. Subunits 4a and 4b correspond to metre-scale clinoforms that are characterized by rapid lateral facies changes over a few tens of metres. Such a pattern suggests a shallow proximal setting above fair-weather wave base (Quiquerez & Dromart, Reference Quiquerez and Dromart2006). In these clinothems grain-supported sediments have a limited extension (i.e. a few tens of metres close to the shoreline position), suggesting a low-wave-energy embayment system.
Lithological unit 6 was sampled laterally (Fig. 10). Lateral facies distribution suggests that the inherited topography may have still influenced the sedimentation under a wave-dominated regime. Observed where lithological unit 3 is the thickest, the deposition of an oolitic grainstone suggests a shoreface setting (Fig. 11c). Protected behind these high-energy deposits, mud-supported sediments may have been deposited. Below the fair-weather wave base, offshore deposits correspond to a bivalve-rich packstone. Some of these bivalves shielded underlying siliceous sponges (Fig. 10e) whereas others are vertically oriented, suggesting local turbulent and rotary flows during storms (Pérez-López & Pérez-Valera, Reference Pérez-López and Pérez-Valera2011). This third depositional model suggests a marginal area where sedimentation was either tide or wave dominated.
Facies observed in lithological units 5 and 7 likely reflect more internal peritidal settings compared to deposits of lithological unit 6 (Fig. 11d). Angular cross-stratification, herringbone cross-bedding and local peloidal horizons observed in the upper part of lithological unit 5 indicate a tide-dominant regime. The presence of fenestrae and mud intraclasts may reflect more internal tidal flats. The lower part of lithological unit 5 consists of mud-supported bioclastic sediments rich in echinoderms and quartz (Fig. 7a, c), suggesting deposition in a marine offshore setting of a low-energy embayment system.
5. Transgressive-regressive cycles and shoreline migrations
Stratal stacking patterns and surface geometries observed in Timpoweap Canyon reveal at least three different orders of transgressive-regressive sequences (Fig. 1b). Above the Rock Canyon Conglomerate, the lithological succession can be replaced by a long-term transgressive trend. Inside this transgressive trend, lithological units 2 and 3 illustrate a complete first medium-scale sequence that ends at the erosional truncation surface (Fig. 2). Lithological units 4–7 record the transgressive interval of a second depositional sequence. The regressive interval of this sequence probably corresponds to deposits of the Lower Red Formation. This latter formation overlies the Sinbad Formation (Blakey, Reference Blakey1979; Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007). Four additional small-scale sequences are also well visible within lithological units 4–7 (Figs 1, 12).
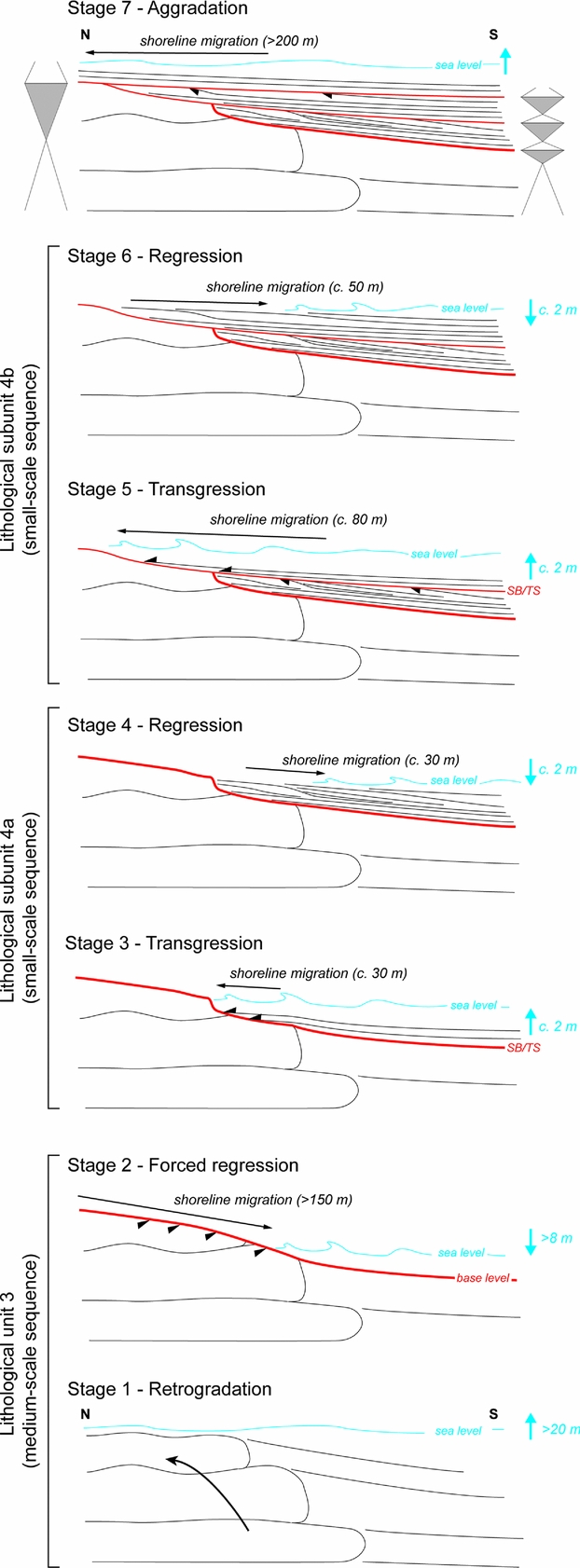
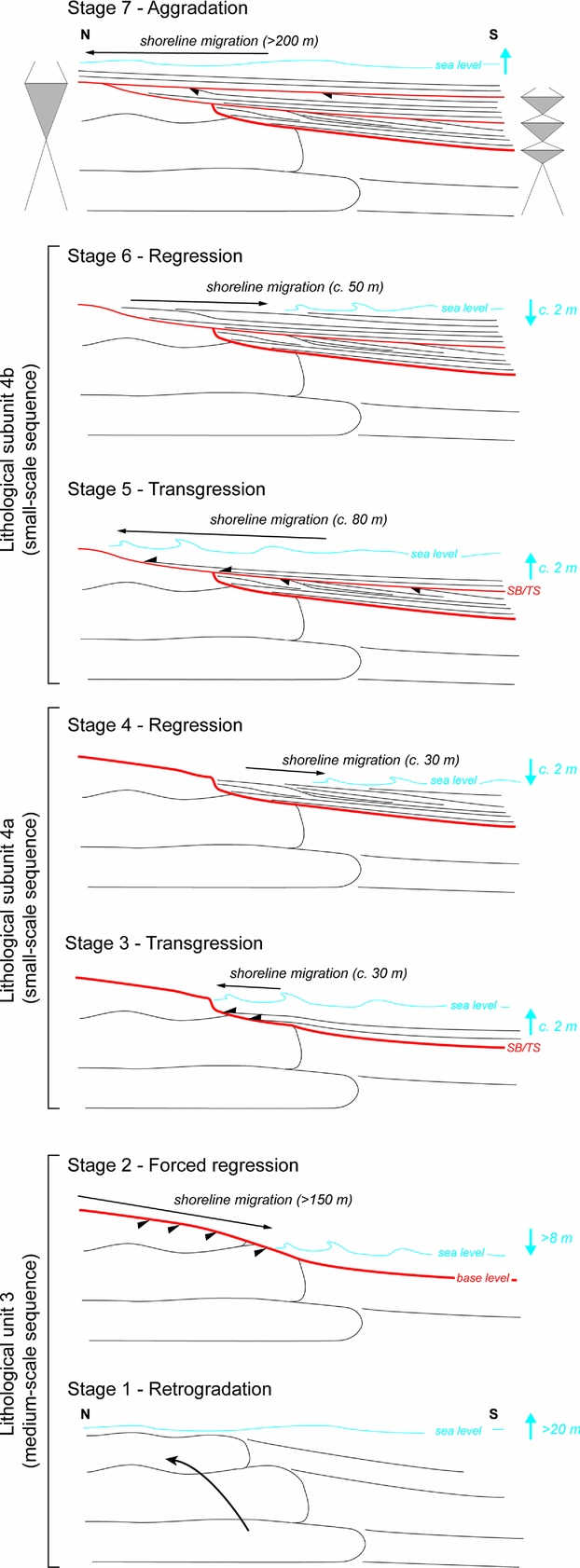
Figure 12. Reconstruction of Early Triassic medium-scale (lithological unit 3) and small-scale (lithological subunits 4a and 4b) sequences, and successive stage of shoreline migrations along the sedimentary succession observed in Timpoweap Canyon.
Deposition of fenestral-microbial limestones (i.e. lithological unit 3) reflects a progressive marine incursion on a chert pebble alluvial plain (Blakey, Reference Blakey1979). Transgression of the Early Triassic sea within central and southern Utah – probably during the early Smithian in this area (Brayard et al. Reference Brayard, Bylund, Jenks, Stephen, Olivier, Escarguel, Fara and Vennin2013) – resulted in deposition of the limestones of the Sinbad Formation (Nielson, Reference Nielson1991). This first long- and medium-scale transgressive interval is highlighted by a retrogradational stacking pattern of deposits in the lower part of lithological unit 3 (stage 1, Fig. 12). Microbial patchy morphologies have larger dimensions in the middle part of this unit, suggesting higher accommodation and thus an interval of maximum flooding for the first identified medium-scale transgressive-regressive depositional sequence (Fig. 1b). Stratal terminations in the uppermost part of fenestral-microbial unit 3 correspond to a truncation that implies a net accommodation loss associated with an erosional phase. Such an erosional surface reflects a forced regressive trend (stage 2, Fig. 12). The peritidal setting of fenestral-microbial deposits coupled with relative sea-level changes could have preferentially caused an emersion and erosion up to the formation of a local palaeorelief that gradually dipped to the south (Fig. 2a, b). The shoreline migration probably exceeded 150 m southwards, that is, the lateral extent of the outcrop where truncation is visible at the top of lithological unit 3. Lateral to this fenestral-microbial facies, dolomudstones are progressively enriched in pisoids and finally exhibit an uneven erosional surface in their stratigraphic uppermost part. As toplap terminations are no longer observed, this erosional surface probably reflects wave scouring in a shallow marine setting during the later stages of base level fall. This erosional surface is interpreted as the sequence boundary of the first identified medium-scale transgressive-regressive sequence (Figs 1b, 5a, c, 6).
A thin double bed formed of pisoids with abundant shrinkage structures overlies this surface (Fig. 5d, e). These pisoids were probably removed from subaerial unconformity at the top of lithological unit 3 and redeposited in a shallow subtidal setting. Considering a negligible compaction for deposits of lithological unit 3 – absence of stylolites coupled with a rapid lithification of microbial deposits – this forced regressive trend resulted in a sea-level fall of more than 8 m. This estimate corresponds to the thickness of fenestral-microbial limestone unit 3 that exhibits the truncation.
A first small-scale transgressive-regressive sequence is recorded within lithological subunit 4a. The transgressive interval corresponds to successive beds that onlapped against the truncated surface at the top of lithological unit 3 (Figs 2d, 5a, c). A putative berm crest may reflect the shoreline position at the end of the transgression (Fig. 2d). In that case, the shoreline migration accompanying this transgressive trend can be estimated to be c. 30 m northwards (stage 3, Fig. 12). The corresponding relative sea-level rise is estimated to be c. 2 m. Successive clinoforms mark a normal regressive trend of this first small-scale sequence (stage 4, Fig. 12). The latter erosional transgressive surface makes it difficult to estimate the position of the shoreline at the end of the regression. Shoreline position should not exceed the more distal truncated clinobed, which probably reflects wave scouring in a shallow marine setting during the later stages of a base level fall. In that case, the shift in shoreline during this regressive trend was probably close to 30 m southwards, and the corresponding sea-level fall was close to 2 m.
The second small-scale sequence corresponds to subunit 4b. The basal part of this subunit displays successive beds that onlapped against the erosional transgressive surface at the top of lithological unit 4a (stage 5, Fig. 12). A new putative berm crest marks the shoreline position at the end of the transgression (Fig. 2e). The shoreline position shifted about 80 m northwards and the corresponding sea-level rise did not exceed c. 2 m. Similar to the previous small-scale sequence, successive clinoforms indicate a regression. The southernmost eroded clinobed observed in the uppermost part of subunit 4b suggests a shoreline migration of <50 m and a sea-level fall of <2 m (stage 6, Fig. 12).
With vertical stacking patterns, lithological units 5–7 do not allow us to further reconstruct shoreline trajectories (Fig. 2). However, this sedimentary succession suggests two additional small-scale sequences of relative sea-level changes. The first (i.e. third small-scale sequence; Fig. 1b) is recorded in lithological unit 5. At the base of the unit, a multi-centimetre-thick interval of limestones, rich in coarse echinoderm fragments and sparse siliceous sponges, was deposited during a relative sea-level rise (Fig. 7a). The corresponding transgressive trend was marked by a shoreline migration that exceeded 200 m – i.e. the lateral extent of the outcrop where lithological unit 5 is visible over lithological unit 3 (stage 7, Fig. 12). A thin marly horizon probably reflects the maximum flooding interval. A dolomicrite limestone with sparse echinoderms and overlying tidal bivalve-rich grainstones may indicate a relative sea-level fall (Figs 7a, 11d). The last small-scale sequence includes deposits of lithological units 6 and 7. Wave-dominated bivalve-rich limestones of lithological unit 6 were deposited during a relative sea-level rise, whereas deposition of tide-dominated bivalve-rich limestones reflects a relative sea-level fall.
The erosional phase leading to truncation at the top of lithological unit 3 triggered an increment of accommodation over a few tens of metres along a N–S proximal-distal axis (Fig. 2). Two small-scale sequences with successive clinoforms are observed at a southern (i.e. distal) position, whereas these are absent from a proximal position (Fig. 12). During the transgressive trend at the base of lithological unit 5 the antecedent relief was largely filled by deposits of lithological unit 4, leading to a shoreline migration exceeding several hundreds of metres (stage 7, Fig. 12). Sampling difficulties did not allow us to highlight the influence of palaeotopography during deposition of lithological unit 5. Nevertheless, inherited topography still influenced the sedimentation of lithological unit 6, as suggested by deposition of oolitic grainstones on a residual topographic high (Fig. 10). The Timpoweap sedimentary succession therefore reflects peritidal to shallow subtidal settings, which were adequate to record rapid shoreline migrations. In such a context topographic highs of restricted local extent can be formed, leading to the recording of a different number of depositional sequences according to the position along the coastline (Fig. 12). A complex and irregular coastline is also suggested during the Smithian when comparing the northwards polarity of the observed depositional system with the global trend of the Early Triassic transgression at a regional scale. Indeed, the Smithian advancement of the sea seems to have proceeded southwards along a corridor within Utah (Collinson, Kendal & Marcantel, Reference Collinson, Kendall and Marcantel1976; Nielson, Reference Nielson1991; Paull & Paull, Reference Paull and Paull1993; Dickinson, Reference Dickinson2006; Brayard et al. Reference Brayard, Bylund, Jenks, Stephen, Olivier, Escarguel, Fara and Vennin2013). Moreover, the enlarged palaeogeographical reconstruction of the studied area by Blakey (Reference Blakey1979) indicates another general polarity of the depositional system, with topographical highs located on the west and an open sea on the east. The observed sedimentary succession in Timpoweap Canyon suggests that, at least locally, the continental to marine open depositional system can display a N–S polarity.
6. Relative sea-level changes
Global sea-level curves generally rise across the Permian–Triassic boundary (Haq & Al-Qahtani, Reference Haq and Al-Qahtani2005; Haq & Shutter, Reference Haq and Shutter2008; ICS, unpub. data, 2011: http://www.stratigraphy.org). In this global trend, Early Triassic sea-level changes of higher frequency are not easily dated and correlated, which explains why they are rarely considered (Embry, Reference Embry1997). Lehrmann et al. (Reference Lehrmann, Payne, Pei, Enos, Druke, Steffen, Zhang, Wei, Orchard and Ellwood2007) identify several third-order sea-level sequences in the Beisi Formation of the Chongzuo–Pingguo platform (South China). In SW Utah, the Lower Triassic sedimentary succession recorded three third-order transgressive-regressive sequences that originated in the Griesbachian, Smithian and earliest middle Spathian (Embry, Reference Embry1997; Paull & Paull, Reference Paull, Paull, Dickins, Zunyi, Hongfu, Lucas and Acharyya1997). Deposition of the Rock Canyon Conglomerate could have followed the tectonic uplift linked to the Sonoma Orogeny (Collinson, Kendal & Marcantel, Reference Collinson, Kendall and Marcantel1976; Nielson, Reference Nielson1991; Dickinson, Reference Dickinson2006). The lack of biostratigraphic markers makes it difficult to assign a more precise age, which was dated as either Permian or Early Triassic (Gregory, Reference Gregory1950; Reeside & Bassler, Reference Reeside and Bassler1922; Blakey, Reference Blakey1979). In Timpoweap Canyon, the gradual transition between continental and marine deposits corresponds to the transition between the Rock Canyon Conglomerate and limestones of the Sinbad Formation that was considered as middle–late Smithian in age (Brayard et al. Reference Brayard, Bylund, Jenks, Stephen, Olivier, Escarguel, Fara and Vennin2013). The presence of Owenites sp. in the deposits just above the fenestral-microbial lithological unit 3 allows us to specify a middle Smithian age for the Sinbad Formation. This sedimentary evolution can be placed into the third-order Smithian sea-level rise identified by Embry (Reference Embry1997).
The identification of medium- and small-scale depositional sequences in the Sinbad Formation reflects high-frequency relative sea-level fluctuations. Small-scale depositional sequences observed in lithological units 4–7 are of middle–late Smithian age, representing a total maximum duration of <400 ka (see estimates by Brühwiler et al. Reference Brühwiler, Bucher, Brayard and Goudemand2010). If high-frequency (i.e. Milankovitch and sub-Milankovitch) cycles appear to have been recorded in Lower Triassic sedimentary successions (e.g. Huang et al. Reference Huang, Tong, Hinnov and Chen2011; Wu et al. Reference Wu, Zhang, Feng, Jiang, Li and Yang2012), corresponding sea-level changes were only suspected (Lucas, Krainer & Milner, Reference Lucas, Krainer, Milner, Lucas and Spielmann2007; Chen, Fraiser & Bolton, Reference Chen, Fraiser and Bolton2012). Observed small-scale depositional sequences are asymmetrical with thin transgressive deposits and thicker regressive deposits. Such an asymmetrical pattern of sequences is typical of short-term sea-level variations that are induced by glaciation–deglaciation cycles (e.g. Shackleton, Reference Shackleton1987; Ruddiman, Reference Ruddiman2003; Ridente et al. Reference Ridente, Petrungaro, Falese and Chiocci2012). However, the amplitude of sea-level fluctuations indicated by the small-scale depositional sequences of lithological units 4–7 does not exceed c. 2 m. Such low-amplitude sea-level change is consistent with the absence of evidence for continental glaciation during Early Triassic time (Frakes, Francis & Syktus, Reference Frakes, Francis and Syktus1992; Sano et al. Reference Sano, Onoue, Orchard and Martini2012). In that case, these high-frequency sea-level changes may be interpreted to be the result of intense thermal expansions of the world ocean triggered by global warming (e.g. Kidder & Worsley, Reference Kidder and Worsley2004). Particularly warm seawater temperatures during the middle–late Smithian (cf. Romano et al. Reference Romano, Goudemand, Vennemann, Ware, Schneebeli-Hermann, Hochuli, Brühwiler, Brinkmann and Bucher2013) support such an explanation. Alternatively, a sudden drop in sea level of at least 8 m triggered the formation of a palaeorelief at the top of lithological unit 3. Such a sea-level drop could have a tectonic origin related to the Sonoma Orogeny, which is responsible for regional uplift during the transition from Permian to Triassic (Collinson, Kendal & Marcantel, Reference Collinson, Kendall and Marcantel1976). Thus, large- and medium-scale depositional sequences identified in the Timpoweap Canyon succession are consistent with regional uplifts, whereas small-scale depositional sequences may have been climatically driven.
7. Depositional settings and regional biotic recovery
Within Timpoweap Canyon, the oldest Triassic fossils are documented from lithological unit 3. This unit shows important lateral facies variations with fenestral-microbial limestones in peritidal settings and dolomudstones and bioclastic limestones in shallow marine subtidal settings (Fig. 11a). In the fenestral-microbial limestones, benthic organisms (gastropods, bivalves, ostracods and rare putative siliceous sponges) are poorly diversified. Microbial deposits are responsible for the edification of small patches that do not exceed a few metres in thickness (Figs 2, 4). Associated with rare putative siliceous sponges, these patches do not have a complex framework made of skeletal metazoans as was observed in other Smithian or Spathian reefs from Utah (Brayard et al. Reference Brayard, Vennin, Olivier, Bylund, Jenks, Stephen, Bucher, Hofmann, Goudemand and Escarguel2011 b; Marenco et al. Reference Marenco, Griffin, Fraiser and Clapham2012). Patches observed in Timpoweap Canyon therefore cannot be considered as true metazoan-microbial reefs. This also suggests that Lower Triassic microbial-dominated limestone units (i.e. basal limestones of the Sinbad Formation) can show important differences in terms of biotic associations and frameworks at the scale of the western USA basin.
The benthic assemblage observed here in association with microbial deposits is well known worldwide in the aftermath of the end-Permian mass extinction (Sano & Nakashima, Reference Sano and Nakashima1997; Kershaw et al. Reference Kershaw, Li, Crasquin-Soleau, Feng, Mu, Collin, Reynolds and Guo2007, Reference Kidder and Worsley2012). These metazoans are classically interpreted as representing low-diversity communities whose structure reflects harsh environmental conditions (Yang et al. Reference Yang, Chen, Wang, Tong, Song and Chen2011; Forel et al. Reference Forel, Crasquin, Kershaw and Collin2013). They are typically composed of opportunistic or disaster forms dwelling in microbial ecosystems. However, in Timpoweap Canyon, the peritidal fenestral-microbial limestones are observed laterally to shallow bioclastic and bioturbated limestones with echinoderms, gastropods, serpulids and bivalves. Such a relatively diversified fauna suggests subtidal and marine conditions. It also seems to indicate that the biotic recovery was well underway contemporaneously with deposition of the fenestral-microbial limestones, at least locally. This observation questions the putative anachronistic and disaster character commonly attributed to microbialites in the western USA basin (Schubert & Bottjer, Reference Schubert and Bottjer1992; Pruss & Bottjer Reference Pruss and Bottjer2004; Pruss et al. Reference Pruss, Bottjer, Corsetti and Baud2006; Baud, Richoz & Pruss, Reference Baud, Richoz and Pruss2007; Woods, Reference Woods2009, Reference Woods2013). Another example of a massive microbial mound that laterally passes to bioclastic packstones deposited in normal marine conditions was recently illustrated in the upper Anisian of Iran (Berra et al. Reference Berra, Balini, Levera, Nicora and Salamati2012). Thus, the presence of microbial deposits in western USA does not necessarily indicate widespread harsh marine conditions. Rather, it underlines the preponderant role of the depositional environment upon the facies and biota distribution along a complex regional Early Triassic coastline.
In lithological unit 4, depositional settings evolved to a wave-dominated ramp system implanted on an inherited topography (Fig. 11b). In this facies model, microbial deposits are no longer observed. The absence of a large peritidal area coupled with relative high-energy conditions in an inner ramp setting probably did not allow the installation and stabilization of biofilms. The faunal assemblage – composed of echinoderms, bivalves, gastropods, serpulids and ostracods – did not significantly change during the deposition of unit 4, with the exception of bioturbation and some gastropod accumulations. The occurrence of large-sized gastropods at the base of subunit 4a suggests unstressed marine conditions. Interestingly, it reinforces the previous assertion that a putative Lilliput effect on gastropods was non-effective or overestimated, at least during the Smithian (Brayard et al. Reference Brayard, Nützel, Stephen, Bylund, Jenks and Bucher2010, Reference Brayard, Nützel, Kaim, Escarguel, Hautmann, Stephen, Bylund, Jenks and Bucher2011 a). Bioturbation seems more intense in subunits 4a and 4b compared to lithological unit 3. However, the quality of the outcrop did not allow us to adequately quantify the intensity of bioturbation in this latter unit. Below the fair-weather wave base, deposits of lithological unit 4 display dense, deep and large burrows that affect the entire thickness of the beds (Fig. 9). Colonization by bottom dwellers occurred at several stratigraphic horizons in the same bed. This suggests that successive generations of colonization reflect primary population density and are not linked to an important and long phase of sediment starvation. Although this dense ichnoassemblage is apparently not diversified (Arenicolites and Palaeophycus or Planolites), it is rather unusual not only for the Smithian but also for the entire Early Triassic. It does not entirely fit within traditional models of benthic recovery after the Permian–Triassic mass extinction that assume a progressive reappearance of dense, deep and large burrows only in the Spathian (e.g. Twitchett, Reference Twitchett2006; but see Hofmann et al. Reference Hofmann, Goudemand, Wasmer, Bucher and Hautmann2011, Reference Hofmann, Hautmann, Wasmer and Bucher2013 for alternative models). Indeed, this type of ichnoassemblage within a subtidal setting is more in agreement with the concept of a shallow ‘habitable zone’ as proposed by Beatty, Zonneveld & Henderson (Reference Beatty, Zonneveld and Henderson2008). Overall, it may indicate that environmental conditions were suitable for life (e.g. no anoxia) and stable. It also suggests that nutrients were profusely available during the deposition of these bioturbated beds.
8. Conclusion
Stacking patterns and geometric surfaces observed at Timpoweap Canyon reveal three different orders of relative sea-level fluctuations and high-frequency shoreline migrations. During the transition from Permian to Triassic, regional tectonic uplift related to the Sonoma Orogeny led to the deposition of terrigenous sediments of the Rock Canyon Conglomerate Formation. The first marine incursion, related to third-order sea-level transgression, occurred during the middle Smithian with deposition of limestones of the Sinbad Formation. A truncated surface at the top of the fenestral-microbial Sinbad limestones reflects a sudden sea-level drop, suggesting a tectonic influence still active in the Early Triassic. Inherited topographic highs of restricted local extension led to a different number of depositional sequences being recorded, according to the position along the coastline. Such limited geographic extension of some depositional sequences may distort our knowledge of delayed recovery of complex benthic communities after the end-Permian extinction. These small-scale, possibly climatically driven, depositional sequences resulted from shoreline migrations in excess of a few tens of metres and from sea-level drop and rise at a scale of 1–2 m.
The reconstruction of depositional settings in the Early Triassic of Timpoweap Canyon reflects the evolution of coastline sedimentation. Overlying the Permian–Triassic transition, alluvial fans and braided plains represent continental environments. The observed lateral facies variations indicate a progressive evolution from continental deposits to fenestral-microbial limestones in peritidal domain, and then to shallow subtidal marine environments with bivalves, gastropods, serpulids and echinoderms. Such a lateral facies variation highlights the control of the depositional setting upon the distribution of organisms. It also questions the anachronistic character of Early Triassic microbialites in the western USA basin.
Acknowledgements
The Région Bourgogne and the CNRS INSU Interrvie jointly supported this study. This is also a contribution to the ANR-funded project AFTER (ANR-13-JS06-0001-01). R. Hofmann and N. Goudemand (Zurich) are thanked for their help in ichnofossil and conodont determinations, respectively. D. Stephen is grateful for the ongoing financial support of the College of Science & Health at Utah Valley University. The studied outcrop is located on US public land under the stewardship of the Bureau of Land Management (BLM) of the US Department of the Interior; their management and access to these lands is much appreciated. S. Lucas and an anonymous reviewer provided constructive suggestions that helped us to improve the manuscript.