INTRODUCTION
In agroforestry parkland systems, trees and crops interact with one another in the process of capturing resources from the environment that are required for growth and development (Bayala et al., Reference Bayala, Teklehaimanot and Ouédraogo2002; Ong and Leakey, Reference Ong and Leakey1999; Sanchez, Reference Sanchez1995). The interaction between trees and crops can be complementary, neutral or competitive for both underground and aboveground resources (Bayala et al., Reference Bayala, Teklehaimanot and Ouédraogo2002; Sanchez, Reference Sanchez1995). Trees have been reported to affect light and rainwater distribution as well as soil physical and chemical properties under their crowns (Bayala et al., Reference Bayala, Balesdent, Marol, Zapata, Teklehaimanot and Ouédraogo2006; Rao et al., Reference Rao, Nair and Ong1998). Several previous studies pointed out that tree shade was the major factor causing a decrease in the yield of associated crops such as millet (Bayala et al., Reference Bayala, Teklehaimanot and Ouédraogo2002; Boffa et al., Reference Boffa, Taonda, Dickey and Knudson2000; Kessler, Reference Kessler1992; Maiga, Reference Maiga1987). However, Jonsson et al. (Reference Jonsson, Ong and Odongo1999) reported a positive effect of tree shade on millet yield due to the increase in soil water availability and the reduction in temperature under shade.
Light reduction by trees in parkland systems (Bayala et al., Reference Bayala, Teklehaimanot and Ouédraogo2002; Belsky et al., Reference Belsky, Amundson, Duxbury, Riha, Ali and Mwonga1989; Boffa et al., Reference Boffa, Taonda, Dickey and Knudson2000; Kessler, Reference Kessler1992) has been reported to result in a reduction in temperature under tree crowns (Boffa, Reference Boffa1999; Garcia-Barrios and Ong, Reference Garcia-Barrios and Ong2004; Rao et al., Reference Rao, Nair and Ong1998). Because of the temperature reduction, evapo-transpiration under tree crowns is reduced compared to the open field as reported by Belsky et al. (Reference Belsky, Amundson, Duxbury, Riha, Ali and Mwonga1989) and Boffa (Reference Boffa1999). Despite rainwater interception (Boffa, Reference Boffa1999;, Breman and Kessler, Reference Breman and Kessler1995; Ong and Swallow, Reference Ong, Swallow, van Noordwijk, Cadisch and Ong2003), several workers reported higher soil water availability under tree crowns due to the reduced evapo-transpiration rate (Garcia-Barrios and Ong, Reference Garcia-Barrios and Ong2004; Ong et al., Reference Ong, Black, Marshall, Corlett, Ong and Huxley1996; Ong and Swallow, Reference Ong, Swallow, van Noordwijk, Cadisch and Ong2003). However, trees and crops compete for the available soil water if their rooting systems occupy the same niche. Trees and crops also compete for soil nutrients although higher soil nutrient content under tree crowns than in the open field is common in parklands (Bayala et al., Reference Bayala, Balesdent, Marol, Zapata, Teklehaimanot and Ouédraogo2006; Boffa, Reference Boffa1999; Breman and Kessler, Reference Breman and Kessler1995). The competition for these growth resources may reduce associated crops performance under trees even though, one might think that high soil nutrient and water availability underneath trees provide favourable conditions for crop growth. The performance of any crop under tree crowns, however, depends on its ability to compete with trees for soil water and nutrients and its photosynthetic efficiency under reduced light conditions beneath tree crowns.
Thus, the favourable microclimate and the edaphic conditions created under tree shade may favour shade-tolerant crops such as taro more than shade-intolerant crops such as millet. Although both crops may compete with trees for soil nutrients and water, light reduction under trees may be more favourable to taro than millet. However, contradictory results of shade effect on taro performance were reported in previous studies. Caesar (Reference Caesar1980) found a reduction of taro corm yield because of a combination of shade and water stress effects. In contrast, Miyasaka et al. (Reference Miyasaka, Ogoshi, Tsuji and Kodani2003) observed that low temperature and low light availability did not significantly affect taro corm yield. In addition, Rogers and Iosepha (Reference Rogers and Iosepha1993) obtained best quality of taro corms under shade condition. However, no study comparing the effect of parkland trees on shade-tolerant crops such as taro and shade-intolerant crops such as millet by growing them simultaneously has been reported in the literature.
Therefore, the present research was conducted to assess the shading effect of néré (Parkia biglobosa) and baobab (Adansonia digitata) on the photosynthesis and biomass production of associated millet (Pennisetum glaucum), a shade-intolerant crop, and taro (Colocasia esculenta), a shade-tolerant crop, in an agroforestry parkland system.
MATERIALS AND METHODS
Study site
The study was carried out between 2006 and 2008 in a parkland at Nobéré, a village located at 11°30′ North and 00°58′ West in the south-central region of Burkina Faso (West Africa). The area is Sudanian savanna zone with a rainy season of 4–5 months and annual rainfall of 800–1000 mm (Nikiéma, Reference Nikiéma2005). Figure 1 gives the rainfall and the number of rainy days in 2006, 2007 and 2008. The rainfall in 2006 (843.4 mm) was within the normal range of the region while in 2007 and 2008, the annual rainfall was 46.1 mm and 223.3 mm higher than the normal maximum for the region, respectively (Figure 1).

Figure 1. Rainfall and number of rainy days during 2006, 2007 and 2008 at Nobéré in Burkina Faso.
The soil is luvisol with a sandy loam texture (clay = 11.5%, silt = 20.1% and sand = 68.4%) with low nutrient content (N = 0.69 g kg−1, P = 0.14 g kg−1 and K = 0.50g kg −1) and low organic matter content (1.32%) in the 10-cm upper soil layer.
Tree selection
The two tree species, Adansonia digitata L. and Parkia biglobosa (Jacq.) Benth, were chosen for the study because of their high frequency in parklands of Sudanian savanna zone and also because of their contrasting phenology and light interception. Eight isolated mature trees of each of the two tree species (A. digitata and P. biglobosa) were randomly selected within the 100 ha parkland. Tree characteristics such as height, circumference at 1.30 m aboveground and average crown (north–south and east–west orientations) diameters were measured and used to plan the experimental design. Average height, diameter at 1.30 m and crown diameter were 15.50 ± 1.10 m, 188.32 ± 18.93 cm and 16.52 ± 0.82 m, respectively for baobab and 11.07 ± 0.33 m, 70.07 ± 3.75 cm and 17.14 ± 0.83 m, respectively for néré trees.
Experimental design and layout
Eight mature trees of each of P. biglobosa and A. digitata were randomly selected within an area of 100 ha. The area around each selected tree was then subdivided into three concentric zones according to its crown size as follows:
Zone A – from the trunk of each tree up to half of the radius of the tree crown;
Zone B – from half of the radius of the tree crown up to the edge of the crown;
Zone C – from the edge of the tree crown up to 3 m away;
A control plot (Zone H) of an area of 8 × 8 m was delimited for each sample tree to assess crop performance in the open area. The control plot was situated at least 40 m away from the edge of the crown of the sample tree but not shaded by any of the surrounding trees at any time of the day throughout the cropping season (Figure 2). Dividing the area under tree into concentric zones was preferred to directional transects to reduce directional biases related to leaf and rainfall, and micro-variability of soil fertility and to separate the different influence zones of tree as defined by Rao et al. (Reference Rao, Nair and Ong1998).

Figure 2. Experimental design with concentric zones (A, B, C) around a tree trunk and the control (H).
Taro was grown under four selected trees of each tree species and millet under the other four trees of each species. The association of crops to selected trees was randomly done for each tree species.
Parameters assessment
During three cropping seasons (2006, 2007 and 2008), leaf area index (LAI), photosynthesis parameters and biomass yield of millet and taro were assessed.
Leaf area index. Leaf area was determined digitally using a planimetric method. Thirty (30) leaves were randomly collected per concentric zone and in the control plot for each crop under each studied tree. The outline of each collected leaf was traced on a sheet of paper. Long and large leaves were cut into two or three pieces to fit on a sheet. The drawings were scanned and each leaf area was computed using the software ImageJ (NIH, USA, Freeware, Version 1.34s). An average leaf area was calculated per concentric zone and in the control plot for each crop. The average value was used to calculate the total leaf area per plant by scaling up using the number of leaves for each plant. Then, the LAI was calculated by dividing the total leaf area (m2) per plant by the ground area (m2) occupied by a single plant.
Photosynthesis. Measurements of photosynthetic parameters were made using CIRAS-1 (PP-Systems, UK) a portable infra-red gas analyser. Four plants were randomly selected and labelled in each concentric zone under four trees (two trees associated with taro and two trees with millet) and in the control plot. The second completely expanded leaf from the top of each selected plant was labelled for the measurements of the day. Measurements were made at 1 h intervals, from 7 am to 5 pm for 4 days during vegetative growth phase of crops in the 2008 growing season. Each leaf was enclosed and placed perpendicular to the sun in the leaf chamber PLC-B (CIRAS-1, PP-Systems, UK). Then, net photosynthesis (PN), stomatal conductance (gs), photosynthetically active radiation (PAR) and transpiration (Et) were recorded by the instrument (CIRAS-1).
The daily integral of PN was calculated as the sum of PN per hour of daytime. The PN per hour was the instantaneous PN (μmol CO2 m−2 s−1) measured at the given hour multiplied by 3600 s. The radiation use efficiency (RUE) and water use efficiency (WUE) were calculated as the ratios between integrals of PN and PAR, and between integrals of PN and Et, respectively. In some experiments, RUE was labelled as LUE (or light use efficiency, see Grace et al., Reference Grace, Nichol, Disney, Lewis, Quaife and Bowyer2007). It was expressed as mmol of CO2 captured per mol of PAR absorbed. WUE was defined as mmol of CO2 captured per mol of water lost.
Assessment of crop biomass
At harvest, the biomass production of crops was assessed in each concentric zone and in the control plot. For the biomass production of each zone, straws and grains of millet or tubers of taro were dried in an oven at 80 °C for 48 h and weighed. The biomass production (kg ha−1) was the sum of dry matter of straws and grains or tubers of each zone.
Data analysis
In the experimental design used for the study, it was not possible to randomise the concentric zones that are a representation of the gradient of tree effect. The variation between zones cannot be analysed by ANOVA because it is not possible to calculate a valid estimation of error as residuals of zones are correlated (see Wilson et al., Reference Wilson, Brook and Tomlinson1998). Therefore, the variation of data according to cropping years, tree species and crops were analysed using the ANOVA general linear model (GLM) while the variation between concentric zones and the control plot was tested using pairwise t-tests. All these analyses were performed using modules of the software Minitab 15 (Minitab Inc., USA). After a GLM analysis, means were compared using Tukey's test at a confidence level of 95%. The t-tests were performed assuming that the difference between the two zones compared was equal to zero and this hypothesis was rejected in case of a significant result of the test. A single rectangular hyperbolic regression (Y = aX/(X+b)) was used to compare the PN response curve of millet and taro according to PAR variation using SigmaPlot 12 (Systat software, USA). The correlation between crops biomass and photosynthesis parameters were tested using Pearson's correlation.
RESULTS
Biomass production
Aboveground dry biomass varied significantly between crops, cropping years and tree species (P < 0.001, P < 0.001 and P < 0.01, respectively). Biomass production of millet under baobab (2220.9 ± 188.46 kg ha−1) was not statistically different to the value in the control plot (1820.4 ± 241.70 kg ha−1) but higher as compared to the value under néré (1303.9 ± 111.05 kg ha−1). In contrast, biomass production of taro was higher under néré compared to the values under baobab and in the control plot (1025.3 ± 80.14 kg ha−1 and 752.2 ± 122.50 kg ha−1, respectively). But no significant difference was found between the values in the control plot and under baobab (875.4 ± 105.78 kg ha−1) (Figure 3).

Figure 3. Mean biomass of millet (Pennisetum glaucum) and taro (Colocasia esculenta) for three consecutive cropping seasons (2006–2008) according to zones around baobab (Adansonia digitata), néré (Parkia biglobosa) and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – control plot.
According to the results of pairwise t-test, biomass production of millet was lower in zone A under both tree species compared to the values in zones B and C and the control plot. In contrast, the biomass of taro was higher in zone A under baobab compared to the values in zones B, C and the control plot. Biomass production of taro under néré decreased from the tree trunk to the open area (Figure 3).
Leaf area index
The average LAI varied significantly between the three cropping years (P < 0.01), the two crop species (P < 0.001), the two tree species (P < 0.001) and the three concentric zones under trees (P < 0.001). There were also significant interactive effects of crops and tree species (P < 0.05) on LAI. LAI of millet was higher in 2007 (P < 0.01) than in 2006 and 2008 (1.2 ± 0.06, 0.9 ± 0.08 and 1.0 ± 0.05, respectively) while no significant difference in LAI was found for taro between the cropping years. Taro had a significantly higher LAI over the three cropping years (P < 0.001) compared to millet (1.7 ± 0.14 and 1.0 ± 0.04, respectively). LAI of millet was almost the same under both baobab and néré while LAI of taro was significantly higher (P < 0.001) under néré than baobab (2.6 ± 0.35 and 1.3 ± 0.22, respectively) (Figure 4).

Figure 4. Average leaf area index (LAI) of millet (Pennisetum glaucum) and taro (Colocasia esculenta) for three consecutive cropping seasons (2006–2008) according to zones around baobab (Adansonia digitata), néré (Parkia biglobosa) and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – control plot.
The results of pairwise t-test showed that LAI of millet was significantly higher in zone C (1.4 ± 0.13) under baobab when compared to the values in zones A, B and the control plot (0.8 ± 0.12, 1.0 ± 0.09 and 0.9 ± 0.12, respectively). Except the LAI of millet in zone C that was higher compared to the control plot (1.1 ± 0.08 and 0.9 ± 0.06, respectively), no significant difference in the LAI was noted between the other zones under néré. The LAI of taro decreased significantly from the tree trunk to the open area under both tree species (Figure 4). However, the LAI of taro in zone C under baobab was lower than the value in the control plot (0.5 ± 0.09 and 0.9 ± 0.09, respectively).
Photosynthetically active radiation
The diurnal maximum PAR recorded in the open area at 12 h in 2008 was 1.54 mmol m−2 s−1 (Figure 5). This value was reduced by 82%, 31% and 18% in zone A, B and C of baobab, respectively while the reduction was accented in zones of néré with 92%, 63% and 21% in zone A, B and C, respectively. The PAR availability did not significantly vary according to daytime in zone A of both tree species. In the other zones, PAR increased from 7 to 12 h and then decreased until 17 h (Figure 5).

Figure 5. Diurnal course of photosynthetically active radiation (PAR) recorded under baobab (Adansonia digitata) and néré (Parkia biglobosa) according to zones around trees and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso (2008). Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – control plot.
Net photosynthesis
The mean PN recorded in the control plot was higher for millet plants (24.79 ± 1.621 μmol CO2 m−2 s−1) compared to taro plants (14.3 ± 0.71 μmol CO2 m−2 s−1). The highest PN values for both crops under both trees species were recorded between 10 and 15 h in zone C and the control plot (Figure 6). In the association of millet with baobab tree, the highest values of PN were observed between 8 and 9 h in zone A and between 12 and 13 h in zone B. PN values did not vary diurnally in zones A and B under néré for both crops and under baobab for taro only. The main difference of PN between zones was noted between 9 and 15 h. During this period, the PN of the two crops was reduced under the tree crown (A and B) compared to the values in zone C and the control and the reduction was more noticeable in zones under néré than under baobab (Figure 6).

Figure 6. Diurnal course of the net photosynthesis (PN) in 2008 of millet (Pennisetum glaucum) and taro (Colocasia esculenta) associated with baobab (Adansonia digitata) and néré (Parkia biglobosa) according to zones around trees and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – control plot.
For both crops, the daily integral of PN varied between trees species (P < 0.01 and P < 0.05 for millet and taro, respectively). As shown in Figure 7, the daily integral of PN in zone A under baobab was higher for taro compared to millet (864.7 ± 174.94 mmol CO2 m−2 day−1 and 227.7 ± 31.42 mmol CO2 m−2 day−1, respectively). In zones A and B under néré, the daily integral of PN of taro was higher (566.5 ± 49.16 mmol CO2 m−2 day−1 and 692.3 ± 76.19 mmol CO2 m−2 day−1, respectively) compared to the values of millet (307.1 ± 44.6 mmol CO2 m−2 day−1 and 450.4 ± 39.9 mmol CO2 m−2 day−1, respectively).

Figure 7. Daily integral of net photosynthesis (PN) in 2008 of millet (Pennisetum glaucum) and taro (Colocasia esculenta) associated with baobab (Adansonia digitata) and néré (Parkia biglobosa) according to zones around trees and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – Control plot.
Radiation use efficiency
The radiation use efficiency was significantly different between crop species (P <0.05). Higher RUE was recorded for millet (20.3 ± 2.45 mmol CO2 mol−1 PAR) compared to the value of taro (14.7 ± 1.15 mmol CO2 mol−1 PAR). There was no significant difference in RUE between tree species in both crops. However, RUE values were significantly different between zones for taro but not for millet. RUE values of taro decreased from the tree trunk to the control plot (Figure 8).

Figure 8. Radiation use efficiency (RUE) in 2008 of millet (Pennisetum glaucum) and taro (Colocasia esculenta) associated with baobab (Adansonia digitata) and néré (Parkia biglobosa) according to zones around trees and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – Control plot.
Water use efficiency
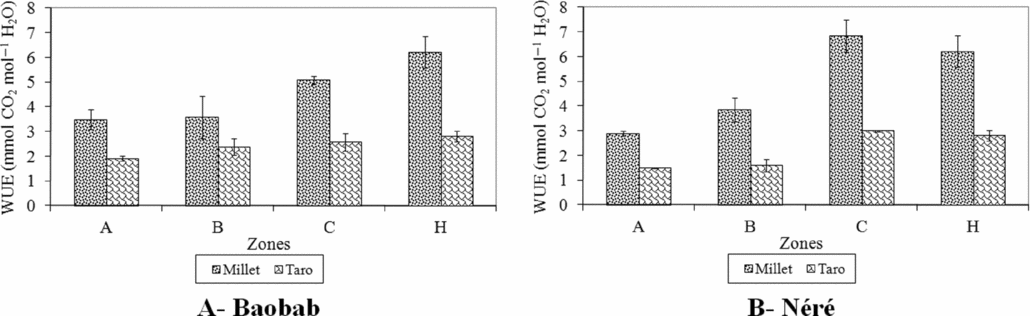
There was significant difference in the water use efficiency (WUE) between the two crops (P < 0.001). WUE of millet was higher compared to the value of taro (4.8 ± 0.45 mmol CO2 mol−1 H2O and 2.3 ± 0.17 mmol CO2 mol−1 H2O, respectively). WUE values did not significantly differ between tree species for both crops. The values of WUE increased from the area under the tree crown (zones A and B) to zone C and the control plot for both crops (Figure 9).
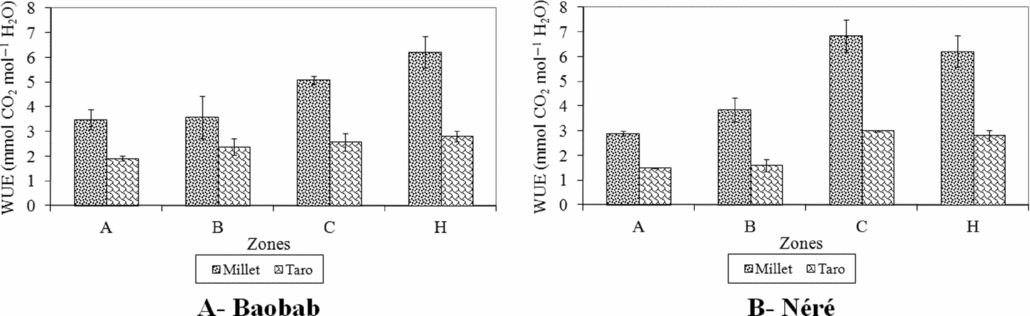
Figure 9. Water use efficiency (WUE) in 2008 of millet (Pennisetum glaucum) and taro (Colocasia esculenta) associated with baobab (Adansonia digitata) and néré (Parkia biglobosa) according to zones around trees and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – Control plot.
Stomatal conductance (gs)
The maximum and minimum stomatal conductances (gs) varied significantly between crops (P < 0.01) but not between tree species. Maximum and minimum values of gs were higher for taro (589.9 ± 67.60 mmol m−2 s−1 and 150.8 ± 22.05 mmol m−2 s−1, respectively) compared to the values for millet (319.4 ± 25.44 mmol m−2 s−1 and 69.6 ± 13.13 mmol m−2 s−1, respectively). The maximum gs of taro was higher compared to millet in all zones while the minimum gs of taro was only higher in zones A and B compared to millet (Figure 10). The gs diurnal course of taro did not vary significantly between zones of baobab but gs values from 13 to 16 h were higher in the control plot compared to zones under néré influence (Zones A, B and C). The gs values of millet were significantly different between zones of both tree species. The main difference of gs between zones was observed between 10 and 15 h. During this period, the gs of millet was lower in zone A of baobab compared to other zones while the gs value of millet was lower in all shaded zones of néré (A and B) compared to the control plot. A significant difference of gs was also noted between shaded zones of néré and the zone C from 10 to 12 h (Figure 10). A slight decrease of gs in the open area (zone H) was observed for both crops around 12 h.

Figure 10. Diurnal course of stomatal conductance (gs) in 2008 of millet (Pennisetum glaucum) and taro (Colocasia esculenta) associated with baobab (Adansonia digitata) and néré (Parkia biglobosa) according to zones around trees and in a control plot in an agroforestry parkland at Nobéré in Burkina Faso. Zone A – from the trunk of each tree up to half of the radius of the tree crown; Zone B – from half of the radius of the tree crown up to the edge of the crown; Zone C – from the edge of the tree crown up to 3 m away; Zone H – Control plot.
Relationships between PAR, photosynthesis parameters (daily integral of PN, RUE and WUE) and the biomass production of crops
There was a strong significant relationship between PN of both crops and PAR availability (P < 0.001). The two regression equations for millet and taro were PN = 91.86 PAR/(PAR + 3.74) (r 2 = 82%) and PN = 26.69 PAR/(PAR + 1.09) (r 2 = 79%), respectively. The initial slope was quite the same for both the response curves of millet and taro, respectively. However, the half-maximum PN is reached by millet for PAR value three times higher compared to the PAR value for half-maximum PN of taro (3.74 mmol m−2 s−1 and 1.09 mmol m−2 s−1, respectively). The PN response curves showed that less increase in PN of taro was recorded when increasing PAR value compared to millet (Figure 11). Therefore, the maximum PN value of millet was three times higher compared to taro (91.86 μmol m−2 s−2 and 26.69 μmol m−2 s−2, respectively).

Figure 11. Relationship between the net photosynthesis (PN) of millet (Pennisetum glaucum) and taro (Colocasia esculenta) and the photosynthetically active radiation (PAR) in an agroforestry parkland at Nobéré in Burkina Faso.
The biomass production of millet was negatively and significantly correlated with RUE (r = −0.37; P < 0.001). No significant correlation was found between daily integral of PN and WUE and the biomass production of millet. In contrast, biomass production of taro was positively and significantly correlated with RUE (r = 0.32; P < 0.01). There was also a significant negative correlation between biomass of taro and WUE (r = −0.38; P <0.001). The daily integral of PN was negatively and significantly correlated to the biomass production of taro (r = −0.38; P < 0.001).
DISCUSSION
Crop biomass production
The highest biomass production of millet was recorded under baobab and was not statistically different from what was obtained in the open field (Zone H) while under néré, the biomass production in zone A was lower compared to the control plot. In contrast, the best performance of taro was obtained under néré where higher biomass production was recorded in zones A and B compared to the control. Under baobab, only biomass production in zone A was higher compared to the control plot. Therefore, the high performance of taro under shade could be due to its adaptation to the microclimate created by the tree shade. In addition, the increase in photorespiration due to the high temperature and water stress, and light saturation could explain its low dry matter accumulation in the open field as previously reported in some C3 plants (Hay and Porter, Reference Hay and Porter2006; Osborne et al., Reference Osborne, Wythe, Ibrahim, Gilbert and Ripley2008; Yang et al., Reference Yang, Zhao and Zhang2004).
Crop LAI
In the present study, an increase in LAI of both crops was observed under tree crowns compared to the control. This corroborates the findings of previous studies which reported that plants usually adapt to shade by increasing their total leaf area and number of leaves (Caesar, Reference Caesar1980; Lin et al., Reference Lin, McGraw, George and Garrett2001; Wilson et al., Reference Wilson, Brook and Tomlinson1998; Wong, Reference Wong, Shelton and Stur1991). The highest LAI of taro was found in the most shaded zones: within zones A and B of néré trees and zone A of the baobab trees. Johnston and Onwueme (Reference Johnston, Onwueme and 1998) observed a similar result of the effect of shade on the leaf area of taro. In contrast, the highest LAI of millet was recorded in zone C of baobab trees while the LAI of millet in all zones under néré was similar to the control plot.
A positive linear relationship between LAI and light interception, which was directly related to crop performance, was reported by Ewert (Reference Ewert2004) and Monteith (Reference Monteith1977). The LAI of taro increased under shade as part of the plant's strategy to enable it to intercept more light and consequently it was able to grow better under shade. In contrast, millet did not show clear differences in LAI under shaded and open areas, suggesting the absence of an adaptation strategy of the crop to shade conditions.
Crop photosynthesis
The diurnal trend of PN of both crops in the control plot showed no peak as also noted in a previous study by Boegh et al. (Reference Boegh, Soegaard, Hanan, Kabat and Lesch1999). Similarly, no peak was observed for stomatal conductance (gs) of both crops. As shown above, the results of gs of both crops in the open area slightly decreased around noon when the peak was expected. The peak of PN was also expected around noon when daylight intensity was optimum, but at this time, temperature was also high that resulted in decrease of gs. Therefore, high temperature could explain the lack of a peak in the diurnal course of PN in the control plot (open area).
The average PN of millet (24.3 ± 5.01 μmol CO2 m−2s−1) recorded in the control plot in the present study was within the range of values of 4.4–62.94 μmol CO2 m−2s−1 reported by previous studies (Boegh et al., Reference Boegh, Soegaard, Hanan, Kabat and Lesch1999; McPherson, Reference McPherson and Slatyer1973; Masojidek et al., Reference Masojidek, Trivedi, Halshaw, Alexiou and Hall1991; Tewolde et al., Reference Tewolde, Dobrenz and Voigt1993; Warner and Edwards, Reference Warner and Edwards1988). The average PN of taro in the control plot (13.8 ± 1.18 μmol CO2 m−2s−1) was also within the range of values (4.1–30.93 μmol CO2 m−2s−1) reported by Sato et al. (Reference Sato, Kawai and Fukuyama1978), Schaffer and O'Hair (Reference Schaffer and O'Hair1987) and Sugimoto et al. (Reference Sugimoto, Fujita, Koesmaryono and Sato1997). The PN of both crops increased from the tree trunk to the open area, showing that the photosynthesis of both crops was limited by PAR availability under tree crowns. However, PN in taro was less affected in the shaded area than millet. Therefore, the photosynthesis system of taro plant was adapted to shade as reported by Onwueme (Reference Onwueme1999) and Onwueme and Johnston (Reference Onwueme and Johnston2000).
In the present study, it was observed that minimum stomatal conductance of taro was the same in shaded zones and in the open field while the minimum stomatal conductance of millet decreased with an increase in shade. Such adaptation to shade condition was noted by Onwueme and Johnston (Reference Onwueme and Johnston2000) who showed an increase in stomatal density of taro leaves under shade compared to the full sunlight. It may be concluded that taro, through such an adaptation, increased its efficiency of PAR capture in the shaded area. Indeed, the results of the analysis of the radiation use efficiency (RUE) of taro showed a decrease from the tree trunk to the open area while no variation was observed in millet between zones. The increase in RUE of taro may have helped it to avoid a drastic reduction of net assimilation production in shade conditions.
The daily integral of PN increased for both crops from tree trunk to the open field. Despite this trend of the daily integral of PN per unit leaf area, higher biomass production of taro was recorded in shaded zones compared to the open. Such production was the result of the increase in the crop LAI in the shaded zones. Therefore, the increase in leaf area in the shaded area may have allowed taro to accumulate more daily assimilate per plant than millet. These results confirm that taro was better adapted to the conditions under tree shade than millet.
The WUE values of both crops were lower in shaded areas (zone A and B) compared to zone C and control plot and WUE value of taro was lower than that of millet. The low WUE values in shaded zones indicate that both crops lost more water per unit PAR captured perhaps due to more stomata that are open. The result of the stomatal conductance indicates that taro had a higher total area of open stomata than millet in all zones because higher stomatal conductance indicates higher number of open stomata or larger aperture of stomata. The stomatal conductance in millet leaves increased from the tree trunk to the control plot while the stomatal conductance did not vary between zones in taro. These results indicate that taro in shaded zones adapted its stomata to harvest more CO2, but at the same time also lost more water as a result of high area of open stomata for gas exchange. In contrast, millet had smaller area of open stomata, less transpiration in shaded areas and captured less amount of PAR. The lack of significant difference between the stomatal conductance of taro leaves in the open area and under shade of baobab could be an expression of the stresses due to high irradiance, high temperature and low soil moisture in the open field. However, high reduction of PAR under néré combined with the decrease of topsoil moisture in the afternoon (13–16 h) resulted in a lower gs of taro in shaded zones compared to the open area. The increase in WUE of millet from the tree trunk to the open indicates that WUE in millet increased with an increase in environmental stress. This was in accordance with the findings of Singh and Singh (Reference Singh and Singh1995) who showed that WUE of millet not only increased with an increase in water stress but also with an increase in light availability which is limiting factor under shade for crops.
Relationship between photosynthesis parameters and crop performance
The curves of the relationship of PAR availability and PN of millet and taro show that millet increased more its PN when increasing PAR compared to taro. The lower increase in PN of taro than millet from shade area to the open field is due to the photosynthetic pathway of the two crops (C4 and C3, for millet and taro, respectively). In fact, under high irradiance such as in the open field of Sahel region, C3 plants are limited by their low light saturation point compared to C4 plants (Hay and Porter, Reference Hay and Porter2006; Osborne et al., Reference Osborne, Wythe, Ibrahim, Gilbert and Ripley2008). In addition, due to the competition between CO2 and O2 fixation in the C3 pathway of photosynthesis combined with the energy cost to convert 3-phosphoglycolate to phosphoglycerate, the quantum yield of C3 plant is lower compared to C4 pathway of photosynthesis.
Accordingly, with the behaviour of taro under shade conditions as reported by Johnston and Onwueme (Reference Johnston, Onwueme and 1998) and Onwueme and Johnston (Reference Onwueme and Johnston2000), the morphological and anatomical adaptations of the crop to shade resulted in a better performance of taro than millet in the shaded area under tree crowns. Indeed, the strong and positive correlation between the biomass production of taro and RUE and daily integral of PN shows a strong adaptation of taro to shade. In contrast, the negative correlation observed between RUE and millet performances may also indicate that millet was indeed shade-intolerant. Contrary to RUE, WUE of taro was negatively correlated with its growth performances. Therefore, taro production in open area seems to be limited by water availability as also observed by Caesar (Reference Caesar1980) who noted the importance of light and water in taro production.
In summary, taro was more efficient than millet in exploiting the favourable conditions (reduced temperature, higher nutrient and water contents of soil) under baobab and néré trees by changing its leaf morphology and anatomy. Therefore, it could be a worthy strategy to replace millet by taro under dense tree crowns in agroforestry parkland systems. The adoption of such a practice will certainly help to maintain high density of tree on farmed parklands of semi-arid areas.
Acknowledgements
This study was funded by the project SAFRUIT/INCO Contract Number 015465 of the European Commission. Yonli H. Hermann is acknowledged for field assistance. We wish to thank the farmers in Nobéré for their permission and participation in the field experiment.