


The effect of feeding an all-TMR diet v. an all pasture diet to dairy cows has been examined previously (Kolver & Muller, Reference Kolver and Muller1998; Tucker et al. Reference Tucker, Rude and Wittayakun2001). Others have examined the merit of including supplemental concentrate to cows on pasture (White et al. Reference White, Benson, Washburn and Green2002; Fontaneli et al. Reference Fontaneli, Sollenberger, Littell and Staples2005) as well as some combination of TMR plus pasture (Soriano et al. Reference Soriano, Polan and Miller2001; Bargo et al. Reference Bargo, Muller, Delahoy and Cassidy2002). Our present study was designed to evaluate lactation performance when replacing incremental amounts of TMR with forage from pasture. To our knowledge, there are no published studies that examine animal performance under increasing TMR replacement with pasture. Research from the eastern USA comparing dairy confinement systems with pasture-based systems (with or without supplementation) has consistently shown reduced dry matter intake (DMI), milk production and body condition scores (BCS) for pasture-based dairying (Kolver & Muller, Reference Kolver and Muller1998; Soriano et al. Reference Soriano, Polan and Miller2001; Bargo et al. Reference Bargo, Muller, Delahoy and Cassidy2002; White et al. Reference White, Benson, Washburn and Green2002; Fontaneli et al. Reference Fontaneli, Sollenberger, Littell and Staples2005). Our objectives were to determine (1) the effect of increasing the proportion of forage from pasture on pasture and total intakes and (2) the level of pasture inclusion in TMR diets to optimize feed efficiency and lactation performance. Two independent experiments were conducted to monitor the effects of fall pasture (Experiment 1) and spring pasture (Experiment 2) on intakes, milk yield, and milk composition.
Materials and Methods
Two separate, 8-week experiments (each using 30 Holstein cows) were conducted to evaluate total and pasture DMI and lactation performance from cows fed different combinations of TMR and pasture. Experiment 1 evaluated pasture grazed in the fall and Experiment 2, with a different set of cows, evaluated pasture grazed in the spring. Unless stated otherwise, materials and methods common to both experiments are described below.
Cows and treatments
The experiments were conducted at the North Carolina State University Lake Wheeler Dairy Educational Unit, Raleigh NC, USA. Thirty lactating Holstein cows were blocked by milk yield, parity and days in milk (DIM). Cows were randomly assigned to either a nutritionally balanced TMR fed ad libitum (100:00, n=6, full nutrient positive control, no access to pasture) or one of the following three formulated partial mixed rations (PMR) with access to pasture (n=8) and targeted to be (1) 85% TMR and 15% pasture, (2) 70% TMR and 30% pasture and (3) 55% TMR and 45% pasture. Cows fed 100:00 had ad-libitum access to TMR to assure a daily feed refusal of approximately 10% of that offered. All cows were milked twice daily at 6.00 and 17.00 and milk production was recorded at each milking during the entire 8-week periods (Westfalia Dairy Plan, Westfalia Surge, Inc. Naperville IL, USA). Cows were weighed on two consecutive days at 15.00 during week 1 and weeks 5–8 inclusive. In the same weeks, body condition scores (BCS) were obtained by more than one independent scorer within each experiment, with at least one common scorer present for both experiments and all used a five-point scale (Wildman et al. Reference Wildman, Jones, Wagner, Boman, Troutt and Lesch1982).
Confinement and pasture feeding management
Four weeks prior to the initiation of the experiments, cows were trained to individual feeding gates (American Calan Inc., Northwood NH, USA) and then assigned to a corn-silage based TMR (Table 1). Cows were fed twice daily using a mixing cart (Uebler Manufacturing, Vernon NY, USA). The week prior to the initiation of each experiment, all 30 cows were confined and fed the control all-TMR diet. An average 48-h TMR consumption from all 30 cows on the last 2 d was used to arrive at the restricted TMR levels for the PMR treatments. One-third of the daily feed was offered after the morning milking and the rest after the afternoon milking. Daily TMR intakes for individual cows were calculated from the difference between the amount offered and amount refused. All cows had access to water in the barn, and cows on 100:00 treatment had access to a bare lot for daily exercise.
Table 1. Ingredients of the total mixed ration (TMR) offered to all cows during Experiments 1 and 2

† Blend of poultry by-product meal, hydrolysed poultry feathers, meat and bone meal, blood meal, and fish meal (Nutrimax Inc., Greensboro NC, USA)
‡ Soybean meal
§ Volclay® (American colloid Co., Arlington Heights IL, USA)
The three grazing groups had access to annual ryegrass (Lolium multiflorum Lam.; var. Marshall) that was seeded on 13 September 2004. Annual ryegrass was selected as the pasture owing to its nutritive value and agronomic performance (i.e., yield, length of growing season, seeding establishment and persistence under close grazing) in southeastern USA (Ellis & Lippke, Reference Ellis, Lippke and Holt1976). Cows grazed once daily as a single group (n=24) between the a.m. and p.m. milking for approximately 7 h/d (8.00–15.00) after which they were milked and housed overnight in free stall barns along with cows on 100:00. Cows were offered fresh pasture (i.e., strip grazing) and water was available in each paddock. One field of 4·9 ha was used during the experiments. To maximize nutritive value and avoid shortages during the grazing window, the amount of pasture offered was approximately 3-times more than the targeted pasture DMI for the cows on the 55:45 TMR:Pasture treatment (Bargo et al. Reference Bargo, Muller, Kolver and Delahoy2003).
Pasture intake measurements
Pasture intakes were measured according to the herbage disappearance method (HDM) based on the difference between pre- and post-grazing herbage mass (Lantinga et al. Reference Lantinga, Neuteboom, Meijs and Penning2004). Each week a cow was randomly selected from each of the three grazing treatments and assigned randomly to an individual plot. Sampling of herbage mass (pre- and post-grazing) occurred weekly by clipping eight 0·25-m2 quadrats in each plot before and immediately after the grazing period. Clippings were taken to ground level with the aid of battery-powered sheep-shearing heads (Sunbeam, Botany, New South Wales, Australia) and care was taken to avoid soil contamination. Clippings were made at the same time of the day, approximately 15.00 on two consecutive days, positioning the post-grazing frame approximately 10 cm from, but adjacent to, the previous, pre-grazing frame. Because pre- and post-grazing measurements were taken within a short window of time (24 h), growth during this period was ignored (Meijs et al. Reference Meijs, Walters, Keen and Leaver1982). Samples were placed in plastic bags, kept on ice, transported to the laboratory, and separated into three subsamples. One subsample was weighed and subsequently dried at 60°C in a forced-air oven until constant weight to determine DM content. A second subsample was separated into ryegrass green leaf, green stem, and other components (i.e., brown tissue and weeds) and dried at 60°C in a forced-air oven. The third subsample was freeze dried for later characterization of nutritive value. Ratios of leaf to stem provided additional information about pasture management and characterization.
Pasture intakes were also estimated using the NRC (2001) prediction equations as outlined by Macoon et al. (Reference Macoon, Sollenberger, Moore, Staples, Fike and Portier2003). Briefly, energy requirements included net energy (NE, Mcal/d) requirements for: (1) maintenance (NEM); (2) lactation (NEL); (3) body weight changes (NEBW); (4) walking activity (NEW); and (5) grazing activity (NEG). Total NE requirements were obtained by summing NEM, NEL, NEBW, NEW, and NEG. The NE from pasture intake was estimated as total NE requirements minus NE supplied by TMR intake.
Sample collection and analyses
Pasture samples were selectively hand plucked twice a week simulating animal grazing behaviour and stored at −20°C until freeze-dried. Weekly composites were ground in a Wiley mill (Thomas Scientific, Swedesboro NJ, USA) to pass through a 1-mm sieve. Freeze-dried weights were used to calculate DM. Total N was determined using a Kjeldahl procedure with an automated, colorimetric quantification of ammonia in digested samples (AOAC, 1990) and multiplied by 6·25 to estimate crude protein (CP). Neutral detergent fibre (NDF) and acid detergent fibre (ADF) were sequentially analysed using an Ankom fibre extractor (Ankom Technologies, Fairport NY, USA) as described by Van Soest et al. (Reference Van Soest, Robertson and Lewis1991). Non-protein N (NPN) was determined according to Licitra et al. (Reference Licitra, Hernandez and Van Soest1996). In-vitro true DM digestibility (IVTDMD) was determined in an Ankom II Daisy batch analyser for a period of 48 h in a solution containing 1600 ml of McDougall's buffer (Tilley & Terry, Reference Tilley and Terry1963) and 400 ml of strained ruminal fluid. Microbial residues were removed from indigestible residues by extraction with a neutral detergent solution. Samples of TMR were taken once a week, stored at −20°C, and subsequently freeze-dried. Samples were chemically analysed as described previously, with the exception of a heat-stable α-amylase that was added to the NDF procedure.
Individual milk samples were collected weekly and shipped to the regional Dairy Herd Improvement laboratory (United DHIA, Blacksburg VA, USA) where they were analysed for fat, protein, lactose, milk urea nitrogen (MUN) and somatic cell counts (SCC) using infrared spectrophotometry (Fossomatic 360, Foss Electronic, Slangerupgade, Denmark). Weighted composite milk samples from a.m. and p.m. milkings, as well as pasture and TMR samples, were analysed for fatty acid (FA) composition using gas–liquid chromatography (model CP 3380; GC Varian, Walnut Creek CA, USA). Milk samples (2 ml) were methylated (Kramer et al. Reference Kramer, Fellner, Dugan, Sauer, Mossoba and Yurawecz1997) and fatty acid methyl esters (FAME) were separated and quantified using a CP–Sil 88 capillary column (Chrompack, Middleburg, The Netherlands).
Experiment 1 (Fall pasture)
Experiment 1 was conducted from 26 October to 17 December 2004. Cows averaged 32·4±2·5 kg/d milk, 87·1±9·2 DIM, 1·6±0·2 lactations, 561±12 kg BW and 3·03±0·06 BCS at the initiation of the experiment. Although the PMR were initially targeted to be 85% TMR and 15% pasture, 70% TMR and 30% pasture and 55% TMR and 45% pasture, the corresponding actual PMR consumed and the ones used to describe the dietary treatments were 79% TMR and 21% pasture (79:21), 68% TMR and 32% pasture (68:32) and 59% TMR and 41% pasture (59:41).
Owing to a slower growth of pasture in the fall a second pasture (paddock 2, 4·1 ha) was used in addition to paddock 1 (4·9 ha) following the first grazing cycle. Paddock 2 was used for 9 d during this experiment and was located approximately 600 m from the free stall housing. Fresh pasture offered daily averaged 0·18 ha (75·3 m2 per cow) for 51 d. Paddocks 1 and 2 were fertilized before the beginning of Experiment 1 (4 October 2004) with 32·6 and 48·4 kg N/ha, respectively, and were spray-irrigated in November 2004 with lagoon waste delivering the equivalent of 22·6 and 7·8 kg N/ha, respectively. Estimates of pasture DMI using the HDM (n=6) were obtained once a week, with the exception of weeks 1 and 5 which had to be deferred because of incessant rain. Individual plot size varied according to herbage mass and averaged 144·6 m2/cow.
Experiment 2 (Spring pasture)
Experiment 2 was conducted from 22 March to 13 May 2005. Cows averaged 36·6±1·5 kg milk, 125·7±6·7 DIM, 1·9±0·2 lactations, 607±12 kg BW and 2·88±0·06 BCS at the initiation of the experiment. Instead of the targeted PMR of 85% TMR and 15% pasture, 70% TMR and 30% pasture and 55% TMR and 45% pasture, corresponding actual PMR consumed were 89% TMR and 11% pasture (89:11), 79% TMR and 21% pasture (79:21) and 65% TMR and 35% pasture (65:35).
Fresh pasture offered daily averaged 0·15 ha (63·3 m2 per cow) for 46 d. The pasture received no supplemental N during the spring of 2005 owing to State regulations that require a reduction in N-loading in the area (Neuse River Basin, Natural Resources Conservation Service, NRCS). Estimates of pasture DMI using the HDM (n=8) were obtained once a week during the 8-week trial. Individual plot size varied according to herbage mass and averaged 107·7 m2/cow.
Blood samples were collected once a week from coccygeal vessels. Initial samples were collected during week 1 and thereafter from weeks 5–8 before the p.m. milking. Plasma was analysed for glucose (YSI 2700 Select, Yellow Springs Instrument Co. Inc., Yellow Springs OH, USA), urea N (Technicon AutoAnalyzer II, Technicon Industrial method No. 334–74A/A, Technicon Industrial Systems, Tarrytown NY, USA) and non-esterified fatty acids (NEFA; Wako NEFA C kit No. 994–75409 E, Wako Chemicals USA, Inc., Richmond VA, USA).
Statistical analyses
Data from both experiments were analysed according to a randomized complete block design using the PROC MIXED procedure of SAS (2002), with a 3-week adaptation period (weeks 1–3) and a 5-week data collection period (weeks 4–8). The model included the effect of treatment in addition to a random block term and a residual error term. Linear, quadratic and lack-of-fit effects were tested by partitioning the degrees of freedom for treatment into single degree of freedom contrasts. For each variable analysed, covariates for pretreatment milk yield, DIM, parity, SCC, BW and BCS were included and kept in the model when significant (P<0·05). Least squares means and sem are reported for all data, and significance was established at P⩽0·05 and trends at P⩽0·10.
Results
Nutrient composition (Experiments 1 and 2)
Pasture CP averaged 23·7% in Experiment 1, and 13·9% in Experiment 2 (Table 2). Total soluble protein (Fraction A) comprised a greater proportion of the CP in the pasture compared with TMR. Linoleic (18:2) and oleic (18:1) comprised the main fatty acids in TMR, whereas linolenic (18:3) was the predominant fatty acid in pasture.
Table 2. Chemical composition (mean±se) of the TMR and pasture offered to all cows in Experiments 1 and 2 (DM basis)

† In vitro true DM disappearance
‡ A=non-protein N; B1=rapidly degraded fraction; B2=intermediately degraded fraction; B3=slowly degraded fraction; C=unavailable fraction
Experiment 1
Weather data:
Average monthly temperatures (°C) at soil level and at a 2-m height were 24·4 and 21·7 in September, 20·3 and 16·7 in October, 14·6 and 11·9 in November, and 9·4 and 8·5 in December (first 18 d), respectively, with an average of 13·4 and 11·4 for the experiment. This compares with a long-term mean (1982–2005) of 23·8 and 21·4 in September, 17·9 and 15·5 in October, 13·2 and 11·8 in November and 7·4 and 5·8 in December, respectively. Precipitation during September, October, November and December was 113·0, 36·2, 80·1 and 28·8 mm, respectively, and accumulated a total of 108·9 mm during the experiment. Long-term averages (1982–2005) for each of the 4 months were 128·0, 77·0, 46·2 and 62·5 mm, respectively (State Climate Office of North Carolina, NC CRONOS Database, data retrieval from Lake Wheeler Rd. Field Lab, Raleigh NC, USA).
Intakes, BW and BCS:
Intake of TMR was greatest for cows on 100:00 (25·8 kg/d) and decreased with increasing proportions of pasture in the diets (Table 3). Pasture intake tended to increase linearly (P=0·06) with decreasing TMR and averaged 4·6, 7·1 and 9·2 kg/d for the three PMR (21, 32 and 41% dietary pasture, respectively). BW changes (week 8 minus week 5) showed a quadratic (P=0·05) response, ranging from −4 (100:00) to 14 kg (79:21). BCS changes increased linearly (P=0·04) with increasing pasture, with changes ranging from −0·20 (100:00) to 0·25 (59:41).
Table 3. Intakes of dry matter (DM) and crude protein (CP), and adjusted body weights (BW) and body condition scores (BCS) of cows offered varying amounts of TMR and pasture in Experiment 1

† Actual diets represented by ratios of TMR to pasture
‡ L=linear; Q=quadratic
§ Estimates of pasture DMI based on a herbage disappearance method (HDM)
¶ Measurements taken on 2 consecutive days. Initial BW was used as a covariate
†† Measurements taken on 2 consecutive days. Initial BCS was used as a covariate
Milk production and milk composition:
Milk yields and milk constituents did not differ among treatments except for milk protein concentration and somatic cell scores (SCS) (Table 4). Milk protein concentration was greatest for cows on 68:32 (3·12%) and least for cows on 59:41 (2·84%). The concentration of 16:0 in milk fat decreased quadratically (P=0·002) with increasing intakes of pasture. The concentration of cis 18:1, cis-9, trans-11 CLA, 18:3 and unsaturated fatty acids (UFA, the sum of 18:1, 18:2, CLA and 18:3) in milk fat increased linearly (P<0·001) with increasing intakes of pasture, whereas the concentration of saturated fatty acids (SFA, the sum of 10:0, 12:0, 14:0, 16:0 and 18:0) in milk fat decreased linearly (P<0·001) with increasing intakes of pasture.
Table 4. Milk production, milk composition and milk fatty acid composition of cows offered varying amounts of TMR and pasture in Experiment 1

† Actual diets represented by ratios of TMR to pasture
‡ L=linear; Q=quadratic
§ Somatic cell score=((log10(SCC/1000)−2)/log10(2))+3
¶ Sum of C6 to C22 inclusive
†† Saturated fatty acids: Sum of 10:0, 12:0, 14:0, 16:0 and 18:0
‡‡ Unsaturated fatty acids: Sum of 18:1, 18:2, CLA and 18:3
Experiment 2
Weather data:
Average temperatures (°C) at soil level and at a 2-m height were 8·5 and 9·1 in March, 16·6 and 15·8 in April and 19·0 and 16·7 in May (first 13 d), respectively, with overall soil and 2-m average temperatures of 16·4 and 15·6 for the experiment. This compares with a long-term mean (1982–2005) of 11·5 and 11·4 in March, 16·5 and 19·1 in April and 23·0 and 22·6 in May, respectively. Precipitation during these months was 9·9, 8·7 and 50·0 mm and accumulated a total of 62·2 mm during the experiment compared with long-term averages (1982–2005) of 104·3, 50·5 and 33·7 mm for the respective months.
Intakes, BW and BCS:
Intake of TMR was greatest for cows on 100:00 (24·9 kg/d) and decreased with increasing proportions of pasture in the diets (Table 5). Pasture intake increased linearly (P<0·001) with decreasing TMR and averaged 2·2, 4·2 and 6·9 kg/d for the three PMR (11, 21 and 35% dietary pasture, respectively). Total daily DMI and daily CP intake decreased quadratically (P<0·01) with increasing intakes of pasture. BW changes (week 8 minus week 5) showed a quadratic (P<0·001) response, attributed to a net BW loss of cows on 100:00 (−16 kg) whereas cows on PMR gained BW. BCS, however, did not differ between treatments.
Table 5. Intakes of dry matter (DM) and crude protein (CP), and adjusted body weights (BW) and body condition scores (BCS) of cows offered varying amounts of TMR and pasture in Experiment 2

† Actual diets represented by ratios of TMR to pasture
‡ L=linear; Q=quadratic
§ Estimates of pasture DMI based on a herbage disappearance method (HDM)
¶ Measurements taken on 2 consecutive days. Initial BW was used as a covariate
†† Measurements taken on 2 consecutive days. Initial BCS was used as a covariate
Milk production and milk composition
Milk yield and 4% FCM yields decreased linearly (P<0·02) with increasing pasture (Table 6). Milk fat concentration tended (P=0·10) to increase with increasing pasture and ranged from 3·31% (100:00) to 3·68% (65:35). Although milk fat yields were unaffected by treatment, milk protein yields decreased linearly (P<0·01) with increasing intakes of pasture. MUN decreased quadratically (P<0·001) with increasing intakes of pasture. Concentrations of 16:0, 18:2 and SFA in milk fat decreased linearly (P<0·05) with increasing intakes of pasture. Conversely, the concentration of cis 18:1, cis-9, trans-11 CLA and 18:3 in milk fat increased linearly (P<0·05) with increasing intakes of pasture. Concentrations of trans 18:1 showed a quadratic (P<0·001) response to varying levels of TMR and pasture.
Table 6. Milk production, milk composition and milk fatty acid composition of cows offered varying amounts of TMR and pasture in Experiment 2

† Actual diets represented by ratios of TMR to pasture
‡ L=linear; Q=quadratic
§ Somatic cell score=((log10(SCC/1000)−2)/log10(2))+3
¶ Sum of C6 to C22 inclusive
†† Saturated fatty acids: Sum of 10:0, 12:0, 14:0, 16:0 and 18:0
‡‡ Unsaturated fatty acids: Sum of 18:1, 18:2, CLA and 18:3
Blood metabolites:
Plasma glucose concentration averaged 61·5 mg/dl in 100:00 and decreased linearly (P=0·002) with increasing intakes of pasture (Table 7). Plasma NEFA concentration at the initiation of the data collection period tended (P=0·09) to increase with increasing pasture, but was reduced in cows on PMR compared with cows on 100:00 by the end of the experiment (92 v. 209 μeq/l, respectively). Similarly, plasma urea nitrogen (PUN) decreased quadratically (P<0·001) with increasing intakes of pasture, presumably from the sharp decrease in PUN between 100:00 (13·9 mg/dl) and 89:11 (9·0 mg/dl).
Table 7. Plasma metabolites of cows offered varying amounts of TMR and pasture in Experiment 2
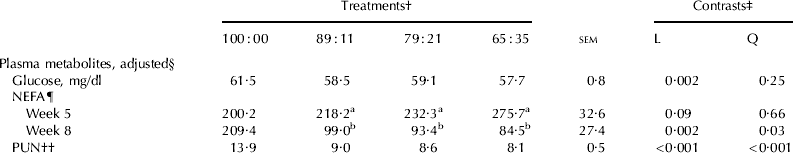
† Actual diets represented by ratios of TMR to pasture
‡ L=linear; Q=quadratic
§ Samples were collected once a week during weeks 5 to 8. Week 1 data were used as covariates
¶ Non-esterified fatty acids, μEq/l
†† Plasma urea nitrogen, mg/dl
a,b Least squares means in the same column with different superscripts are significantly different (P⩽0·05)
Experiments 1 and 2
Estimates of pasture DMI:
In Experiment 1, estimates of daily pasture DMI determined using the HDM were similar to those predicted using NRC equations (Table 8). In Experiment 2, however, estimates of daily pasture DMI obtained using the HDM method were consistently lower (P<0·05) than estimates obtained using the NRC equations. Also, in Experiment 2, both methods of estimating pasture DMI showed a linear increase (P<0·001) in pasture intake with increasing proportion of pasture in the diet.
Table 8. Estimates of pasture dry matter intake using a herbage disappearance method (HDM) and NRC estimates (NRC, 2001) in Experiments 1 and 2

† Actual diets represented by ratios of TMR to pasture
‡ L=linear; Q=quadratic
§ Estimates of pasture DMI based on a herbage disappearance method (HDM)
¶ Estimates of pasture DMI based on NRC estimates
a,b Least squares means in the same column with different superscripts are significantly different (P⩽0·05)
Discussion
Dietary nutrient composition
The TMR was formulated to provide 16% CP (38·8% Rumen undegradable protein, 6·1% ether extract; NRC 2001). The nutritive value of annual ryegrass varied from week 4 to week 8 during both experiments (only means are presented in Table 2). As the fall grazing season progressed, pasture DM concentration increased from 15·9% (week 4) to 21·1% (week 8) (data not shown). Similarly, DM concentration in Experiment 2 increased from 21·3% (week 4) to 25·7% (week 8) during the spring grazing season (data not shown).
In Experiment 1, pasture CP ranged from 21·6 to 25·8% (DM basis) and increased markedly from week 4 to week 5 which corresponded with the timing of N application. In Experiment 2, pasture CP ranged from 11·8% (week 6) to 15·1% (week 4). With cool-season grasses a rapid rate of decline in CP concentration seems to occur during the early stages of growth and the rate of decline slows as the pasture becomes more mature (Fick & Onstad, Reference Fick and Onstad1988). In Experiment 2 (spring pasture), rainfall was less and coupled with warmer temperature, forage maturation was accelerated resulting in reduced CP concentration compared with the fall forage (Experiment 1).
Pasture and total nutrient intakes
Total DMI for cows fed 100:00 averaged 25·8 kg/d in Experiment 1 and 24·9 kg/d in Experiment 2. Total DMI for the three PMR was similar within each experiment, and averaged 21·7 kg/d in Experiment 1 and 19·8 kg/d in Experiment 2. Reducing the proportion of TMR in the diets increased pasture consumption of cows on all PMR, but reduced total DMI compared with cows on 100:00. Reduced DMI from cows offered pasture most likely arises from a combination of restricted time spent grazing, grazing window (08.00–15.00) not including normal grazing pattern and the inherently more arduous grass harvesting compared with indoor, chopped feeding associated with TMR. Given extremely low refusals of TMR, cows offered pasture did not seem to substitute pasture for the TMR portion of their feed. All experimental cows were in the regular milking herd and offered an all-TMR ration prior to the initiation of the studies. They had all been exposed to pastures and had some grazing experience as growing heifers and during their dry periods. Reports from studies on feeding a combination of TMR and pasture show similar intake patterns (Soriano et al. Reference Soriano, Polan and Miller2001; Bargo et al. Reference Bargo, Muller, Delahoy and Cassidy2002). Despite having access to high-quality pasture, Bargo et al. (Reference Bargo, Muller, Delahoy and Cassidy2002) report that cows on pasture plus TMR that are offered 17 kg DM of TMR daily consume almost all of what was offered (15·5 kg DM consumed). Similarly, Soriano et al. (Reference Soriano, Polan and Miller2001) report TMR intakes of 26·6 kg DM/d for cows offered unrestricted TMR, 17·5 kg/d for cows offered TMR plus access to pasture in the afternoon, and 20·3 kg/d for cows offered TMR plus access to pasture in the morning.
HDM and NRC predictions for estimating DMI were positively correlated (P<0·05) with r=0·51 in Experiment 1 and r=0·61 in Experiment 2, and were of the magnitude reported by Macoon et al. (Reference Macoon, Sollenberger, Moore, Staples, Fike and Portier2003). The coefficients of determination (r 2) and se of prediction (Sxy) were 0·26 and 2·9 kg DM, respectively, for Experiment 1, and 0·37 and 1·7 kg DM, respectively, for Experiment 2. Also, in Experiment 1 no differences were reported between pasture DMI estimates using the HDM compared with those predicted from NRC, but our estimates were numerically greater than those obtained using NRC predictions, particularly as the proportion of pasture in the diet increased. In contrast, pasture DMI estimates based on HDM in Experiment 2 were lower than those predicted by NRC equations, but the magnitude of the difference was less as the proportion of pasture in the diet increased. The use of a HDM for estimating pasture DMI proved to be successful for rotational stocking and short grazing periods (Macoon et al. Reference Macoon, Sollenberger, Moore, Staples, Fike and Portier2003; Fontaneli et al. Reference Fontaneli, Sollenberger, Littell and Staples2005). Both the HDM and reverse calculations based on animal performance provide biologically reasonable estimates of DMI of cows grazing rye and ryegrass or ryegrass, crimson clover and red clover mixed pastures (Macoon et al. Reference Macoon, Sollenberger, Moore, Staples, Fike and Portier2003) and intensively managed kikuyu (Pennisetum clandestinum) pastures (Reeves et al. Reference Reeves, Fulkerson, Kellaway and Dove1996).
Milk production and composition
Although cows fed an all-TMR diet supported greater milk yields than the PMR combination the difference was significant only in Experiment 2. In Experiment 1, the decline in milk production of cows consuming 32% pasture was unexpected since increasing pasture to 41% showed no further decrease. In Experiment 2, however, the two high-pasture diets (21 and 35% pasture) resulted in significantly less milk production. Cows that consumed 100:00 in Experiment 1 produced 14% more milk and 5% more FCM than cows consuming 68:32. Likewise, cows that consumed 100:00 in Experiment 2 produced 15% more milk and 9% more FCM than cows consuming 79:21. Others report similar effects on milk production, but the magnitude of the differences vary, and are most likely attributable to a combination of DMI, dietary ingredients, pasture nutritive value, and the energetics involved in walking and grazing activities. Based on the NE concentration in milk and the extra daily activity of cows on the PMR (NRC, 2001), daily maintenance energy requirements for cows grazing in Experiment 1 were increased by 0·97, 0·85 and 0·61 Mcal for cows on 59:41, 68:32, and 79:21, respectively, compared with cows on 100:00. This potentially accounted for 1·4, 1·1, and 0·9 kg of extra milk for the corresponding grazing groups. Similarly, in Experiment 2, daily maintenance energy requirements for cows grazing were increased by 0·76, 0·60 and 0·43 Mcal for cows on 65:35, 79:21, and 89:11, respectively, compared with cows fed all-TMR, accounting for 1·1, 0·9 and 0·6 kg of milk for cows on the corresponding grazing groups. As expected, some combination of increased activity and decreased energy intake accounts for much of the difference in milk production among dietary treatments (Kolver & Muller, Reference Kolver and Muller1998; Bargo et al. Reference Bargo, Muller, Delahoy and Cassidy2002). Interestingly, cows that had access to pasture showed increased feed efficiencies (kg feed/kg 4% FCM) compared with cows consuming all-TMR. Feed conversion efficiencies of cows fed all-TMR were 0·80 in Experiment 1 and 0·76 in Experiment 2, compared with 0·68 and 0·63 across all PMR diets, respectively.
MUN in both experiments was within the normal range of 10–15 mg/dl (Moore & Varga, Reference Moore and Varga1996). Because MUN reflects CP intake (Wittwer et al. Reference Wittwer, Gallardo, Reyes and Opitz1999) MUN concentrations were greater for the three grazing groups during Experiment 1 (fall) than during Experiment 2 (spring). Factors such as the proportion of rumen degradable and rumen undegradable protein, and ratios of protein to energy in the diet also affect MUN (Baker et al. Reference Baker, Ferguson and Chalupa1995).
Milk fatty acid composition
Reduced TMR and increased pasture consumption in both experiments decreased total dietary fat, decreased the proportion of 18:1 and 18:2, and increased the proportion of 18:3 in the diets. The TMR fed to cows was relatively high in 16:0 and 18:2 whereas the pasture was high in 18:3 (Table 2). The FA profile of milk from both experiments was consistent with the practice of feeding stored v. fresh forages, and has been attributed to the loss of precursor FA during the silage- or hay-making process (Dewhurst et al. Reference Dewhurst, Scollan, Youell, Tweed and Humphreys2001). The FA composition reported for annual ryegrass was within the range of values reported elsewhere (Schroeder et al. Reference Schroeder, Gagliostro, Bargo, Delahoy and Muller2004).
Milk from grazing cows contains more trans fatty acids than milk from cows fed a TMR, primarily because of an increase in trans-11 18:1 concentration (Loor et al. Reference Loor, Soriano, Lin, Herbein and Polan2003). The increased concentration of trans-11 18:1 in milk from cows that graze was not seen in our study, suggesting limited impact of pasture on the biohydrogenation activity for the diets tested. The concentration of cis-9, trans-11 CLA in milk fat increased linearly with increasing pasture in both experiments and was consistent with the literature (Dhiman et al. Reference Dhiman, Anand, Satter and Pariza1999; Loor et al. Reference Loor, Soriano, Lin, Herbein and Polan2003; Schroeder et al. Reference Schroeder, Couderc, Bargo and Rearte2005).
Body weights, body condition scores and blood metabolites
A common feature of both experiments was the association between BW losses (−4 and −16 kg) and BCS losses (−0·2 and −0·07) for treatment 100:00 compared with the groups that had access to pasture. Despite these features, our results were unexpected, and are somewhat in disagreement with some of the findings reported previously. Cows fed TMR exclusively either maintain (Kolver & Muller, Reference Kolver and Muller1998; Bargo et al. Reference Bargo, Muller, Delahoy and Cassidy2002) or gain BCS (Tucker et al. Reference Tucker, Rude and Wittayakun2001) whereas cows consuming pasture-only diets lose BCS (Kolver & Muller, Reference Kolver and Muller1998; Tucker et al. Reference Tucker, Rude and Wittayakun2001).
Plasma glucose concentration was greatest (P=0·05) for 100:00 and least for 89:11 and 65:35 in week 5 (Experiment 2), but remained similar across treatments during the week that followed (data not shown). This pattern of plasma glucose concentration indicates a smaller discrepancy between energy demand and energy supply as the spring season progressed. These results are consistent with initially lower plasma glucose concentrations (60 v. 65 mg/dl) for cows transitioning from a TMR to a pasture-only diet, but become similar over time (Kolver & Muller, Reference Kolver and Muller1998). Also, Bargo et al. (Reference Bargo, Muller, Delahoy and Cassidy2002) report similar plasma glucose concentrations of cows consuming pasture plus concentrate, pasture plus TMR and TMR, averaging 65 mg/dl.
Plasma NEFA concentrations, a measure of adipose tissue mobilization, only tended to differ in week 5. However, by week 8, NEFA concentrations decreased quadratically with increasing intakes of pasture. Concurrently, BW losses in Experiment 2 (week 8 minus week 5) were only reported for treatment 100:00 (−16 kg) whereas all three grazing groups gained BW, indicating a greater body fat mobilization for cows on 100:00 to support greater levels of milk production.
In conclusion, including pasture as a major proportion in partial mixed rations is a viable option to confinement dairying. Most dairy enterprises in the eastern USA have a land resource that can be readily shifted into grass, or legume-based pastures that will provide an economic source of forage with adequate nutritive value. The producer has the freedom to vary the amount of pasture in the partial mixed ration to meet the economics of production while providing milk with enhanced CLA concentrations and hence a healthier consumer product.
We thank Sarah Jo McLeod for assisting with sample handling and analyses, and Pete Thompson, Justin Garrett, Weston McCorkle and Wayne McLamb for assistance in conducting these experiments.








