


Introduction
Continental sediments of upper Campanian (Upper Cretaceous) age are broadly exposed throughout Canada, Mexico, and the United States and have yielded exceptionally rich turtle faunas consisting of adocids, chelydroids, helochelydrids, macrobaenids, nanhsiungchelyids, trionychids, and paracryptodires (see Brinkman, Reference Brinkman2003, Reference Brinkman2005; Brinkman and Rodriguez de la Rosa, Reference Brinkman and Rodriguez de la Rosa2006; Hutchison et al., Reference Hutchison, Knell and Brinkman2013; Sullivan et al., Reference Sullivan, Jasinski and Lucas2013; Lively, Reference Lively2016; and Lyson et al., Reference Lyson, Joyce and Sertich2017 for recent summaries). Soft-shelled turtles (Pan-Trionychidae) are an important component of these faunas, but the majority of finds are fragmentary, making it difficult to rigorously assess their alpha taxonomy and phylogenetic relationships (Vitek and Joyce, Reference Vitek and Joyce2015). The pan-trionychid faunas from the late Campanian of Alberta, New Mexico, and Utah are the best understood to date. The Oldman and Dinosaur Park formations of Alberta, Canada, have yielded particularly rich material, including complete shells and skeletons, that allows recognition of four taxa, the plastomenid Aspideretoides foveatus (Leidy, Reference Leidy1857), the trionychine Axestemys splendidus (Hay, Reference Hay1908), and the indeterminate ‘Trionyx’ allani (Gilmore, Reference Gilmore1923) and ‘Trionyx’ latus (Gilmore, Reference Gilmore1919) (Gardner et al., Reference Gardner, Russell and Brinkman1995; Brinkman, Reference Brinkman2005; Vitek, Reference Vitek2012; Vitek and Joyce, Reference Vitek and Joyce2015). (The pan-trionychid fauna of the Fruitland and Kirtland formations of New Mexico, by contrast, mostly consists of fragments that hint at the presence of at least four trionychids, in particular two unnamed taxa and the indeterminate ‘Trionyx’ austerus (Hay, Reference Hay1908) and ‘Trionyx’ robustus (Gilmore, Reference Gilmore1919) (Sullivan et al., Reference Sullivan, Jasinski and Lucas2013; Vitek and Joyce, Reference Vitek and Joyce2015). The Kaiparowits Formation of neighboring Utah has so far yielded mostly fragmentary remains of four unnamed pan-trionychids with uncertain affinities (Hutchison et al., Reference Hutchison, Knell and Brinkman2013; Vitek and Joyce, Reference Vitek and Joyce2015). Although the turtle faunas of Alberta, New Mexico, and Utah are broadly contemporaneous (Roberts et al., Reference Roberts, Deino and Chan2005, Reference Roberts, Sampson, Deino, Bowring and Buchwaldt2013), clear faunal links are still missing with the exception of an unnamed taxon with a striated carapace reported from the lower Kirtland Formation of northwestern New Mexico (Plastomeninae indet. of Sullivan et al., Reference Sullivan, Jasinski and Lucas2013) and the Kaiparowits Formation of southern Utah (Trionychidae indet. type B of Hutchison et al., Reference Hutchison, Knell and Brinkman2013). The purpose of this contribution is to formally name this taxon as a new species of plastomenid pan-trionychid based on a nearly complete shell from the Fossil Forest Member of the Fruitland Formation of northwestern New Mexico. In addition, we provide an initial assessment of the phylogenetic relationship of the new species and discuss biogeographic and paleoecological implications.
Geologic setting
Vertebrate fossils, including abundant remains of turtles, have been recovered from the upper Fruitland and lower Kirtland formations for more than a century. These remains have traditionally been grouped into the ‘Hunter Wash Local Fauna,’ defined as the fossiliferous horizons in the upper 12.2 meters of the Fruitland Formation and the lower 16.8 m of the Kirtland Formation in the Hunter Wash area (Clemens, Reference Clemens1973; Sullivan and Lucas, Reference Sullivan and Lucas2003, Reference Sullivan and Lucas2006). This definition, however, is dependent on the recognition of a clear interformational contact. Considerable disagreement persists over the nature and stratigraphic location of the contact between the Fruitland and Kirtland formations. As first defined, the Fruitland/Kirtland contact was recognized as gradational, though the Fruitland Formation was observed to be typically sandier than the overlying Kirtland Formation (Bauer, Reference Bauer1916). Later work by Fassett and Hinds (Reference Fassett and Hinds1971) examined the contact in outcrop and subsurface logs across the San Juan Basin and defined the contact as the top of the highest coal or carbonaceous shale. This definition was largely followed by later work (Fassett, Reference Fassett2000, Reference Fassett2010), though the gradational nature of the contact between the lower shale member of the Kirtland formation (‘Hunter Wash Member’ of Hunt and Lucas, Reference Hunt and Lucas1992) and the upper Fruitland Formation make the definition geologically useless, with the only easily definable contact at the base of the overlying Farmington Sandstone Member (‘Farmington Member’ of Hunt and Lucas, Reference Hunt and Lucas1992). Other attempts to define the contact (e.g., Hunt and Lucas, Reference Hunt and Lucas1992; Lucas et al., Reference Lucas, Hunt and Sullivan2006) have placed the top of the Fruitland at the base of the ‘Bisti Member’ or ‘Bisti Bed,’ an indurated but laterally discontinuous sandstone horizon. Regardless of the definition used, the Gilmoremys gettyspherensis type locality (DMNH loc. 5812) is stratigraphically below the Bisti Bed by at least 10 m and falls comfortably within the Fruitland Formation, within the recognized horizons of the Hunter Wash Local Fauna.
The locality is likely bracketed by dateable ash horizons, one in the lower horizons of the Fruitland Formation just above its contact with the underlying Pictured Cliffs sandstone (‘Dog Eye Pond’ [DEP] ash of Fassett and Steiner, Reference Fassett and Steiner1997) and the other in the lower Kirtland Formation (Ash 2 of Fassett and Steiner, Reference Fassett and Steiner1997). The originally published dates for both of these ashes, 75.56 ± 0.41 Ma and 74.56 ± 0.13 Ma, respectively (Fassett and Steiner, Reference Fassett and Steiner1997), have more recently been recalculated using updated standards. Interestingly, these recalculations are in marked disagreement: DEP = 75.16 ± 0.41 Ma and Ash 2 = 74.17 ± 0.13 Ma (Roberts et al., Reference Roberts, Sampson, Deino, Bowring and Buchwaldt2013); DEP = 76.029 ± 0.41 Ma and Ash 2 = 75.023 ± 0.13 Ma (Fowler, Reference Fowler2017). Refining the geochronology of the Fruitland Formation through identification and dating of new ash beds, and redating known ash beds, is imperative for the comparison of the Hunter Wash Local Fauna with other Western Interior Basin faunas. Regardless of the precise date, all published and recalculated ages firmly place the type locality in the late Campanian.
The holotype specimen was recovered from a blocky, carbonaceous mudstone horizon likely representing a ponded, overbank depositional environment located in San Juan County, New Mexico, USA (Fig. 1). Disarticulation of the carapace and plastron, accompanied by the vertical displacement and irregular orientation of recovered elements at the site, suggests soft sediment bioturbation typical of shallow pond or slow water environments on a fluvial floodplain. It is likely the postmortem remains settled to the bottom of a low-energy, shallow-water environment and experienced in situ disarticulation with negligible transport.

Figure 1 Location of DMNH loc. 5812, type locality of Gilmoremys gettyspherensis n. sp. (1) Overview of surface exposures (green) of the upper Campanian Fruitland and Kirtland formations in the San Juan Basin of northwestern New Mexico; (2) approximate placement of DMNH loc. 5812 (star) in the upper horizons of the Fruitland Formation in the North Escavada Study Area.
Materials and methods
All materials used herein were collected over the course of the past decades by independent teams from several institutions from Upper Cretaceous (Campanian) sediments exposed in New Mexico and Utah. Previously published materials from New Mexico (Sullivan et al., Reference Sullivan, Jasinski and Lucas2013) and Utah (Hutchison et al., Reference Hutchison, Knell and Brinkman2013) are housed at the State Museum of Pennsylvania and Utah Museum of Natural History, respectively. New material presented herein is housed at the Denver Museum of Nature and Science.
Repositories and institutional abbreviations
Types, figured, and other specimens examined in this study are deposited in the following institutions: Denver Museum of Nature and Science (DMNH), Denver, Colorado, USA; State Museum of Pennsylvania (SMP), Harrisburg, Pennsylvania, USA; Utah Museum of Natural History (UMNH), Salt Lake City, Utah, USA.
Systematic paleontology
Testudinata Batsch, Reference Batsch1788, sensu Joyce et al., Reference Joyce, Parham and Gauthier2004
Trionychidae Gray, Reference Gray1825, sensu Joyce et al., Reference Joyce, Parham and Gauthier2004
Gilmoremys Joyce and Lyson, Reference Joyce and Lyson2011
Type species
Aspideretes lancensis Gilmore, Reference Gilmore1916.
Gilmoremys gettyspherensis new species

Figure 2 DMNH EPV.125905, Gilmoremys gettyspherensis, holotype, Late Cretaceous (late Campanian) of New Mexico. (1) Photograph and illustration of carapace in dorsal view; (2) photograph and illustration of carapace in ventral view. co = costal bone.
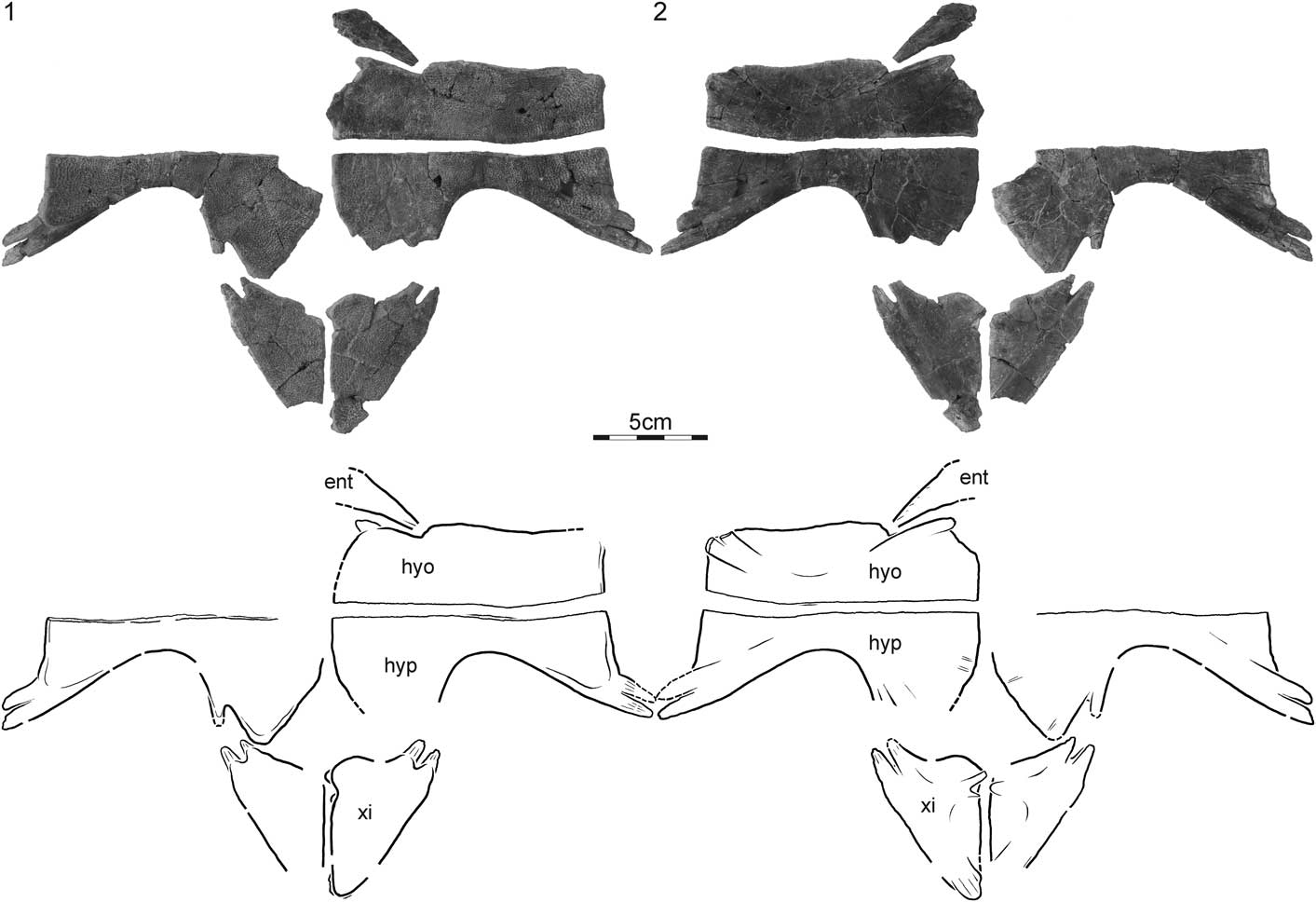
Figure 3 DMNH EPV.125905, Gilmoremys gettyspherensis, holotype, Late Cretaceous (late Campanian) of New Mexico. (1) Photograph and illustration of plastron in ventral view; (2) photograph and illustration of carapace in dorsal view. ent = entoplastron; hyo = hyoplastron; hyp = hypoplastron; xi = xiphiplastron.
Holotype
DMNH EPV.125905, a partial, disarticulated shell consisting of right costals I–III, V, VI, and VIII, left costals V, VII, and VIII, the left half of the entoplastron, the right hypo- and xiphiplastron, and the left hyo-, hypo-, and xiphiplastron (Figs. 2, 3); DMNH loc. 5812, North Escavada Study Area, San Juan County, New Mexico, USA (Fig. 1); Fossil Forest Member, Fruitland Formation, late Campanian, Late Cretaceous.
Diagnosis
Gilmoremys gettyspherensis n. sp. can be diagnosed as a representative of Pan-Trionychidae, among other characters, by the absence of peripherals, pygals, and scutes, and as a representative of Gilmoremys by a relatively thin shell; carapacial sculpturing consisting of fine pits combined with extended sinusoidal ridges or grooves that cross the length of several carapacial elements; costal ribs that protrude beyond the margin of the carapacial disk; the inferred presence of a preneural; a distally constricted costal I and distally expanded costal II; two lateral hyoplastral processes; low hyoplastral shoulders (i.e., anteriorly protruding lappets); and full midline contact of the elongate xiphiplastra. Gilmoremys gettyspherensis n. sp. can be differentiated from Gilmoremys lancensis by smaller size; the presence of raised sinusoidal ridges, not grooves; less distally expanded costal II; and less elongate xiphiplastra.
Occurrence
San Juan County, New Mexico, USA, Fossil Forest Member, Fruitland Formation, late Campanian, Late Cretaceous (type occurrence); San Juan County, New Mexico, USA, Hunter Wash Member, Kirtland Formation, late Campanian, Late Cretaceous (Plastomeninae indet. of Sullivan et al., Reference Sullivan, Jasinski and Lucas2013, fig. 20.17); Kane and Garfield County, Utah, USA, Kaiparowits Formation, late Campanian, Late Cretaceous (Trionychidae type B of Hutchison et al., Reference Hutchison, Knell and Brinkman2013, fig. 13.17).
Description
The carapace of DMNH EPV.125905, the type specimen, includes right costals I–III, V, VI, and VIII, left costals V, VII, and VIII (Fig. 2). All elements with the exception of the nuchal, the preneural, the neurals, and costals IV are therefore present. The available elements indicate that the outline of the carapace was rounded, with the exception of a broad but shallow pygal notch formed by costals VIII. Although the costal rib is only completely preserved for right costal VI, well-developed breaks along the distal ends of all other preserved costals indicate that all costal ribs would have protruded significantly beyond the margins of the carapacial disk. We estimate the carapacial disk to have been about 22.5 cm along the midline, including the nuchal. The costals vary from about 3.5 mm in thickness toward the midline to about 4.5 mm toward their margins, not including the ribs. The margins of the costals are rounded and show no evidence for ‘splitting’ (i.e., separation of surficial and deepest layers of the costal by a trough). The carapace is primarily decorated by a surface pattern consisting of blunt, circular depressions that fade toward the midline. In addition, the carapace is decorated by a series of sinuous ridges that are arranged parallel to the midline and span the length of three to four costals. The ridges are faint and few on costals I, but they become increasingly dense and distinct toward the back of the carapace.
The nuchal is not preserved, but its preserved sutural attachment site with right costal I suggests that the metaplastic portion of the nuchal formed a callosity about half the width of the carapacial disk and that suprascapular fontanelles were absent. The neural series, including the preneural, is not preserved in DMNH EPV.125905, but the medial margins of the available costals provide enough information to allow reconstruction of the most important aspects. The presence of two facets along the medial edge of costal I indicate the presence of a preneural, which was shorter than neural I. The width of this bone is unknown. Costals II, V, and VI have two facets as well, of which the anterior is shorter than the posterior one. This allows the inference that neurals I–VI were hexagonal and had short posterior sides. The medial margins of costal VII are unfortunately damaged and it is therefore unclear whether neural VII was square (i.e., laterally only contacted costals VII) or hexagonal with short posterior sides. The anteromedial margins on both costals VIII clearly hint at the former presence of a small neural VIII. There is an apparent gap between the anterior three and posterior four available costals, and DMNH EPV.125905 can therefore be reconstructed with confidence to have had eight pairs of costals. Costals I are slightly bowed to the anterior and narrower distally than proximally. Costals II mediate between the anteriorly oriented costals I and the straight costals III and are therefore expanded distally. Costals III, V, VI, and likely costal IV are similar in proportions, expand only slightly distally, and mediate between the anteriorly directly anterior costals and the posteriorly oriented posterior costals. Costals VII are clearly directly posterolaterally, have similar anteroposterior dimensions to the more anterior costals, but are noticeably shorter. Costals VIII are slightly broader than long, have the greatest anteroposterior dimensions of the costal series, contact one another along the midline for much of their anteroposterior length, and frame the shallow pygal notch.
No thoracic vertebrae are preserved in DMNH EPV.125905. The ribs of all available costals are greatly eroded, with the exception of right costal VI, which still preserves much of the distal end of its rib. It appears all but certain by reference to the available scars that all costal ribs extended significantly beyond the margin of the carapacial disk. Whereas costal rib VII clearly did not underlap costal VI, costal rib VIII may have underlapped costal VII.
The plastron of DMNH EPV.125905 includes the left half of the entoplastron, the right hypo- and xiphiplastron, and the left hyo-, hypo-, and xiphiplastron (Fig. 3). The entoplastron is greatly damaged medially, and it is therefore not possible to assess the original shape of this bone with confidence. We here nevertheless conclude by reference to other trionychids with an entoplastral callosity that enough is preserved to infer that such a callosity is absent.
Much of the hyo-, hypo, and xiphiplastra are covered by broad callosities that are decorated with a fine pattern of low, raised tubercles that are particularly closely packed near the inguinal buttresses, but otherwise coalesce in groups of twos or threes. The strap-like processes that form the deep tissue component of these bones are fully exposed along the lateral margins of the hyo- and hypoplastron but are otherwise partially to fully covered by metaplastic bone. Whereas it is apparent that the xiphiplastra contacted one another along their full anteroposterior length, the hyo- and hypoplastra at best had a blunt midline contact.
The lateral processes of the only available hyoplastron are damaged, but the cross section of the break indicates that two processes were present. A single, medial process is apparent along the anteromedial margin of the hyoplastron that protrudes just beyond the margin of the metaplastic bone. The hyoplastral callosity is otherwise rectangular in shape and shows a modest anterior lappet that lightly embraces the entoplastron. The hypoplastra have two well-developed lateral processes and two faintly developed posterior processes for articulation with the xiphiplastron, but we can only discern faint traces of fine, medial processes. The hypoplastra form a triangular median fontanelle. The xiphiplastra have two anterior processes that interfinger with the hypoplastral processes and a single posteromedial process. The left xiphiplastron furthermore has two anteromedial processes that interfinger with a single anteromedial process of the right xiphiplastron.
Etymology
In reference to the ‘Gettysphere,’ a circle of positive energy that surrounded Mike Getty, finder of the type specimen and recently deceased paleontologist, preparator, friend, and human being extraordinaire. Though figurative, the Gettysphere was a place, and the species epithet is therefore formed with the ending ‘-ensis.’
Phylogenetic analysis
We explore the phylogenetic relationships of Gilmoremys gettyspherensis n. sp. by incorporating it into the character/taxon matrix of Joyce and Lyson (Reference Joyce and Lyson2017), which in turn is based on the successive phylogenetic analyses of Meylan (Reference Meylan1987), Joyce et al. (Reference Joyce, Revan, Lyson and Danilov2009, Reference Joyce, Lyson and Williams2016), Joyce and Lyson (Reference Joyce and Lyson2010, Reference Joyce and Lyson2011), and Brinkman et al. (Reference Brinkman, Rabi and Zhao2017). To help resolve relationships, we added a single character (character 95) to the matrix that encodes the absence (0) or presence (1) of well-developed anteroposterior striation in the carapace of adults. The final data file, which includes the character/taxon matrix consisting of 95 characters scored for 37 taxa, the full list of characters and character states, and the backbone constraint, is provided in Supplemental data set 1.
We performed a parsimony analysis using the software TNT (Goloboff et al., Reference Goloboff, Farris and Nixon2008). Characters 1, 3, 5, 14, 20, 22, 41, 54, 79, 81, and 94 form morphoclines and were therefore ordered. Whereas the relationships of all extant trionychids were constrained using the molecular topology of Le et al. (Reference Le, Duong, Dinh, Nguyen, Pritchard and McCormack2014), all fossil trionychids were allowed to float. Following the recommendations of Goloboff et al. (Reference Goloboff, Torres and Arias2017), characters were lightly weighted using a k factor of 12. The matrix was subjected to 1,000 replicates of random addition sequences followed by a second round of tree bisection-reconnection. The strict consensus tree resulting from the six equally most parsimonious trees with a score of 12.79441 is provided in Figure 4. The full strict consensus tree, common synapomorphies, and Bremer support values are provided in Supplemental data set 2.

Figure 4 Time-calibrated strict consensus cladograms retrieved from the phylogenetic analysis performed herein. The full tree is provided in Supplementary data set 2.
Discussion
Alpha taxonomy
Although a total of 14 fossil trionychid species have been named from Campanian sediments exposed throughout North America, only six are currently recognized as valid (Gardner et al., Reference Gardner, Russell and Brinkman1995; Vitek, Reference Vitek2012; Sullivan et al., Reference Sullivan, Jasinski and Lucas2013; Vitek and Joyce, Reference Vitek and Joyce2015): Aspideretoides foveatus (Leidy, Reference Leidy1857), Axestemys splendidus (Hay, Reference Hay1908), ‘Trionyx’ allani (Gilmore, Reference Gilmore1923), ‘Trionyx’ austerus (Hay, Reference Hay1908), ‘Trionyx’ latus (Gilmore, Reference Gilmore1919), and ‘Trionyx’ robustus (Gilmore Reference Gilmore1919).
Gilmoremys gettyspherensis n. sp. resembles Aspideretoides foveatus (as described by Gardner et al., Reference Gardner, Russell and Brinkman1995; Brinkman, Reference Brinkman2005) in overall size, outline of the carapace, and presence of a preneural, but Aspideretoides foveatus differs, among other characters, by being thicker shelled and by possessing a carapacial surface texture consisting of coarse pits that fade toward the midline, fewer neurals, smaller costals VIII, a broader entoplastron, only a single lateral hyoplastral process, and only poorly developed xiphiplastral callosities.
Gilmoremys gettyspherensis n. sp. resembles Axestemys splendidus (as described by Gardner et al., Reference Gardner, Russell and Brinkman1995) in carapacial outline, presence of a preneural, proportions of costals VIII, and the overall arrangement of the hyo-, hypo, and xiphiplastra. However, Axestemys splendidus differs by being about twice as large, by being thicker shelled, by having a coarsely netted carapacial surface ornamentation, fewer neurals, and less elongated xiphiplastra, and by lacking hyoplastral shoulders.
Gilmoremys gettyspherensis n. sp. resembles ‘Trionyx’ allani (as described by Gardner et al., Reference Gardner, Russell and Brinkman1995) in the overall arrangement of the carapace, including presence of a preneural, but ‘Trionyx’ allani differs substantially by being about twice as large, having a coarsely netted carapacial sculpturing, possessing open suprascapular fontanelles, and exhibiting substantially smaller plastral callosities that lack a surface ornamentation.
Gilmoremys gettyspherensis n. sp. resembles ‘Trionyx’ latus (as described by Gardner et al., Reference Gardner, Russell and Brinkman1995) in overall size and number of neural elements, but ‘Trionyx’ latus differs by lacking a sutural contact between the nuchal and the remaining carapacial disk, exhibiting well-developed suprascapular fontanelles, lacking a preneural, possessing a finely knit carapacial surface sculpturing that is not developed along the margins of the shell, and having significantly smaller costals VII and VIII.
Gilmoremys gettyspherensis n. sp. resembles ‘Trionyx’ robustus (as described by Gilmore, Reference Gilmore1919) in overall carapace outline, presence of a preneural, and proportions of costals VIII, but ‘Trionyx’ robustus differs by being about twice as large, being thick shelled, having a carapacial surface sculpturing consisting of deep, rounded pits, and having much better developed hyo/hypoplastral callosities that fully cover the medial and lateral processes.
The morphology of ‘Trionyx’ austerus (as described by Hay, Reference Hay1908) is still poorly characterized, but this taxon can nevertheless be easily distinguished from Gilmoremys gettyspherensis n. sp. by being much larger and extremely thick shelled.
These comparisons highlight that Gilmoremys gettyspherensis n. sp. can be distinguished easily from all previously named Campanian taxa from North America.
Although up to eight fossil trionychids may compose the Maastrichtian trionychid assemblage of North America (Holroyd et al., Reference Holroyd, Wilson and Hutchison2014), only six are currently recognized by name (Vitek and Joyce, Reference Vitek and Joyce2015): Axestemys splendida (Hay, Reference Hay1908), Gilmoremys lancensis (Gilmore, Reference Gilmore1916), Helopanoplia distincta Hay, Reference Hay1908, Hutchemys sterea (Hutchison, Reference Hutchison2009), Hutchemys tetanetron (Hutchison, Reference Hutchison2009), and ‘Trionyx’ beecheri Hay, Reference Hay1904. The two named Maastrichtian species of Hutchemys (as described by Hutchison, Reference Hutchison2009) are easily distinguished from Gilmoremys gettyspherensis n. sp., among others, by being smaller and thicker shelled, by exhibiting a pitted shell sculpturing, and, most significantly, by possessing well-developed hyoplastral shoulders that suturally articulate with well-developed entoplastral callosities. Helopanoplia distincta (as described by Joyce and Lyson, Reference Joyce and Lyson2017) most readily differs from Gilmoremys gettyspherensis n. sp. by being larger, by lacking free rib ends for costal ribs I–VI, by exhibiting a carapacial and plastral surface sculpturing that ranges from netting to isolated tubercles, by only possessing only a single lateral hyoplastral process and much more broadly developed hyo-, hypo-, and xiphiplastral callosities. Differences with Axestemys splendida are already listed. ‘Trionyx’ beecheri (as described by Hay, Reference Hay1908; personal observation of the type specimen) finally differs by possessing a carapacial surface texture consisting of fine netting, a plastral texture consisting of fine raised tubercles, and large hyo/hypoplastral callosities that cover the lateral processes completely.
Gilmoremys gettyspherensis n. sp. broadly overlaps in morphology with Gilmoremys lancensis (as described by Joyce and Lyson, Reference Joyce and Lyson2011): both species have a relatively thin shell, carapacial sculpturing consisting of fine pits combined with extended sinusoidal ridges or grooves, costal ribs that protrude beyond the margin of the carapacial disk, a preneural, distally constricted costal I and distally expanded costal II, two lateral hyoplastral processes, only slightly developed hyoplastral shoulders, and full midline contact of the elongate xiphiplastra. It is therefore not surprising that our phylogenetic analysis retrieves them as immediate sisters (see the following). A small number of systematic differences are nevertheless apparent: the Maastrichtian material is consistently larger, costals II are more expanded distally, the xiphiplastra are more elongated, and most significantly, the sinusoidal ornamentation is developed in the form of grooves, not elevated ridges. We find the available character evidence to be sufficient to establish a new species for the Campanian morphotype that we herein name Gilmoremys gettyspherensis.
The comparisons made in the preceding highlight that Gilmoremys gettyspherensis n. sp. is the only known named species from the Late Cretaceous of North America to have surface sculpturing consisting of raised ridges. The only other previously reported material with such ridges has been reported from the Campanian of New Mexico (Plastomeninae indet. of Sullivan et al., Reference Sullivan, Jasinski and Lucas2013) and Utah (Trionychidae indet. type B of Hutchison et al., Reference Hutchison, Knell and Brinkman2013). The most informative figured specimen from Utah is a left costal VIII (Hutchison et al., Reference Hutchison, Knell and Brinkman2013, fig. 13.17c) that corresponds fully with that of the holotype of Gilmoremys gettyspherensis n. sp. by being wider than long and exhibiting well-developed raised ridges. The previously described specimen from New Mexico, recovered from the lowermost horizons of the Kirtland Formation (Hunter Wash Local Fauna) is a partial shell. The preserved costals once again exhibit well-developed raised ridges (Sullivan et al., Reference Sullivan, Jasinski and Lucas2013, fig. 20.17a, b). The preserved portion of the hyo/hypoplastra (Sullivan et al., Reference Sullivan, Jasinski and Lucas2013, fig. 20.17c, d) are consistent with the holotype of Gilmoremys gettyspherensis n. sp. in regard to sculpturing, but the hyoplastral shoulder is better developed. As this specimen appears to be slightly larger than the holotype, this difference can be attributed to a greater ontogenetic age. We therefore refer the previously described material from New Mexico and Utah to Gilmoremys gettyspherensis n. sp. with confidence.
Phylogenetic relationships
Our phylogenetic analysis resulted in six equally parsimonious trees. The relationships among plastomenid soft-shelled turtles are fully resolved in the strict consensus tree (Fig. 3), highlighting that the six equally parsimonious trees did not reveal any topological conflict within this clade. The topology generally resembles those of previous analyses (e.g., Joyce and Lyson, Reference Joyce and Lyson2011, Reference Joyce and Lyson2017; Joyce et al., Reference Joyce, Lyson and Williams2016) with exception of a more derived placement of Atoposemys superstes (Russell, Reference Russell1930) as the sister to the Helopanoplia/Hutchemys clade and placement of Gilmoremys gettyspherensis n. sp. as sister to Gilmoremys lancensis. A literal interpretation of the tree implies that at least three plastomenid taxa existed during the Campanian: Aspideretes foveatus, Gilmoremys gettyspherensis, as the ancestral lineage of the clade formed by Atoposemys superstes, Helopanoplia distincta, Hutchemys spp., and Plastomenus thomasii (Cope, Reference Cope1872). This number is certainly too conservative as an additional, unnamed plastomenid has been reported from the late Campanian of Utah (Helopanoplia sp. of Hutchison et al., Reference Hutchison, Knell and Brinkman2013) and as some previously described species from the Campanian of North American exhibit plastomenid characteristics, in particular ‘Trionyx’ (originally Plastomenus) robustus (Gilmore, Reference Gilmore1919). We therefore infer with confidence that the group originated prior to the late Campanian.
The hypothesized sister group relationship between the Campanian Gilmoremys gettyspherensis n. sp. and the Maastrichtian Gilmoremys lancensis is not surprising, as the morphology of these two species corresponds broadly. As the Campanian morphotype does not exhibit any characters that must be inferred to be apomorphic, it is possible to conclude that it may represent the ancestral morphology of the Maastrichtian morphotype and that the two form a single, persistent anagenetic lineage. This assertion can be tested in the future by finding both morphotypes in contemporaneous sediments.
It is relatively easy to establish the presence of Gilmoremys gettyspherensis n. sp. in a fauna, as the shells of adults are decorated by sinusoidal ridges. Although the study of turtle remains from the Campanian of New Mexico and Utah is still in its infancy, we here document multiple specimens referable to Gilmoremys gettyspherensis n. sp. Although the fossil turtle faunas of more northern basins of the Western Interior, in particularly those in Alberta and Montana, have been sampled heavily and are much better understood (see Brinkman, Reference Brinkman2005 for a summary), we are unaware of any fragments of Gilmoremys gettyspherensis n. sp. reported from those localities. We therefore conclude that this taxon was restricted to the southern basins during the late Campanian.
The majority of plastomenid remains are recovered from siltstone and mudstones and the group is therefore often believed to have preferred ponded environments (e.g., Joyce and Lyson, Reference Joyce and Lyson2017). The discovery of the type specimen of Gilmoremys gettyspherensis n. sp. in a mudstone once again confirms this conclusion.
Acknowledgments
We thank the late M. Getty (DMNH) for finding and collecting the holotype specimen. Special thanks are due to the late D. McCuan, DMNH volunteer, for preparation of the fossil. Our fieldwork in the San Juan Basin has been generously supported by several anonymous donors to the DMNH Laramidia Project. Immense gratitude is extended to the large volunteer corps at DMNH, who contributed thousands of hours of work in the field, laboratories, and collections resulting, among others, in this publication. Thanks also to BLM Regional Paleontologist P. Gensler and BLM Field Office Paleontologist S. Landon for their logistical support. The holotype specimen described herein was collected under BLM permit NM14-04S. P. Meylan is thanked for providing thoughtful comments that helped improve the quality of the manuscript.
Accessibility of supplemental data
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.7h9p04r.




