





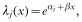

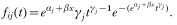







Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Dreyer, Gerusa
Addiss, David
and
Norões, Joaquim
2005.
Does longevity of adult Wuchereria bancrofti increase with decreasing intensity of parasite transmission? Insights from clinical observations.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 99,
Issue. 12,
p.
883.
Stolk, Wilma A
de Vlas, Sake J
and
Habbema, J Dik F
2006.
Advances and challenges in predicting the impact of lymphatic filariasis elimination programmes by mathematical modelling.
Filaria Journal,
Vol. 5,
Issue. 1,
Wetten, Sally
Collins, Richard C.
Vieira, Juan Carlos
Marshall, Clare
Shelley, Anthony J.
and
Basáñez, María-Gloria
2007.
Vector competence for Onchocerca volvulus in the Simulium (Notolepria) exiguum complex: Cytoforms or density-dependence?.
Acta Tropica,
Vol. 103,
Issue. 1,
p.
58.
Lefèvre, Thierry
and
Thomas, Frédéric
2008.
Behind the scene, something else is pulling the strings: Emphasizing parasitic manipulation in vector-borne diseases.
Infection, Genetics and Evolution,
Vol. 8,
Issue. 4,
p.
504.
Dawes, Emma J
Churcher, Thomas S
Zhuang, Shijie
Sinden, Robert E
and
Basáñez, María-Gloria
2009.
Anopheles mortality is both age- and Plasmodium-density dependent: implications for malaria transmission.
Malaria Journal,
Vol. 8,
Issue. 1,
Paily, K. P.
Hoti, S. L.
and
Das, P. K.
2009.
A review of the complexity of biology of lymphatic filarial parasites.
Journal of Parasitic Diseases,
Vol. 33,
Issue. 1-2,
p.
3.
Michael, Edwin
Snow, Lucy C.
and
Bockarie, Moses J.
2009.
Ecological Meta-Analysis of Density-Dependent Processes in the Transmission of Lymphatic Filariasis: Survival of Infected Vectors.
Journal of Medical Entomology,
Vol. 46,
Issue. 4,
p.
873.
Churcher, Thomas S
Dawes, Emma J
Sinden, Robert E
Christophides, George K
Koella, Jacob C
and
Basáñez, María-Gloria
2010.
Population biology of malaria within the mosquito: density-dependent processes and potential implications for transmission-blocking interventions.
Malaria Journal,
Vol. 9,
Issue. 1,
Cornet, Stéphane
Gandon, Sylvain
and
Rivero, Ana
2013.
Patterns of phenoloxidase activity in insecticide resistant and susceptible mosquitoes differ between laboratory-selected and wild-caught individuals.
Parasites & Vectors,
Vol. 6,
Issue. 1,
Slater, Hannah C.
Gambhir, Manoj
Parham, Paul E.
Michael, Edwin
and
Antia, Rustom
2013.
Modelling Co-Infection with Malaria and Lymphatic Filariasis.
PLoS Computational Biology,
Vol. 9,
Issue. 6,
p.
e1003096.
McCall, John W.
Hodgkins, Elizabeth
Varloud, Marie
Mansour, Abdelmoneim
and
DiCosty, Utami
2017.
Blocking the transmission of heartworm (Dirofilaria immitis) to mosquitoes (Aedes aegypti) by weekly exposure for one month to microfilaremic dogs treated once topically with dinotefuran-permethrin-pyriproxyfen.
Parasites & Vectors,
Vol. 10,
Issue. S2,
Xu, Zhijing
Graves, Patricia M.
Lau, Colleen L.
Clements, Archie
Geard, Nicholas
and
Glass, Kathryn
2019.
GEOFIL: A spatially-explicit agent-based modelling framework for predicting the long-term transmission dynamics of lymphatic filariasis in American Samoa.
Epidemics,
Vol. 27,
Issue. ,
p.
19.