Introduction
Certain U.S. vegetable growers and processors are working toward producing edamame [soybean, Glycine max (L.) Merr.] on a commercial scale to address consumer demand for domestically grown product. Initially, weed interference was identified as a major obstacle to domestic production. Some herbicides have since been registered for use on edamame (Williams and Nelson Reference Williams and Nelson2014), and the vegetable industry now has nascent weed management systems in the crop (Williams Reference Williams2015a). Further development of integrated weed management (IWM) systems are needed, particularly IWM systems that combat herbicide resistance by taking greater advantage of biological, cultural, and physical weed management tactics (Harker and O’Donovan Reference Harker and O’Donovan2013).
Several fall-seeded cover crops suppress weed density and biomass through biological (e.g., seed predation), chemical (e.g., allelopathy), and physical (e.g., light interception) mechanisms (Wortman et al. Reference Wortman, Francis, Bernards, Blankenship and Lindquist2013). Cover crop residues have been examined for their weed management contributions in no-till grain-type soybean for decades (Liebl et al. Reference Liebl, Simmons, Wax and Stoller1992; Moore et al. Reference Moore, Gillespie and Swanton1994). Dense residues from cover crops such as cereal rye (Secale cereale L.) reduce preplant weed emergence and biomass in excess of 90% (Forcella Reference Forcella2013). Even low levels of certain residues, including cereal rye and oilseed radish (Raphanus sativus L.) provide early-season benefits to weed management (Moore et al. Reference Moore, Gillespie and Swanton1994; Ryan et al. Reference Ryan, Mirsky, Mortensen, Teasdale and Curran2011). However, cover crops can also lower soybean stand and yield by reducing available soil moisture or interfering with seedling establishment (Davis Reference Davis2010; Forcella Reference Forcella2013). Moreover, dense cover crop residues can interfere with crop-planting equipment and reduce crop emergence (Forcella Reference Forcella2013).
Edamame are larger seeded than grain-type soybean and can exhibit low crop emergence (Duppong and Hatterman-Valenti Reference Duppong and Hatterman-Valenti2005; Sánchez et al. Reference Sánchez, Kelley and Butler2005). Regardless of weed-suppressive benefits, cover crop residues would not be a viable management tactic if they further compromised edamame emergence. Currently, the potential role of fall-seeded cover crops for weed management in edamame is unknown. Therefore, the first objective of this study was to determine the extent to which cover crop–residue management systems influence edamame emergence while selectively suppressing weed density and biomass. Because seed size of edamame is not only larger than grain-type soybean, but also more variable among cultivars, the second objective was to determine whether cultivars differed in their emergence in cover crop–residue management systems.
Materials and Methods
Experimental Design
A field experiment was conducted at the University of Illinois Vegetable Crop Farm near Urbana, IL (40.076623N, −88.239866W), for edamame growing seasons in 2014 to 2016. Different fields were used each year. The soil was a Flanagan silt loam (fine, smectitic, mesic Aquic Argiudolls) with an average organic matter content of 3.4% and an average pH of 5.9. The previous crop of each field was grain-type soybean.
The experiment was a split-plot randomized complete block design with three replications. Main plots were assigned one of six fall-seeded cover crop–residue management systems, hereafter simply called “cover crop treatments,” including a bare-soil control. Cover crops were planted with hand and drill planters in late summer in 2013 to 2015, after the previous crop was harvested (Table 1). Cover crop treatments were selected to create a gradient of residues at planting and included one radish treatment (due to winterkill, a low-residue treatment), two canola (Brassica napus L.) treatments (i.e., early-killed and late-killed canola; medium-residue treatments), and two cereal rye treatments (early-killed and late-killed cereal rye; high-residue treatments; Table 1). Different timings of a burndown application with glyphosate (Roundup WeatherMax®, 1,094 g ae ha−1, Monsanto, St Louis, MO) were used to create early-killed and late-killed treatments. Main plots measured 36 m2. Subplot treatments were assigned one of 12 soybean cultivars and included 11 edamame cultivars and one grain-type soybean cultivar (Table 2). Selection of edamame cultivars was based on seed availability of the more commonly used cultivars among commercial growers in the United States. Each subplot consisted of one 2.5-m row of 64 crop seeds. There was an inadequate supply of edamame seed for the larger plots necessary for accurately measuring crop yield.
Table 1 Details of cover crop treatments and cultural practices in field trials near Urbana, IL across 2014–2016 edamame growing seasons.

a For radish, winter-killed dates are based on first occurrence of four consecutive nighttime temperatures ≤3.9 C (Weil et al. Reference Weil, White and Lawley2009). For all other treatments, dates are when glyphosate was applied at a rate of 1,094 g ae ha−1.
b Planted using an adjustable hand seeder followed by incorporation with a two-gang cultipacker.
Table 2 Germplasm information for 1 soybean and 11 edamame cultivars used in each year of the study.

The entire study area was sprayed with a burndown application of glyphosate at 1,094 g ae ha−1 1 wk before planting to kill emerged weeds; this application was in addition to the cover crop kill application in cereal rye and canola. All cultivar seeds were treated with mefenoxam (3.37 g per 100 kg seed) and fludioxonil (2.27 g per 100 kg seed; Apron Maxx, Syngenta Crop Protection, Greensboro, NC) fungicides before planting. Cultivars were direct seeded into cover crop treatments using a no-till cone planter on May 27, 2014, May 28, 2015, and May 23, 2016.
Weeds emerging in the experiment were from the naturally occurring soil seedbank. Aside from suppression provided by cover crop residues and crop interference, no other weed management tactic was applied after crop planting.
Data Collection
Aboveground plant residues were measured 3 d before cultivar planting by harvesting two 0.5-m2 areas from each main plot. Weeds present before planting were included in the cover crop residue biomass sampling, because it was difficult to distinguish preplant weed biomass from cover crop biomass; however, weed occurrence was minimal at the sampling time, given it was early in the growing season and there had been at least one burndown application of glyphosate, even in the no-cover crop treatments. In each main plot, five 2.0-cm-diameter soil cores also were taken from a depth of 20 cm the day before edamame planting. Soil cores were composited by main plot, and soil moisture was determined gravimetrically.
The 100-seed mass and germination of cultivar seed lots were determined before planting. After planting, cultivar emergence was assessed daily across each subplot until all plants had at least one fully emerged trifoliate leaf. Weed emergence by species was assessed weekly in four randomly placed, permanent 0.25-m2 quadrats within main plots until 6 wk after planting. Weed seedlings were immediately removed from the permanent quadrats after species emergence was assessed. At 6 wk after planting, total aboveground weed biomass of each main plot was sampled from 1-m2 sampling areas. Weeds were sorted by species, dried to constant mass, and weighed. Because the objectives of this study focused on cultivar emergence and early-season weed density and biomass, the study was terminated 6 wk after planting. Growth responses beyond 6 wk after planting were at risk of being confounded by the single-row cultivar subplots.
Data Analysis
Data were analyzed with ANOVA in SAS (v. 9.4, SAS Institute, Cary, NC) using PROC MIXED. Data met assumptions of normality, equality of variance, and independence based on residual analysis (Hox Reference Hox2002). Two ANOVA models were used. Cover crop treatment was the fixed effect in the ANOVA model used to analyze cover crop biomass, soil moisture, weed emergence, and weed biomass data. Both cover crop and cultivar were fixed effects in the ANOVA model used to analyze crop emergence data. Year and replicate nested within year were treated as random effects in both models after initial analysis, as a fixed effect revealed no significant or biologically meaningful interactions with treatment effects. Main effects and interactions (when appropriate) were examined. Treatment comparisons were made using a protected Tukey’s means separation test at α=0.05.
Results and Discussion
Cover Crops
The goal of creating a gradient of cover crop residues was accomplished. Differences in residue biomass were observed among cover crop treatments before crop planting (P<0.001). Few preplant weeds were observed at glyphosate application, as evidenced by the bare-soil control averaging 43 kg ha−1 of plant residue. Residue biomass in other cover crop treatments ranged from 438 kg ha−1 for winter-killed radish to 9,003 kg ha−1 for late-killed cereal rye (Table 3). Cover crop biomass levels were comparable with results reported in previous research in the upper Midwest (Davis Reference Davis2010; Hill et al. Reference Hill, Renner, Sprague and Davis2016).
Table 3 Average decaying cover crop–residue biomass 3 d before planting and average gravimetric soil water content 1 d before planting in field trials near Urbana, IL, across 2014–2016 edamame growing seasons.Footnote a

a Values followed by different letters are significantly different at α=0.05 based on a protected Tukey’s means separation test.
Cover crop treatments influenced soil moisture at the time of planting. Soil moisture of late-killed cereal rye was highest, followed by early-killed cereal rye. All other cover crop treatments were comparable in soil moisture with the bare-soil treatment (Table 3). This result suggests that cover crop growth during the fallow season did not deplete surface soil moisture reserves and that the greater surface residue provided by cereal rye served to reduce evaporative soil water loss between cover crop killing and planting. Elevated soil moisture in a cereal rye cover crop, relative to bare soil, is common in central Illinois but varies regionally with soil physical properties and weather (Davis Reference Davis2010; Haramoto and Brainard Reference Haramoto and Brainard2012).
Crop Emergence
Crop emergence varied with both cover crop treatment and cultivar (P<0.001); however, there was no interaction (P=0.993). Therefore, while emergence differed among cultivars, their responses to cover crop treatments were similar.
With one exception, cover crop treatments resulted in similar total crop emergence as the bare-soil control, which was 71.1% (Table 4). Crop emergence in the late-killed cereal rye treatment was only 38.5%. The late-killed cereal rye treatment could have lowered soybean emergence by several mechanisms. Poor seed to soil contact is a common problem when planting into high levels (≥6,000 kg ha−1) of cover crop residues (Forcella Reference Forcella2013). Despite efforts to optimize planter performance in the present study, approximately 30% of crop seed in the late-killed cereal rye treatment was observed within cereal rye residue on the soil surface. The high level of residue in the late-killed cereal rye treatment (averaging 9,003 kg ha−1) resulted in poor seed-to-soil contact and was likely the primary mechanism reducing crop emergence in that treatment. Similar cereal rye–residue planting interference has been observed in other crops, including cotton (Gossypium hirsutum L.) (Kornecki et al. Reference Kornecki, Raper, Arriaga, Schwab and Bergtold2009) and corn (Zea mays L.) (Lawley et al. Reference Lawley, Weil and Teasdale2011). Additional mechanisms by which cover crop residues reduce crop emergence include increased seed predation, reduced available light for seedling growth, increased allelopathic chemical concentrations in the seed zone, and physical interference (Forcella Reference Forcella2013; Moore et al. Reference Moore, Gillespie and Swanton1994; Wortman et al. Reference Wortman, Francis, Bernards, Blankenship and Lindquist2013).
Table 4 Average crop emergence as influenced by cover crop treatment and cultivar 3 wk after planting in field trials near Urbana, IL, across 2014–2016 edamame growing seasons, with significance (P value) of main effects and their interaction included.

a Within a response variable, values followed by different letters are significantly different at α=0.05 based on a protected Tukey’s means separation test.
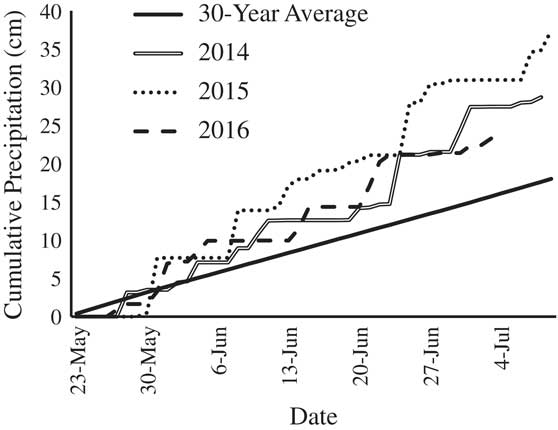
Four edamame cultivars, specifically ‘Gardensoy 42,’ ‘Misono Green,’ ‘Mojo Green,’ and ‘WSU 910a,’ had lower emergence than the grain-type cultivar, ‘Asgrow AG-3253’ (Table 4). Edamame emergence as low as 35% is not uncommon in field trials (Duppong and Hatterman-Valenti Reference Duppong and Hatterman-Valenti2005; Sánchez et al. Reference Sánchez, Kelley and Butler2005; Williams Reference Williams2015b). In the present study, edamame emergence ranged from 56.1% to 71.1% when averaged across all cover crop treatments (Table 4). Weather conditions were favorable after planting and may have accounted for the higher crop emergence than in previous edamame research. For instance, spring air temperatures were well above the minimum 12 C needed for soybean germination, and precipitation was above the 30-yr average, indicating cold stress, drought stress, and soil crusting were unlikely during crop germination and emergence (Figure 1).
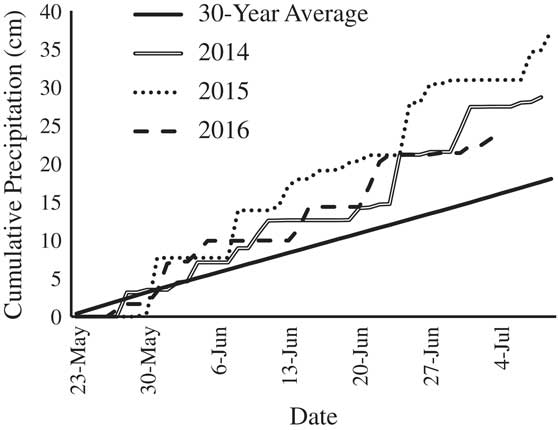
Figure 1 Cumulative daily precipitation by year, including the 30-yr average, from planting until the end of the experiment.
Weed Density
Predominant weed species included common lambsquarters (Chenopodium album L.), common purslane (Portulaca oleracea L.), common waterhemp [Amaranthus rudis (Moq.) J. D. Sauer], Palmer amaranth (Amaranthus palmeri S. Watson), and velvetleaf (Abutilon theophrasti Medik.). To minimize misidentification of newly emerged seedlings, A. rudis and A. palmeri were combined into an “Amaranthus species” group. In addition, a few other species were infrequently observed (Table 5). Portulaca oleracea accounted for 79% to 89% of emerged weeds in cover crop treatments with one exception; in the late-killed cereal rye treatment, P. oleracea accounted for only 52% of emerged weeds (Table 5).
Table 5 Effect of cover crop treatment on average total weed density, species-specific weed density, and total weed biomass in field trials near Urbana, IL, across 2014–2016 edamame growing seasons.Footnote a

a Within a column, means followed by different letters are significantly different at α=0.05 based on a protected Tukey’s means separation test.
b Species abbreviations: ABUTH, velvetleaf (Abutilon theophrasti Medik.); AMARA, Amaranthus spp., including common waterhemp [Amaranthus rudis (Moq.) J. D. Sauer], and Palmer amaranth (Amaranthus palmeri S. Watson); CHEAL, common lambsquarters (Chenopodium album L.); GRASS includes all grass species; OTHER includes infrequent observations of carpetweed (Mollugo verticillata L.), dandelion (Taraxacum officinale F. H. Wigg), horseweed (Erigeron canadensis L.), kochia [Bassia scoparia (L.) A. J. Scott], and Venice mallow (Hibiscus trionum L.); POROL, common purslane (Portulaca oleracea L.).
Total weed density varied across cover crop treatments. Weed density in cover crop treatments, ranked low to high, were late-killed cereal rye<early-killed cereal rye<bare soil<winter-killed radish<early-killed canola<late-killed canola (Table 5). The bare-soil treatment had an average weed density of 96.0 weeds m−2. Weed densities in the winter-killed radish, early-killed canola, and late-killed canola treatments were an average of 17.4, 50.2, and 74.8 weeds m−2 higher, respectively. There are several possible explanations for the higher weed emergence in the winter-killed radish and the two canola treatments compared with the bare-soil treatment. Although there were no significant soil moisture differences between radish, canola, and bare-soil treatments at crop planting, soil moisture is dynamic, and there may have been other times (not sampled) when soil moisture was more favorable for weed emergence in the radish and canola treatments compared with the bare-soil treatment. Haramoto and Brainard (Reference Haramoto and Brainard2012) report that nonincorporated oat (Avena sativa L.) cover crop residues can increase soil moisture relative to incorporated oat residues. Surface residues of radish and canola may have increased water infiltration and reduced evaporation from the soil surface, thereby conserving soil moisture relative to the bare-soil treatment and favoring weed seed germination.
The high weed density in the canola treatment was surprising, because a meta-analysis on Brassica cover crops such as canola found that incorporated Brassica residues produce allelochemicals that suppress weed emergence, but the effects were variable across environments and were short-lived (Haramoto and Gallandt Reference Haramoto and Gallandt2004). Given that canola residues in the present study were not incorporated and that weed density was not measured until several weeks after canola cover crop termination, it is unlikely allelopathy would have been observed, even if it had played a role in this study. Soil nitrogen also could affect weed density results. Decomposing cereal rye residues typically reduce soil nitrogen availability, which suppresses weed emergence (Hill et al. Reference Hill, Renner, Sprague and Davis2016). However, radish residues, which may also reduce soil nitrogen availability, have shown mixed effects on weed emergence. Hill et al. (Reference Hill, Renner, Sprague and Davis2016) reported that decomposing radish residues suppressed weed emergence, but Belfrey et al. (Reference Belfrey, Trueman, Vyn, Loewen and Van Eerd2017) and Lawley et al. (Reference Lawley, Teasdale and Weil2012) reported that, under certain conditions, radish residues, on the surface or incorporated into the soil, stimulated early spring (January to March) emergence of common chickweed [Stellaria media (L.) Vill.], C. album, common ragweed (Ambrosia artemisiifolia L.), and redroot pigweed (Amaranthus retroflexus L.). Both Belfrey et al. (Reference Belfrey, Trueman, Vyn, Loewen and Van Eerd2017) and Lawley et al. (Reference Lawley, Teasdale and Weil2012) hypothesized the stimulatory effect of radish on weed emergence was due to the higher soil nitrate levels relative to the control treatment. Lawley et al. (Reference Lawley, Teasdale and Weil2012) found that soil nitrate levels in the radish treatments were more than three times higher than the nitrate levels in the control treatment. Perhaps soil nitrate released from decomposing winter-killed radish residues in the present study contributed to increased weed emergence relative to bare soil.
Cover crop treatments that stimulate in-crop weed emergence would be problematic, in part because herbicide efficacy declines with weed density under certain conditions. Myers et al. (Reference Myers, Curran, Vangessel, Majek, Scott, Mortensen, Calvin, Karsten and Roth2005) reported that crop and weed plants with more biomass can interfere with herbicide coverage, potentially blocking herbicide delivery to small plants. As such, positive correlations have been observed between weed density and herbicide failure (Scursoni et al. Reference Scursoni, Forcella and Gunsolus2007). Moreover, positive relationships between soybean yield loss and weed density are well documented (Zimdahl Reference Zimdahl2004).
Both cereal rye treatments suppressed weed density relative to bare soil. The weed densities in the early-killed and late-killed cereal rye treatments were 18.3 and 88.7 weeds m−2 lower, respectively, than the bare-soil treatment. There are several possible reasons for lower weed density in cereal rye treatments. The late-killed cereal rye treatment had three times or more greater surface residue than all other cover crop treatments. Several studies report a negative relationship between residue biomass and weed emergence (Price et al. Reference Price, Monks, Culpepper, Duzy, Kelton, Marshall, Steckel, Sosnoskie and Nichols2016; Ryan et al. Reference Ryan, Mirsky, Mortensen, Teasdale and Curran2011). However, the amount of residue at crop planting does not fully account for differences in weed emergence among cover crop treatments in the present study; early-killed cereal rye and late-killed canola had similar biomass before crop planting, yet very different weed densities. The cereal rye treatments resulted in a thick, generally uniform, residue mat. In contrast, canola treatments resulted in standing stems and scattered leaves that decomposed quickly. Differences in residue pattern and persistence may have accounted for some of the differences in weed densities. Additionally, cereal rye is well known for suppressing a variety of summer annual weeds (DeVore et al. Reference DeVore, Norsworthy and Brye2013; Mehring et al. Reference Mehring, Stenger and Hatterman-Valenti2016; Moore et al. Reference Moore, Gillespie and Swanton1994), and allelochemicals are often implicated in weed suppression. Another potential factor limiting weed emergence in the cereal rye treatments could be the low soil nitrogen resulting from the decomposing cereal rye biomass. Cereal rye residues have a high carbon to nitrogen ratio, which can reduce soil nitrogen and emergence of nitrophilic species (Hill et al. Reference Hill, Renner, Sprague and Davis2016).
Midseason Weed Biomass
All cover crop treatments reduced weed biomass relative to the bare-soil treatment, which averaged 1,554 kg ha−1. Weed biomass, ranked low to high, was late-killed cereal rye=early-killed cereal rye<late-killed canola<early-killed canola=winter-killed radish<bare soil (Table 5). Radish and canola treatments, despite having higher weed densities than bare soil, exhibited a strong suppressive effect on weed growth. Cereal rye treatments had the lowest weed density (77.7 and 7.3 weeds m−2, early- and late-killed, respectively) and resulted in the lowest weed biomass 6 wk after planting (226 and 104 kg ha−1, early- and late-killed, respectively).
Recent studies report that as cereal rye residues increase, Amaranthus species control increases (Price et al. Reference Price, Monks, Culpepper, Duzy, Kelton, Marshall, Steckel, Sosnoskie and Nichols2016; Ryan et al. Reference Ryan, Mirsky, Mortensen, Teasdale and Curran2011). Several Amaranthus species, including A. rudis and A. palmeri, are especially problematic for producers, because they have high fecundity and herbicide resistance is becoming more common. Ryan et al. (Reference Ryan, Mirsky, Mortensen, Teasdale and Curran2011) found a negative relationship between cereal rye–residue levels and biomass of smooth pigweed (Amaranthus hybridus L.) and A. retroflexus, but the relationship was not linear at low cereal rye–residue levels. Price et al. (Reference Price, Monks, Culpepper, Duzy, Kelton, Marshall, Steckel, Sosnoskie and Nichols2016) found that the use of cover crops reduced herbicide-resistant A. palmeri populations relative to winter-fallow systems. Price et al. (Reference Price, Monks, Culpepper, Duzy, Kelton, Marshall, Steckel, Sosnoskie and Nichols2016) also reported a negative relationship between cover crop–residue biomass and A. palmeri survival. Ryan et al. (Reference Ryan, Mirsky, Mortensen, Teasdale and Curran2011) reported giant foxtail (Setaria faberi Herrm.) and Amaranthus species emergence was completely suppressed at cereal rye–residue levels above 15,000 kg ha−1, which is considerably higher than residue levels in the present research.
Only one cover crop treatment in this study appeared to have potential for weed management in edamame. The early-killed cereal rye treatment was the best candidate for edamame production, because it provided measurable weed suppression without compromising crop emergence. Terminating cereal rye in early- to mid-April, 6 to 7 wk before edamame planting, limited cereal rye residue to 2,375 kg ha−1, yet still provided a 20% reduction in weed density and an 85% reduction in weed biomass without any loss of crop emergence relative to the bare-soil control. Additional research is needed on coupling the early-killed cereal rye cover crop with other weed management tactics, including in-crop weed management using additional chemical, cultural, and physical control tactics.
Acknowledgments
The authors wish to thank Jim Moody, Nick Hausman, Michael Douglas, and the many undergraduate students who helped conduct the field experiment. No conflicts of interest have been declared.