


Introduction
Lichen species often produce more than one colour morph. Cyanomorphs and chloromorphs in Lobaria (e.g. Jørgensen Reference Jørgensen1998) are an example where even one individual mycelium produces two colour variants. Several examples are also known of non-pigmented phenotypes that occur rarely, but randomly, throughout normally pigmented populations. These include: specimens with unpigmented ascomata in species with normally dark fruit bodies (Gilbert Reference Gilbert1996); specimens with unpigmented thalli in for example, Caloplaca verruculifera (Vain.) Zahlbr. (Søchting Reference Søchting1973) and Arctoparmelia centrifuga (L.) Hale (Santesson Reference Santesson1970); and the citrine-green morphotypes in various species of Candelariella (Gilbert et al. Reference Gilbert, Henderson and James1981). Such variation may be due to occasional mutations blocking specific metabolic pathways. On the other hand, greyish thalli of normally yellow Xanthoria species are usually considered to be ecologically induced phenotypic expressions occurring in shaded and wet populations.
Here, we present an example of two sorediate ‘species’ of the genus Caloplaca (Teloschistaceae) recognized because of the different colours, grey versus yellow, of their thalli.
Caloplaca phlogina (Ach.) Flagey
This is a yellow, sorediate morphotype containing anthraquinones (Fig. 1A) that grows on bark (rarely on concrete). It is known from Scandinavia (Arup Reference Arup2006), Canada (Richardson et al. Reference Richardson, Lucas and Anderson2009; specimen confirmed by U. Søchting) and Romania (Vondrák et al. Reference Vondrák, Říha, Arup and Søchting2009). Records not confirmed by molecular data are also known from Western and Central Europe (Crespo et al. Reference Crespo, Barreno, Rico and Bueno1980; Sérusiaux et al. Reference Sérusiaux, Diederich, Brand and van den Boom1999; Vondrák et al. Reference Vondrák, Kocourková, Palice and Liška2007; Fletcher & Laundon Reference Fletcher, Laundon, Smith, Aptroot, Coppins, Fletcher, Gilbert, James and Wolseley2009), and India (Joshi & Upreti Reference Joshi and Upreti2008).
Caloplaca scythica Khodosovtsev & Søchting
This is a whitish, or white-green, sorediate morphotype lacking anthraquinones in the thallus (Fig. 1B) that grows on the maritime shrubs Halocnemum strobilaceum and Limonium suffruticosum, plant debris and loess soil on the northern coast of the Black Sea (Kondratyuk et al. Reference Kondratyuk, Søchting, Khodosovtsev and Kärnefelt1998; Vondrák et al. Reference Vondrák, Říha, Arup and Søchting2009a, sub C. phlogina). It also occurs on tree bark in Scandinavia, where it often grows side by side with yellow-sorediate morphotypes. Surprisingly, this morphotype was recently collected from a weathered piece of leather at a maritime site in southern Chile (Isla Navarino, Zeuthen Søgaard 84, C). Its ITS sequence was nearly identical to those of C. scythica (Fig. 2).

Fig. 1. Caloplaca phlogina, thallus and apothecia. A, C. phlogina type with anthraquinones in the thallus [Denmark, Nekselø, Søchting 11263 (C)]; B, C. scythica type (Russia, Black Sea coast, CBFS JV7059). Scales: A & B = 1 mm.
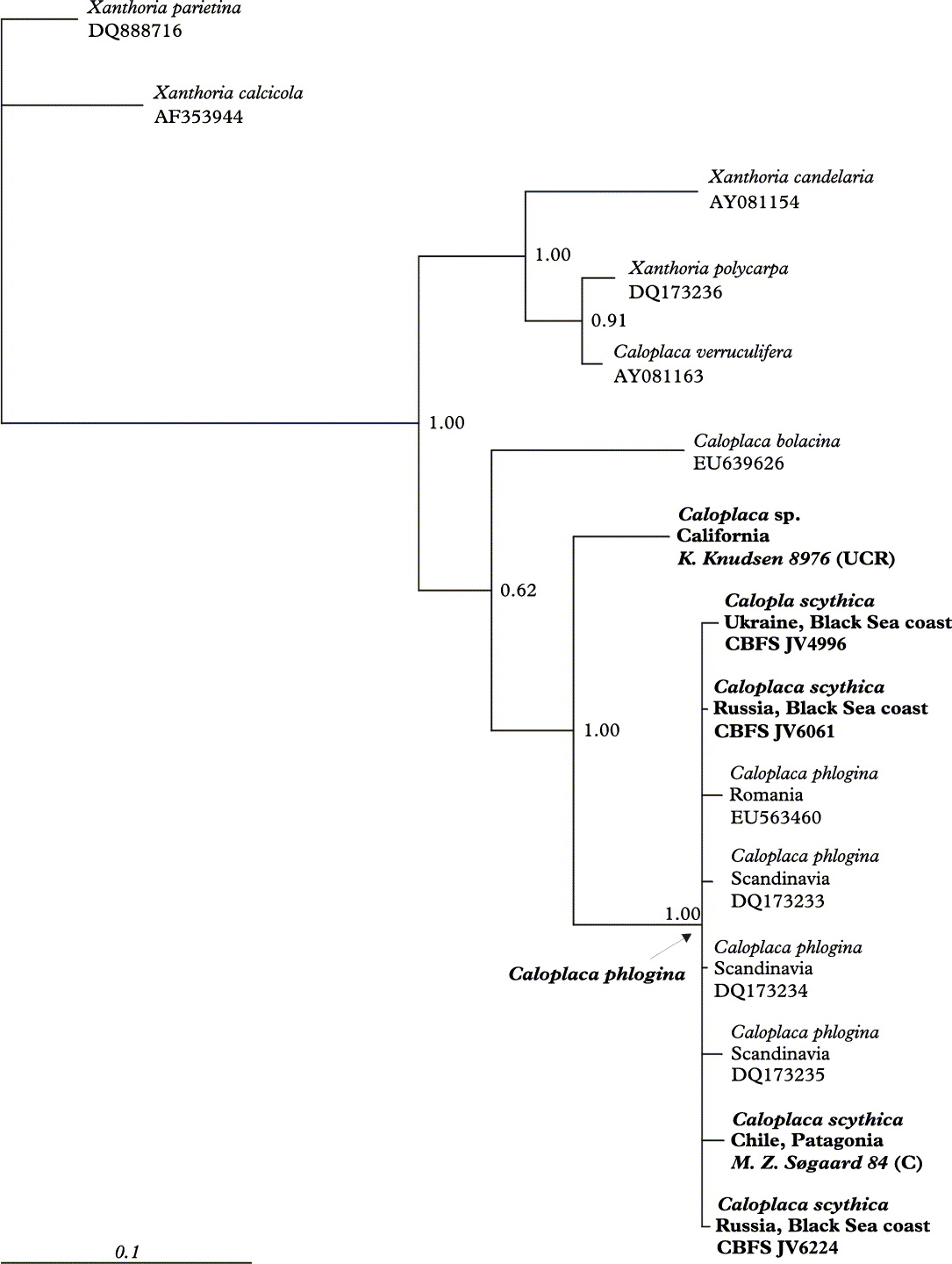
Fig. 2. Bayesian consensus phylogeny of the clade including Caloplaca phlogina. Node support values are Bayesian posterior probabilities. Newly obtained ITS sequences are in bold.
Our aim was to investigate the status of the Caloplaca morphotypes using phenotype studies and phylogenetic analysis of the ITS regions of nuclear rDNA.
Materials and Methods
Specimens studied
Morphotypes with yellow soralia. Denmark: Bornholm: Arnager, on bark, 2006, R. S. Larsen & J. Vondrák (CBFS JV4713). Jutland: Ranum, on bark, T. Jensen (LD 1081648). Sjælland: Haraldsted, 1946, O. Almborn (LD 1081712). Sjælland: Kildebrønde, on wood, 1943, M. Skytte-Christiansen (LD 1081584).— France: Seine-et-Marne: Fontainebleau, on bark, 1949, O. Almborn (LD 1094801).—Germany: Oldenburg, on wood, 1889, H. Sandstede (LD 1093457).—Romania: Black Sea coast: Constanta, Mangalia, on concrete, 2007, J. Vondrák (CBFS JV3437).— Sweden: Blekinge, Ringamåla, on bark, 1989, U. Arup & S. Ekman (LD Reference Arup1025279). Halland: Släp, on bark, 1989, U. Arup & S. Ekman (LD 1025983); Närke, Götlunda, Sickelsjö, on bark, 1952, G. Kjellmert (LD 104335). Skåne: Bollerup, on bark, 1992, L. Lindblom (LD 1068814); Hallands Väderö, 1990, L. Lindblom (LD 1068878); Kåseberga, on bark, 1997, P. Frödén (LD 1023711) Vittskövle, on bark, 1943, O. Almborn (LD 1067982).
Morphotypes with soralia lacking anthraquinones. Chile: Isla Navarino: Puerto Williams, on weathered leather, 2008, M. Z. Søgaard (C).—Russia: Black Sea coast: Taman Peninsula, on loess and maritime shrubs, 2007, J. Vondrák (CBFS JV6060, Reference Arup6061, 6224, 7385, 7500).—Sweden: Skåne: on bark, 1942, O. Almborn (LD 1067918); ibid., on bark, 1964, S.W. Sundell (LD 1068110, anthraquinones present in spots); Kristianstad, on bark, 1935, O. Almborn (LD 1024223, 1068942); Kyrkheddinge, on bark, 2002, D. Göransson (LD 1262223).—Ukraine: Crimea: Syvash salt lagoon, on maritime shrubs, 2006 & 2009, J. Vondrák (CBFS JV4648, Reference Arup4653, 4996, 7209); ibid.: 2003, A. Khodosovtsev (KHER 2918). Kherson region: Sadove, Nikolaivka, on maritime shrubs, 2009, J. Vondrák (CBFS JV7143). Nikolaev region: Kinburnskaya kosa, on maritime shrubs, 2009, J. Vondrák (CBFS JV7132, Reference Arup7133).
Samples with both morphotypes: Sweden: Halland: Vallda, on bark, 1932, A. H. Magnusson (Magnusson: Lich. sel. scand. exs. 143; LD 1067470). Skåne: Ivotofta, 1942, O. Almborn (LD 1023967, 1067854); Mölleberga, on bark, 1938, O. Almborn (LD 1023903); Kristineberg, on bark, 1995, P. Frödén (LD 1023775); Träne, on bark, 1946, O. Almborn (LD 1024031). Södermanland: Trosa, on maritime wood, 1925, G. O. A. Malme (LD 1015744). Västmanland: Norlund, on wood, 1950, G. Kjellmert (LD 1015552).
DNA extraction and amplification
Direct PCR was used for PCR-amplification of the ITS regions including the 5.8S gene of the nuclear rDNA following Arup (Reference Arup2006). Primers for amplification were ITS1F (Gardes & Bruns Reference Gardes and Bruns1993) and ITS4 (White et al. Reference White, Bruns, Lee, Taylor, Innis, Gelfand, Sninsky and White1990). PCR cycling parameters followed Ekman (Reference Ekman2001).
Phylogenetic analysis
Five newly obtained ITS sequences (Table 1) were included in the phylogenetic analysis along with ten sequences from GenBank selected to illustrate the phylogenetic position of Caloplaca scythica in relation to the Caloplaca phlogina clade. Sequences were aligned using MAFFT 6 (on-line version in the Q-INS-i mode; see Katoh et al. Reference Katoh, Kuma, Toh and Miyata2002) and manually cut to eliminate the unaligned ends and ambiguously aligned regions of ITS1 and ITS2; 506 positions were retained. Bayesian phylogenetic analysis was carried out using the program MrBayes 3.1.1 (Ronquist & Huelsenbeck Reference Ronquist and Huelsenbeck2003). The optimal nucleotide substitution model (HKY+G) was found using the program MrModeltest v2.3 (Nylander Reference Nylander2004) with the Akaike Information Criterion and the hierarchical likelihood ratio test (Posada & Crandall Reference Posada and Crandall1998). The MCMC analysis was run for five million generations, performed in two runs, each with four chains starting from a random tree and using the default temperature of 0·2. Every 100th tree was sampled, and the first 174 000 generations were discarded as burn-in, using standard deviation of splits between runs less than 0·01 as a convergence criterion.
Table 1. Voucher specimens and GenBank accession numbers of the new ITS sequences used in the phylogenetic analysis.

Phenotype investigations were made according to Vondrák et al. (Reference Vondrák, Říha, Arup and Søchting2009) and the phenotypic characters selected for the study are listed in Table 2. Anthraquinone content was analyzed in one selected sample by HPLC analysis (for details see Søchting Reference Søchting1997).
Table 2. Phenotypic data for both colour variants of Caloplaca phlogina; literature data and our data (bold) are shown.

* no. of measurements
Results
The Bayesian phylogenetic analysis of the ITS nrDNA region revealed that bothphenotypic variants together form one well-supported phylogenetic clade (PP =1·0) without any internal diversification (Fig. 2). Only 10 variable positions (of 506) occurred within this clade. The closest relative we found is a North American species resembling C. pyracea (USA: California: Channel Islands, on Eucalyptus bark, 2007, K. Knudsen 8976, UCR, dupl. in CBFS), but which is not related to C. pyracea (Ach.) Th. Fr. s. str.
Phenotypic appraisals did not reveal any significant differences between the two variants (Table 2). Both morphotypes are very similar in the 13 characters examined; the colour of their soralia remains the only difference.
However, there are marked differences in the ecological and distributional pattern of the two variants. They grow together on tree bark in southern Scandinavia, but the white-green sorediate morphotype is probably less common there. In contrast, on shrubs in salt marshes on the Black Sea coast, the yellow sorediate morphotype is exceptionally rare, whereas the grey one is very common.
Based on both molecular and phenotypic data, we consider both morphotypes to be conspecific and we propose to reduce Caloplaca scythica into synonymy with C. phlogina.
Discussion
The molecular data show Caloplaca phlogina to be rather closely related to the morphologically very dissimilar C. bolacina (Tuck.) Herre, whereas it is morphologically ex-tremely similar to several species from the unrelated Caloplaca citrina group. For example, Caloplaca citrina (Hoffm.) Th. Fr. and C. limonia Nimis & Poelt are sometimes morphologically indistinguishable from C. phlogina. Ecology can help us to determine the species and we can achieve almost 100 % certainty in identification of C. phlogina in its ‘typical’ habitats: bark of broad-leaved trees in southern Sweden or shrubs in salt marshes at the Black Sea coast. Samples from other habitats (e.g., concrete) may be confused with other species and we are sceptical about some records not confirmed by molecular data (e.g. Crespo et al. Reference Crespo, Barreno, Rico and Bueno1980; Vondrák et al. Reference Vondrák, Kocourková, Palice and Liška2007).
Caloplaca phlogina – a species with two different ecologies
In Scandinavia, C. phlogina mainly grows inland, has no clear tendency to be maritime (Arup Reference Arup2006), and grows mainly on tree trunks, not twigs and branches. Its distribution and ecology elsewhere in Europe are not well known, but it is probable that there are other, scattered, non-maritime populations. In contrast, the Black Sea populations are strongly maritime and grow mainly on twigs of shrubs in salt marshes. It might be justified to regard the Black Sea population with white soralia as distinct from the Scandinavian one at an infra-specific level, but we prefer to wait for convincing molecular data before making any taxonomic proposals.
Founder effects in distant populations
The distant maritime populations of C. phlogina may have originated from one or a few diaspores. This hypothesis is based mainly on the observations that the species is ecologically and phenotypically uniform in maritime conditions where the yellow-sorediate morphotype is extremely rare and with a low anthraquinone content. We consider the Black Sea populations to be established through distribution from some European population where the species grows on a variety of substrata, in both phenotypic variants and even inland or along sea shores. Even though the distribution of C. phlogina in South America is poorly known, we also suspect that the only known Chilean record is a result of young distribution from the source populations in Europe.
The examples listed in the introduction concern phenotypes which appear to have lost their ability to produce anthraquinones or other pigments. This phenomenon, which may be due to a single mutation somewhere in the synthesis pathway, differs from the situation in C. phlogina. Here, in the grey thallus phenotype, the anthraquinones are still produced in the apothecia but the synthesis is not expressed in the soralia.
Kerry Knudsen and Alexander Khodosovtsev kindly provided their lichen material for this study. Our research was supported by the Grant Agency of the Academy of Sciences of the Czech Republic (KJB 601410701). B. Abbot kindly improved the English.




